In Vitro and In Vivo Effect of Peptides Derived from 14-3-3 Paracoccidioides spp. Protein

 , ,
, ,  , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Prediction of Binding Peptides from Paracoccidioides spp. 14-3-3 Protein to Major Histocompatibility Complex (MHC) Class II Mouse Alleles
2.2. Microorganisms and Culture Conditions
2.3. Cytotoxicity of 14-3-3 Peptides
2.4. Antifungal Activity of 14-3-3 Peptides
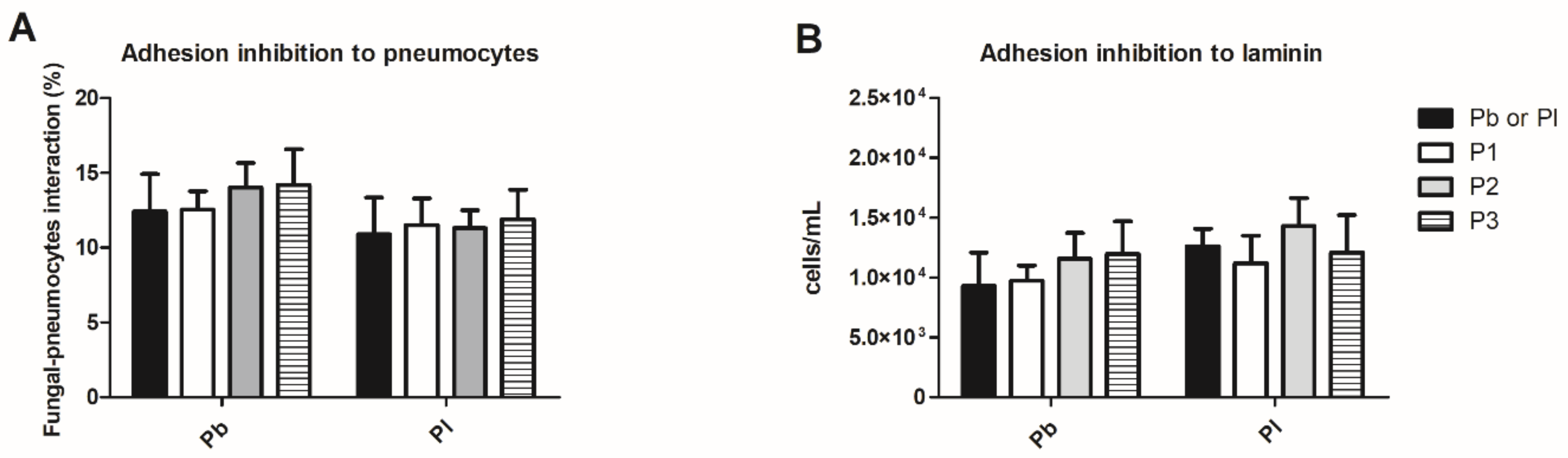
2.5. Paracoccidioides spp. Adhesion Inhibition Assay to the Extracellular Matrix Component or Pneumocytes (A549) Cells
2.6. G. mellonella Rearing and Experiment Conditions
2.6.1. Toxicity Evaluation of 14-3-3 Peptides
2.6.2. Hemocytes Concentration
2.6.3. Induction of Phenoloxidase Activity by Peptides
2.6.4. Analysis of the Expression of Antimicrobial Peptides
2.6.5. Efficacy of 14-3-3 Peptides in G. mellonella Model
2.7. Effect of 14-3-3 Peptides on the Induction of C. elegans Antimicrobial Peptides
2.8. Statistical Analysis
3. Results
3.1. In Silico Prediction of Binding Peptides from Paracoccidioides spp. 14-3-3 Protein to Mouse MHC Class II Alleles
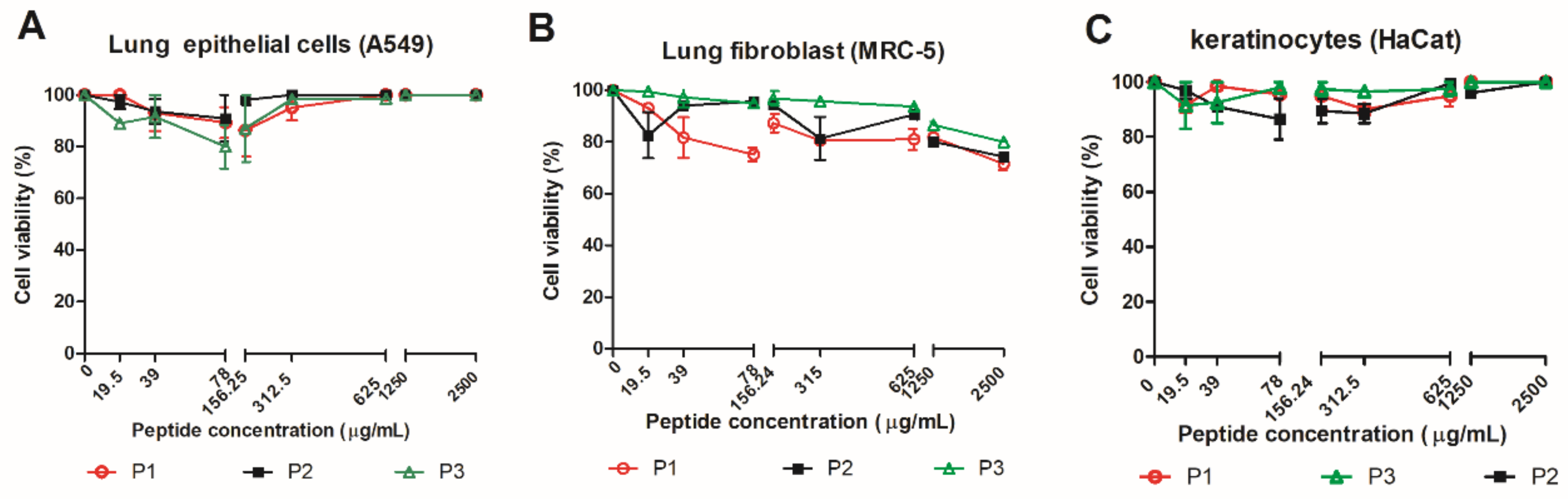
3.2. Effect of 14-3-3 Peptides in Mammalian Cell
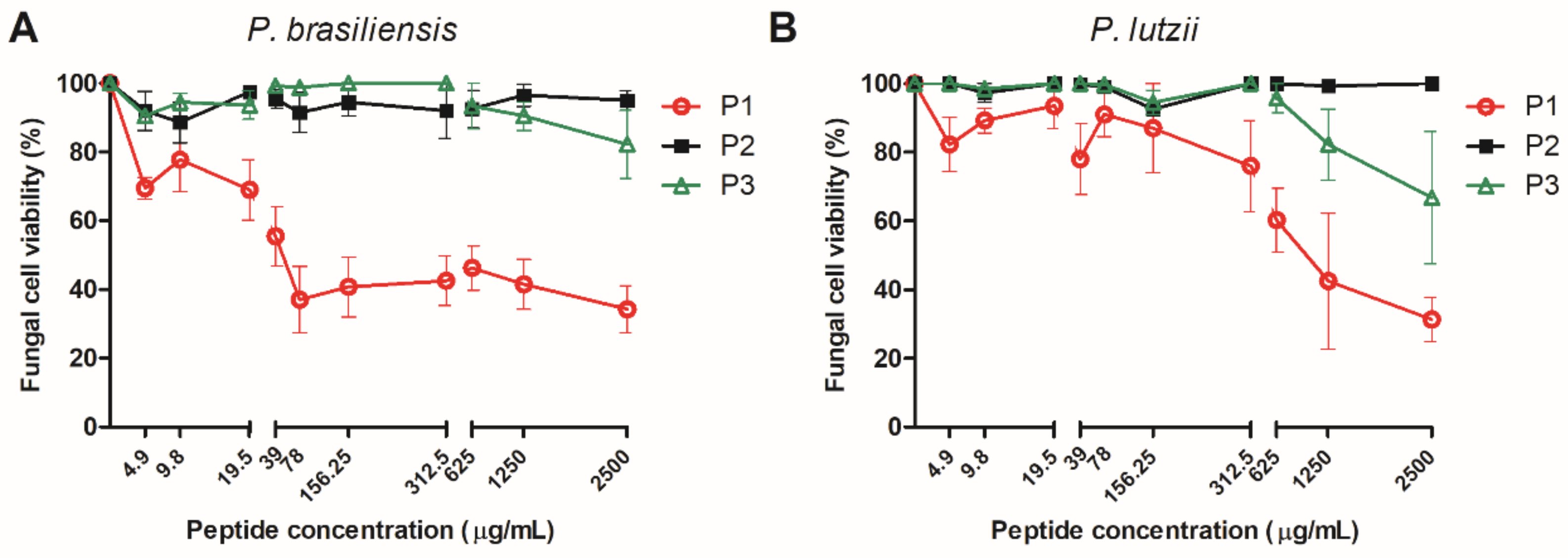
3.3. Effect of 14-3-3 Peptides on Paracoccidioides spp. Cells
3.4. Evaluation of the Immunomodulatory and Antifungal Effect in G. mellonella
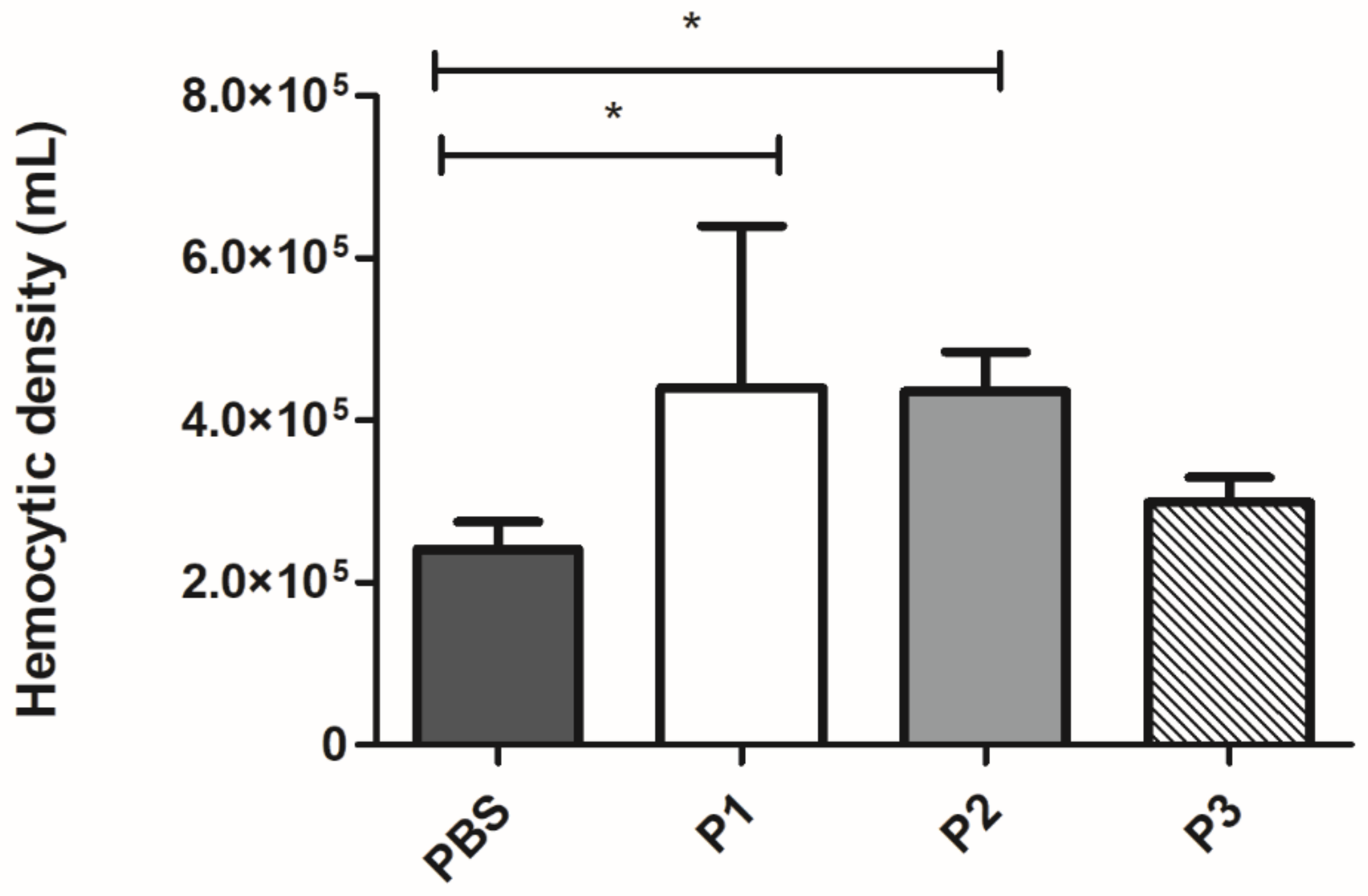
3.4.1. Effect of 14-3-3 Peptides in Hemocytes Concentration
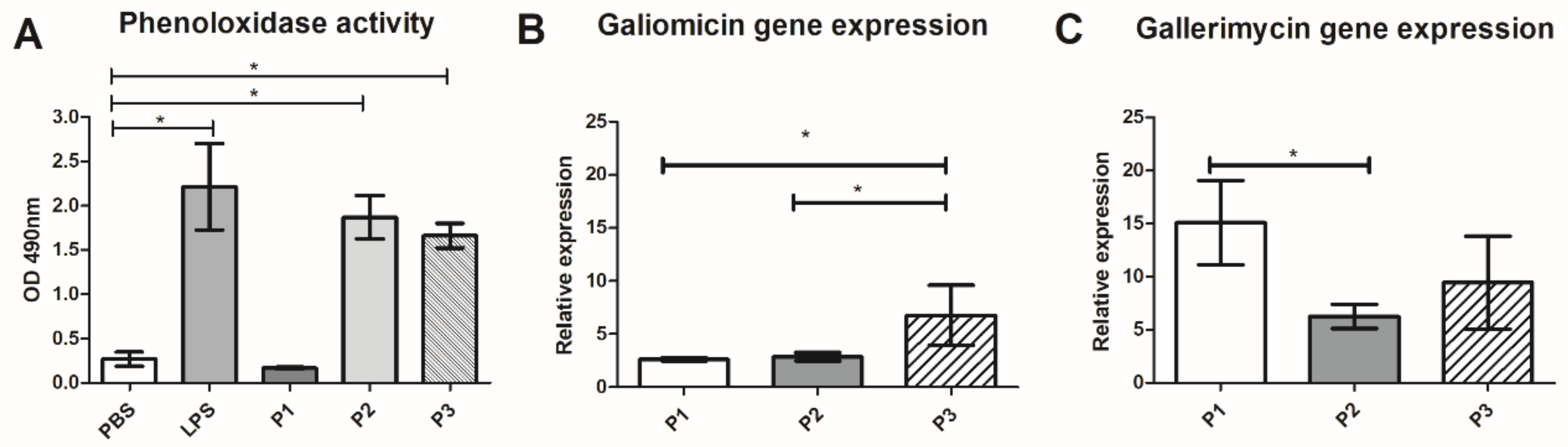
3.4.2. Effect of 14-3-3 Protein Peptides in Humoral Response of G. mellonella
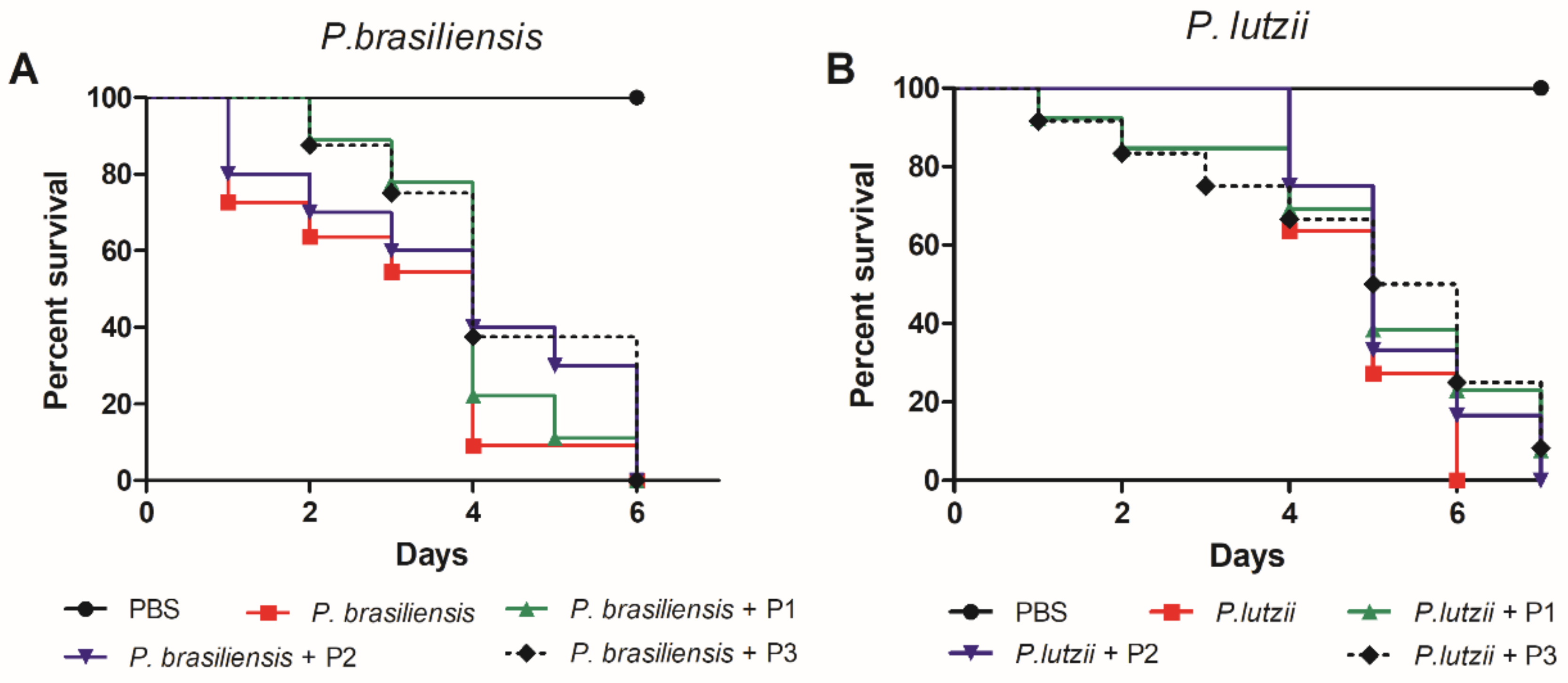
3.4.3. Antifungal Protective Effect of 14-3-3 Protein Peptides against Paracoccidioides spp. Infection
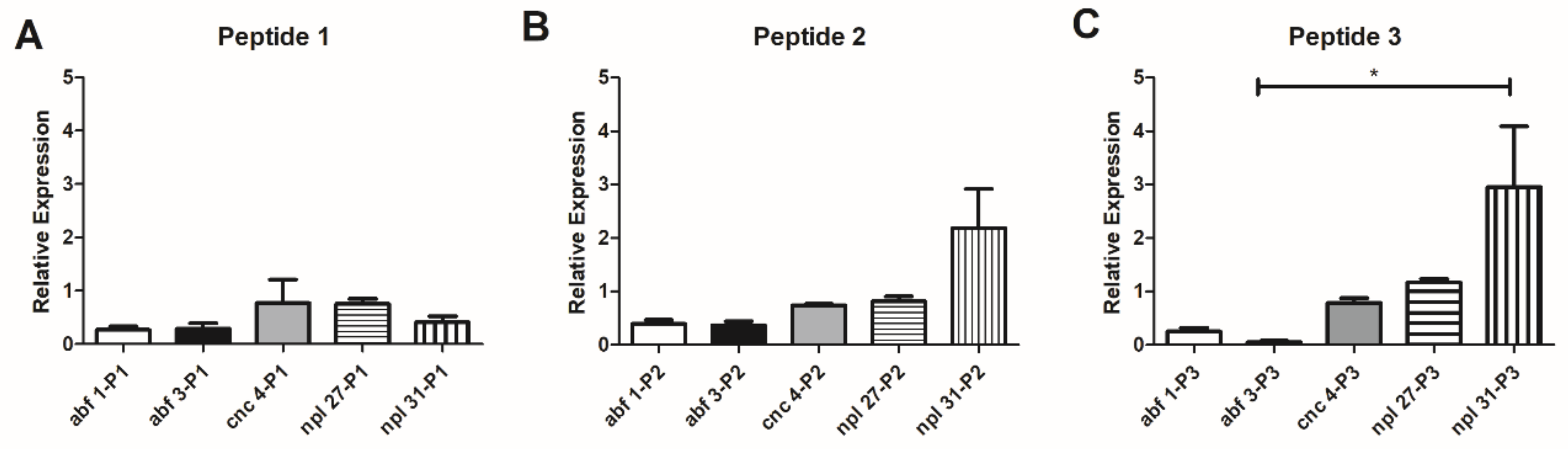
3.5. Evaluation of C. elegans Antimicrobial Peptide Expression after Treatment with 14-3-3 Peptides
4. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [Green Version]
- Jampilek, J. How can we bolster the antifungal drug discovery pipeline? Future Med. Chem. 2016, 8, 1393–1397. [Google Scholar] [CrossRef]
- Teixeira, M.M.; Theodoro, R.C.; De Carvalho, M.J.; Fernandes, L.; Paes, H.C.; Hahn, R.C.; Mendoza, L.; Bagagli, E.; San-Blas, G.; Felipe, M.S. Phylogenetic analysis reveals a high level of speciation in the Paracoccidioides genus. Mol. Phylogenet. Evol. 2009, 52, 273–283. [Google Scholar] [CrossRef]
- Munoz, J.F.; Farrer, R.A.; Desjardins, C.A.; Gallo, J.E.; Sykes, S.; Sakthikumar, S.; Misas, E.; Whiston, E.A.; Bagagli, E.; Soares, C.M.; et al. Genome Diversity, Recombination, and Virulence across the Major Lineages of Paracoccidioides. mSphere 2016, 1, e00213-16. [Google Scholar] [CrossRef] [Green Version]
- Turissini, D.A.; Gomez, O.M.; Teixeira, M.M.; McEwen, J.G.; Matute, D.R. Species boundaries in the human pathogen Paracoccidioides. Fungal Genet. Biol. 2017, 106, 9–25. [Google Scholar] [CrossRef]
- Martinez, R. New Trends in Paracoccidioidomycosis Epidemiology. J. Fungi 2017, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; Valle, A.C.F.D.; Bagagli, E.; Benard, G.; et al. Brazilian guidelines for the clinical management of paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2017, 50, 715–740. [Google Scholar] [CrossRef]
- Martinez, R. Epidemiology of paracoccidioidomycosis. Rev. Inst. Med. Trop. Sao Paulo 2015, 57 (Suppl. 19), 11–20. [Google Scholar] [CrossRef] [Green Version]
- Filler, S.G.; Sheppard, D.C. Fungal invasion of normally non-phagocytic host cells. PLoS Pathog. 2006, 2, e129. [Google Scholar] [CrossRef]
- Pigosso, L.L.; Parente, A.F.; Coelho, A.S.; Silva, L.P.; Borges, C.L.; Bailão, A.M.; Soares, C.M. Comparative proteomics in the genus Paracoccidioides. Fungal Genet. Biol. 2013, 60, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Mendes-Giannini, M.J.; Hanna, S.A.; Da Silva, J.L.; Andreotti, P.F.; Vincenzi, L.R.; Benard, G.; Lenzi, H.L.; Soares, C.P. Invasion of epithelial mammalian cells by Paracoccidioides brasiliensis leads to cytoskeletal rearrangement and apoptosis of the host cell. Microbes Infect. 2004, 6, 882–891. [Google Scholar] [CrossRef]
- Marcos, C.M.; De Fátima da Silva, J.; De Oliveira, H.C.; Moraes da Silva, R.A.; Mendes-Giannini, M.J.; Fusco-Almeida, A.M. Surface-expressed enolase contributes to the adhesion of Paracoccidioides brasiliensis to host cells. FEMS Yeast Res. 2012, 12, 557–570. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, J.d.F.; De Oliveira, H.C.; Marcos, C.M.; Da Silva, R.A.; Da Costa, T.A.; Calich, V.L.; Almeida, A.M.; Mendes-Giannini, M.J. Paracoccidoides brasiliensis 30 kDa Adhesin: Identification as a 14-3-3 Protein, Cloning and Subcellular Localization in Infection Models. PLoS ONE 2013, 8, e62533. [Google Scholar] [CrossRef]
- Nogueira, S.V.; Fonseca, F.L.; Rodrigues, M.L.; Mundodi, V.; Abi-Chacra, E.A.; Winters, M.S.; Alderete, J.F.; De Almeida Soares, C.M. Paracoccidioides brasiliensis enolase is a surface protein that binds plasminogen and mediates interaction of yeast forms with host cells. Infect. Immun. 2010, 78, 4040–4050. [Google Scholar] [CrossRef] [Green Version]
- Torres, I.; Hernandez, O.; Tamayo, D.; Muñoz, J.F.; Leitão, N.P.; García, A.M.; Restrepo, A.; Puccia, R.; McEwen, J.G. Inhibition of PbGP43 expression may suggest that gp43 is a virulence factor in Paracoccidioides brasiliensis. PLoS ONE 2013, 8, e68434. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Neto, B.R.; De Fátima da Silva, J.; Mendes-Giannini, M.J.; Lenzi, H.L.; De Almeida Soares, C.M.; Pereira, M. The malate synthase of Paracoccidioides brasiliensis is a linked surface protein that behaves as an anchorless adhesin. BMC Microbiol. 2009, 9, 272. [Google Scholar] [CrossRef] [Green Version]
- Assato, P.A.; Da Silva, J.d.F.; De Oliveira, H.C.; Marcos, C.M.; Rossi, D.; Valentini, S.R.; Mendes-Giannini, M.J.; Zanelli, C.F.; Fusco-Almeida, A.M. Functional analysis of Paracoccidioides brasiliensis 14-3-3 adhesin expressed in Saccharomyces cerevisiae. BMC Microbiol. 2015, 15, 256. [Google Scholar] [CrossRef] [Green Version]
- Marcos, C.M.; Da Silva, J.d.F.; De Oliveira, H.C.; Assato, P.A.; Singulani, J.L.; Lopez, A.M.; Tamayo, D.P.; Hernandez-Ruiz, O.; McEwen, J.G.; Mendes-Giannini, M.J.; et al. Decreased expression of 14-3-3 in Paracoccidioides brasiliensis confirms its involvement in fungal pathogenesis. Virulence 2016, 16, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Marcos, C.M.; De Oliveira, H.C.; Assato, P.A.; De Andrade, C.R.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Paracoccidioides brasiliensis 14-3-3 protein is important for virulence in a murine model. Med. Mycol. 2019, 57, 900–904. [Google Scholar] [CrossRef]
- Rossi, S.A.; Trevijano-Contador, N.; Scorzoni, L.; Mesa-Arango, A.C.; De Oliveira, H.C.; Werther, K.; De Freitas Raso, T.; Mendes-Giannini, M.J.; Zaragoza, O.; Fusco-Almeida, A.M. Impact of Resistance to Fluconazole on Virulence and Morphological Aspects of Cryptococcus neoformans and Cryptococcus gattii Isolates. Front. Microbiol. 2016, 7, 153. [Google Scholar] [CrossRef]
- Gago, S.; García-Rodas, R.; Cuesta, I.; Mellado, E.; Alastruey-Izquierdo, A. Candida parapsilosis, Candida orthopsilosis, and Candida metapsilosis virulence in the non-conventional host Galleria mellonella. Virulence 2014, 5, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Scorzoni, L.; Alves de Paula e Silva, A.C.; Singulani, J.d.L.; Leite, F.S.; De Oliveira, H.C.; Moraes da Silva, R.A.; Fusco-Almeida, A.M.; Soares Mendes-Giannini, M.J. Comparison of virulence between Paracoccidioides brasiliensis and Paracoccidioides lutzii using Galleria mellonella as a host model. Virulence 2015, 6, 766–776. [Google Scholar] [CrossRef] [Green Version]
- Maurer, E.; Browne, N.; Surlis, C.; Jukic, E.; Moser, P.; Kavanagh, K.; Lass-Flörl, C.; Binder, U. Galleria mellonella as a host model to study Aspergillus terreus virulence and amphotericin B resistance. Virulence 2015, 6, 591–598. [Google Scholar] [CrossRef] [Green Version]
- Maurer, E.; Hortnagl, C.; Lackner, M.; Grassle, D.; Naschberger, V.; Moser, P.; Segal, E.; Semis, M.; Lass-Florl, C.; Binder, U. Galleria mellonella as a model system to study virulence potential of mucormycetes and evaluation of antifungal treatment. Med. Mycol. 2019, 57, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Pereira, T.C.; De Barros, P.P.; Fugisaki, L.R.O.; Rossoni, R.D.; Ribeiro, F.C.; De Menezes, R.T.; Junqueira, J.C.; Scorzoni, L. Recent Advances in the Use of Galleria mellonella Model to Study Immune Responses against Human Pathogens. J. Fungi 2018, 4, 128. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, G.; Kavanagh, K. Analysis of the early cellular and humoral responses of Galleria mellonella larvae to infection by Candida albicans. Virulence 2018, 9, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Browne, N.; Heelan, M.; Kavanagh, K. An analysis of the structural and functional similarities of insect hemocytes and mammalian phagocytes. Virulence 2013, 4, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Wojda, I. Immunity of the greater wax moth Galleria mellonella. Insect Sci. 2017, 24, 342–357. [Google Scholar] [CrossRef]
- Souza, A.C.R.; Fuchs, B.B.; Alves, V.S.; Jayamani, E.; Colombo, A.L.; Mylonakis, E. Pathogenesis of the Candida parapsilosis Complex in the Model Host Caenorhabditis elegans. Genes 2018, 9, 401. [Google Scholar] [CrossRef] [Green Version]
- Desalermos, A.; Tan, X.; Rajamuthiah, R.; Arvanitis, M.; Wang, Y.; Li, D.; Kourkoumpetis, T.K.; Fuchs, B.B.; Mylonakis, E. A multi-host approach for the systematic analysis of virulence factors in Cryptococcus neoformans. J. Infect. Dis. 2015, 211, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Zaborin, A.; Romanowski, K.; Gerdes, S.; Holbrook, C.; Lepine, F.; Long, J.; Poroyko, V.; Diggle, S.P.; Wilke, A.; Righetti, K.; et al. Red death in Caenorhabditis elegans caused by Pseudomonas aeruginosa PAO1. Proc. Natl. Acad. Sci. USA 2009, 106, 6327–6332. [Google Scholar] [CrossRef] [Green Version]
- Espinal, P.; Pantel, A.; Rolo, D.; Marti, S.; Lopez-Rojas, R.; Smani, Y.; Pachon, J.; Vila, J.; Lavigne, J.P. Relationship Between Different Resistance Mechanisms and Virulence in Acinetobacter baumannii. Microb. Drug Resist. 2019, 25, 752–760. [Google Scholar] [CrossRef]
- Manoharan, R.K.; Lee, J.H.; Kim, Y.G.; Kim, S.I.; Lee, J. Inhibitory effects of the essential oils alpha-longipinene and linalool on biofilm formation and hyphal growth of Candida albicans. Biofouling 2017, 33, 143–155. [Google Scholar] [CrossRef]
- De Barros, P.P.; Scorzoni, L.; Ribeiro, F.C.; Fugisaki, L.R.O.; Fuchs, B.B.; Mylonakis, E.; Jorge, A.O.C.; Junqueira, J.C.; Rossoni, R.D. Lactobacillus paracasei 28.4 reduces in vitro hyphae formation of Candida albicans and prevents the filamentation in an experimental model of Caenorhabditis elegans. Microb. Pathog. 2018, 117, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Madende, M.; Albertyn, J.; Sebolai, O.; Pohl, C.H. Caenorhabditis elegans as a model animal for investigating fungal pathogenesis. Med. Microbiol. Immunol. 2020, 209, 1–13. [Google Scholar] [CrossRef]
- Scorzoni, L.; De Lucas, M.P.; Singulani, J.L.; De Oliveira, H.C.; Assato, P.A.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Evaluation of Caenorhabditis elegans as a host model for Paracoccidioides brasiliensis and Paracoccidioides lutzii. Pathog. Dis. 2018, 76. [Google Scholar] [CrossRef]
- Mallo, G.V.; Kurz, C.L.; Couillault, C.; Pujol, N.; Granjeaud, S.; Kohara, Y.; Ewbank, J.J. Inducible antibacterial defense system in C. elegans. Curr. Biol. 2002, 12, 1209–1214. [Google Scholar] [CrossRef] [Green Version]
- Muhammed, M.; Fuchs, B.B.; Wu, M.P.; Breger, J.; Coleman, J.J.; Mylonakis, E. The role of mycelium production and a MAPK-mediated immune response in the C. elegans-Fusarium model system. Med. Mycol. 2012, 50, 488–496. [Google Scholar] [CrossRef] [Green Version]
- Ong, C.W.; Chen, S.C.; Clark, J.E.; Halliday, C.L.; Kidd, S.E.; Marriott, D.J.; Marshall, C.L.; Morris, A.J.; Morrissey, C.O.; Roy, R.; et al. Diagnosis, management and prevention of Candida auris in hospitals: Position statement of the Australasian Society for Infectious Diseases. Intern. Med. J. 2019. [Google Scholar] [CrossRef]
- Benedict, K.; Jackson, B.R.; Chiller, T.; Beer, K.D. Estimation of Direct Healthcare Costs of Fungal Diseases in the United States. Clin. Infect. Dis. 2019, 68, 1791–1797. [Google Scholar] [CrossRef] [Green Version]
- Hendrickson, J.A.; Hu, C.; Aitken, S.L.; Beyda, N. Antifungal Resistance: A Concerning Trend for the Present and Future. Curr. Infect. Dis. Rep. 2019, 21, 47. [Google Scholar] [CrossRef]
- Perfect, J.R. The antifungal pipeline: A reality check. Nat. Rev. Drug Discov. 2017, 16, 603–616. [Google Scholar] [CrossRef] [Green Version]
- Pavan, F.R.; Da S Maia, P.I.; Leite, S.R.; Deflon, V.M.; Batista, A.A.; Sato, D.N.; Franzblau, S.G.; Leite, C.Q. Thiosemicarbazones, semicarbazones, dithiocarbazates and hydrazide/hydrazones: Anti-Mycobacterium tuberculosis activity and cytotoxicity. Eur. J. Med. Chem. 2010, 45, 1898–1905. [Google Scholar] [CrossRef]
- De Paula e Silva, A.C.; Oliveira, H.C.; Silva, J.F.; Sangalli-Leite, F.; Scorzoni, L.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J. Microplate alamarBlue assay for Paracoccidioides susceptibility testing. J. Clin. Microbiol. 2013, 51, 1250–1252. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, H.C.; Da Silva, J.d.F.; Scorzoni, L.; Marcos, C.M.; Rossi, S.A.; De Paula E Silva, A.C.; Assato, P.A.; Da Silva, R.A.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J. Importance of adhesins in virulence of Paracoccidioides spp. Front. Microbiol. 2015, 6, 303. [Google Scholar] [CrossRef]
- Jorjão, A.L.; Oliveira, L.D.; Scorzoni, L.; Figueiredo-Godoi, L.M.A.; Prata, M.C.A.; Jorge, A.O.C.; Junqueira, J.C. From moths to caterpillars: Ideal conditions for Galleria mellonella rearing for in vivo microbiological studies. Virulence 2018, 9, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Laughton, A.M.; Siva-Jothy, M.T. A standardised protocol for measuring phenoloxidase and prophenoloxidase in the honey bee, Apis mellifera. Apidologie 2011, 42, 140–149. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-ΔΔC(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wojda, I.; Jakubowicz, T. Humoral immune response upon mild heat-shock conditions in Galleria mellonella larvae. J. Insect Physiol. 2007, 53, 1134–1144. [Google Scholar] [CrossRef]
- Bergin, D.; Murphy, L.; Keenan, J.; Clynes, M.; Kavanagh, K. Pre-exposure to yeast protects larvae of Galleria mellonella from a subsequent lethal infection by Candida albicans and is mediated by the increased expression of antimicrobial peptides. Microbes Infect. 2006, 8, 2105–2112. [Google Scholar] [CrossRef] [Green Version]
- Alper, S.; McBride, S.J.; Lackford, B.; Freedman, J.H.; Schwartz, D.A. Specificity and complexity of the Caenorhabditis elegans innate immune response. Mol. Cell. Biol. 2007, 27, 5544–5553. [Google Scholar] [CrossRef] [Green Version]
- Zugasti, O.; Ewbank, J.J. Neuroimmune regulation of antimicrobial peptide expression by a noncanonical TGF-beta signaling pathway in Caenorhabditis elegans epidermis. Nat. Immunol. 2009, 10, 249–256. [Google Scholar] [CrossRef]
- Pujol, N.; Zugasti, O.; Wong, D.; Couillault, C.; Kurz, C.L.; Schulenburg, H.; Ewbank, J.J. Anti-fungal innate immunity in C. elegans is enhanced by evolutionary diversification of antimicrobial peptides. PLoS Pathog. 2008, 4, e1000105. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.D.; Denning, D.W.; Levitz, S.M. Tackling Human Fungal Infections. Science 2012, 336, 647. [Google Scholar] [CrossRef] [Green Version]
- Boniche, C.; Rossi, S.A.; Kischkel, B.; Barbalho, F.V.; Moura, A.N.D.; Nosanchuk, J.D.; Travassos, L.R.; Taborda, C.P. Immunotherapy against Systemic Fungal Infections Based on Monoclonal Antibodies. J. Fungi 2020, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Travassos, L.R.; Taborda, C.P. Linear Epitopes of Paracoccidioides brasiliensis and Other Fungal Agents of Human Systemic Mycoses As Vaccine Candidates. Front. Immunol. 2017, 8, 224. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, L.B.R.; Taborda, C.P.; Nosanchuk, J.D. Advances in Fungal Peptide Vaccines. J. Fungi 2020, 6, 119. [Google Scholar] [CrossRef]
- Kischkel, B.; Rossi, S.A.; Santos, S.R.; Nosanchuk, J.D.; Travassos, L.R.; Taborda, C.P. Therapies and Vaccines Based on Nanoparticles for the Treatment of Systemic Fungal Infections. Front. Cell. Infect. Microbiol. 2020, 10, 463. [Google Scholar] [CrossRef]
- Rittner, G.M.; Muñoz, J.E.; Marques, A.F.; Nosanchuk, J.D.; Taborda, C.P.; Travassos, L.R. Therapeutic DNA vaccine encoding peptide P10 against experimental paracoccidioidomycosis. PLoS Negl. Trop. Dis. 2012, 6, e1519. [Google Scholar] [CrossRef] [Green Version]
- Bergin, D.; Brennan, M.; Kavanagh, K. Fluctuations in haemocyte density and microbial load may be used as indicators of fungal pathogenicity in larvae of Galleria mellonella. Microbes Infect. 2003, 5, 1389–1395. [Google Scholar] [CrossRef] [Green Version]
- Fallon, J.P.; Troy, N.; Kavanagh, K. Pre-exposure of Galleria mellonella larvae to different doses of Aspergillus fumigatus conidia causes differential activation of cellular and humoral immune responses. Virulence 2011, 2, 413–421. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, H.C.; Michaloski, J.S.; Da Silva, J.d.F.; Scorzoni, L.; De Paula E Silva, A.C.; Marcos, C.M.; Assato, P.A.; Yamazaki, D.S.; Fusco-Almeida, A.M.; Giordano, R.J.; et al. Peptides Derived from a Phage Display Library Inhibit Adhesion and Protect the Host against Infection by Paracoccidioides brasiliensis and Paracoccidioides lutzii. Front. Pharm. 2016, 7, 509. [Google Scholar] [CrossRef]
- Gandra, R.M.; McCarron, P.; Viganor, L.; Fernandes, M.F.; Kavanagh, K.; McCann, M.; Branquinha, M.H.; Santos, A.L.S.; Howe, O.; Devereux, M. In vivo Activity of Copper(II), Manganese(II), and Silver(I) 1,10-Phenanthroline Chelates Against Candida haemulonii Using the Galleria mellonella Model. Front. Microbiol. 2020, 11, 470. [Google Scholar] [CrossRef] [Green Version]
- Rossoni, R.D.; De Barros, P.P.; Mendonca, I.D.C.; Medina, R.P.; Silva, D.H.S.; Fuchs, B.B.; Junqueira, J.C.; Mylonakis, E. The Postbiotic Activity of Lactobacillus paracasei 28.4 Against Candida auris. Front Cell Infect. Microbiol. 2020, 10, 397. [Google Scholar] [CrossRef]
- Cerenius, L.; Lee, B.L.; Soderhall, K. The proPO-system: Pros and cons for its role in invertebrate immunity. Trends Immunol. 2008, 29, 263–271. [Google Scholar] [CrossRef]
- Binggeli, O.; Neyen, C.; Poidevin, M.; Lemaitre, B. Prophenoloxidase activation is required for survival to microbial infections in Drosophila. PLoS Pathog. 2014, 10, e1004067. [Google Scholar] [CrossRef]
- Vertyporokh, L.; Wojda, I. Immune response of Galleria mellonella after injection with non-lethal and lethal dosages of Candida albicans. J. Invertebr. Pathol. 2020, 170, 107327. [Google Scholar] [CrossRef]
- Wu, G.; Zhao, Z.; Liu, C.; Qiu, L. Priming Galleria mellonella (Lepidoptera: Pyralidae) larvae with heat-killed bacterial cells induced an enhanced immune protection against Photorhabdus luminescens TT01 and the role of innate immunity in the process. J. Econ. Entomol. 2014, 107, 559–569. [Google Scholar] [CrossRef] [Green Version]
- Barros, P.P.; Rossoni, R.D.; Ribeiro, F.C.; Silva, M.P.; Souza, C.M.; Jorge, A.O.C.; Junqueira, J.C. Two sporulated Bacillus enhance immunity in Galleria mellonella protecting against Candida albicans. Microb. Pathog. 2019, 132, 335–342. [Google Scholar] [CrossRef]
- Ermolaeva, M.A.; Schumacher, B. Insights from the worm: The C. elegans model for innate immunity. Semin. Immunol. 2014, 26, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Pukkila-Worley, R.; Ausubel, F.M.; Mylonakis, E. Candida albicans infection of Caenorhabditis elegans induces antifungal immune defenses. PLoS Pathog. 2011, 7, e1002074. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, A.; Ferreira, K.S.; Almeida, S.R.; Nosanchuk, J.D.; Travassos, L.R.; Taborda, C.P. Prophylactic and therapeutic vaccination using dendritic cells primed with peptide 10 derived from the 43-kilodalton glycoprotein of Paracoccidioides brasiliensis. Clin. Vaccine Immunol. 2012, 19, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, J.E.; Luft, V.D.; Amorim, J.; Magalhães, A.; Thomaz, L.; Nosanchuk, J.D.; Travassos, L.R.; Taborda, C.P. Immunization with P10 peptide increases specific immunity and protects immunosuppressed BALB/c mice infected with virulent yeasts of Paracoccidioides brasiliensis. Mycopathologia 2014, 178, 177–188. [Google Scholar] [CrossRef]
- Silva, L.B.R.; Dias, L.S.; Rittner, G.M.G.; Munoz, J.E.; Souza, A.C.O.; Nosanchuk, J.D.; Travassos, L.R.; Taborda, C.P. Dendritic Cells Primed with Paracoccidioides brasiliensis Peptide P10 Are Therapeutic in Immunosuppressed Mice with Paracoccidioidomycosis. Front. Microbiol. 2017, 8, 1057. [Google Scholar] [CrossRef]
- Silva, L.B.R.; Taira, C.L.; Dias, L.S.; Souza, A.C.O.; Nosanchuk, J.D.; Travassos, L.R.; Taborda, C.P. Experimental Therapy of Paracoccidioidomycosis Using P10-Primed Monocyte-Derived Dendritic Cells Isolated From Infected Mice. Front. Microbiol. 2019, 10, 1727. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | References |
|---|---|---|---|
| S7 | ATG TGC CAA TGC CCA AGT TG | GTG GCT AGG CTT GGG AAG AAT | [49] |
| Galiomicin | TCG TAT CGT CAC CGC AAA ATG | GCC GCA ATG ACC ACC TTT ATA | [50] |
| Gallerimycin | TATCAT TGG CCT TCT TGG CTG | GCA CTCGTA AAA TAC ACA TCC GG | [50] |
| Gene | Forward Primer | Reverse Primer | Reference |
|---|---|---|---|
| abf-1 | GTACAGCACAGAAATGCATGACCGG | GGCGTTTGAACAACCTCCACAGAAGC | [51] |
| abf-3 | GGTGTCGAATAAGGCAGTGTGGACCT | GGCATTTCCATAGCTATCCCTGTAGC | [51] |
| cnc-4 | ACAATGGGGCTACGGTCCATAT | ACTTTCCAATGAGCATTCCGAGGA | [52] |
| nlp-27 | CGGTGGAATGCCATATGGTG | ATCGAATTTACTTTCCCCATCC | [53] |
| nlp-31 | GGTGGATATGGAAGAGGTTATGGAG | GTCTATGCTTTTACTTTCCCC | [53] |
| act-1 | CCATCATGAAGTGCGACATTG | CATGGTTGATGGGGCAAGAG | [53] |
| Peptide | Mammalian Cell Toxicity | G. mellonella Toxicity | Antifungal Activity | Hemocytes Density | Phenoloxidase Activity | Expression G. mellonella Antimicrobial Peptides | Protective effect against Paracoccidioides spp. infection in G. mellonella model | Expression of Antimicrobial Peptides of C. elegans |
|---|---|---|---|---|---|---|---|---|
| P1 | Non-toxic (A549, HACat and MRC-5) | Non-toxic (10–100 µg/larva) | P. brasiliensis (30.5–66% of viability reduction) P. lutzii (22–70% of viability reduction) | 1.8-fold increase | No statistical difference | Galiomicin: 2.6-fold-increase Gallerimycin:15-fold-increase | Delay in the larvae death | abf-1: 0.26-fold-increase abf-3: 0.29-fold- increase cnc-4: 0.77-fold- increase npl-27: 0.75-fold- increase npl-31: 0.40-fold increase |
| P2 | Non-toxic (A549, HACat and MRC-5) | Non-toxic (10–100 µg/larva) | No antifungal activity against Paracoccidioides spp. | 1.8-fold increase | 7-fold increase | Galiomicin: 2.8 fold-increase Gallerimycin: 6.2 fold-increase | Delay in the larvae death | abf-1: 0.39-fold-increase abf-3: 0.36-fold- increase cnc-4: 0.73-fold- increase npl-27: 0.81-fold- increase npl-31: 2.17-fold increase |
| P3 | Non-toxic (A549, HACat and MRC-5) | Non-toxic (10–100 µg/larva) | P. brasiliensis (20% of viability reduction) P. lutzii (30% of viability reduction at 2500 µg/mL) | No statistical difference | 6.3-fold increase | Galiomicin: 6.7 folds-increase Gallerimycin: 6.8-fold increase | Delay in the larvae death | abf-1: 0.20-fold-increase abf-3: 0.05-fold- increase cnc-4: 0.78-fold- increase npl-27: 1.16-fold- increase npl-31: 2.9-fold increase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scorzoni, L.; Alves de Paula e Silva, A.C.; de Oliveira, H.C.; Tavares dos Santos, C.; de Lacorte Singulani, J.; Akemi Assato, P.; Maria Marcos, C.; Teodoro Oliveira, L.; Ferreira Fregonezi, N.; Rossi, D.C.P.; et al. In Vitro and In Vivo Effect of Peptides Derived from 14-3-3 Paracoccidioides spp. Protein. J. Fungi 2021, 7, 52. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010052
Scorzoni L, Alves de Paula e Silva AC, de Oliveira HC, Tavares dos Santos C, de Lacorte Singulani J, Akemi Assato P, Maria Marcos C, Teodoro Oliveira L, Ferreira Fregonezi N, Rossi DCP, et al. In Vitro and In Vivo Effect of Peptides Derived from 14-3-3 Paracoccidioides spp. Protein. Journal of Fungi. 2021; 7(1):52. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010052
Chicago/Turabian StyleScorzoni, Liliana, Ana Carolina Alves de Paula e Silva, Haroldo Cesar de Oliveira, Claudia Tavares dos Santos, Junya de Lacorte Singulani, Patricia Akemi Assato, Caroline Maria Marcos, Lariane Teodoro Oliveira, Nathália Ferreira Fregonezi, Diego Conrado Pereira Rossi, and et al. 2021. "In Vitro and In Vivo Effect of Peptides Derived from 14-3-3 Paracoccidioides spp. Protein" Journal of Fungi 7, no. 1: 52. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010052