1. Introduction
The European black truffle (
Tuber melanosporum Vittad., Pezizales) is an ectomycorrhizal fungus that in nature mostly grows in association with Angiosperm plants (e.g.,
Fagaceae). In cultivated orchards, the most common hosts are
Quercus species. Truffle cultivation has advanced greatly in recent years, although it is not completely domesticated yet, and many biological and ecological aspects of the several processes involved still need clarification [
1,
2]. Black truffle has also attracted attention as a model ectomycorrhizal ascomycetous species for genomic studies, research on the mating process and population genetic structure, on fruitbody (FB) nutrition or on aroma [
2,
3,
4,
5]. Black truffle fruiting is a multigene-mediated process that follows specific and organized differentiation patterns and requires several months to reach completion [
3,
6,
7]. The sequential morphogenetic stages leading to the FB formation can be classified into mating process, FB growing stage, and maturation [
1,
6,
7,
8]. However, very little is known about the intrinsic or environmental signaling pathways regulating truffle FB morphogenesis [
1,
7,
9,
10].
The mating process (from the stimulation of the formation of the mating structures to the mating itself) typically happens throughout late spring, apparently in several flushes [
1] (
Table 1). The precise location along the mycelial network where the mating event between mycelia of opposite mating types happens will determine the soil depth of the full-grown FB [
2,
11]. After the mating event, the FB starts to develop and its structure becomes gradually complex as the weight rapidly increases [
7,
8]. Growing below ground, the FB swelling and its final shape will be influenced by the soil mechanical constraints, with a relevant role of the characteristic pyramidal warts of the peridium [
6]. At the end of the intense growth stage, the FB has practically achieved its final size. It is then, that the maturation stage begins, with the spores acquiring their characteristic pigmentation and the FB developing its unique aroma [
7,
12,
13]. Maturation begins in late autumn, and the subsequent senescence processes set the moment in which dogs can localize the ripe FB. The FBs are harvested during several months throughout the winter, evidencing that the volatile compounds that attract dogs are not formed simultaneously in all FBs [
5] (
Table 1). Each one of the spots localized by a dog is excavated by the harvester: in most of the digs only one FB appears (single FBs), whereas in others, a cluster of FBs grow in very close proximity. Little scientific attention is usually paid to truffle FBs growing in clusters [
14,
15], although growing within these clusters could either affect FB formation patterns or could trade off with size due to localized resource depletion or inhibition mechanisms.
The outcome of the FB formation process relies upon how this sequence of stages (mating, growing and maturation) proceeds. The developmental patterns of FBs are affected not only by environmental but also by endogenous factors at different stages, such as the expression of certain enzymes related with melanin-synthesis pathways [
10,
16]. This raises the question of whether the moment and conditions in which a morphogenetic stage occurs might influence the following ones. Research on the relationships among FB development characters could help shed light on this aspect. As outlined above, a number of FB development characters that include fruiting depth, weight, shape, and spore maturity can be primarily linked to particular morphogenetic stages (
Table 1). Since these characters define or influence the commercial quality standards of truffle FBs [
17], understanding the relationships among development characters and the processes that shape them may also open new perspectives for pre-harvest quality management through improved farming practices.
Here, we aimed to: (i) build a causal model to explain how development characters of truffle FB influence one another, and (ii) test whether these relationships are consistent across different soils and dig typologies (single FBs and FB clusters). We tested several alternative models for each dig typology in three replicate blocks along a soil texture gradient that is representative of common truffle orchard soils, and compared these mineral soils with the FBs growing within a peat-based substrate amendment. The causal models were built considering the linkages between the studied characters and the sequential morphogenetic stages of truffle FBs: fruiting depth linked to the mating process, weight and shape linked to the growing stage, and spore maturity linked to maturation [
1,
7,
8] (
Table 1). We hypothesized that: (i) fruiting depth would have a positive effect on weight and maturity, because soil depth buffers extreme values in temperature and water content, which are particularly variable in Mediterranean climates [
18,
19]; (ii) no relationship between weight and maturity would appear, because dogs usually localize full-developed ripe FBs of sizes from less than 10 g to more than 100 g; (iii) shape of small FBs would we more rounded, because they need to make and occupy less soil volume and are less likely to face mechanical constraints during growth; (iv) in FB clusters, the weight of the largest FB would show a negative relationship with the weight of the remaining FBs, due to the local resource depletion or inhibition mechanisms hypothesized by Moore et al. [
20]; and (v) differences among soils and with substrate would affect relationships among FB development characters, since soil properties and localized substrate amendments are able to influence these characters [
21].
4. Discussion
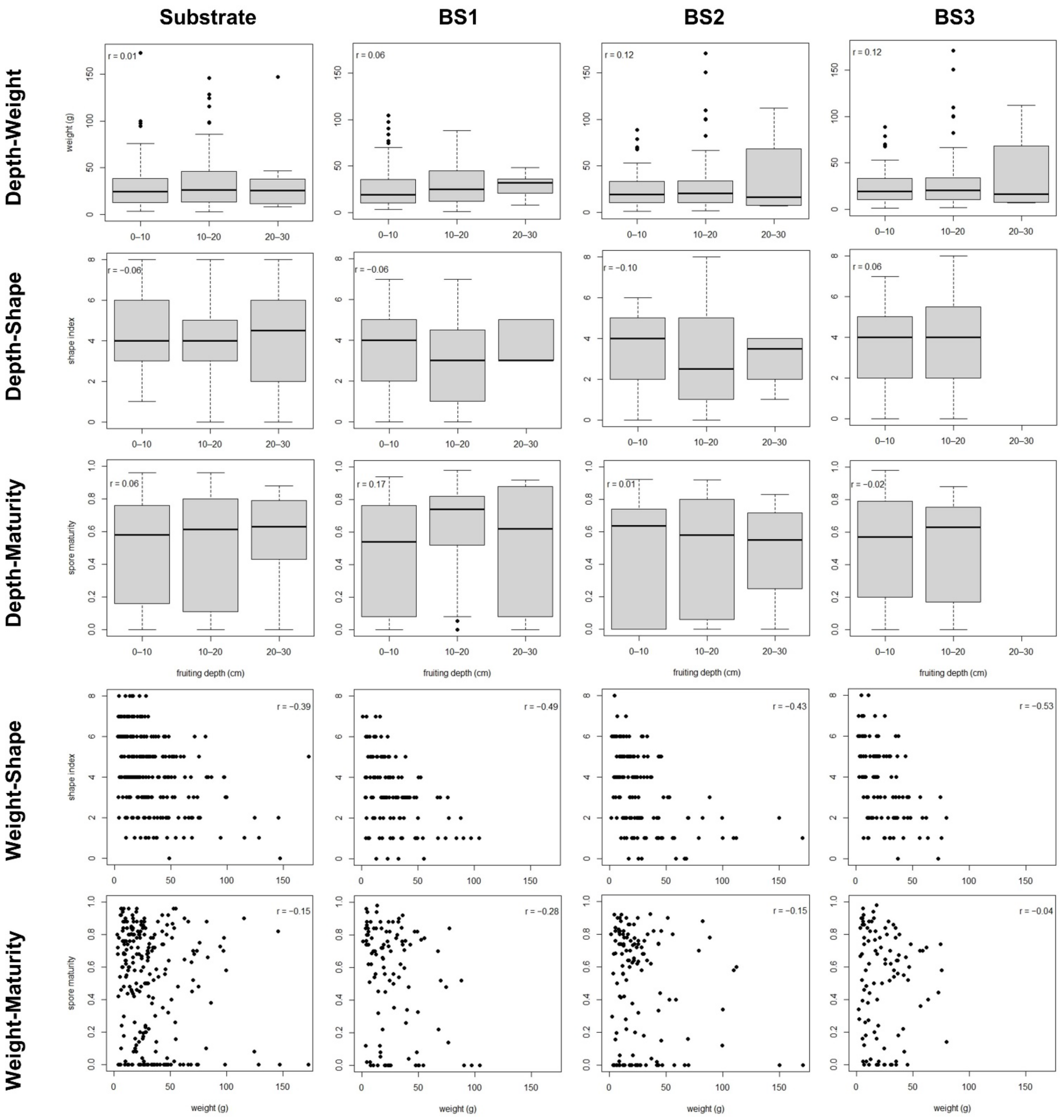
The soil depth at which truffle FBs grew did not show any causal link with FB weight, shape, or spore maturity for any of the soil or dig typologies analyzed. This suggests that, under the experimental conditions, weight and maturity do not depend on FB location throughout the soil profile. This was unexpected, because it is generally accepted that soil depth determines FB sensitivity to bioclimatic damages such as those caused by drought or heat waves [
33]. It is generally accepted that soil temperature and water regime follow marked depth gradients, with more extreme values and more rapid fluctuations near the soil surface [
18,
19]. Montant and Kulifaj [
9] experimentally found that increasing soil temperature during winter and spring made truffle FBs grow earlier—as indicated by the soil surface cracking linked to the swelling of shallow FBs- and ripen earlier—as indicated by the harvesting date [
34]. For other hypogeous ectomycorrhizal fungi, Luoma [
35] and Luoma et al. [
36] found a negative influence of thick layers of organic matter on fruiting, assuming that it was due to poor CO
2 diffusion. However, in our experimental site the existing ranges in microclimatic variables did not trigger changes in weight or maturity throughout the soil profile for any soil typology. Despite this, microclimatic gradients in the soil play a role in the mating process of black truffle and thus on productivity of truffle orchards [
1].
The growth of the truffle FB seems primarily related to the uptake, metabolism, and translocation of carbon from the plant host, and to nitrogen and water balances between fungus and host [
4,
37,
38], although a hypothetical genetic influence on weight could also play a role. Besides, for many fungal species good aeration is associated with successful growth of FBs, in relation to gas exchange with the surrounding soil and to intensification of oxidative metabolism [
20]. Thus, the soil depth at which the truffle grows and its weight seem to be determined by a different set of edaphoclimatic factors, with the host plant playing a more relevant role in the case of weight. The existence of differential gene expression patterns for each development stage is also relevant, and gene expression can also be influenced by environmental factors. Montanini et al. [
39] hypothesized that the abundance of up-regulated transcription factors in
T. melanosporum FB indicates a high degree of functional specialization in the reproductive stage. Moreover, Hacquard et al. [
37] underscored the variety of metabolic pathways reflecting complex genetic processes during the truffle FB formation.
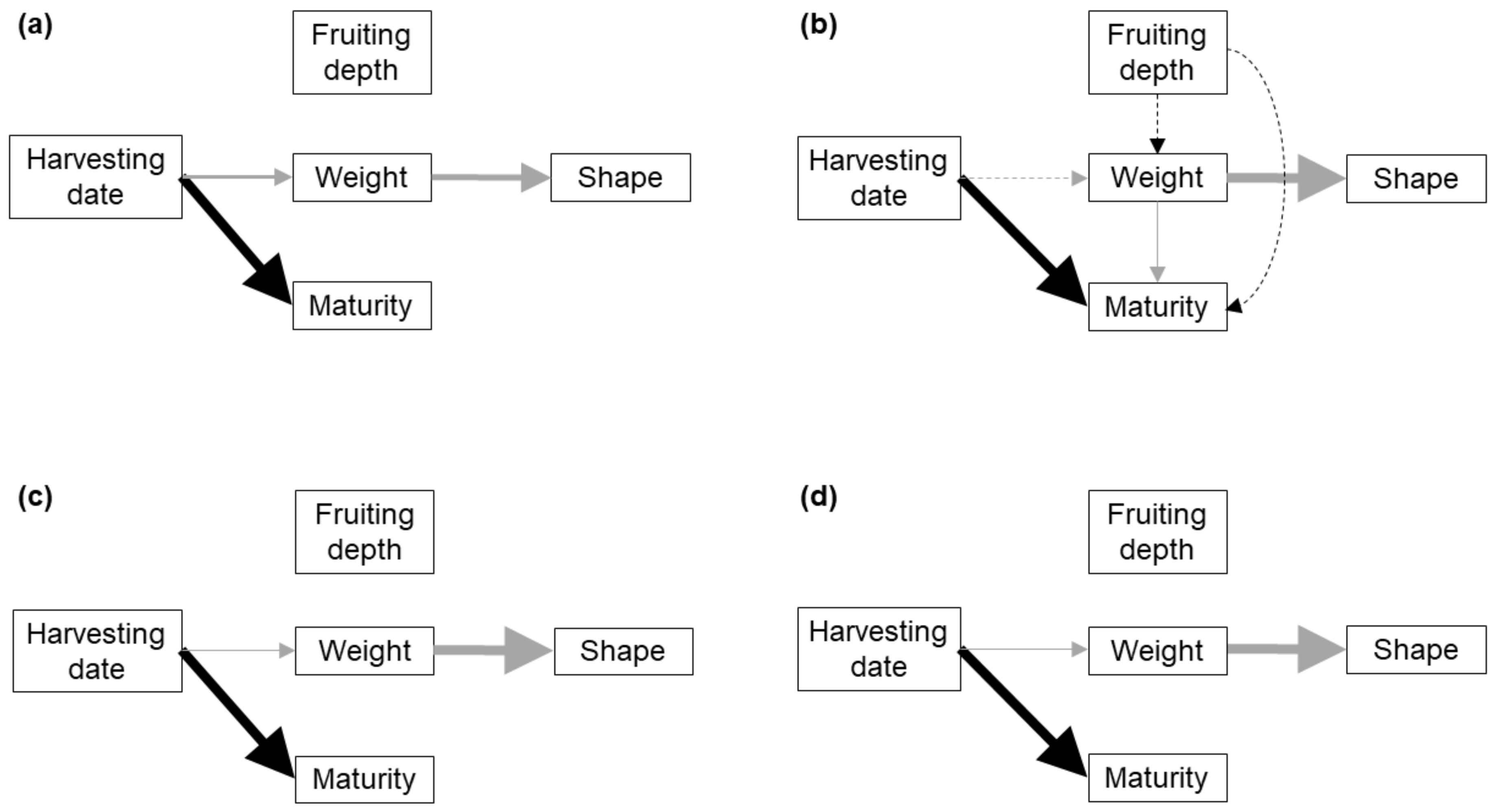
The causal relationship between weight and shape, both linked to the intense growth stage, was clear and general among all the soil and dig typologies, with the exception of BS1 clusters being likely related to a narrower range in weight. The existence of a negative relationship between FB weight and shape index (i.e., bigger FBs showing more irregular, less rounded shapes) is generally accepted between growers and researchers. From early in the development (peridial stage) the peridium warts are formed to help the FB swell [
6,
7], and the mechanical interaction with the surrounding soil matrix (including stones and roots) models the FB shape as it grows. Smaller FBs need to make and occupy less soil volume and are less likely to face mechanical constraints during growth. However, the fact that in all soil typologies the nine shape categories were present marks the great variability of this character (
Tables S6 and S7) and of related mechanical constraints at relatively low spatial scales.
No causal link between FB maturity and weight was found, except for single FBs in BS1. Black truffle growers generally accept that weight and maturity are not related within the usual range of weight for full-developed healthy FBs. We found no evidence that the FBs must reach a minimum size to ripen (or that weight increase and maturation evolve together) for the range of weights studied, with minimum weights of 1–3 g in all soil typologies (
Tables S6 and S7). This size is already attained by some FBs during the veined stage, typically happening in July–August [
7]. Regarding the exception found in BS1, a negative relationship restricted to the smallest FBs was found in this block. This could be related to the fact that spore samples were taken near the peridium. In our experience, during early season it is not uncommon that spore maturity shows a certain degree of heterogeneity within a FB. The negative relationship could be explained by the fact that spore maturity could be more homogeneous in the hymenium of small FBs and our sampling overestimated the differences between small and large FBs, in relation to the fact that the development and maturation of the spores in a FB is asynchronous and progresses from the center of fertile veins outward and from the center of the gleba outward [
6]. This underlines how important it might be for future truffle research to develop unbiased indicators and non-destructive sampling methods for assessing spore maturity [
40].
We found no evidence that the maturation stage of the truffle FB is influenced by its weight, whereas previous research pointed that it is regulated through a decrease in tyrosinase activity [
12,
16]. The transition from the growing to the maturation stage could also be regulated by environmental cues, as suggested by the fact that November-December temperatures influence the mean date of truffle harvesting [
41]. Several genes coding for photoreceptors and light-dependent regulators have been found in
T. melanosporum, suggesting that seasonal variations in light, temperature, and/or oxygen concentration could mediate FB morphogenesis [
3]. However,
T. melanosporum apparently lacks regulators of circadian rhythmicity, which is a common indicator of seasons in plants [
3].
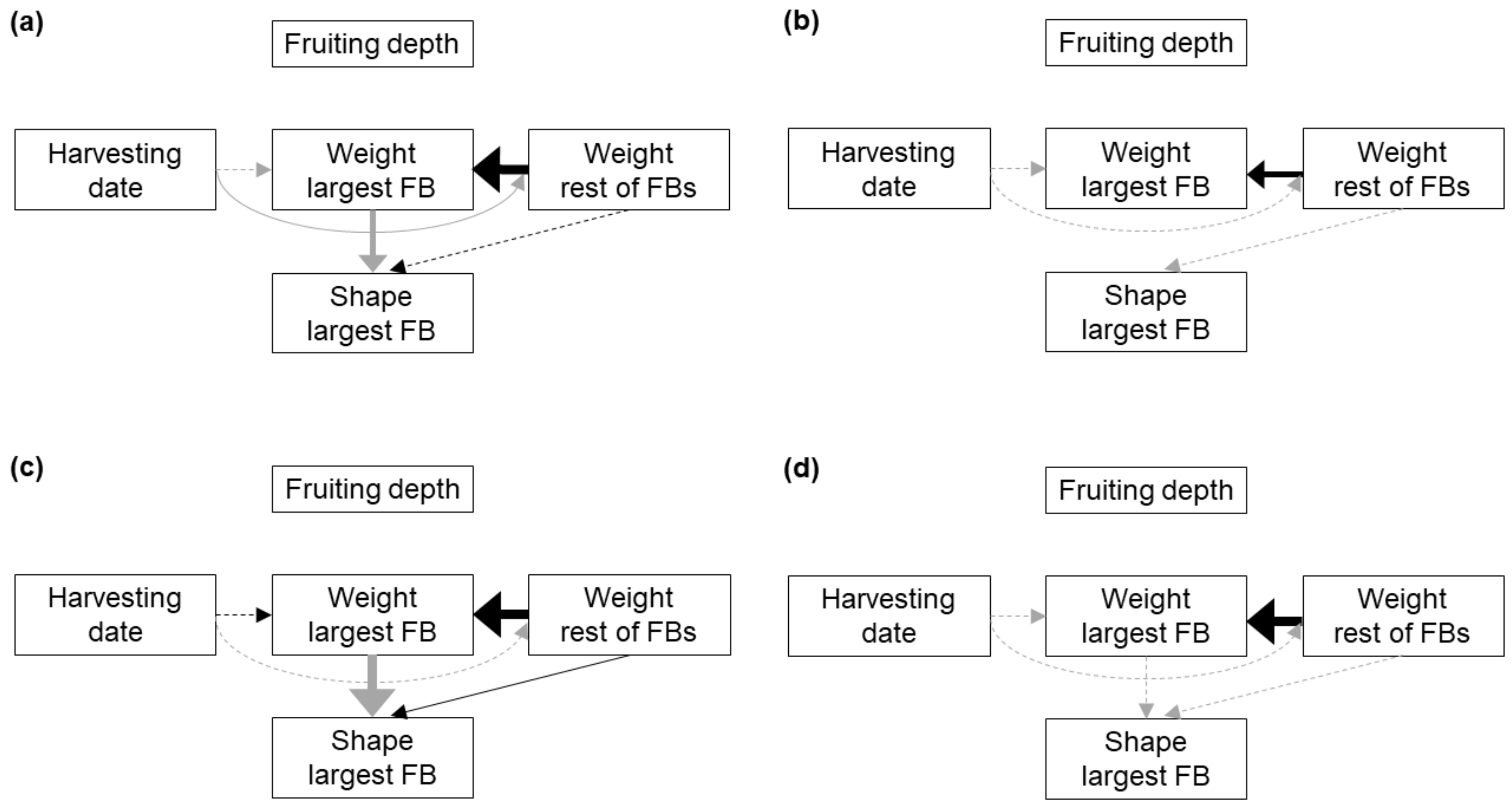
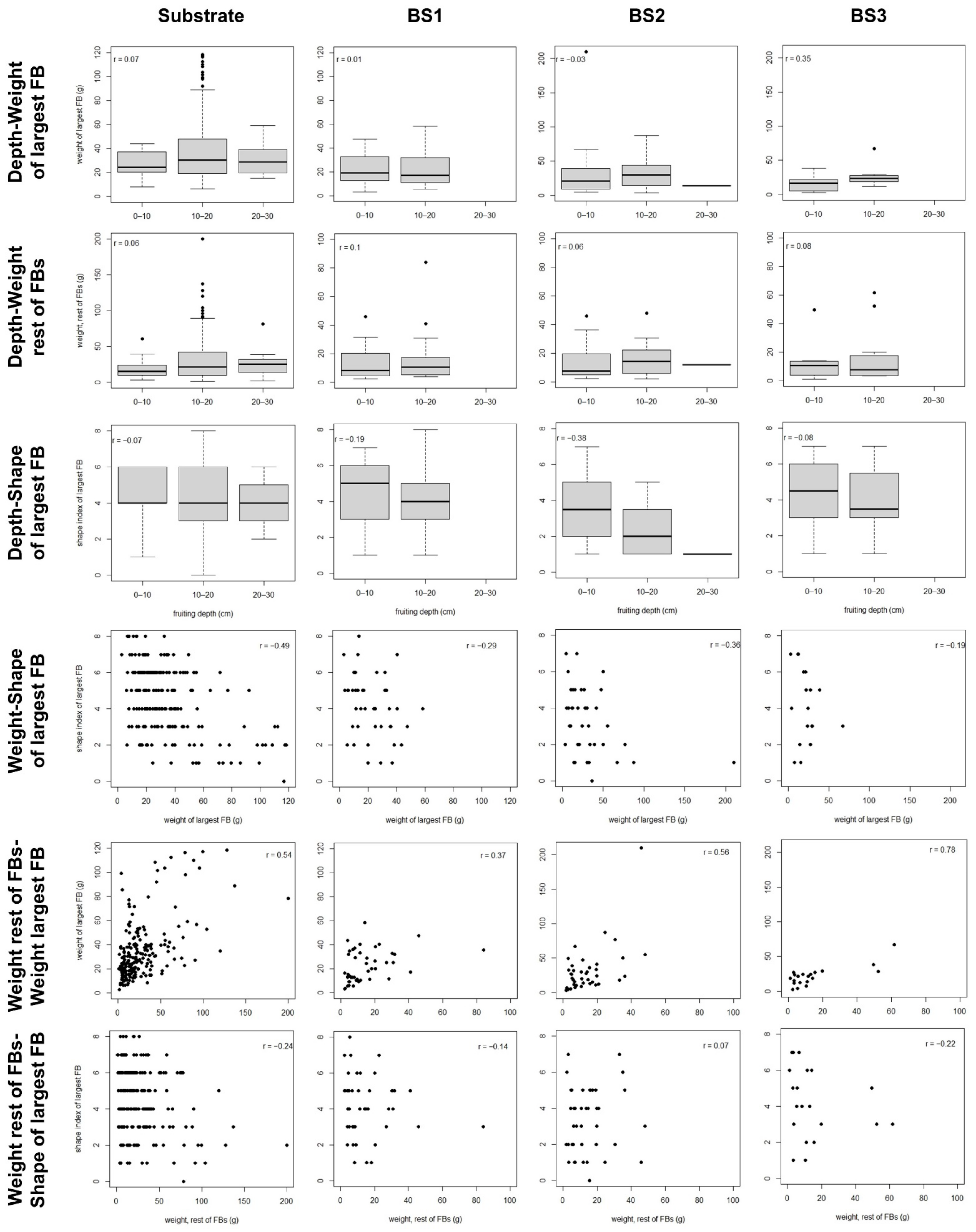
Finally, a positive relationship between the weight of the largest FB and the weight of the remaining FBs was clear among the FB clusters of all the analysed soils. This was not expected. We had initially hypothesized that local resource depletion would trigger compensating mechanisms between FB survival and FB size in a dig, in consonance with the fact that, before summer, the density of
T. melanosporum FBs (still immature) in the soil is much higher than the density of ripe FBs localized by dogs in the fruiting season [
1,
42]. The positive relationship we found could be explained if the growth of the largest FB was enhanced by the remaining FBs loosening the surrounding soil. However, the relationship between these development characters is very strong in the substrate, which is much looser than the bulk soil.
Alternatively, the positive relationship between the weights of the FBs in a dig could be related to local conditions in the soil microenvironment or to carbon transfer from the host. For other fungi, Moore et al. [
20] posited that once a fungal genet begins to fruit the distribution of FBs is controlled by a flow of resources towards particular FBs rather than by local depletion of nutrients or by inhibition mechanisms. For hypogeous fungal communities in natural forests, Hunt and Trappe [
43] and Luoma [
35] found that, at a small plot level (4 m
2), the relationship between total biomass and number of FBs was positive and not plateauing (although with low R
2), either for the whole hypogeous community or for particular species. The hypothetical genetic influence on weight could play a role, considering that FBs in a cluster are likely to share maternal genetic material [
2].
Overall, our results were noticeably consistent among the studied soil typologies, despite the fact that environmental gradients linked to soil depth and soil typologies influence the mating, FB survival, weight and number of FBs per dig [
1,
21]. The relationships among FB characters linked to different stages were, in general, not significant or of low magnitude. The lack of causal links among subsequent stages could be generalized for
T. melanosporum, with the existence of morphogenetic stage switches governing which environmental or physiological cues play a role in each stage. Alternatively, the lack of causal links among stages could be specific to low-stress conditions for FB formation. Cultural practices (e.g., irrigation, soil tillage, and peat amendments) reduce the impact of stressful, limiting factors (e.g., plant competition for light, rhizosphere diversity, nutrients or space, summer aridity or high soil resistance to penetration). In herbaceous crops, compensating mechanisms among yield-related traits are typically higher in strong-stress than in low-stress conditions [
27]. It would be interesting to test whether our results can be generalized to high-stress environments such as rainfed orchards or wild truffle stands. The genetic structure of
T. melanosporum population in the experimental site could also play a role, considering that truffle orchards usually show high genetic diversity due to nursery and field practices [
2].
Our approach did not allow to identify which specific environmental or physiological conditions influence development characters of FBs. Among the environmental factors potentially involved, not only abiotic factors but also microbial communities could play a critical role. The nature of the relationships between a fungus and the associated microbial communities is complex, ranging from defense against competition to modulation of physiological or developmental processes [
44,
45]. Kues and Liu [
46] suggested that in certain saprophytic taxa, such as cultivated
Agaricus spp., the presence of pseudomonads was essential for fruiting events. In other cases, some microorganisms influence the fruiting process by eliminating inhibitory compounds [
47]. In this regard, truffle FBs and mycorrhizae harbor a poorly-understood microflora that could play a role in the mating, growing or maturation [
48,
49,
50]. Further research on transcriptional gene expression throughout the distinct morphogenetic stages would greatly help identify the environmental and/or physiological factors governing each phase [
7]. This would also help understanding how this genetic program is switched on, whether it is expressed constitutively or cumulatively, and the intensity with which the fungus perceives the different environmental inputs that model these morphogenetic processes.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}