Differential miRNA Expression in Human Macrophage-Like Cells Infected with Histoplasma capsulatum Yeasts Cultured in Planktonic and Biofilm Forms

 ,
, 
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. H. capsulatum Strains
2.2. Ethics Statement
2.3. H. capsulatum Planktonic Cultures and Biofilm Formation
2.4. Polysaccharide Extraction and Quantification
2.5. THP-1 Cell Culture and Differentiation
2.6. THP-1 Mø-Like Cell Viability Assay after H. capsulatum Yeast Infection
2.7. THP-1 Cells Infection
2.8. Determination of Infected THP-1 Mø-Like Cells Using Flow Cytometry
2.9. Total RNA Isolation, Quantification, and Integrity Assessment
2.10. qRT-PCR
2.11. Bioinformatics and Statistical Analyses
3. Results
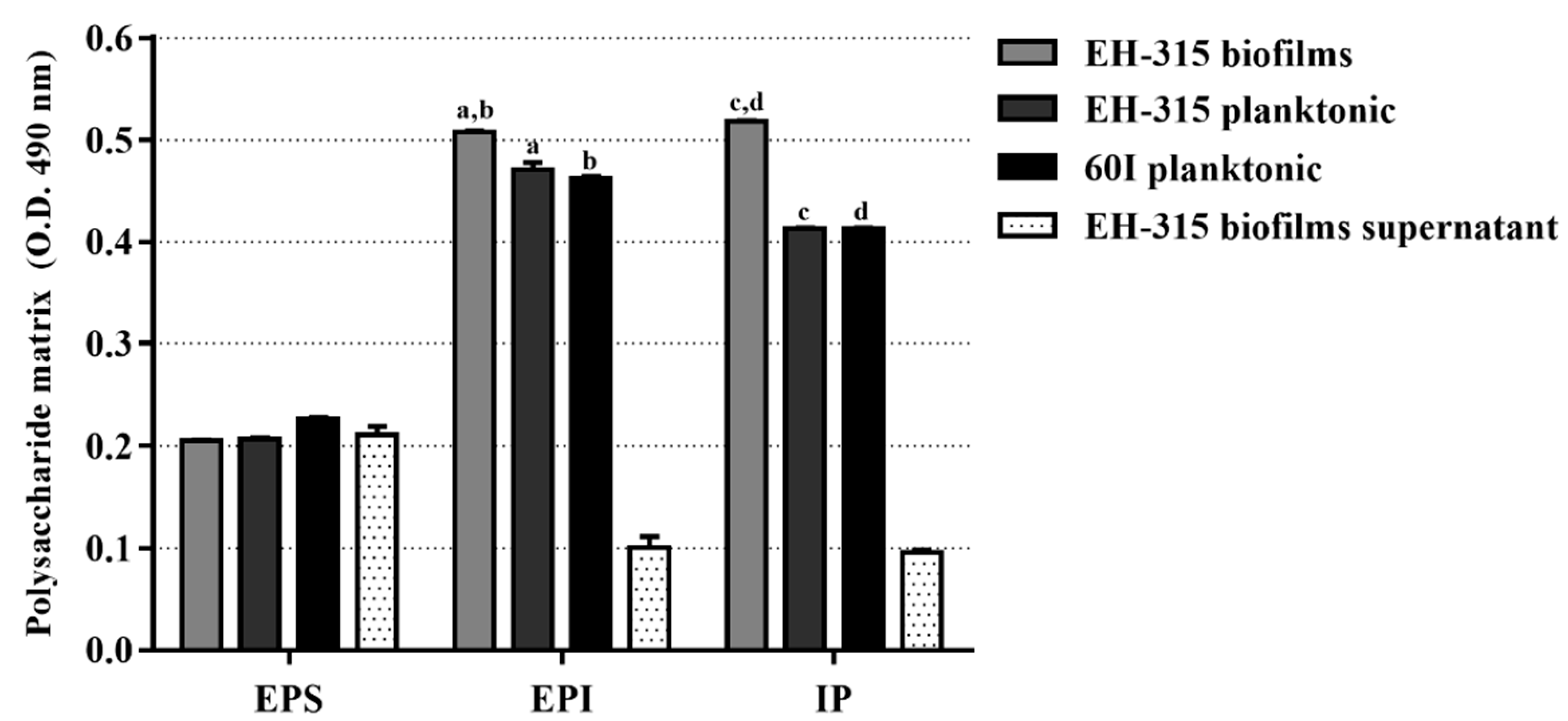
3.1. Polysaccharide Matrix from H. capsulatum Yeasts
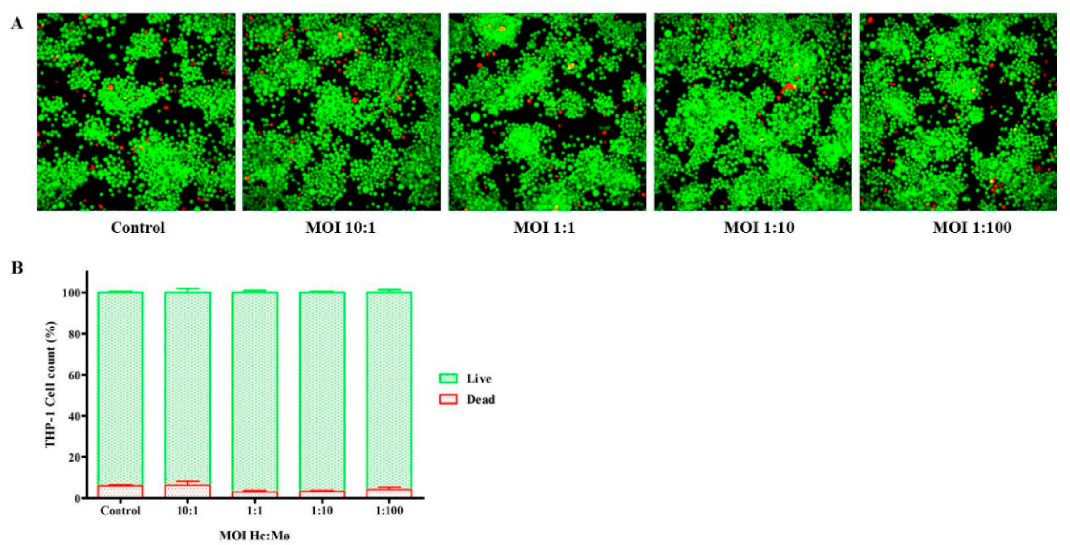
3.2. Viability of Infected THP-1 Mø-Like Cells
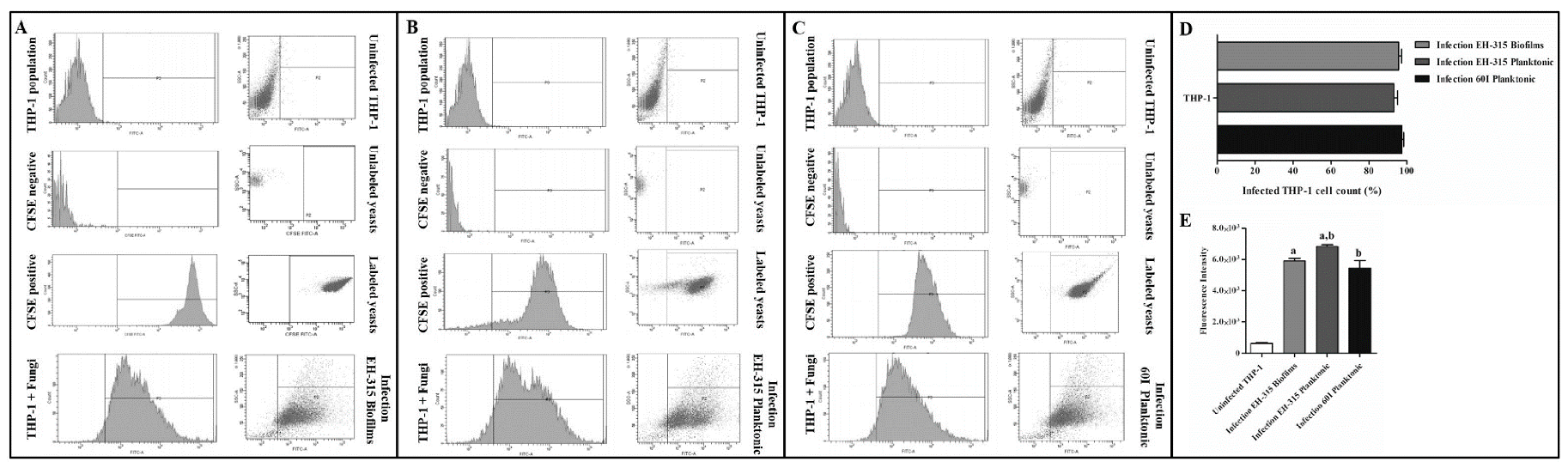
3.3. Determination of Infected THP-1 Mø-Like Cells
3.4. Quantification and Assessment of RNA Samples
3.5. Differential microRNA Expression
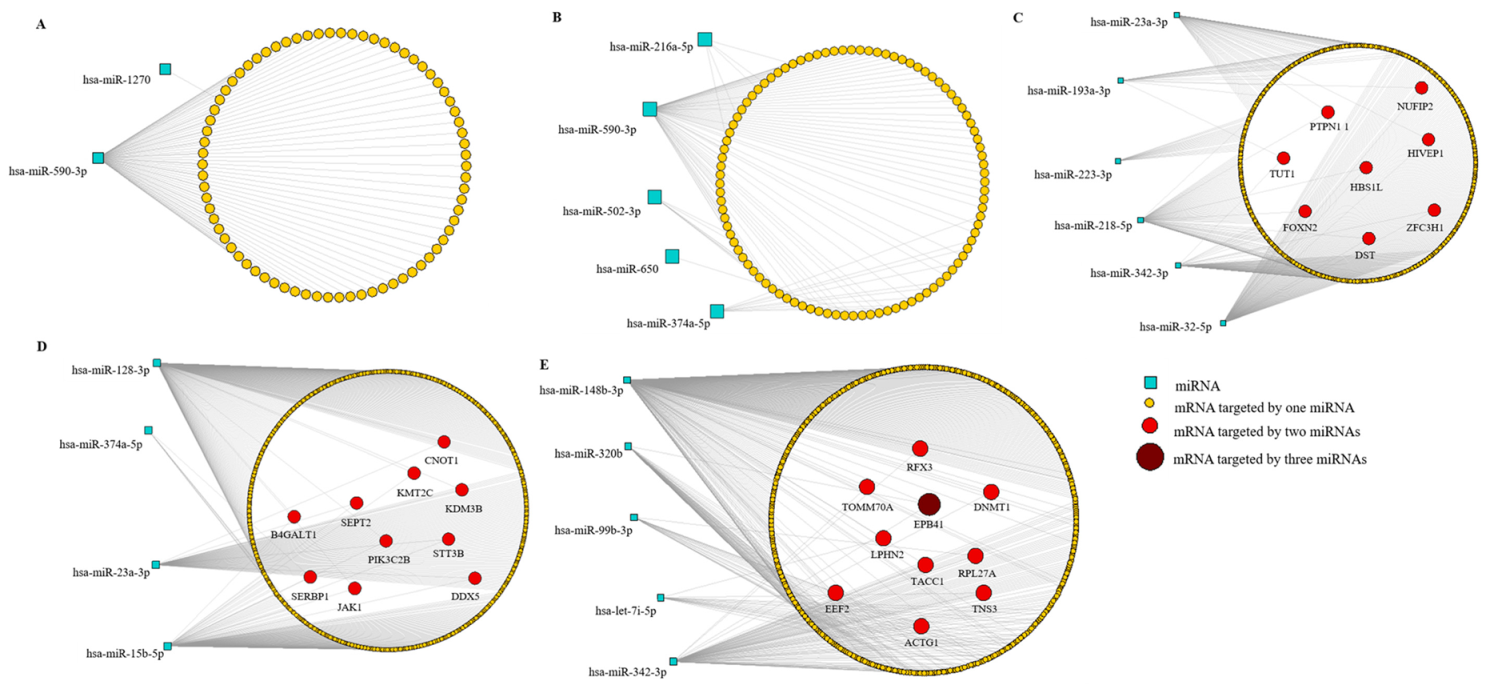
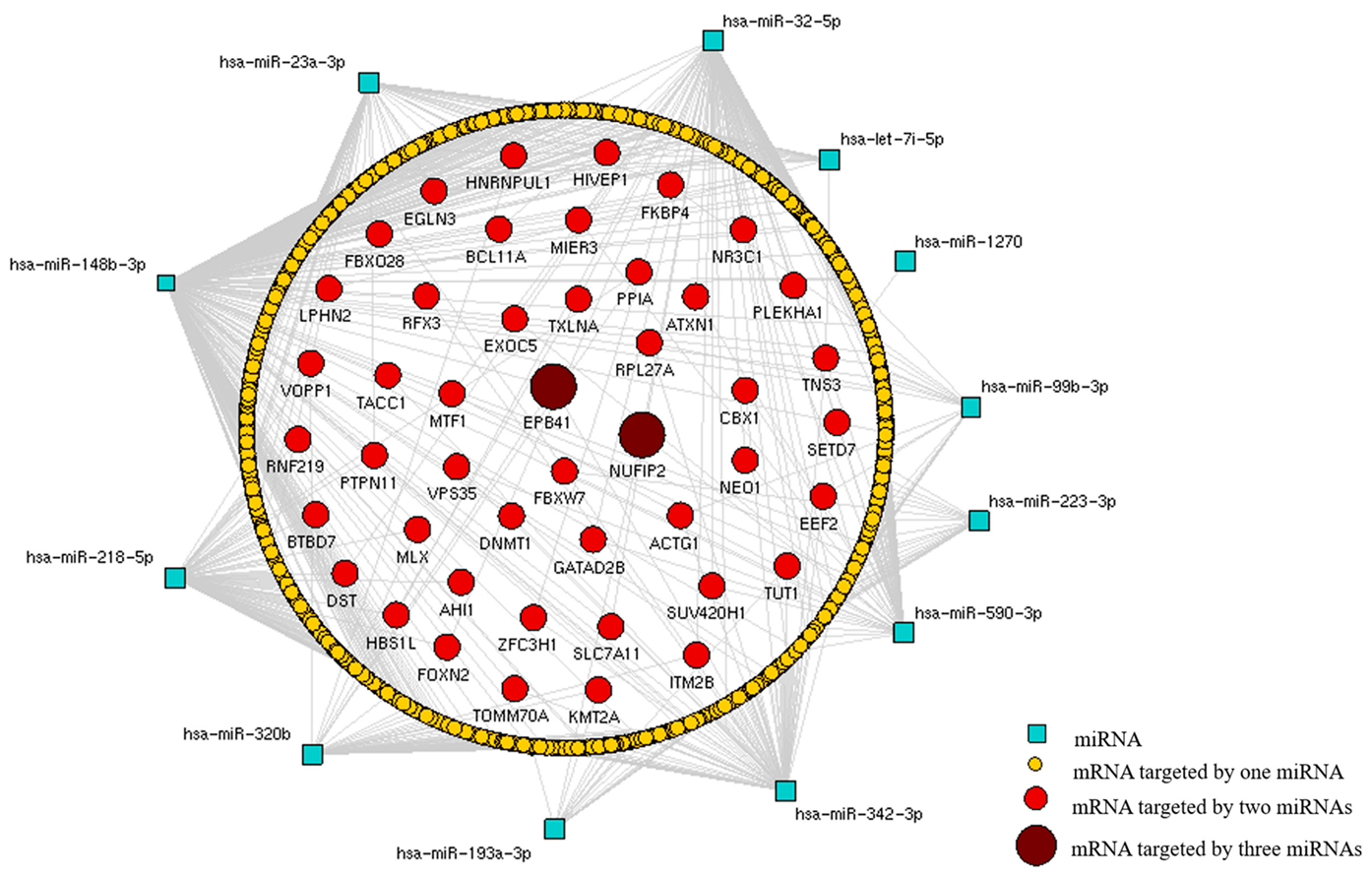
3.6. MiRNA-mRNA Interactions
3.7. In Silico Recognition of Signaling Pathways Hypothetically Regulated by Differentially Expressed miRNAs
3.8. In Silico Detection of Target Genes Regulated by Differentially Expressed miRNAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reyes-Montes, M.R.; Valle, M.B.-D.; Martínez-Rivera, M.A.; Rodríguez-Arellanes, G.; Maravilla, E.; Sifuentes-Osornio, J.; Taylor, M.L. Relatedness Analyses of Histoplasma capsulatum Isolates from Mexican Patients with AIDS-Associated Histoplasmosis by Using Histoplasmin Electrophoretic Profiles and Randomly Amplified Polymorphic DNA Patterns. J. Clin. Microbiol. 1999, 37, 1404–1408. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.L.; Ruíz-Palacios, G.M.; Reyes-Montes, M.R.; Rodríguez-Arellanes, G.; Carreto-Binaghi, L.E.; Duarte-Escalante, E.; Hernández-Ramírez, A.; Pérez, A.; Suã¡rez-Alvarez, R.O.; Suárez-Alvarez, N.Y.A.; et al. Identification of the infectious source of an unusual outbreak of histoplasmosis, in a hotel in Acapulco, state of Guerrero, Mexico. FEMS Immunol. Med. Microbiol. 2005, 45, 435–441. [Google Scholar] [CrossRef] [Green Version]
- Samayoa, B.; Medina, N.; Chiller, T.; Lau-Bonilla, D.; Cleveland, A.A.; Gómez, B.L.; Arathoon, E.; Roy, M.; Scheel, C.M. High Mortality and Coinfection in a Prospective Cohort of Human Immunodeficiency Virus/Acquired Immune Deficiency Syndrome Patients with Histoplasmosis in Guatemala. Am. J. Trop. Med. Hyg. 2017, 97, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Damasceno, L.S.; Teixeira, M.D.M.; Barker, B.M.; Almeida, M.A.; Muniz, M.D.M.; Pizzini, C.V.; Mesquita, J.R.L.; Rodríguez-Arellanes, G.; Ramírez, J.A.; Vite-Garín, T.; et al. Novel clinical and dual infection by Histoplasma capsulatum genotypes in HIV patients from Northeastern, Brazil. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antinori, S. Histoplasma capsulatum: More Widespread than Previously Thought. Am. J. Trop. Med. Hyg. 2014, 90, 982–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suã¡rez-Alvarez, R.O.; Sahaza, J.H.; Berzunza-Cruz, M.; Becker, I.; Curiel-Quesada, E.; Pérez-Torres, A.; Reyes-Montes, M.D.R.; Taylor, M.L. Dimorphism and Dissemination of Histoplasma capsulatum in the Upper Respiratory Tract after Intranasal Infection of Bats and Mice with Mycelial Propagules. Am. J. Trop. Med. Hyg. 2019, 101, 716–723. [Google Scholar] [CrossRef] [Green Version]
- Medoff, G.; Maresca, B.; Lambowitz, A.M.; Kobayashi, G.; Painter, A.; Sacco, M.; Carratù, L. Correlation between pathogenicity and temperature sensitivity in different strains of Histoplasma capsulatum. J. Clin. Investig. 1986, 78, 1638–1647. [Google Scholar] [CrossRef]
- Nemecek, J.C. Global Control of Dimorphism and Virulence in Fungi. Science 2006, 312, 583–588. [Google Scholar] [CrossRef]
- Nguyen, V.Q.; Sil, A. Temperature-induced switch to the pathogenic yeast form of Histoplasma capsulatum requires Ryp1, a conserved transcriptional regulator. Proc. Natl. Acad. Sci. USA 2008, 105, 4880–4885. [Google Scholar] [CrossRef] [Green Version]
- Mittal, J.; Ponce, M.G.; Gendlina, I.; Nosanchuk, J.D. Histoplasma Capsulatum: Mechanisms for Pathogenesis. Fungal Physiol. Immunopathogenesis 2018, 422, 157–191. [Google Scholar] [CrossRef]
- Ray, S.C.; Rappleye, C.A. Flying under the radar: Histoplasma capsulatum avoidance of innate immune recognition. Semin. Cell Dev. Biol. 2019, 89, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Isotalo, P.; Chan, K.L.; Rubens, F.; Beanlands, D.S.; Auclair, F.; Veinot, J. Prosthetic valve fungal endocarditis due to histoplasmosis. Can. J. Cardiol. 2001, 17, 297–303. [Google Scholar] [PubMed]
- Carreto-Binaghi, L.E.; Damasceno, L.S.; Pitangui, N.S.; Almeida, A.M.F.; Mendes-Giannini, M.J.S.; Zancopé-Oliveira, R.M.; Taylor, M.L. CouldHistoplasma capsulatumBe Related to Healthcare-Associated Infections? BioMed. Res. Int. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa-Orlandi, C.B.; Sardi, J.C.O.; Pitangui, N.S.; De Oliveira, H.C.; Scorzoni, L.; Galeane, M.C.; Medina-Alarcón, K.P.; Melo, W.C.M.A.; Marcelino, M.Y.; Braz, J.D.; et al. Fungal Biofilms and Polymicrobial Diseases. J. Fungi 2017, 3, 22. [Google Scholar] [CrossRef] [PubMed]
- Pitangui, N.; Sardi, J.; Silva, J.; Benaducci, T.; Da Silva, R.M.; Rodríguez-Arellanes, G.; Taylor, M.; Mendes-Giannini, M.; Fusco-Almeida, A.M. Adhesion ofHistoplasma capsulatumto pneumocytes and biofilm formation on an abiotic surface. Biofouling 2012, 28, 711–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sardi, J.D.C.O.; Pitangui, N.D.S.; Rodríguez-Arellanes, G.; Taylor, M.L.; Fusco-Almeida, A.M.; Giannini, M.J.S.M. Highlights in pathogenic fungal biofilms. Rev. Iberoam. Micol. 2014, 31, 22–29. [Google Scholar] [CrossRef]
- Cordeiro, R.D.A.; Marques, F.J.D.F.; Da Silva, M.R.; Malaquias, A.D.M.; De Melo, C.V.S.; Mafezoli, J.; Oliveira, M.D.C.F.; Brilhante, R.S.N.; Rocha, M.F.G.; Bandeira, T.D.J.P.G.; et al. Synthesis and Antifungal Activity In Vitro of Isoniazid Derivatives against Histoplasma capsulatum var. capsulatum. Antimicrob. Agents Chemother. 2014, 58, 2504–2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brilhante, R.S.N.; De Lima, R.A.C.; Marques, F.J.D.F.; Silva, N.F.; Caetano, É.P.; Castelo-Branco, D.D.S.C.M.; Bandeira, T.D.J.P.G.; Moreira, J.L.B.; Cordeiro, R.D.A.; Monteiro, A.J.; et al. Histoplasma capsulatum in planktonic and biofilm forms: In vitro susceptibility to amphotericin B, itraconazole and farnesol. J. Med. Microbiol. 2015, 64, 394–399. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, M.; Di Leva, G.; Croce, C. microRNAs as Anti-Cancer Therapy. Curr. Pharm. Des. 2014, 20, 5328–5335. [Google Scholar] [CrossRef]
- Leung, A.; Natarajan, R. Noncoding RNAs in vascular disease. Curr. Opin. Cardiol. 2014, 29, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.M.; Xu, Z.; Shek, F.H.; Wong, K.-F.; Lee, N.P.; Poon, R.T.; Chen, J.; Luk, J.M. miR-122 Targets Pyruvate Kinase M2 and Affects Metabolism of Hepatocellular Carcinoma. PLoS ONE 2014, 9, e86872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rebane, A.; Akdis, C.A. MicroRNAs in Allergy and Asthma. Curr. Allergy Asthma Rep. 2014, 14, 424. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Cao, X.; Li, Y. Roles of microRNAs in allergic airway diseases. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2014, 36, 114–118. [Google Scholar] [PubMed]
- O’connell, R.M.; Rao, D.S.; Chaudhuri, A.A.; Baltimore, D. Physiological and pathological roles for microRNAs in the immune system. Nat. Rev. Immunol. 2010, 10, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Chitwood, D.H.; Timmermans, M.C.P. Target mimics modulate miRNAs. Nat. Genet. 2007, 39, 935–936. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.C.; Leitão, A.L.; Enguita, F.J. Biogenesis and Mechanism of Action of Small Non-Coding RNAs: Insights from the Point of View of Structural Biology. Int. J. Mol. Sci. 2012, 13, 10268–10295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monk, C.E.; Hutvagner, G.; Arthur, J.S.C. Regulation of miRNA Transcription in Macrophages in Response to Candida albicans. PLoS ONE 2010, 5, e13669. [Google Scholar] [CrossRef] [Green Version]
- Das Gupta, M.; Fliesser, M.; Springer, J.; Breitschopf, T.; Schlossnagel, H.; Schmitt, A.-L.; Kurzai, O.; Hünniger, K.; Einsele, H.; Loeffler, J. Aspergillus fumigatus induces microRNA-132 in human monocytes and dendritic cells. Int. J. Med. Microbiol. 2014, 304, 592–596. [Google Scholar] [CrossRef]
- Muhammad, S.A.; Fatima, N.; Syed, N.-I.-H.; Wu, X.; Yang, X.F.; Chen, J.Y. MicroRNA Expression Profiling of Human Respiratory Epithelium Affected by Invasive Candida Infection. PLoS ONE 2015, 10, e0136454. [Google Scholar] [CrossRef] [Green Version]
- Agustinho, D.P.; De Oliveira, M.A.; Tavares, A.H.; Derengowski, L.; Stolz, V.; Guilhelmelli, F.; Mortari, M.R.; Kuchler, K.; Silva-Pereira, I. Dectin-1 is required for miR155 upregulation in murine macrophages in response toCandida albicans. Virulence 2017, 8, 41–52. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Jin, Y.; Chen, H.; Liao, N.; Wang, Y.; Chen, J. MicroRNA-mediated inflammatory responses induced by Cryptococcus neoformans are dependent on the NF-κB pathway in human monocytes. Int. J. Mol. Med. 2017, 39, 1525–1532. [Google Scholar] [CrossRef] [PubMed]
- Singulani, J.D.L.; Silva, J.D.F.D.; Gullo, F.P.; Costa, M.C.; Fusco-Almeida, A.M.; Enguita, F.J.; Giannini, M.J.S.M. Preliminary evaluation of circulating microRNAs as potential biomarkers in paracoccidioidomycosis. Biomed. Rep. 2017, 6, 353–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitangui, N.D.S.; Sardi, J.D.C.O.; Evoltan, A.R.; Santos, C.T.E.; Silva, J.D.F.D.; Da Silva, R.A.M.; Souza, F.O.; Soares, C.P.; Erodríguez-Arellanes, G.; Taylor, M.L.; et al. An Intracellular Arrangement of Histoplasma capsulatum Yeast-Aggregates Generates Nuclear Damage to the Cultured Murine Alveolar Macrophages. Front. Microbiol. 2016, 6, 1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasuga, T.; White, T.J.; Koenig, G.; McEwen, J.; Restrepo, A.; Castañeda, E.; Lacaz, C.D.S.; Heins-Vaccari, E.M.; De Freitas, R.S.; Zancopé-Oliveira, R.M.; et al. Phylogeography of the fungal pathogen Histoplasma capsulatum. Mol. Ecol. 2003, 12, 3383–3401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, M.D.M.; Patané, J.S.L.; Taylor, M.L.; Gómez, B.L.; Theodoro, R.C.; De Hoog, S.; Engelthaler, D.M.; Zancopé-Oliveira, R.M.; Felipe, M.S.S.; Barker, B.M. Worldwide Phylogenetic Distributions and Population Dynamics of the Genus Histoplasma. PLoS Negl. Trop. Dis. 2016, 10, e0004732. [Google Scholar] [CrossRef] [Green Version]
- Gannon, W.L.; Sikes, R.S. Guidelines of the American Society of Mammalogists for the Use of Wild Mammals in Research. J. Mammal. 2007, 88, 809–823. [Google Scholar] [CrossRef]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, W.J.; Gonçalves, L.M.; Seneviratne, J.; Parahitiyawa, N.; Samaranayake, L.P.; Cury, A.A.D.B. Exopolysaccharide matrix of developed Candida albicans biofilms after exposure to antifungal agents. Br. Dent. J. 2012, 23, 716–722. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.Y.; Gilles, K.; Hamilton, J.K.; Rebers, P.A.; Smith, F.G. A Colorimetric Method for the Determination of Sugars. Nat. Cell Biol. 1951, 168, 167. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Vlachos, I.S.; Kostoulas, N.; Vergoulis, T.; Georgakilas, G.; Reczko, M.; Maragkakis, M.; Paraskevopoulou, M.D.; Prionidis, K.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA miRPath v.2.0: Investigating the combinatorial effect of microRNAs in pathways. Nucleic Acids Res. 2012, 40, W498–W504. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.-P.; Lee, C.-Y.; Tsai, M.-H.; Chiu, Y.-C.; Hsiao, C.K.; Lai, L.-C.; Chuang, E.Y. miRSystem: An Integrated System for Characterizing Enriched Functions and Pathways of MicroRNA Targets. PLoS ONE 2012, 7, e42390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dweep, H.; Gretz, N. miRWalk2.0: A comprehensive atlas of microRNA-target interactions. Nat. Methods 2015, 12, 697. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulos, D.; Rotem, A.; Struhl, K. Inhibition of miR-193a Expression by Max and RXR Activates K-Ras and PLAU to Mediate Distinct Aspects of Cellular Transformation. Cancer Res. 2011, 71, 5144–5153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.-W.; Wu, Y.; Wang, D.; Qin, Z. MicroRNA network analysis identifies key microRNAs and genes associated with precancerous lesions of gastric cancer. Genet. Mol. Res. 2014, 13, 8695–8703. [Google Scholar] [CrossRef] [PubMed]
- Köpke, S.; Buhrke, T.; Lampen, A. miRNA Expression in Human Intestinal Caco-2 Cells is Comparably Regulated by cis- and trans -Fatty Acids. Lipids 2015, 50, 227–239. [Google Scholar] [CrossRef]
- Cook, J.R.; MacIntyre, D.A.; Samara, E.; Kim, S.H.; Singh, N.; Johnson, M.R.; Bennett, P.R.; Terzidou, V. Exogenous oxytocin modulates human myometrial microRNAs. Am. J. Obstet. Gynecol. 2015, 213, 65.e1–65.e9. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Shu, R.; Jiang, S.; Liu, D.L.; Zhang, X. Comparison of microRNA profiles of human periodontal diseased and healthy gingival tissues. Int. J. Oral Sci. 2011, 3, 125–134. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Dong, Y.; Wu, C.W.; Zhao, Z.; Ng, S.S.M.; Chan, F.K.L.; Sung, J.J.Y.; Yu, J. MicroRNA-218 Inhibits Cell Cycle Progression and Promotes Apoptosis in Colon Cancer by Downregulating BMI1 Polycomb Ring Finger Oncogene. Mol. Med. 2012, 18, 1491–1498. [Google Scholar] [CrossRef]
- Heikham, R.; Shankar, R. Flanking region sequence information to refine microRNA target predictions. J. Biosci. 2010, 35, 105–118. [Google Scholar] [CrossRef]
- Fourie, N.H.; Peace, R.M.; Abey, S.K.; Sherwin, L.B.; Rahim-Williams, B.; Smyser, P.A.; Wiley, J.W.; Henderson, W.A. Elevated circulating miR-150 and miR-342-3p in patients with irritable bowel syndrome. Exp. Mol. Pathol. 2014, 96, 422–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brogaard, L.; Larsen, L.E.; Heegaard, P.M.H.; Anthon, C.; Gorodkin, J.; Dürrwald, R.; Skovgaard, K. IFN-λ and microRNAs are important modulators of the pulmonary innate immune response against influenza A (H1N2) infection in pigs. PLoS ONE 2018, 13, e0194765. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Jiang, S.; Yoshida, N.; Sakamoto, R.; Nishizawa, M. Interferon-alpha competing endogenous RNA network antagonizes microRNA-1270. Cell. Mol. Life Sci. 2015, 72, 2749–2761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.; Zhang, Z.-Y.; Fu, S.-L.; Xie, J.-G.; Liu, X.-S.; Xu, Y.-J.; Zhao, J.-P.; Xiong, W. Retracted Article: Hsa-miR-623 suppresses tumor progression in human lung adenocarcinoma. Cell Death Dis. 2016, 7, e2388. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; Lee, S.Y.; Lee, S.Y.; Kim, Y.J.; Park, J.Y.; Kwon, S.J.; Na, M.J.; Lee, E.J.; Jeon, H.S.; Son, J.W. microRNA-99b acts as a tumor suppressor in non-small cell lung cancer by directly targeting fibroblast growth factor receptor 3. Exp. Ther. Med. 2011, 3, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.G.; Shi, Y.; Hong, D.F.; Song, M.; Huang, D.; Wang, C.Y.; Zhao, G. MiR-148b suppresses cell proliferation and invasion in hepatocellular carcinoma by targeting WNT1/β-catenin pathway. Sci Rep. 2015, 5, 8087. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhang, M.; Huang, Y.; Feng, L.; Chen, H.; Huarong, C.; Chen, H.; Zhang, K.; Zheng, L.; Zheng, S. MicroRNA-320b promotes colorectal cancer proliferation and invasion by competing with its homologous microRNA-320a. Cancer Lett. 2015, 356, 669–675. [Google Scholar] [CrossRef] [Green Version]
- Rai, K.; Takigawa, N.; Ito, S.; Kashihara, H.; Ichihara, E.; Yasuda, T.; Shimizu, K.; Tanimoto, M.; Kiura, K. Liposomal Delivery of MicroRNA-7–Expressing Plasmid Overcomes Epidermal Growth Factor Receptor Tyrosine Kinase Inhibitor-Resistance in Lung Cancer Cells. Mol. Cancer Ther. 2011, 10, 1720–1727. [Google Scholar] [CrossRef] [Green Version]
- Weng, L.; Ziliak, D.; Lacroix, B.; Geeleher, P.; Huang, R.S. Integrative “Omic” Analysis for Tamoxifen Sensitivity through Cell Based Models. PLoS ONE 2014, 9, e93420. [Google Scholar] [CrossRef]
- Adyshev, D.M.; Elangovan, V.R.; Moldobaeva, N.; Mapes, B.; Sun, X.; Garcia, J.G. Mechanical stress induces pre-B-cell colony-enhancing factor/NAMPT expression via epigenetic regulation by miR-374a and miR-568 in human lung endothelium. Am. J. Respir. Cell Mol. Biol. 2014, 50, 409–418. [Google Scholar]
- Looney, A.-M.; Walsh, B.H.; Moloney, G.M.; Egrenham, S.; Fagan, A.; O’Keeffe, G.W.; Clarke, G.; Cryan, J.F.; Dinan, T.G.; Boylan, G.B.; et al. Downregulation of Umbilical Cord Blood Levels of miR-374a in Neonatal Hypoxic Ischemic Encephalopathy. J. Pediatr. 2015, 167, 269–273.e2. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Green, B.; Marrone, A.; Guo, Y.; Kadlubar, S.; Lin, D.; Fuscoe, J.; Pogribny, I.; Ning, B. Suppression of CYP2C9 by MicroRNA hsa-miR-128-3p in Human Liver Cells and Association with Hepatocellular Carcinoma. Sci. Rep. 2015, 5, srep08534. [Google Scholar] [CrossRef] [PubMed]
- Ye, E.-A.; Steinle, J.J. miR-15b/16 protects primary human retinal microvascular endothelial cells against hyperglycemia-induced increases in tumor necrosis factor alpha and suppressor of cytokine signaling 3. J. Neuroinflammation 2015, 12, 44–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haenisch, S.; Laechelt, S.; Bruckmueller, H.; Werk, A.; Noack, A.; Bruhn, O.; Remmler, C.; Cascorbi, I. Down-Regulation of ATP-Binding Cassette C2 Protein Expression in HepG2 Cells after Rifampicin Treatment Is Mediated by MicroRNA-379. Mol. Pharmacol. 2011, 80, 314–320. [Google Scholar] [CrossRef]
- Cazzoli, R.; Buttitta, F.; Di Nicola, M.; Malatesta, S.; Marchetti, A.; Rom, W.N.; Pass, H.I. microRNAs Derived from Circulating Exosomes as Noninvasive Biomarkers for Screening and Diagnosing Lung Cancer. J. Thorac. Oncol. 2013, 8, 1156–1162. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.; Ma, L.; Chen, X.-L.; Huang, X.; Wang, Q. Downregulation of LncRNAH19 and MiR-675 promotes migration and invasion of human hepatocellular carcinoma cells through AKT/GSK-3β/Cdc25A signaling pathway. Acta Acad. Med. Wuhan 2014, 34, 363–369. [Google Scholar] [CrossRef]
- Chen, S.; Li, F.; Chai, H.; Tao, X.; Wang, H.; Ji, A. miR-502 inhibits cell proliferation and tumor growth in hepatocellular carcinoma through suppressing phosphoinositide 3-kinase catalytic subunit gamma. Biochem. Biophys. Res. Commun. 2015, 464, 500–505. [Google Scholar] [CrossRef]
- Mraz, M.; Dolezalova, D.; Plevova, K.; Kozubik, K.S.; Mayerova, V.; Cerna, K.; Musilova, K.; Tichy, B.; Pavlova, S.; Borsky, M.; et al. MicroRNA-650 expression is influenced by immunoglobulin gene rearrangement and affects the biology of chronic lymphocytic leukemia. Blood 2012, 119, 2110–2113. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Shi, L.-Y.; Lei, Y.-M.; Bao, Y.-H.; Li, Z.-Y.; Ding, F.; Zhu, G.-T.; Wang, Q.-Q.; Huang, C.-X. Radiosensitization effect of hsa-miR-138-2-3p on human laryngeal cancer stem cells. PeerJ 2017, 5, e3233. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhao, L.; Shen, Q.; Lv, Q.; Jin, M.; Ma, H. Down-regulation of KIAA1199/CEMIP by miR-216a suppresses tumor invasion and metastasis in colorectal cancer. Int. J. Cancer 2017, 140, 2298–2309. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Liu, Y.; Zhu, H.; Chen, D.; Hu, W.; Nie, X.; Zheng, X.; Huang, S.; Zhou, P. Downregulation of miR-216a-5p by long noncoding RNA PVT1 suppresses colorectal cancer progression via modulation of YBX1 expression. Cancer Manag. Res. 2019, 11, 6981–6993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Croston, T.L.; Nayak, A.P.; Lemons, A.R.; Goldsmith, W.T.; Gu, J.K.; Germolec, D.R.; Beezhold, D.H.; Green, B.J. Influence ofAspergillus fumigatusconidia viability on murine pulmonary microRNA and mRNA expression following subchronic inhalation exposure. Clin. Exp. Allergy 2016, 46, 1315–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seneviratne, C.J.; Jin, L.; Samaranayake, L.P. Biofilm lifestyle of Candida: A mini review. Oral Dis. 2008, 14, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Singh, Y.; Kaul, V.; Mehra, A.; Chatterjee, S.; Tousif, S.; Dwivedi, V.P.; Suar, M.; Van Kaer, L.; Bishai, W.R.; Das, G. Mycobacterium tuberculosisControls MicroRNA-99b (miR-99b) Expression in Infected Murine Dendritic Cells to Modulate Host Immunity. J. Biol. Chem. 2013, 288, 5056–5061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, K.; Tong, D.; Zhang, S.; Cai, D.; Wang, L.; Yang, Y.; Gao, L.; Chang, S.; Guo, B.; Song, T.; et al. miRNA-99b-3p functions as a potential tumor suppressor by targeting glycogen synthase kinase-3β in oral squamous cell carcinoma Tca-8113 cells. Int. J. Oncol. 2015, 47, 1528–1536. [Google Scholar] [CrossRef] [PubMed]
- Lina, T.T.; Luo, T.; Velayutham, T.-S.; Das, S.; McBride, J.W. Ehrlichia Activation of Wnt-PI3K-mTOR Signaling Inhibits Autolysosome Generation and Autophagic Destruction by the Mononuclear Phagocyte. Infect. Immun. 2017, 85, e00690-17. [Google Scholar] [CrossRef] [Green Version]
- Almazov, V.P.; Kochetkov, D.V.; Chumakov, P.M. The use of p53 as a tool for human cancer therapy. Mol. Biol. 2007, 41, 947–963. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Du, Q.; Han, C.; Wang, Z.; Zhang, X.; Wang, T.; Zhao, X.; Huang, Y.; Tong, D. p53 signaling modulation of cell cycle arrest and viral replication in porcine circovirus type 2 infection cells. Veter Res. 2016, 47, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, T.; Thuer, E.; Heijink, M.; Tóth, R.; Bodai, L.; Vágvölgyi, C.; Giera, M.; Gabaldón, T.; Gácser, A. Eicosanoid biosynthesis influences the virulence of Candida parapsilosis. Virulence 2018, 9, 1019–1035. [Google Scholar] [CrossRef] [Green Version]
- Schulz, W.A.; Ingenwerth, M.; Djuidje, C.; Hader, C.; Rahnenfuehrer, J.; Engers, R. Changes in cortical cytoskeletal and extracellular matrix gene expression in prostate cancer are related to oncogenic ERG deregulation. BMC Cancer 2010, 10, 505. [Google Scholar] [CrossRef] [Green Version]
- Bish, R.; Cuevas-Polo, N.; Cheng, Z.; Hambardzumyan, D.; Munschauer, M.; Landthaler, M.; Vogel, C. Comprehensive Protein Interactome Analysis of a Key RNA Helicase: Detection of Novel Stress Granule Proteins. Biomolecules 2015, 5, 1441–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buetepage, M.; Eckei, L.; Verheugd, P.; Lüscher, B. Intracellular Mono-ADP-Ribosylation in Signaling and Disease. Cells 2015, 4, 569–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rooij, E.; Marshall, W.S.; Olson, E.N. Toward MicroRNA–Based Therapeutics for Heart Disease. Circ. Res. 2008, 103, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Rappleye, C.A.; Engle, J.T.; Goldman, W.E. RNA interference in Histoplasma capsulatum demonstrates a role for α-(1,3)-glucan in virulence. Mol. Microbiol. 2004, 53, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Bohse, M.L.; Woods, J.P. RNA Interference-Mediated Silencing of the YPS3 Gene of Histoplasma capsulatum Reveals Virulence Defects. Infect. Immun. 2007, 75, 2811–2817. [Google Scholar] [CrossRef] [Green Version]
- Dubois, J.C.; Smulian, A.G. Sterol Regulatory Element Binding Protein (Srb1) Is Required for Hypoxic Adaptation and Virulence in the Dimorphic Fungus Histoplasma capsulatum. PLoS ONE 2016, 11, e0163849. [Google Scholar] [CrossRef] [Green Version]
- Dade, J.; Dubois, J.C.; Pasula, R.; Donnell, A.M.; Caruso, J.A.; Smulian, A.G.; Deepe, G.S. HcZrt2, a zinc responsive gene, is indispensable for the survival of Histoplasma capsulatum in vivo. Med. Mycol. 2016, 54, 865–875. [Google Scholar] [CrossRef] [Green Version]
- Pakshir, K.; Badali, H.; Nami, S.; Mirzaei, H.; Ebrahimzadeh, V.; Morovati, H. Interactions between immune response to fungal infection and microRNAs: The pioneer tuners. Mycoses 2019, 63, 4–20. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
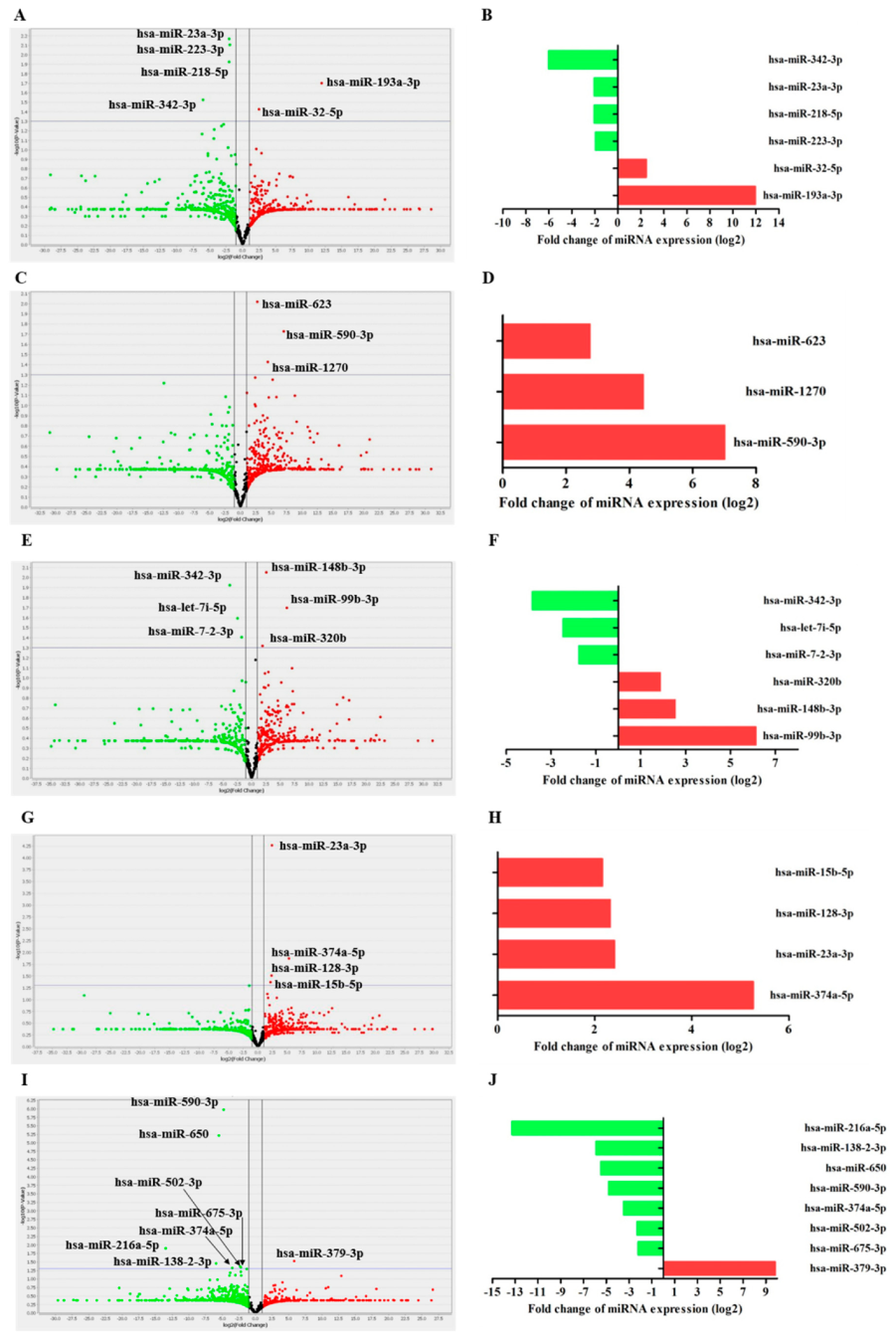
| MiRNA ID | Fold Change (RQ) | p Value | Process or Function Categories Involved | References |
|---|---|---|---|---|
| THP-1 Mø-like cells infected with the EH-315 strain vs. uninfected cells | ||||
| hsa-miR-193a-3p | 3961.50 | 0.0199 | Inhibits cellular transformation | [44,45] |
| hsa-miR-32-5p | 5.50 | 0.0373 | Regulation of apoptosis | [46] |
| hsa-miR-223-3p | 0.25 | 0.0078 | Monocyte-macrophage differentiation; cell proliferation; Induction of apoptosis | [47,48] |
| hsa-miR-218-5p | 0.24 | 0.0119 | Inhibits cell cycle proliferation; induction of apoptosis | [49] |
| hsa-miR-23a-3p | 0.24 | 0.0068 | T-cell signaling; regulation of apoptosis | [50] |
| hsa-miR-342-3p | 0.01 | 0.0297 | Inflammation; pain signaling | [51] |
| THP-1 Mø-like cells infected with the 60I strain vs. uninfected cells | ||||
| hsa-miR-590-3p | 128.33 | 0.01 | Regulation of apoptosis | [52] |
| hsa-miR-1270 | 21.65 | 0.03 | Regulation of IFNα1 | [53] |
| hsa-miR-623 | 6.69 | 0.009 | Cell proliferation, migration, and invasion of tumorous tissues | [54] |
| THP-1 Mø-like cells infected with EH-315 strain biofilms vs. uninfected cells | ||||
| hsa-miR-99b-3p | 69.97 | 0.02 | Cell proliferation | [55] |
| hsa-miR-148b-3p | 5.78 | 0.008 | Induce cell apoptosis and inhibiting cell invasion | [56] |
| hsa-miR-320b | 3.65 | 0.04 | Proliferation and cell invasion | [57] |
| hsa-miR-7-2-3p | 0.29 | 0.03 | Overexpression inhibits cell growth in lung cells line | [58] |
| hsa-let-7i-5p | 0.18 | 0.02 | Induces sensitivity to antineoplastic agents | [59] |
| hsa-miR-342-3p | 0.07 | 0.01 | Inflammation; pain signaling | [51] |
| THP-1 Mø-like cells infected with EH-315 strain biofilms vs. cells infected with the EH-315 strain | ||||
| hsa-miR-374a-5p | 38.54 | 0.01 | Hypoxia; Reduction of vascular permeability of lung tissue | [60,61] |
| hsa-miR-23a-3p | 5.32 | 0.0001 | T-cell signaling; regulation of apoptosis | [50] |
| hsa-miR-128-3p | 4.99 | 0.03 | Proliferation and cell invasion; apoptosis | [62] |
| hsa-miR-15b-5p | 4.46 | 0.04 | Downregulation of TNFα levels; apoptosis | [63] |
| THP-1 Mø-like cells infected with the EH-315 strain vs. cells infected with the 60I strain | ||||
| hsa-miR-379-3p | 903.56 | 0.02 | Regulation of cell adhesion; therapeutic response; drug resistance profiles | [64,65] |
| hsa-miR-675-3p | 0.21 | 0.04 | Migration and cell invasion | [66] |
| hsa-miR-502-3p | 0.20 | 0.04 | Inhibition of autophagy, cell growth and cell cycle progression | [67] |
| hsa-miR-374a-5p | 0.08 | 0.04 | Hypoxia; reduction of vascular permeability of lung tissue | [60,61] |
| hsa-miR-590-3p | 0.03 | 0.0 | Regulation of apoptosis | [52] |
| hsa-miR-650 | 0.02 | 0.0 | Inhibits cell cycle progression; influences the proliferation capacity of B cells | [68] |
| hsa-miR-138-2-3p | 0.01 | 0.03 | Increases the proportion of early and late apoptosis; raises G1 phase arrest; and down-regulation of the S stage in the cell cycle | [69] |
| hsa-miR-216a-5p | 0.0001 | 0.01 | Decreases migration, invasion, cell viability and induces cell apoptosis | [70,71] |
| KEGG Pathway | Number of Genes | P Value |
|---|---|---|
| THP-1 Mø-like cells infected with the EH-315 strain from planktonic growth form vs. uninfected cells | ||
| ECM-receptor interaction | 14 | 1.1 × 10−6 |
| Glycosaminoglycan biosynthesis | 6 | 1.7 × 10−12 |
| Ubiquitin mediated proteolysis | 26 | 0.01 |
| Focal adhesion | 32 | 0.01 |
| MAPK signaling (mitogen-activated protein kinase) | 35 | 0.02 |
| Apoptosis | 12 | 0.04 |
| Gap junction | 17 | 0.04 |
| Regulation of actin cytoskeleton | 29 | 0.04 |
| THP-1 Mø-like cells infected with the 60I strain from planktonic growth form vs. uninfected cells | ||
| Glycosaminoglycan biosynthesis | 7 | 1.9 × 10−11 |
| ECM-receptor interaction | 21 | 7.9 × 10−11 |
| Lysine degradation | 14 | 2.6 × 10−5 |
| TGF-β signaling | 21 | 0.0004 |
| THP-1 Mø-like cells infected with the EH-315 strain from biofilms growth form vs. uninfected cells | ||
| Glycosaminoglycan biosynthesis | 7 | 2.1 × 10−15 |
| ECM-receptor interaction | 19 | 1.8 × 10−10 |
| Lysine degradation | 14 | 1.2 × 10−6 |
| TGF-β signaling | 26 | 2.1 × 10−5 |
| Focal adhesion | 49 | 0.001 |
| Wnt signaling pathway | 39 | 0.0002 |
| Gap junction | 19 | 0.009 |
| Adherence junction | 17 | 0.01 |
| MAPK signaling | 56 | 0.01 |
| Apoptosis | 18 | 0.01 |
| N-Glycan biosynthesis | 10 | 0.04 |
| p53 signaling pathway | 18 | 0.04 |
| THP-1 Mø-like cells infected with the EH-315 strain in biofilms growth form vs. cells infected with the EH-315 strain in planktonic growth form | ||
| Glycosaminoglycan biosynthesis | 6 | 5.1 × 10−41 |
| TGF-β signaling | 26 | 3.4 × 10−7 |
| p53 signaling pathway | 22 | 0.0001 |
| Wnt signaling pathway | 38 | 0.0008 |
| Focal adhesion | 46 | 0.001 |
| MAPK signaling | 56 | 0.002 |
| Ubiquitin mediated proteolysis | 32 | 0.01 |
| T cell receptor signaling pathway | 25 | 0.02 |
| THP-1 Mø-like cells infected with the EH-315 strain vs. cells infected with the 60I strain, both in planktonic growth form | ||
| ECM-receptor interaction | 26 | 2.6 × 10−9 |
| TGF-β signaling | 35 | 1.2 × 10−8 |
| Glycosaminoglycan biosynthesis | 7 | 1.5 × 10−5 |
| Biosynthesis of unsaturated fatty acids | 7 | 4.05 × 10−5 |
| Lysine degradation | 14 | 0.006 |
| Ubiquitin mediated proteolysis | 41 | 0.01 |
| Valine, leucine, and isoleucine degradation | 12 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pitangui, N.d.S.; de Lacorte Singulani, J.; Sardi, J.d.C.O.; de Souza, P.C.; Rodríguez-Arellanes, G.; García-Pérez, B.E.; Enguita, F.J.; Pavan, F.R.; Taylor, M.L.; Mendes-Giannini, M.J.S.; et al. Differential miRNA Expression in Human Macrophage-Like Cells Infected with Histoplasma capsulatum Yeasts Cultured in Planktonic and Biofilm Forms. J. Fungi 2021, 7, 60. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010060
Pitangui NdS, de Lacorte Singulani J, Sardi JdCO, de Souza PC, Rodríguez-Arellanes G, García-Pérez BE, Enguita FJ, Pavan FR, Taylor ML, Mendes-Giannini MJS, et al. Differential miRNA Expression in Human Macrophage-Like Cells Infected with Histoplasma capsulatum Yeasts Cultured in Planktonic and Biofilm Forms. Journal of Fungi. 2021; 7(1):60. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010060
Chicago/Turabian StylePitangui, Nayla de Souza, Junya de Lacorte Singulani, Janaina de Cássia Orlandi Sardi, Paula Carolina de Souza, Gabriela Rodríguez-Arellanes, Blanca Estela García-Pérez, Francisco Javier Enguita, Fernando R. Pavan, Maria Lucia Taylor, Maria José Soares Mendes-Giannini, and et al. 2021. "Differential miRNA Expression in Human Macrophage-Like Cells Infected with Histoplasma capsulatum Yeasts Cultured in Planktonic and Biofilm Forms" Journal of Fungi 7, no. 1: 60. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010060