Taxonomy, Diversity and Cultivation of the Oudemansielloid/Xeruloid Taxa Hymenopellis, Mucidula, Oudemansiella, and Xerula with Respect to Their Bioactivities: A Review



 , , and
, , and
Abstract
:1. Introduction
2. Taxonomic Aspects of Oudemansielloid/Xeruloid Genera
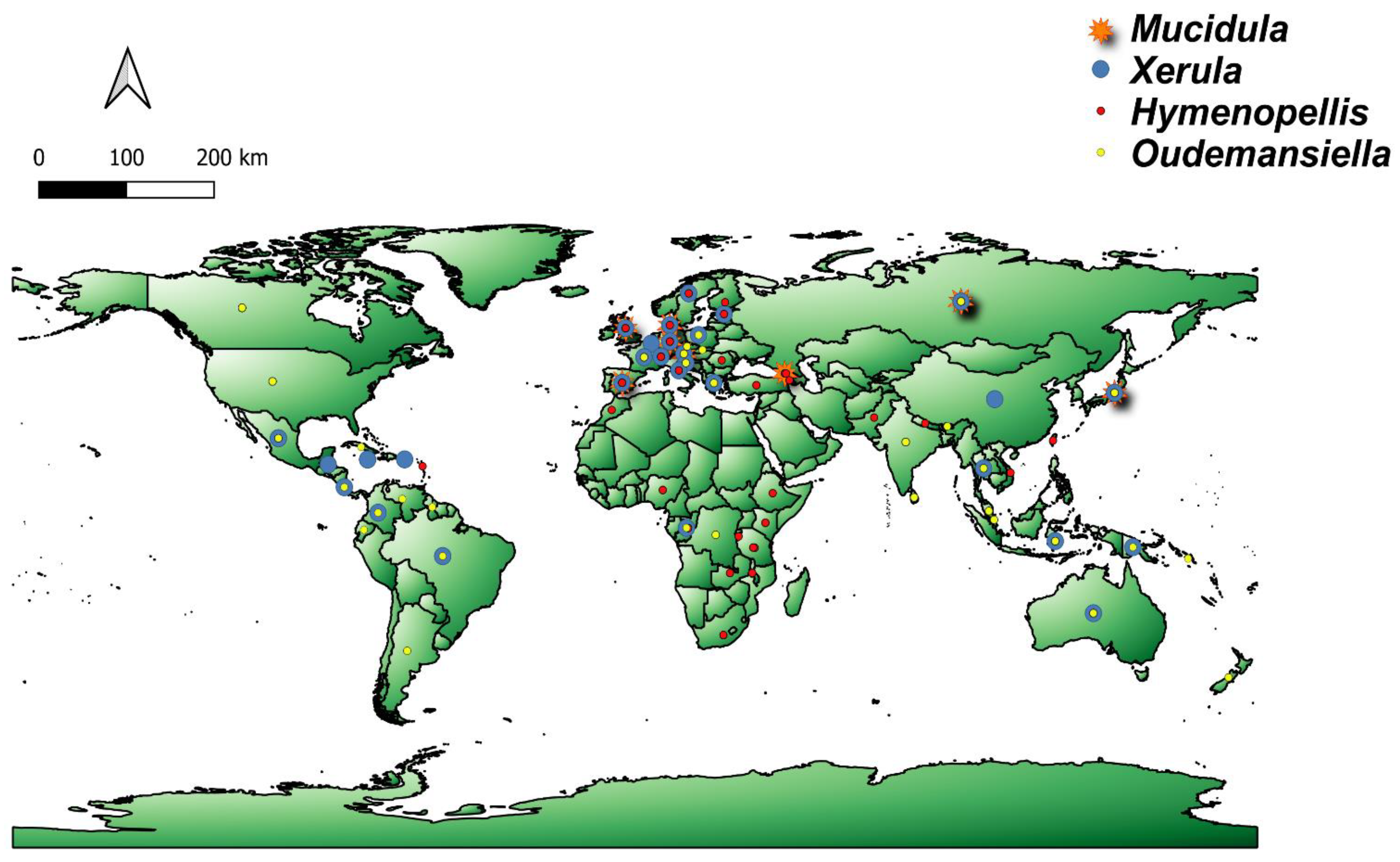
3. Geographical Distribution and Diversity of the Genera
3.1. Hymenopellis
3.2. Mucidula
3.3. Oudemansiella
3.4. Xerula
4. Cultivation of Important Species with Bioactivities
4.1. Cultivation of Hymenopellis
4.2. Cultivation of Mucidula
4.3. Cultivation of Oudemansiella
4.4. Cultivation of Xerula
5. Bioactivities and Mode of Action
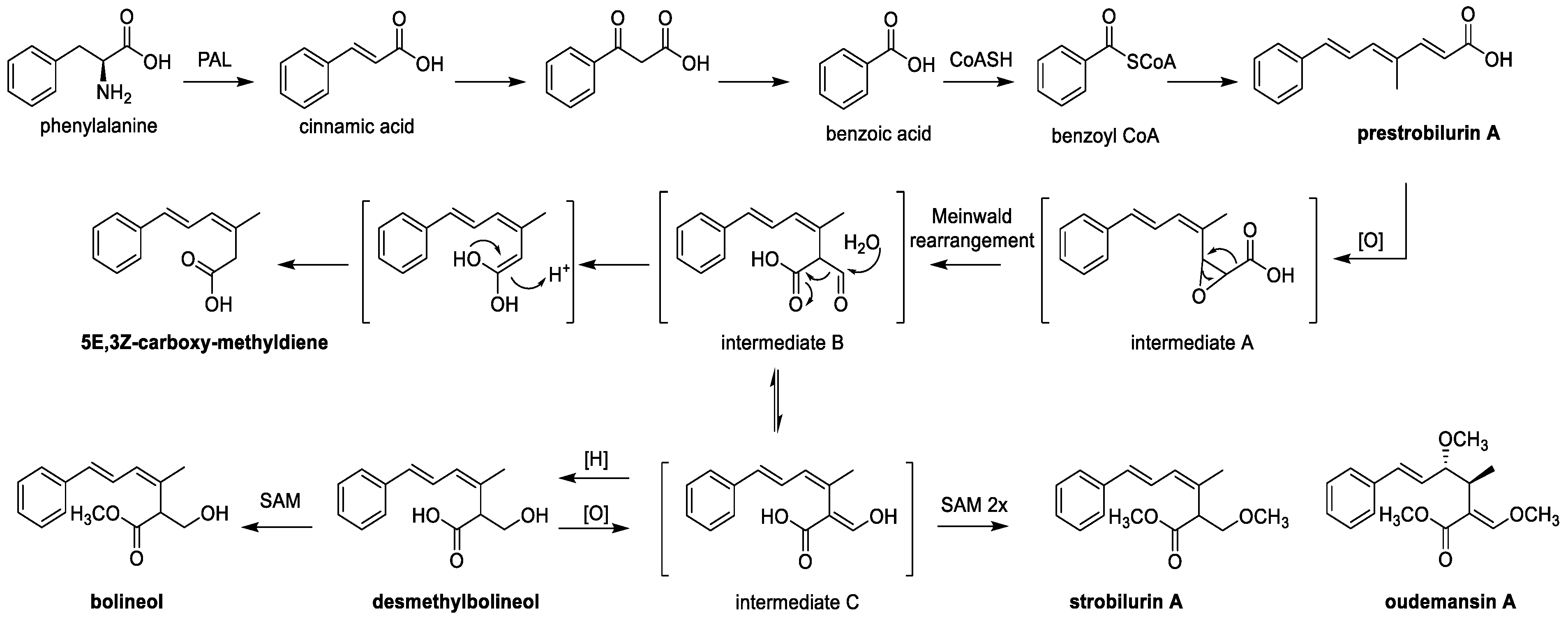
6. Biosynthesis of Strobilurins and Total Synthesis of Xerulins
7. Market and Commercialization
8. Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, J.X.; Chen, Q.; Huang, C.Y.; Gao, W.Q.J. History, current situation and trend of edible mushroom industry development. Mycosystema 2015, 34, 524–540. [Google Scholar]
- Hyde, K.D.; Xu, J.; Rapior, S.; Jeewon, R.; Lumyong, S.; Niego, A.G.T.; Abeywickrama, P.D.; Aluthmuhandiram, J.V.S.; Brahamanage, R.S.; Brooks, S.; et al. The amazing potential of fungi: 50 ways we can exploit fungi industrially. Fungal Divers. 2019, 97, 77–160. [Google Scholar] [CrossRef] [Green Version]
- De Silva, D.D.; Rapior, S.; Fons, F.; Bahkali, A.H.; Hyde, K.D. Medicinal mushrooms in supportive cancer therapies: An approach to anti-cancer effects and putative mechanisms of action. Fungal Divers. 2012, 55, 1–35. [Google Scholar] [CrossRef]
- De Silva, D.D.; Rapior, S.; Hyde, K.D.; Bahkali, A.H. Medicinal mushrooms in prevention and control of diabetes mellitus. Fungal Divers. 2012, 56, 1–29. [Google Scholar] [CrossRef]
- Sandargo, B.; Chepkirui, C.; Cheng, T.; Chaverra-Muñoz, L.; Thongbai, B.; Stadler, M.; Hüttel, S. Biological and chemical diversity go hand in hand: Basidiomycota as source of new pharmaceuticals and agrochemicals. Biotechnol. Adv. 2019, 37. [Google Scholar] [CrossRef] [PubMed]
- McMorris, T.C.; Anchel, M. Fungal Metabolites. The structures of the novel Sesquiterpenoids Illudin-S and -M. J. Am. Chem. Soc. 1965, 87, 1594–1600. [Google Scholar] [CrossRef] [PubMed]
- Novak, R.; Shlaes, D.M. The pleuromutilin antibiotics: A new class for human use. Curr. Opin. Investig. Drugs 2010, 11, 11. [Google Scholar]
- Gressler, M.; Löhr, N.A.; Schäfer, T.; Lawrinowitz, S.; Seibold, P.S.; Hoffmeister, D. Mind the mushroom: Natural product biosynthetic genes and enzymes of Basidiomycota. Nat. Prod Rep. 2021, in press. [Google Scholar] [CrossRef]
- Wang, Y.; Jia, J.; Ren, X.; Li, B.; Zhang, Q. Extraction, preliminary characterization and in vitro antioxidant activity of polysaccharides from Oudemansiella radicata mushroom. Int. J. Biol. Macromol. 2018, 120, 1760–1769. [Google Scholar] [CrossRef]
- Petersen, R.H.; Hughes, K.W. The Xerula/Oudemansiella Complex (Agaricales); Beihefte zu Nova Hedw; J. Cramer: Stuttgart, Germany, 2010; Volume 137, ISBN 978-3-443-51059-6. [Google Scholar]
- He, M.Q.; Zhao, R.L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; Sánchez-Ramírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar] [CrossRef] [Green Version]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Tang, L.Z.; Aptroot, A.; Castañeda-Ruiz, R.F.; Druzhinina, I.S.; Cai, F.; Ekanayaka, A.H.; Erdoğdu, M.; et al. A dynamic portal for a community-driven, continuously updated classification of Fungi and fungus-like organisms: Outlineoffungi. org. Mycosphere 2020, 11, 1514–1526. [Google Scholar] [CrossRef]
- Alves, M.J.; Ferreira, I.C.F.R.; Martins, A.; Pintado, M. Antimicrobial activity of wild mushroom extracts against clinical isolates resistant to different antibiotics. J. Appl. Microbiol. 2012, 113, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.H.; Cota, B.B.; Machado, K.M.G.; Rosa, C.A.; Zani, C.L. Antifungal and other biological activities from Oudemansiella canarii (Basidiomycota). World J. Microbiol. Biotechnol. 2005, 21, 983–987. [Google Scholar] [CrossRef]
- Rashid, S.; Aminuzzaman, F.; Islam, M.; Rahaman, M.; Rumainul, M. Biodiversity and Distribution of wild Mushrooms in the Southern Region of Bangladesh. J. Adv. Biol. Biotechnol. 2016, 9, 1–25. [Google Scholar] [CrossRef]
- Spegazzini, P. Sobré la Oudemansia platensis. Ann. Soc. Cien. Argent. 1880, 10, 279–280. [Google Scholar]
- Spegazzini, P.A. Fungi Argentini additis nonnullis Brasiliensibus Montevideensibusque. Pugillus Quartus. An. la Soc. Científica Argent. 1881, 12, 13–30. [Google Scholar]
- Moser, M. Studiem zur Gattung Oudemansiella Speg., Schleim- und Sammetrüblinge. Zeitschr Pilzk 1955, 21, 4–11. [Google Scholar]
- Singer, R. Diagnoses fungorum novorum Agaricalium II. Sydowia 1962, 15, 45–83. [Google Scholar]
- Singer, R. New genera of fungi. VIII. Persoonia 1962, 2, 407–415. [Google Scholar]
- Singer, R. Oudemansiellinae, Macrocystidinae, Pseudohiatulinae in South America. Darwiniana 1964, 13, 145–190. [Google Scholar]
- Clémençon, H. Taxonomic structure of the genus Oudemansiella (Agaricales). Sydowia 1979, 32, 74–80. [Google Scholar]
- Dörfelt, H. Taxonomische Studien in der Gattung Xerula R. Mre. (III). Feddes Repert. 1980, 91, 415–438. [Google Scholar] [CrossRef]
- Boekhout, T.; Bas, C. Notulae ad Floram agaricinam neerlandicam—XII. Some notes on the genera Oudemansiella and Xerula. Pers. Mol. Phylogeny Evol. Fungi 1986, 13, 45–56. [Google Scholar]
- Redhead, S.A.; Ginns, J.S.R. The Xerula (Collybia, Oudemansiella) radicata complex in Canada. Mycotaxon 1987, 30, 357–405. [Google Scholar]
- Petersen, R.H.; Nagasawa, E. The genus Xerula in temperate east Asia. Rep. Tottori Mycol. Inst. 2006, 43, 1–49. [Google Scholar]
- Petersen, R.H.; Baroni, T.J. Xerula hispida and Xerula setulosa (comb. nov.), two similar subtropical New World agarics. Mycotaxon 2007, 101, 113–136. [Google Scholar]
- Petersen, R.H. Species of Xerula from sub-Saharan Africa. Fungal Divers. 2008, 30, 121–147. [Google Scholar]
- Petersen, R.H.; Halling, R.E. Mating systems in the Xerulaceae: Oudemansiella. Trans Mycol Soc Japan 1993, 34, 409–421. [Google Scholar]
- Petersen, R.H.; Methven, A.S. Mating systems in the Xerulaceae: Xerula. Can. J. Bot. 1994, 72, 1151–1163. [Google Scholar] [CrossRef]
- Petersen, R.H. Scanning electron microscope images of basidiospores of Xerula (Physalacriaceae, Agaricales). Mycoscience 2008, 49, 19–34. [Google Scholar] [CrossRef]
- Corner, E.J.H. On the agaric genera Hohenbuehelia and Oudemansiella. Part. II: Oudemansiella Speg. Gard. Bull Singap. 1994, 46, 49–75. [Google Scholar]
- Contu, M. II genere Xerula (Basidiomyctonia, Agaricales) nell’ Italia centro-meridionale ed insulare. Micol. Veget. Mediterr. 2000, 15, 18–39. [Google Scholar]
- Mueller, G.M.; Wu, Q.X.; Huang, Y.Q.; Guo, S.Y.; Aldana-Gomez, R.; Vilgalys, R. Assessing biogeographic relationships between North American and Chinese macrofungi. J. Biogeogr. 2001, 28, 271–281. [Google Scholar] [CrossRef]
- Pegler, D.N.; Young, T.W.K. Classification of Oudemansiella (Basidiomycota: Tricholomataceae), with special reference to spore structure. Trans. Br. Mycol. Soc. 1986, 87, 583–602. [Google Scholar] [CrossRef]
- Rexer, K.H.; Kost, G.W. Zur Systematik der agaricoiden Hymenomycetes I. Zur Morphologie. Anatomie und Ontogenèse der Fruchtkôrper von Megacolhbia, Oudemansiella, Strobilurus. Zeitschr. Mykol. 1989, 55, 207–240. [Google Scholar]
- Rexer, K.H.; Kost, G.W. Zur Systematik der agaricoiden Hymenomyceten II. Verwandschaftliche Beziehungen von, Megacollybia, Oudemansiella, Strobilurus zur anderen Basidiomyceten, nebst Studien zur Ultrastruktur ihrer Cystiden. Zeitschr. Mykol. 1989, 55, 241–252. [Google Scholar]
- Yang, Z.L.; Zang, M. Classification of the genus Oudemansiella Speg. in Southwest China. Acta Mycol. Sin. 1993, 12, 16–27. [Google Scholar]
- Yang, Z.L. Further notes on the genus Oudemansiella from Southwestern China. Mycotaxon 2000, 74, 357–366. [Google Scholar]
- Mizuta, Y. Two new species and one new variety of Oudemansiella (Agaricales) from Japan. Mycoscience 2006, 47, 380–384. [Google Scholar] [CrossRef]
- Yang, Z.L.; Zhang, L.F.; Mueller, G.M.; Kost, G.W.; Rexer, K.H. A new systematic arrangement of the genus Oudemansiella s. str. (Physalacriaceae, Agaricales). Mycosystema 2009, 28, 1–13. [Google Scholar]
- Patouillard, N. Matériaux pour l’histoire des champignons. I. Les Hyménomycètes d’Europe 1887, 1, 1–166. [Google Scholar]
- Hoehnel, F. Fragmente zur Mykologie XII. Akademie der Wissenschaften in Wien Mathem. Naturw. Kl. 1910, 119, 877–958. [Google Scholar]
- Boursier, M. Note sur Ie genre Mucidula Pat. Bull. Trimest. La Soc. Mycol. Fr. 1924, 40, 332–333. [Google Scholar]
- Maire, R. Fungi Catalaunici. Treb. Junta Mus. Cienc. Nat. Barc. 1933, 3, 1–120. [Google Scholar]
- Singer, R. Das System der Agaricales. Ann. Mycol. 1936, 34, 286. [Google Scholar]
- Singer, R. The’Agaricales’ (Mushrooms) in modern taxonomy. Lilloa 1951, 22, 1–832. [Google Scholar]
- Relhan, R. Agaricus radicatus Relhan. Fl. Cantab. Suppl. 1786, 28. [Google Scholar]
- Index Fungorum Database. Available online: http://www.indexfungorum.org/ (accessed on 20 December 2020).
- Ronikier, A. Revision of the genus Xerula Maire (Basidiomycetes, Agaricales) in Poland. Acta Soc. Bot. Pol. 2003, 72. [Google Scholar] [CrossRef] [Green Version]
- Berkeley, M. Decades XXV-XXX. Sikkim Himalaya fungi, collected by J.D. Hooker. Lond. J. Bot. 1850, 2, 42–51. [Google Scholar]
- Ushijima, S.; Shimomura, N.; Nagasawa, E.; Maekawa, N. Taxonomic reevaluation of a fungus described as “Mucidula mucida” in Japan. Mushroom Sci. Biotechnol. 2012, 20, 22–30. [Google Scholar] [CrossRef]
- Petersen, R.H.; Gordon, S.A. Mating systems in hymenomycetes: New reports and new species. Mycologia 1994, 86, 743–757. [Google Scholar] [CrossRef]
- Liu, J.K.; Hyde, K.D.; Zhao, R.L. Four species of Oudemansiella and Xerula newly recorded from Thailand. Cryptogam. Mycol. 2009, 30, 341–353. [Google Scholar]
- Petersen, R.H.; Desjardin, D.E.; Krüger, D. Three type specimens designated in Oudemansiella. Fungal Divers. 2008, 32, 81–96. [Google Scholar]
- Berkeley, M.J.; Curtis, M.A. Characters of new fungi, collected in the North Pacific Exploring Expedition by Charles Wright. Proc. Amer. Acad. Arts. Sci. 1860, 4, 111–130. [Google Scholar]
- Lebel, T. Nomenclatural changes and corrections for some previously described AustralAsian truffle-like fungi (Basidiomycetes). Muelleria 2017, 36, 8–14. [Google Scholar]
- Zając, A.; Zając, M. Atlas Rozmieszczenia Roślin Naczy Niowych w Polsce; Pracownia Chorologii Kompute Rowej Instytutu Botaniki Uniwersytetu Jagiellońskiego: Kraków, Poland, 2001; ISBN 8391516113. [Google Scholar]
- Chandrasrikul, A.; Suwanarit, P.; Sangwanit, U.; Lumyong, S.; Payapanon, A.; Sanoamuang, N.; Pukahuta, C.; Petcharat, V.; Sardsud, U.; Duengkae, K.; et al. Checklist of Mushrooms (Basidiomycetes) in Thailand; Office of Natural Resources and Environmental Policy and Planning: Bangkok, Thailand, 2011.
- Dörfelt, H. Taxonomische Studien in der Gattung Xerula R. Mre. (IX). Feddes Repert. 1984, 95, 189–200. [Google Scholar]
- Métrod, G. Sur le genre Xerula M. Maire. Bull. Soc. Mycol. Fr. 1952, 68, 178–180. [Google Scholar]
- Clémençon, H. Neue Arten von Agaricalien und Notizen zu bemerkenswerten Funden aus der Schweiz. Nov. Hedwig. 1977, 28, 1–44. [Google Scholar]
- Murrill, W.A. Agaricaceae (pars). N. Am. Flora 1916, 9, 297–374. [Google Scholar]
- Singer, R. Type studies on Basidiomycetes VI. Lilloa 1953, 26, 59–159. [Google Scholar]
- Alexopoulos, C.J.; Mims, C.W.B. Introductory Mycology, 4th ed.; John Wiley & Sons Inc.: New York, NY, USA, 1996. [Google Scholar]
- Gupta, S.; Summuna, B.; Gupta, M.; Annepu, S.K. Edible Mushrooms: Cultivation, Bioactive Molecules, and Health Benefits. In Bioactive Molecules in Food; Springer: Heidelberg, Germany, 2018; pp. 1–33. [Google Scholar]
- Boddy, L. Chapter 9—Interactions with humans and other animals. In The Fungi; Academic Press: Amsterdam, The Netherlands, 2016; pp. 293–336. [Google Scholar]
- Zervakis, G.I.; Koutrotsios, G.; Katsaris, P. Composted versus raw olive mill waste as substrates for the production of medicinal mushrooms: An assessment of selected cultivation and quality parameters. Biomed. Res. Int. 2013, 2013, 546830. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.O.; Chang, K.C.; Kim, T.H.; Lee, Y.S.; Lee, U.Y.; Lee, T.S.; Lee, M.W. The fruiting body formation of Oudemansiella radicata in the sawdust of oak (Quercus variabilis) mixed with rice bran. Mycobiology 2006, 34, 30–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z. The domestication and cultivation of O. radicata. Edible Fungi China 2000, 19, 5–6. [Google Scholar]
- Lee, G.W.; Jaysinghe, C.; Imtiaj, A.; Shim, M.J.; Hur, H.; Lee, M.W.; Lee, K.R.; Kim, S.H.; Kim, H.Y.; Lee, U.Y.; et al. The Artificial Cultivation of Oudemansiella mucida on the Oak Sawdust Medium. Mycobiology 2007, 35, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira Ruegger, M.J.; Tornisielo, S.M.T.; Bononi, V.L.R.; Capelari, M. Cultivation of the edible mushroom Oudemansiella canarii (Jungh.) Höhn. in lignocellulosic substrates. Brazilian J. Microbiol. 2001, 32, 211–214. [Google Scholar] [CrossRef]
- Xu, F.; Li, Z.; Liu, Y.; Rong, C.; Wang, S. Evaluation of edible mushroom Oudemansiella canarii cultivation on different lignocellulosic substrates. Saudi J. Biol. Sci. 2016, 23, 607–613. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-B.; Kim, S.-H.; Lee, K.-R.; Shim, J.-O.; Lee, M.-W.; Shim, M.-J.; Lee, U.-Y.; Lee, T.-S. The Optimal Culture Conditions for the Mycelial Growth of Oudemansiella radicata. Mycobiology 2005, 33, 230–234. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.J.; Zhao, Q.; Wang, S.X.; Yang, Z.L. What is the radicate Oudemansiella cultivated in China? Phytotaxa 2016, 286, 1–12. [Google Scholar] [CrossRef]
- Ning, X.; LiGuo, F.; ChunHui, W.; ZhaoLi, D.; ShiCheng, Z.; Huan, L. Optimization of culture conditions of fermentation technology for Oudemansiella raphanipes liquid strain fermenter. J. South. Agric. 2019, 50, 344–349. [Google Scholar]
- Musílek, V.; Černá, J.; Šašek, V.; Semerdžieva, M.; Vondráček, M. Antifungal antibiotic of the basidiomycete Oudemansiella mucida—I. Isolation and cultivation of a producing strain. Folia Microbiol. 1969, 14, 377–387. [Google Scholar] [CrossRef]
- Magingo, F.S.; Oriyo, N.M.; Kivaisi, A.K.; Danell, E. Cultivation of Oudemansiella tanzanica nom. prov. on agricultural solid wastes in Tanzania. Mycologia 2004, 96, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhang, L.; Zhang, M.; Xu, Z.; Shang, X.; Tan, Q. Domestication and cultivation of Oudemansiella submucida. Acta Edulis Fungi 2012, 19, 45–48. [Google Scholar]
- Houjiang, L.; Guomin, H.; Zhao, L. Artificial Cultivation Method for Xerula pudens (Pers.:Fr.) Sing. CN Patent CN104396561A, 21 October 2014. [Google Scholar]
- Karwehl, S.; Stadler, M. Exploitation of fungal biodiversity for discovery of novel antibiotics. Curr. Top. Microbiol. Immunol. 2016, 398, 303–338. [Google Scholar] [CrossRef] [PubMed]
- Esser, K.; Bennett, J. The Mycota: A Comprehensive Treatise of Fungi as Experimental System for Basic and Applied Research; Springer: Berlin, Germany, 2002. [Google Scholar]
- Nofiani, R.; de Mattos-Shipley, K.; Lebe, K.E.; Han, L.C.; Iqbal, Z.; Bailey, A.M.; Willis, C.L.; Simpson, T.J.; Cox, R.J. Strobilurin biosynthesis in basidiomycete fungi. Nat. Commun. 2018, 9, 3940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiteller, P. Chemical ecology of fungi. Nat. Prod. Rep. 2015, 32, 971–993. [Google Scholar] [CrossRef] [PubMed]
- Pelley, R.P.; Strickland, F.M. Plants, polysaccharides, and the treatment and prevention of neoplasia. Crit. Rev. Oncog. 2000, 11, 189–225. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-N.; Chang, C.-S.; Hung, M.-H.; Chen, S.; Wang, W.; Tai, C.-J.; Lu, C.-L. The effect of mushrooms beta-glucans from solid culture of Ganoderma lucidum on inhibition of the primary tumor metastasis. Evid. Based. Complement. Altern. Med. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Baldassano, S.; Accardi, G.; Vasto, S. Beta-glucans and cancer: The influence of inflammation and gut peptide. Eur. J. Med. Chem. 2017, 142, 486–492. [Google Scholar] [CrossRef]
- Sivanandhan, S.; Khusro, A.; Paulraj, M.G.; Ignacimuthu, S.; Al-Dhabi, N.A. Biocontrol properties of basidiomycetes: An overview. J. Fungi 2017, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Tsantrizos, Y.S.; Yang, X.; McClory, A. Studies on the biosynthesis of the fungal metabolite oudenone. 2. Synthesis and enzymatic cyclization of an α-diketone, open-chain precursor into oudenone in cultures of Oudemansiella radicata. J. Org. Chem. 1999, 64, 6609–6614. [Google Scholar] [CrossRef]
- Umezawa, H.; Takeuchi, T.; Iinuma, H.; Suzuki, K.; Ito, M.; Matsuzaki, M.; Nagatsu, T.; Tanabe, O. A new microbial product, oudenone, inhibiting tyrosine hydroxylase. J. Antibiot. 1970, 23, 514–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Z.; Li, J.; Song, X.; Zhang, J.; Wang, X.; Jing, H.; Ren, Z.; Li, S.; Zhang, C.; Jia, L. Antioxidative, anti-inflammation and lung-protective effects of mycelia selenium polysaccharides from Oudemansiella radicata. Int. J. Biol. Macromol. 2017, 104, 1158–1164. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ng, T.; Wang, H. Isolation and characterization of a novel lectin from the wild mushroom Oudemansiella radicata (Relhan.: Fr.) sing. Biotechnol. Bioprocess Eng. 2013, 18, 465–471. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Ranadive, K.; Belsare, J.; Deokule, M.; Jagtap, S.; Jadhav, N.; Vaidya, H. Glimpses of antimicrobial activity of fungi from World. J. New Biol. Reports 2013, 2, 142–162. [Google Scholar]
- Hleba, L.; Kompas, M.; Hutková, J.; Rajtar, M.; Petrová, J.; Čuboň, J.; Kántor, A.; Kačániová, M. Antimicrobial activity of crude ethanolic extracts from some medicinal mushrooms. J. Microbiol. Biotechnol. Food Sci. 2016, 5, 60–63. [Google Scholar] [CrossRef] [Green Version]
- Rosa, L.H.; Gomes Machado, K.M.; Jacob, C.C.; Capelari, M.; Augusto Rosa, C.; Leomar Zani, C. Screening of Brazilian Basidiomycetes for antimicrobial activity. Mem. Inst. Oswaldo Cruz. 2003, 98, 967–974. [Google Scholar] [CrossRef] [Green Version]
- Anke, T.; Werle, A.; Bross, M.; Steglich, W. The strobilurins—New antifungal antibiotics from the basidiomycete Strobilurus tenacellus (Pers. ex Fr.) Sing. J Antibiot. 1977, 30, 806–810. [Google Scholar] [CrossRef]
- Anke, T.; Jürgen Hecht, H.; Schramm, G.; Steglich, W. Antibiotics from basidiomycetes. IX. Oudemansin, an antifungal antibiotic from Oudemansiella mucida (Schrader ex Fr.) Hoehnel (Agaricales). J. Antibiot. 1979, 32, 1112–1117. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, Z.; Han, L.C.; Soares-Sello, A.M.; Nofiani, R.; Thormann, G.; Zeeck, A.; Cox, R.J.; Willis, C.L.; Simpson, T.J. Investigations into the biosynthesis of the antifungal strobilurins. Org. Biomol. Chem. 2018, 16, 5524–5532. [Google Scholar] [CrossRef]
- Anke, T.; Werle, A.; Bross, M.; Steglich, W. Antibiotics from basidiomycetes XXXIII. Oudemansin X, a new antifungal E-β-methoxyacrylate from Oudemansiella radicata (Relhan ex Fr.) Sing. J. Antibiotics 1990, 43, 1010–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, W.; Anke, T.; Steffan, B.; Steglich, W. Antibiotics from basidiomycetes: XXXII. Strobilurin E: A new cytostatic and antifungal (E)-β-methoxyacrylate antibiotic from Crepidotus fulvotomentosus Peck. J. Antibiot. 1990, 43, 207–212. [Google Scholar] [CrossRef] [PubMed]
- von Jagow, G.V.; Gribble, G.W.; Trumpower, B.L. Mucidin and strobilurin A are identical and inhibit electron transfer in the Cytochrome bc1 complex of the mitochondrial respiratory chain at the same site as myxothiazol. Biochemistry 1986, 25, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Balba, H. Review of strobilurin fungicide chemicals. J. Environ. Sci. Health Part B Pestic. Food Contam. Agric. Wastes 2007, 42, 441–451. [Google Scholar] [CrossRef]
- Šubík, J.; Behúň, M.; Šmigáň, P.; Musílek, V. Mode of action of mucidin, a new antifungal antibiotic produced by the basidiomycete Oudemansiella mucida. BBA Gen. Subj. 1974, 343, 363–370. [Google Scholar] [CrossRef]
- Ying, J.; Mao, X.; Ma, Q.; Zong, Y.; Wen, H. Icons of Medicinal Fungi from China (translated, Yuehan X); Science Press: Beijing, China, 1987. [Google Scholar]
- Vondracek, M.; Vondrackova, J.; Sedmera, P.; Musilek, V. Another antibiotic from the basidiomycete Oudemansiella mucida. Collect. Czechoslov. Chem. Commun. 1983, 48, 1508–1512. [Google Scholar] [CrossRef]
- Tsantrizos, Y.S.; Zhou, F.; Famili, P.; Yang, X. Biosynthesis of the hypotensive metabolite oudenone by Oudemansiella radicata. 1. Intact incorporation of a tetraketide chain elongation intermediate. J. Org. Chem. 1995, 60, 6922–6929. [Google Scholar] [CrossRef]
- Anke, T.; Besl, H.; Mocek, U.; Steglich, W. Antibiotics from basidiomycetes. XVIII. Strobilurin C and oudemansin B, two new antifungal metabolites from Xerula species (Agaricales). J. Antibiot. 1983, 36, 661–666. [Google Scholar] [CrossRef] [Green Version]
- Zou, X. Optimization of nutritional factors for exopolysaccharide production by submerged cultivation of the medicinal mushroom Oudemansiella radicata. World J. Microbiol. Biotechnol. 2005, 21, 1267–1271. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Ke, Y.; Li, C.; Zhang, Z.; Wu, Y.; Hu, B.; Liu, A.; Luo, Q.; Wu, W. In vitro saliva-gastrointestinal digestion and fecal fermentation of Oudemansiella radicata polysaccharides reveal its digestion profile and effect on the modulation of the gut microbiota. Carbohydr. Polym. 2021, 251, 117041. [Google Scholar] [CrossRef]
- Kuhnt, D.; Anke, T.; Besl, H.; Bross, M.; Herrmann, R.; Mocek, U.; Steffan, B.; Steglich, W. Antibiotics from basidiomycetes. XXXVII. New inhibitors of cholesterol biosynthesis from cultures of Xerula melanotricha Dörfelt. J. Antibiot. 1990, 43, 1413–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, W.; Anke, T.; Bross, M.; Steglich, W. Strobilurin D and strobilurin F: Two new cytostatic and antifungal (E)-β-methoxyacrylate antibiotics from Cyphellopsis anomala. Planta Med. 1990, 56, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Sadorn, K.; Saepua, S.; Boonyuen, N.; Laksanacharoen, P.; Rachtawee, P.; Pittayakhajonwut, P. Antimicrobial activity and cytotoxicity of polyketides isolated from the mushroom: Xerula sp. BCC56836. RSC Adv. 2016, 6, 94510–94523. [Google Scholar] [CrossRef]
- Grabley, S.; Thiericke, R. Drug Discovery from Nature; Springer: Berlin/Heidelberg, Germany, 1999. [Google Scholar]
- Lorenzen, K.; Anke, T. Basidiomycetes as a source for new bioactive natural products. Curr. Org. Chem. 1998, 2, 329–364. [Google Scholar]
- Liu, Q.; Zhu, M.; Geng, X.; Wang, H.; Ng, T.B. Characterization of polysaccharides with antioxidant and hepatoprotective activities from the edible mushroom Oudemansiella radicata. Molecules 2017, 22, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, K.; Nandi, S.; Dutta, A.K. Microanatomical and physicochemical characterization and antioxidative activity of methanolic extract of Oudemansiella canarii (Jungh.) Höhn. Turkish J. Pharm. Sci. 2019, 16, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Liu, X.; Wang, W.; Yang, Q.; Dong, Y.; Xu, N.; Zhang, C.; Song, X.; Ren, Z.; Zhao, F.; et al. Characteristic anti-inflammatory and antioxidative effects of enzymatic- and acidic- hydrolysed mycelium polysaccharides by Oudemansiella radicata on LPS-induced lung injury. Carbohydr. Polym. 2019, 204, 142–151. [Google Scholar] [CrossRef]
- Leong, R.W.L.; Chan, F.K.L. Drug-induced side effects affecting the gastrointestinal tract. Expert Opin. Drug Saf. 2006, 5, 585–592. [Google Scholar] [CrossRef]
- Philpott, H.L.; Nandurkar, S.; Lubel, J.; Gibson, P.R. Drug-induced gastrointestinal disorders. Postgrad. Med. J. 2014, 90, 411–419. [Google Scholar] [CrossRef]
- Fernandes, D.C.R.; Norman, A.J. Drug-induced gastrointestinal disorders. Medcine 2019, 47, 301–308. [Google Scholar] [CrossRef]
- Elsayed, E.A.; El Enshasy, H.; Wadaan, M.A.M.; Aziz, R. Mushrooms: A potential natural source of anti-inflammatory compounds for medical applications. Mediat. Inflamm. 2014, 2014, 805841. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, B.; Grzywacz-Kisielewska, A.; Kała, K.; Gdula-Argasińska, J. Anti-inflammatory properties of edible mushrooms: A review. Food Chem. 2018, 243, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Zhang, C.; Tian, C.; Ren, Z.; Song, X.; Wang, X.; Xu, N.; Jing, H.; Li, S.; Liu, W.; et al. Characterization, antioxidation, anti-inflammation and renoprotection effects of selenized mycelia polysaccharides from Oudemansiella radicata. Carbohydr. Polym. 2018, 181, 1224–1234. [Google Scholar] [CrossRef] [PubMed]
- C.Ooi, V.; Liu, F. Immunomodulation and anti-cancer activity of polysaccharide-protein complexes. Curr. Med. Chem. 2012, 7, 715–729. [Google Scholar] [CrossRef] [Green Version]
- Sobieralski, K.; Siwulski, M.; Lisiecka, J.; Jedryczka, M.; Sas-Golak, I.; Fruzyńska-Jóźwiak, D. Fungi-derived β-glucans as a component of functional food. Acta Sci. Pol. Hortorum Cultus 2012, 11, 111–128. [Google Scholar]
- Novak, M.; Vetvicka, V. β-glucans, history, and the present: Immunomodulatory aspects and mechanisms of action. J. Immunotoxicol. 2008, 5, 47–57. [Google Scholar] [CrossRef]
- Ayeka, P.A. Potential of mushroom compounds as immunomodulators in cancer immunotherapy: A review. Evid. Based Complement. Altern. Med. 2018, 2018, 7271509. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.A.A.; Rouf, R.; Tiralongo, E.; May, T.W.; Tiralongo, J. Mushroom lectins: Specificity, structure and bioactivity relevant to human disease. Int. J. Mol. Sci. 2015, 16, 7802–7838. [Google Scholar] [CrossRef] [Green Version]
- Siegel, K.; Brückner, R. First synthesis of Xerulin, an inhibitor of the biosynthesis of cholesterol. Synlett 1999, 1227–1230. [Google Scholar] [CrossRef]
- Lebe, K.E.; Cox, R.J. O -Methylation steps during strobilurin and bolineol biosynthesis. RSC Adv. 2019, 9, 31527–31531. [Google Scholar] [CrossRef] [Green Version]
- Siegel, K.; Brückner, R. First total synthesis of dihydroxerulin, a potent inhibitor of the biosynthesis of cholesterol. Chem. Eur. J. 1998, 4, 1116–1122. [Google Scholar] [CrossRef]
- Negishi, E.I.; Alimardanov, A.; Xu, C. An efficient and stereoselective synthesis of xerulin via Pd-catalyzed cross coupling and lactonization featuring (E)-iodobromoethylene as a novel two-carbon synthon. Org. Lett. 2000, 2, 65–67. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest Manag. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, E.T.; Lopes, I.; Pardal, M.Â. Occurrence, fate and effects of azoxystrobin in aquatic ecosystems: A review. Environ. Int. 2013, 53, 18–28. [Google Scholar] [CrossRef]
- Henn, A. Fungicides Listed for Use on Pumpkin Diseases in Mississippi; Efficacy is from the Southeastern U.S. In Southeastern US 2016 Vegetable Crop Handbook; Kuhar, T.P., Arancibia, R.A., Straw, A., Rideout, S.L., Reiter, M.S., Kemble, J.M., Eds.; Virginia Polytechnic Institute and State University: Blacksburg, VA, USA, 2016. [Google Scholar]
- Pohanish, R.P. Sittig’s Handbook of Pesticides and Agricultural Chemicals, 2nd ed.; William Andrew Publishing: Norwich, NY, USA, 2015; ISBN 9781455731572. [Google Scholar]
- Amaro, A.C.E.; Ramos, A.R.P.; Macedo, A.C.; Ono, E.O.; Rodrigues, J.D. Effects of the fungicides azoxystrobin, pyraclostrobin and boscalid on the physiology of Japanese cucumber. Sci. Hortic. 2018, 228, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Bahkali, A.H.; Moslem, M.A. Fungi—An unusual source for cosmetics. Fungal Divers. 2010, 43, 1–9. [Google Scholar] [CrossRef]
- Thakur, M.P.; Singh, H.K. Mushrooms, their bioactive compounds and medicinal uses: A review. Med. Plants 2013, 5, 1–20. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author | Year | Arrangement | Adopted |
|---|---|---|---|
| Spegazzini [16] | 1880 | Initially proposed Oudemansia to accommodate a single species, Agaricus platensis Speg. | |
| Spegazzini [17] | 1881 | Changed the name to Oudemansiella | |
| Patouillard [42] | 1887 | Erected Mucidula to separate Agaricus mucidus from both Collybia (Fr.) Kummer and Armillaria (Fr.) Kummer based on the presence of velar layers and the voluminous spores | |
| Hoehnel [43] | 1910 | Emended Oudemansiella to include species with velar layers, a gelatinized pileipellis, and large cystidia and spores | |
| Boursier [44] | 1924 1924 | Expanded Mucidula to include Collybia radicata (Relhan: Fr.) Quel. and C. longipes (Bull.) Kummer, emphasizing morphological similarities (spores, basidia, cystidia, hymenioderm) | |
| Maire [45] | 1933 | Separated C. longipes from Mucidula and proposed the new genus Xerula | Singer [46,47] |
| Moser [18] | 1955 | Merged Xerula and Mucidula into Oudemansiella | Singer [19,20,21] |
| Clémençon [22] | 1979 | Treated Xerula as one of the 5 subgenera of Oudemansiella | |
| Dörfelt [23] | 1980 | Retained Oudemansiella and Xerula as independent genera | Boekhout & Bas [24], Redhead et al. [25], Petersen & Halling [29], Petersen & Methven [30], Corner [32], Contu [33], Mueller et al. [34], Petersen & Nagasawa [26], Petersen & Baroni [27], Petersen [28,31] |
| Pegler & Young [35] | 1987 | Divided Oudemansiella into five sections under the subgenera Oudemansiella and Xerula | Rexer & Kost [36,37], Yang & Zang [38], Yang [39], Mizuta [40] |
| Yang et al. [41] | 2009 | Divided Oudemansiella into four sections (Oudemansiella, Mucidula, Dactylosporina and Radicatae) | |
| Petersen & Hughes [10] | 2010 | Introduction of four new genera (Hymenopellis, Paraxerula, Ponticulomyces, Protoxerula) and reconfiguration of other genera such Dactylosporina, Mucidula, Oudemansiella and Xerula | accepted until now |
| Species | Substrate | Biological Efficiency (%) | References |
|---|---|---|---|
| Hymenopellis radicata | Oak sawdust | – | Shim et al. [69] |
| Sawdust | 100 | Gao [70] | |
| Mucidula mucida | Oak sawdust | – | Lee et al. [71] |
| Oudemansiella canarii | Sugar-cane bagasse | 55.66 | Silveira Ruegger et al. [72] |
| Eucalyptus sawdust | 19.51 | ||
| Cottonseed hull | 113.64 | Xu et al. [73] | |
| Corncob | 105.65 | ||
| Sawdust | 85.49 |
| Species | Bioactive Compounds | Biological Activities | References |
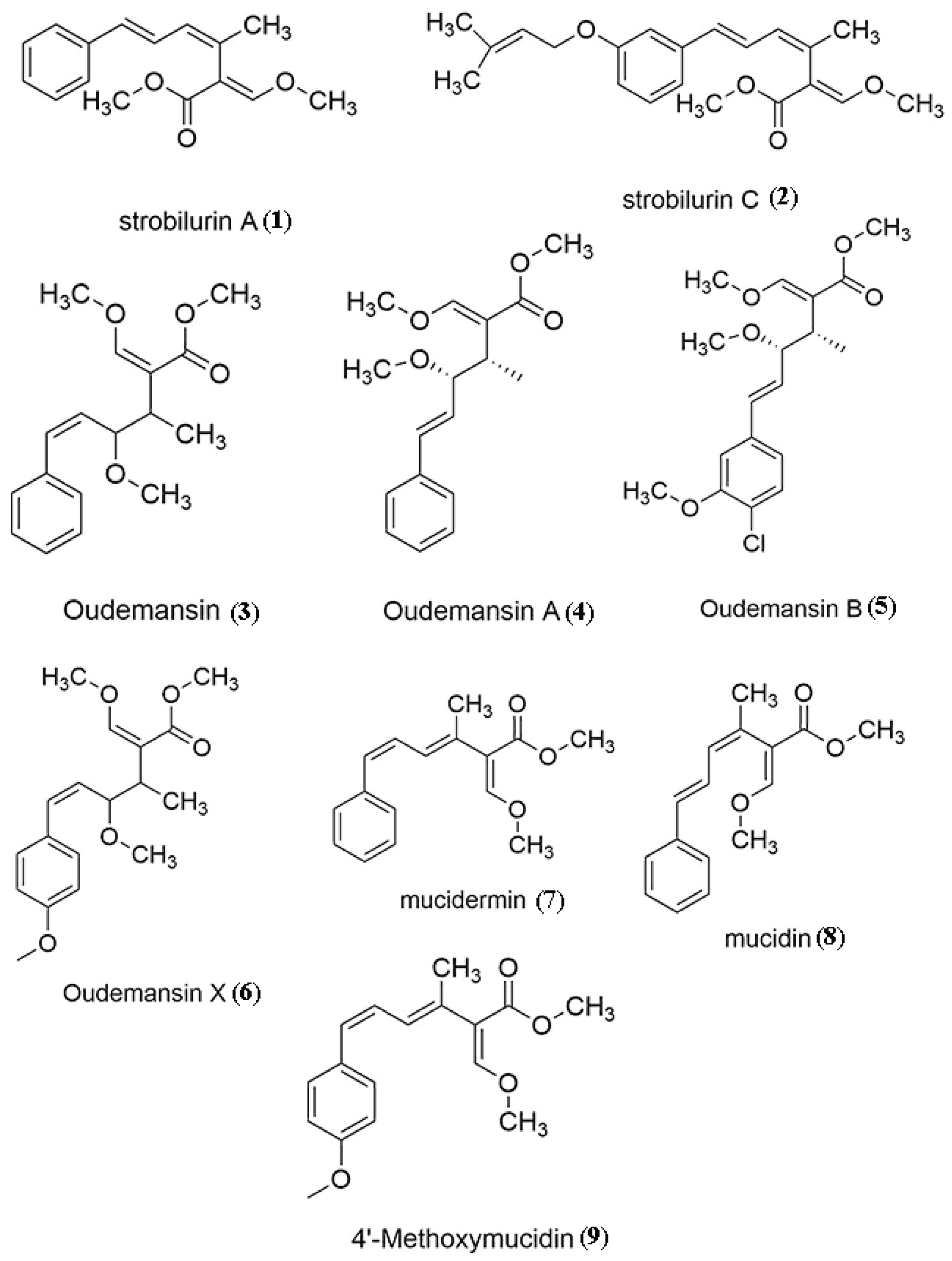
|---|---|---|---|
| Mucidula mucida | Mucidin/strobilurin A/mucidermin | Antifungal | Musilek et al. [77], Anke et al. [98], Subik et al. [104] |
| Strobilurins | Antifungal | Iqbal et al. [99], Anke et al. [98] | |
| Oudemansins | Cytotoxic | Ying et al. [105] | |
| Antifungal | Vondracek [106], Anke et al. [100] | ||
| Strobilurin X, 4’-methoxymucidin | Anke et al. [107] | ||
| Hymenopellis radicata | Oudemansin X | Anke et al. [107] Umezawa et al. [90], Tsantrizos et al. [90,107] | |
| Strobilurins | Antifungal | ||
| Oudemansins | Antihypertensive | ||
| Oudenone | Hemagglutinating activity | Liu et al. [92] | |
| Lectin | Antifungal | Anke et al. [100,104,108] | |
| Mucidin | Antioxidative, anti-inflammatory, lung-protective effects | Gao et al. [91] | |
| SMPS, MPS (mycelia polysaccharides) | Antioxidant; antifungal | Wang et al. [9], Zou [109], Liu et al. [110] | |
| Polysaccharides | Antifungal | Rosa et al. [14,96] | |
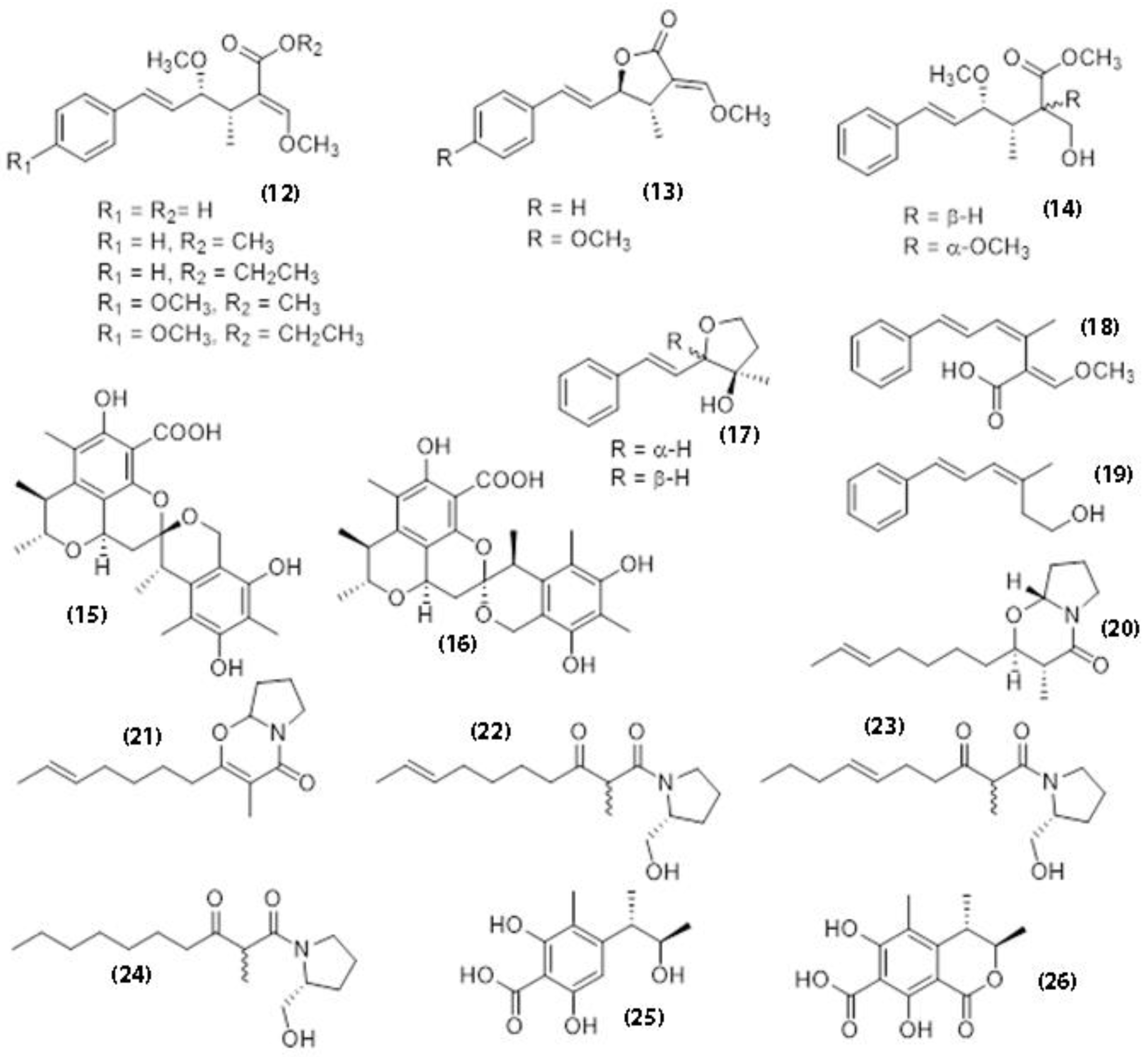
| Oudemansiella canarii | Oudemansin A | Antifungal, inhibitor of eucaryotic respiration | Anke et al. [108] |
| Oudemansiella melanotricha | Oudemansin B, strobilurin C | Inhibitor of cholesterol biosynthesis | Kuhnt & Anke [111] |
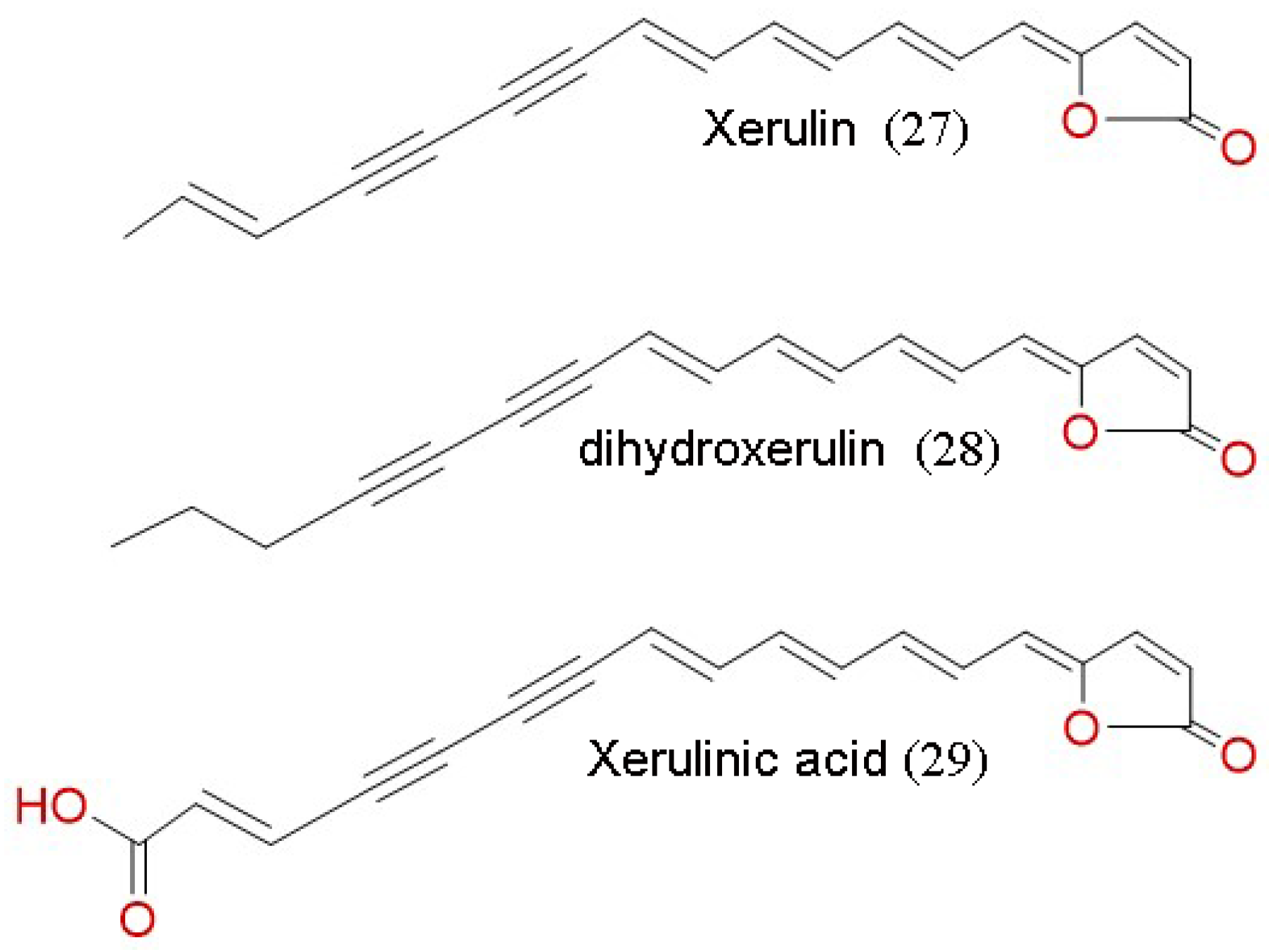
| Xerulin, di-hydro-xerulin, xerulinic acid | Antifungal | Weber et al. [112] | |
| Hydroxy-strobilurin D | Antifungal, inhibitor of eucaryotic respiration | Anke et al. [108] | |
| Xerula longipes | Oudemansin B, strobilurin C | Antifungal | Sivanandhan et al. [88] |
| Xerula pudens | Strobilurin C | Antifungal | Sivanandhan et al. [88] Sadorn et al. [113] |
| Oudemansin B | Antimalarial, antifungal, cytotoxic | ||
| Xerula sp. BCC56836 | Oudemansins | Antibacterial | Sadorn et al. [113] |
| Strobilurin derivatives | Antibacterial, antifungal | ||
| Scalusamides A | Enzyme-inhibitory activity; antifouling activity | ||
| Phenol A acids | Enzyme-inhibitory activity | ||
| Dihydro-citrinone | Antibacterial | ||
| Xerulins | Antibacterial | ||
| Xerucitrinic acid A | Antimicrobial, insecticidal | ||
| 2-(5-Heptenyl)-6,7,8,8a-tetrahydro-3-methyl-4H-pyrrolo [2,1-b][1,3]oxazin-4-one (17) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niego, A.G.; Raspé, O.; Thongklang, N.; Charoensup, R.; Lumyong, S.; Stadler, M.; Hyde, K.D. Taxonomy, Diversity and Cultivation of the Oudemansielloid/Xeruloid Taxa Hymenopellis, Mucidula, Oudemansiella, and Xerula with Respect to Their Bioactivities: A Review. J. Fungi 2021, 7, 51. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010051
Niego AG, Raspé O, Thongklang N, Charoensup R, Lumyong S, Stadler M, Hyde KD. Taxonomy, Diversity and Cultivation of the Oudemansielloid/Xeruloid Taxa Hymenopellis, Mucidula, Oudemansiella, and Xerula with Respect to Their Bioactivities: A Review. Journal of Fungi. 2021; 7(1):51. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010051
Chicago/Turabian StyleNiego, Allen Grace, Olivier Raspé, Naritsada Thongklang, Rawiwan Charoensup, Saisamorn Lumyong, Marc Stadler, and Kevin D. Hyde. 2021. "Taxonomy, Diversity and Cultivation of the Oudemansielloid/Xeruloid Taxa Hymenopellis, Mucidula, Oudemansiella, and Xerula with Respect to Their Bioactivities: A Review" Journal of Fungi 7, no. 1: 51. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010051