
Re-Evaluation of the Podosphaera tridactyla Species Complex in Australia

 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Fungal Sampling and Morphological Characterisation
2.2. DNA Extraction and Quantification
2.3. Library Preparation and Sequencing
2.4. Sequence Analysis
2.5. Phylogenetic Analysis
3. Results
3.1. Sequencing
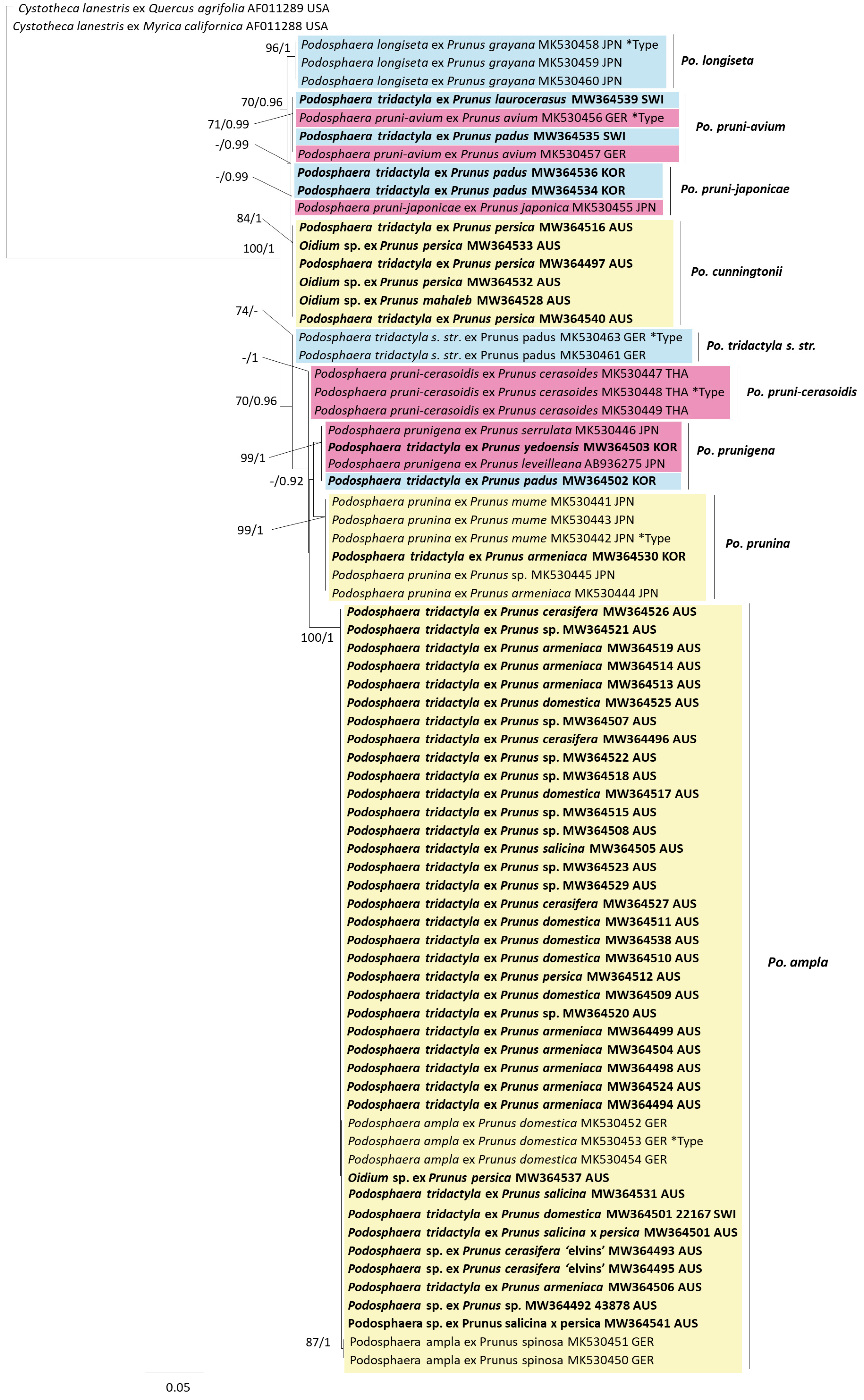
3.2. Phylogeny
3.3. Fungus–Host Relationships
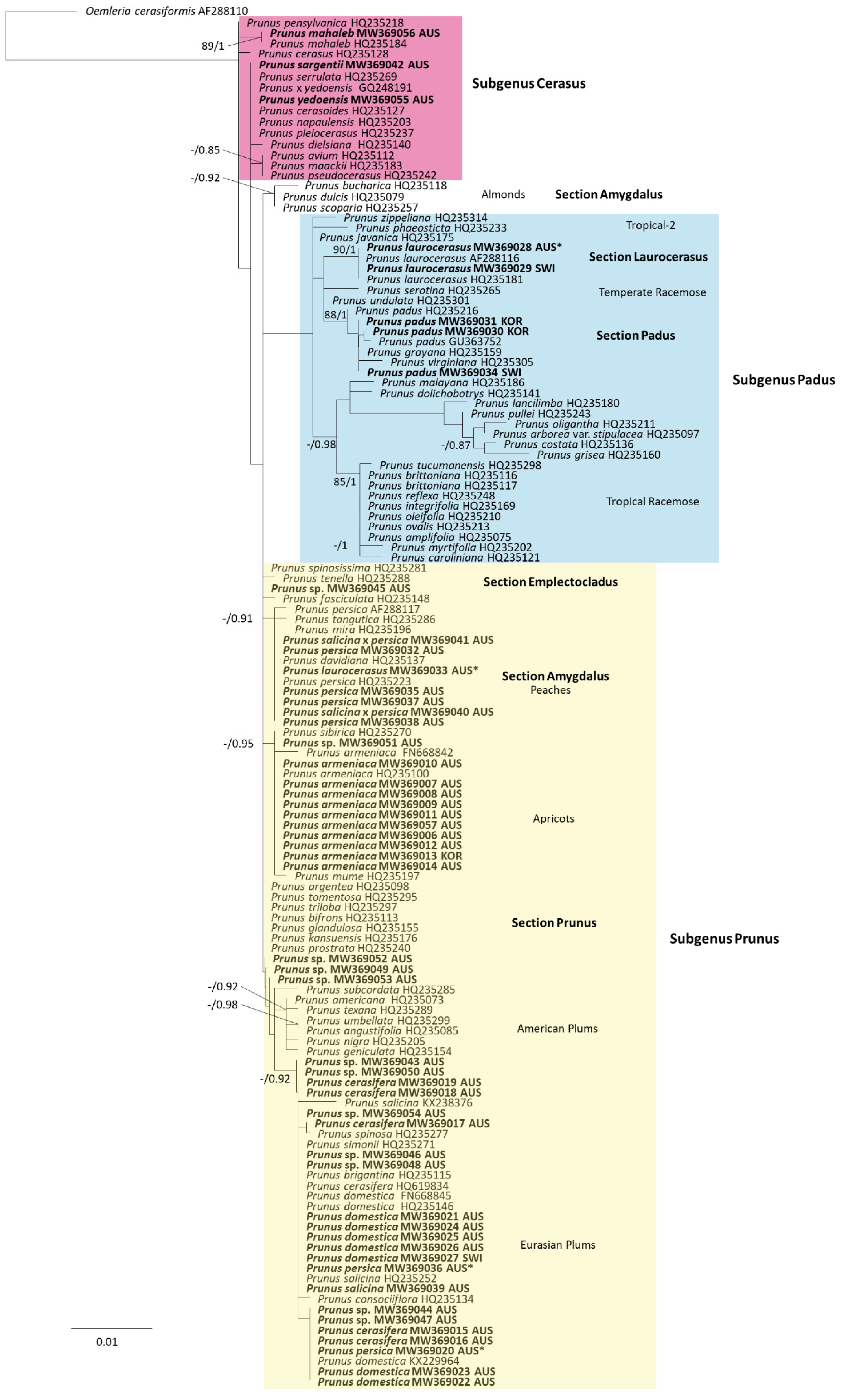
3.4. Plant Host Phylogeny
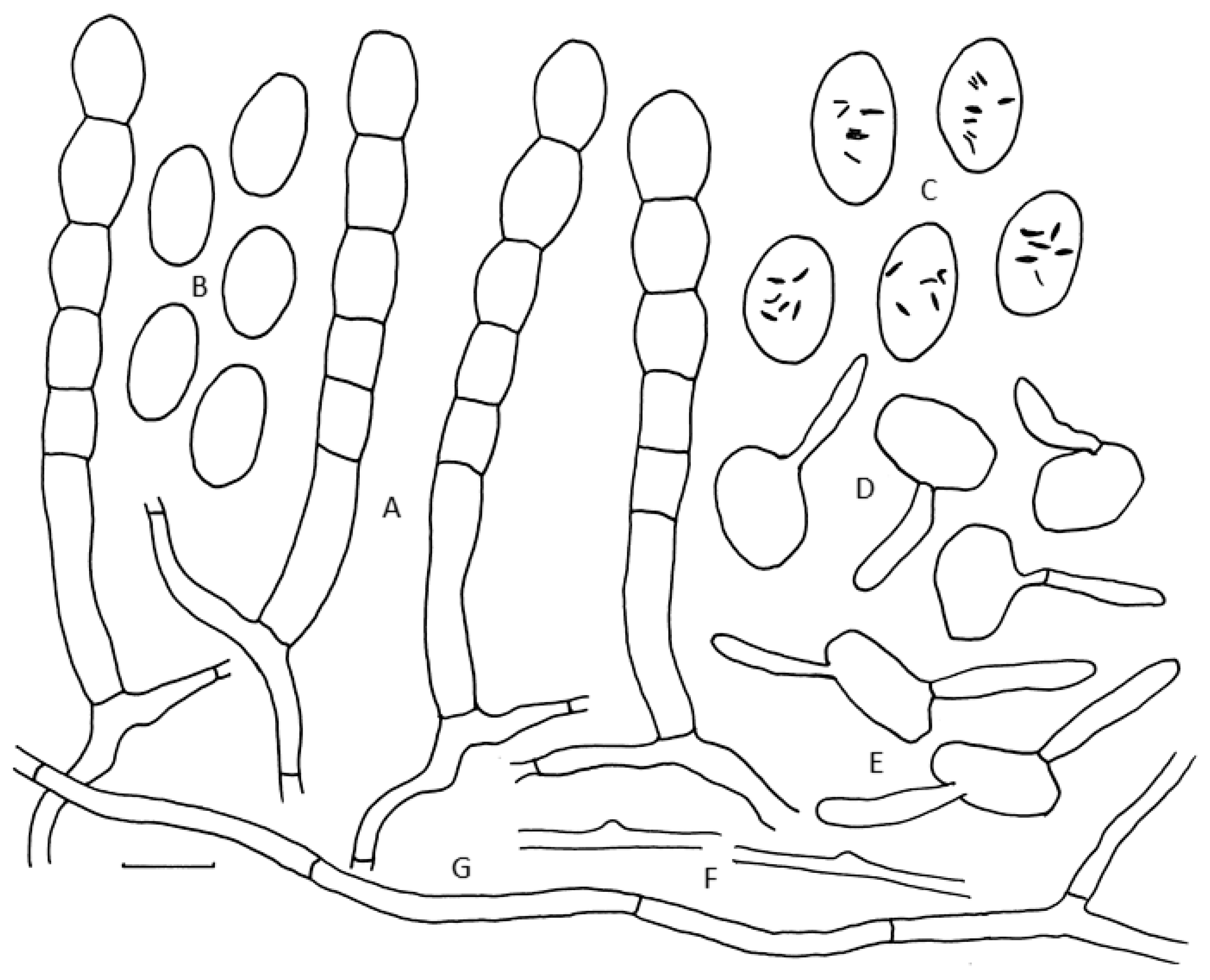
3.5. Morphological Characterisation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jun, W.; Berggren, S.T.; Chung-Hee, L.; Ickert-Bond, S.; Ting-Shuang, Y.; Ki-Oug, Y.; Lei, X.; Joey, S.; Potter, D. Phylogenetic inferences in Prunus (Rosaceae) using chloroplast ndhF and nuclear ribosomal ITS sequences. J. Syst. Evol. 2008, 46, 322–332. [Google Scholar] [CrossRef]
- Jeanes, J.A.; Jobson, P.C. Rosaceae. In Flora of Victoria, Dicotyledons Winteraceae to Myrtaceae; Walsh, N.G., Entwisle, T.J., Eds.; Inkata Press: Melbourne, Australia, 1996; Volume 3. [Google Scholar]
- Shi, S.; Li, J.; Sun, J.; Yu, J.; Zhou, S. Phylogeny and classification of Prunus sensu lato (Rosaceae). J. Integr. Plant Biol. 2013, 55, 1069–1079. [Google Scholar] [CrossRef]
- Chin, S.-W.; Shaw, J.; Haberle, R.; Wen, J.; Potter, D. Diversification of almonds, peaches, plums and cherries–molecular systematics and biogeographic history of Prunus (Rosaceae). Mol. Phylogenetics Evol. 2014, 76, 34–48. [Google Scholar] [CrossRef] [PubMed]
- George, A. Deciduous fruit production in Australia. RAP Publ. (FAO) Eng No. 1999/10. Available online: http://www.fao.org/3/ab985e/ab985e04.htm (accessed on 21 September 2020).
- Horticulture Innovation Australia Limited. Australian Horticulture Statistics Handbook Fruit 2017/2018; Freshlogic: Sydney, Australia, 2019; pp. 91–95. [Google Scholar]
- Braun, U.; Cook, R.T.A. Taxonomic Manual of the Erysiphales (Powdery Mildews); CBS-KNAW Fungal Biodiversity Center: Utrecht, The Netherlands, 2012; Volume 1, p. 2. [Google Scholar]
- Jeon, C.; Kang, S.; Choi, W.; Shin, H.-D. First report of powdery mildew caused by Podosphaera tridactyla on Prunus davidiana in Korea. Plant Dis. 2019, 103, 158. [Google Scholar] [CrossRef]
- García-Ruiz, M.; Galicia-Buendia, N.; Leyva-Mir, S.; Correia, K.; Nieto-López, E.; Camacho-Tapia, M.; Tovar-Pedraza, J. First report of powdery mildew of European plum caused by Podosphaera tridactyla in Mexico. Plant Dis. 2019, 103, 587. [Google Scholar] [CrossRef]
- Cunnington, J.H.; Lawrie, A.C.; Pascoe, I.G. Genetic variation within Podosphaera tridactyla reveals a paraphyletic species complex with biological specialization towards specific Prunus subgenera. Mycol. Res. 2005, 109, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Meeboon, J.; Takamatsu, S.; Braun, U. Morphophylogenetic analysis revealed that Podosphaera tridactyla constitutes a species complex. Mycologia 2020, 112, 244–266. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.L.; Sawbridge, T.; Mann, R.; Kaur, J.; May, T.W.; Edwards, J. Rediscovering an old foe: Optimised molecular methods for DNA extraction and sequencing applications for fungarium specimens of powdery mildew (Erysiphales). PLoS ONE 2020, 15, e0232535. [Google Scholar] [CrossRef] [PubMed]
- Johnston, A.; Booth, C. Plant Pathologist’s Pocketbook; C.M.I.: Kew, UK, 1983. [Google Scholar]
- Shin, H.-D. Erysiphaceae of Korea. Ph.D. Thesis, Seoul National University, Seoul, Korea, 1988. [Google Scholar]
- Shin, H.-D.; LA, Y.-J. Morphology of edge lines of chained immature conidia on conidiophores in powdery mildew fungi and their taxonomic significance. Mycotaxon 1993, 46, 445–451. [Google Scholar]
- Moparthi, S.; Grove, G.G.; Pandey, B.; Bradshaw, M.; Latham, S.R.; Braun, U.; Meeboon, J.; Romberg, M. Phylogeny and taxonomy of Podosphaera cerasi, sp. nov., and Podosphaera prunicola sensu lato. Mycologia 2019, 111, 647–659. [Google Scholar] [CrossRef]
- Smith, R.L.; May, T.W.; Kaur, J.; Sawbridge, T.; Mann, R.; Edwards, J. Molecular data from up to 130-year-old herbarium specimens do not support the presence of cherry powdery mildew in Australia. Plant Pathol. 2020. [Google Scholar] [CrossRef]
- Biomatters Ltd. Geneious Prime 2020.0.5. Available online: https://www.geneious.com/ (accessed on 3 August 2020).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. Available online: http://146.6.100.192/users/CH391L/Handouts/MRBAYES.pdf (accessed on 15 January 2020). [CrossRef] [Green Version]
- Leus, L.; Dewitte, A.; Van Huylenbroeck, J.; Vanhoutte, N.; Van Bockstaele, E.; Höfte, M. Podosphaera pannosa (syn. Sphaerotheca pannosa) on Rosa and Prunus spp.: Characterization of pathotypes by differential plant reactions and ITS sequences. J. Phytopathol. 2006, 154, 23–28. [Google Scholar] [CrossRef]
- Jankovics, T.; Dolovac, N.; Bulajić, A.; Krstić, B.; Pascal, T.; Bardin, M.; Nicot, P.C.; Kiss, L. Peach rusty spot is caused by the apple powdery mildew fungus, Podosphaera leucotricha. Plant Dis. 2011, 95, 719–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marimon, N.; Eduardo, I.; Martínez-Minaya, J.; Vicent, A.; Luque, J. Disease progression of peach powdery mildew in Catalonia, Spain–Towards a decision support system based on degree-days to initiate fungicide spray programs. 2020. Available online: http://hdl.handle.net/20.500.11824/1148 (accessed on 14 November 2020).
- Lu, W.; Newlands, N.K.; Carisse, O.; Atkinson, D.E.; Cannon, A.J. Disease risk forecasting with Bayesian learning networks: Application to grape powdery mildew (Erysiphe necator) in Vineyards. Agronomy 2020, 10, 622. [Google Scholar] [CrossRef]
- Asalf, B.; Onofre, R.B.; Gadoury, D.M.; Peres, N.A.; Stensvand, A. Pulsed water mists for suppression of strawberry powdery mildew. Plant Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kiss, L.; Vaghefi, N.; Bransgrove, K.; Dearnaley, J.D.W.; Takamatsu, S.; Tan, Y.P.; Marston, C.; Liu, S.-Y.; Jin, D.-N.; Adorada, D.L.; et al. Australia: A continent without native powdery mildews? The first comprehensive catalog indicates recent introductions and multiple host range expansion events, and leads to the re-discovery of Salmonomyces as a new lineage of the Erysiphales. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Walker, J. Pacific mycogeography: Deficiencies and irregularities in the distribution of plant parasitic fungi. Aust. J. Bot. Suppl. Ser. 1983, 13, 89–136. [Google Scholar] [CrossRef]
- Schindel, D.E.; Cook, J.A. The next generation of natural history collections. PLoS Biol. 2018, 16, e2006125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyde, K.D.; Chomnunti, P.; Crous, P.W.; Groenewald, J.Z.; Damm, U.; Ko, T.K.; Shivas, R.G.; Summerell, B.A.; Tan, Y.P. A case for re-inventory of Australia’s plant pathogens. Persoonia 2010, 25, 50–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shivas, R.; Beasley, D.; Pascoe, I.; Cunnington, J.; Pitkethley, R.; Priest, M. Specimen-based databases of Australian plant pathogens: Past, present and future. Australas. Plant Pathol. 2006, 35, 195–198. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank ITS+28S | GenBank matK | |||||
|---|---|---|---|---|---|---|
| Collection Number | Date | Country | Fungus | Host | ||
| BRIP 8323 | 1958 | Australia | Po. tridactyla | MW364487 | Pr. persica (AM) | MW369028 |
| BRIP 15118 | 1986 | Australia | Oidium sp. | MW364537 | Pr. persica (AM) | MW369020 |
| DAR 12478 | 1962 | Australia | Oidium sp. | MW364486 | Pr. armeniaca (PR) | MW369006 |
| DAR 28962 | 1977 | Australia | Oidium sp. | MW364533 | Pr. persica (AM) | MW369032 |
| DAR 28963 | 1977 | Australia | Oidium sp. | MW364528 | Pr. mahaleb (CR) | MW369056 |
| DAR 35281 | 1980 | Australia | Oidium sp. | - | Pr. laurocerasus (LR) | - |
| DAR 64667 | 1989 | Australia | Po. tridactyla | - | Pr. persica (AM) | - |
| DAR 71638 | 1996 | Australia | Oidium sp. | MW364532 | Pr. laurocerasus (LR) | MW369033 |
| VPRI 12495 | 1984 | Australia | Po. tridactyla | MW364488 | Pr. persica (AM) | MW369060 |
| VPRI 18452 | 1992 | Australia | Po. tridactyla | MW364489 | Prunus sp. | MW369058 |
| VPRI 18514 | 1992 | Australia | Po. tridactyla | MW364490 | Pr. domestica (PR) | MW369061 |
| VPRI 18600 | 1992 | Australia | Po. tridactyla | MW364521 | Prunus sp. | MW369043 |
| VPRI 18885 | 1993 | Australia | Po. tridactyla | MW364522 | Prunus sp. | MW369044 |
| VPRI 19000 | 1993 | Australia | Po. tridactyla | MW364523 | Prunus sp. | MW369045 |
| VPRI 19006 | 1993 | Australia | Po. tridactyla | MW364529 | Prunus sp. | MW369050 |
| VPRI 19008 | 1993 | Australia | Po. tridactyla | MW364524 | Pr. armeniaca (PR) | MW369007 |
| VPRI 19015 | 1993 | Australia | Po. tridactyla | MW364538 | Pr. domestica (PR) | MW369021 |
| VPRI 19017 | 1993 | Australia | Po. tridactyla | MW364525 | Pr. domestica (PR) | MW369022 |
| VPRI 19022 | 1993 | Australia | Po. tridactyla | MW364520 | Prunus sp. | MW369046 |
| VPRI 19164 | 1993 | Australia | Po. tridactyla | MW364519 | Pr. armeniaca (PR) | MW369008 |
| VPRI 19238 | 1993 | Australia | Po. tridactyla | MW364526 | Pr. cerasifera (PR) | MW369015 |
| VPRI 19248 | 1993 | Australia | Po. tridactyla | MW364518 | Prunus sp. | MW369047 |
| VPRI 19319 | 1993 | Australia | Po. tridactyla | MW364517 | Pr. domestica (PR) | MW369023 |
| VPRI 19591 | 1993 | Australia | Po. tridactyla | MW364516 | Pr. persica (AM) | MW369035 |
| VPRI 19788 | 1994 | Australia | Po. tridactyla | MW364515 | Prunus sp. | MW369048 |
| VPRI 19837 | 1994 | Australia | Po. tridactyla | MW364527 | Pr. cerasifera (PR) | MW369016 |
| VPRI 19864 | 1994 | Australia | Po. tridactyla | MW364514 | Pr. armeniaca (PR) | MW369009 |
| VPRI 19865 | 1994 | Australia | Po. tridactyla | MW364513 | Pr. armeniaca (PR) | MW369010 |
| VPRI 19868 | 1994 | Australia | Po. tridactyla | MW364512 | Pr. persica (AM) | MW369036 |
| VPRI 19871 | 1994 | Australia | Po. tridactyla | MW364511 | Pr. domestica (PR) | MW369024 |
| VPRI 19872 | 1994 | Australia | Po. tridactyla | MW364510 | Pr. domestica (PR) | MW369025 |
| VPRI 19873 | 1994 | Australia | Po. tridactyla | MW364509 | Pr. domestica (PR) | MW369026 |
| VPRI 20027 | 1994 | Australia | Po. tridactyla | MW364508 | Prunus sp. | MW369049 |
| VPRI 20040 | 1994 | Australia | Po. tridactyla | MW364507 | Prunus sp. | MW369051 |
| VPRI 20041 | 1994 | Australia | Po. tridactyla | MW364506 | Prunus sp. | MW369052 |
| VPRI 20045 | 1994 | Australia | Po. tridactyla | MW364505 | Pr. salicina (PR) | MW369039 |
| VPRI 20097 | 1994 | Australia | Po. tridactyla | MW364504 | Pr. armeniaca (PR) | MW369011 |
| VPRI 20231 | 1990 | South Korea | Po. tridactyla | MW364503 | Pr. yedoensis (CR) | MW369055 |
| VPRI 20233 | 1993 | South Korea | Po. tridactyla | MW364534 | Pr. padus (PD) | MW369030 |
| VPRI 20490 | 1993 | South Korea | Po. tridactyla | MW364536 | Pr. padus (PD) | MW369031 |
| VPRI 20491 | 1993 | South Korea | Po. tridactyla | MW364502 | Pr. sargentii (CR) | MW369042 |
| VPRI 20514 | 1995 | Australia | Po. tridactyla | MW364496 | Pr. cerasifera (PR) | MW369017 |
| VPRI 20687 | 1995 | Australia | Po. tridactyla | MW364491 | Prunus sp. | MW369059 |
| VPRI 20705 | 1995 | Australia | Po. tridactyla | MW364497 | Pr. persica (AM) | MW369037 |
| VPRI 20706 | 1995 | Australia | Po. tridactyla | MW364540 | Pr. persica (AM) | MW369038 |
| VPRI 20797 | 1996 | Australia | Po. tridactyla | MW364498 | Pr. armeniaca (PR) | MW369057 |
| VPRI 20811 | 1996 | Australia | Po. tridactyla | MW364499 | Pr. armeniaca (PR) | MW369012 |
| VPRI 21717 | 1998 | Australia | Po. tridactyla | MW364531 | Prunus sp. | MW369053 |
| VPRI 22156 | 1995 | Switzerland | Po. tridactyla | MW364539 | Pr. laurocerasus (LR) | MW369029 |
| VPRI 22159 | 1994 | Switzerland | Po. tridactyla | MW364535 | Pr. padus (PD) | MW369034 |
| VPRI 22167 | 1995 | Switzerland | Po. tridactyla | MW364500 | Pr. domestica (PR) | MW369027 |
| VPRI 22232 | 2000 | Australia | Po. tridactyla | MW364541 | Pr. salicina x persica (PR/AM) | MW369040 |
| VPRI 22233 | 2000 | Australia | Po. tridactyla | MW364501 | Pr. salicina x persica (PR/AM) | MW369041 |
| VPRI 41641 | 2006 | South Korea | Po. tridactyla | MW364530 | Pr. armeniaca (PR) | MW369013 |
| VPRI 43878 | 2018 | Australia | Podosphaera sp. | MW364492 | Prunus sp. | MW369054 |
| VPRI 43879 | 2018 | Australia | Podosphaera sp. | MW364493 | Pr. cerasifera “elvins” (PR) | MW369018 |
| VPRI 43880 | 2018 | Australia | Podosphaera sp. | MW364495 | Pr. cerasifera “elvins” (PR) | MW369019 |
| VPRI 43881 | 2018 | Australia | Podosphaera sp. | MW364494 | Pr. armeniaca (PR) | MW369014 |
| Collection Number | Original Fungus | Reidentified Fungus | Original Host | Reidentified Host |
|---|---|---|---|---|
| BRIP 8323 | Po. tridactyla | Po. pannosa | Pr. persica (AM) * | Pr. laurocerasus (LR) |
| BRIP 15118 | Oidium sp. | Po. ampla | Pr. persica (AM) * | Pr. domestica group (PR) |
| DAR 12478 | Oidium sp. | Po. pannosa | Pr. armeniaca (PR) | Pr. armeniaca (PR) |
| DAR 28962 | Oidium sp. | Po. cunningtonii | Pr. persica (AM) | Pr. persica (AM) |
| DAR 28963 | Oidium sp. | Po. cunningtonii | Pr. mahaleb (CR) | Pr. mahaleb (CR) |
| DAR 71638 | Oidium sp. | Po. cunningtonii | Pr. laurocerasus (LR) * | Pr. persica (AM) |
| VPRI 12495 | Po. tridactyla | Po. leucotricha | Pr. persica (AM) * | Malus prunifolia |
| VPRI 18452 | Oidium sp. | Po. leucotricha | Pr. laurocerasus (LR) * | M prunifolia |
| VPRI 18514 | Po. tridactyla | Po. leucotricha | Pr. persica (AM) * | M. prunifolia |
| VPRI 18600 | Po. tridactyla | Po. ampla | Prunus sp. | Pr. domestica group (PR) |
| VPRI 18885 | Po. tridactyla | Po. ampla | Pr. domestica (PR) | Pr. domestica group (PR) |
| VPRI 19000 | Po. tridactyla | Po. ampla | Prunus sp. | Pr. domestica group (PR) |
| VPRI 19006 | Po. tridactyla | Po. ampla | Prunus sp. | Pr. domestica group (PR) |
| VPRI 19008 | Po. tridactyla | Po. ampla | Prunus sp. | Pr. armeniaca (PR) |
| VPRI 19015 | Po. tridactyla | Po. ampla | Prunus sp. | Pr. domestica group (PR) |
| VPRI 19017 | Po. tridactyla | Po. ampla | Pr. armeniaca (PR) | Pr. domestica group (PR) |
| VPRI 19022 | Po. tridactyla | Po. ampla | Pr. domestica (PR) | Pr. domestica group (PR) |
| VPRI 19164 | Po. tridactyla | Po. ampla | Pr. domestica (PR) | Pr. armeniaca (PR) |
| VPRI 19238 | Po. tridactyla | Po. ampla | Prunus sp. | Pr. domestica group (PR) |
| VPRI 19248 | Po. tridactyla | Po. ampla | Pr. armeniaca (PR) | Pr. domestica group (PR) |
| VPRI 19319 | Po. tridactyla | Po. ampla | Pr. cerasifera (PR) | Pr. domestica group (PR) |
| VPRI 19591 | Po. tridactyla | Po. cunningtonii | Prunus sp. | Pr. persica (AM) |
| VPRI 19788 | Po. tridactyla | Po. ampla | Pr. domestica (PR) | Pr. domestica group (PR) |
| VPRI 19837 | Po. tridactyla | Po. ampla | Pr. persica (AM) | Pr. domestica group (PR) |
| VPRI 19864 | Po. tridactyla | Po. ampla | Prunus sp. | Pr. armeniaca (PR) |
| VPRI 19865 | Po. tridactyla | Po. ampla | Pr. cerasifera (PR) | Pr. armeniaca (PR) |
| VPRI 19868 | Po. tridactyla | Po. ampla | Pr. armeniaca (PR) * | Pr. domestica group (PR) |
| VPRI 19871 | Po. tridactyla | Po. ampla | Pr. armeniaca (PR) | Pr. domestica group (PR) |
| VPRI 19872 | Po. tridactyla | Po. ampla | Pr. persica (AM) * | Pr. domestica group (PR)* |
| VPRI 19873 | Po. tridactyla | Po. ampla | Pr. domestica (PR) | Pr. domestica group (PR) |
| VPRI 20027 | Po. tridactyla | Po. ampla | Pr. domestica (PR) | Pr. domestica group (PR) |
| VPRI 20040 | Po. tridactyla | Po. ampla | Pr. domestica (PR) | Pr. domestica group (PR) |
| VPRI 20041 | Po. tridactyla | Po. ampla | Prunus sp. | Pr. domestica group (PR) |
| VPRI 20045 | Po. tridactyla | Po. ampla | Prunus sp. | Pr. domestica group (PR) |
| VPRI 20097 | Po. tridactyla | Po. ampla | Prunus sp. | Pr. domestica group (PR) |
| VPRI 20231 | Po. tridactyla | Po. prunigena | Pr. salicina (PR) | Pr. yedoensis (CR) |
| VPRI 20233 | Po. tridactyla | Po. pruni-japonicae | Pr. armeniaca (PR) | Pr. padus (PD) |
| VPRI 20490 | Po. tridactyla | Po. pruni-japonicae | Pr. yedoensis (CR) | Pr. padus (PD) |
| VPRI 20491 | Po. tridactyla | Po. prunigena | Pr. padus (PD) | Pr. sargentii (CR) |
| VPRI 20514 | Po. tridactyla | Po. ampla | Pr. padus (PD) | Pr. domestica group (PR) |
| VPRI 20687 | Po. tridactyla | Po. leucotricha | Pr. sargentii (CR) * | M. prunifolia |
| VPRI 20705 | Po. tridactyla | Po. cunningtonii | Pr. cerasifera (PR) | Pr. persica (AM) |
| VPRI 20706 | Po. tridactyla | Po. cunningtonii | Prunus sp. | Pr. persica (AM) |
| VPRI 20797 | Po. tridactyla | Po. ampla | Pr. persica (AM) | Pr. armeniaca (PR) |
| VPRI 20811 | Po. tridactyla | Po. ampla | Pr. armeniaca (PR) | Pr. armeniaca (PR) |
| VPRI 21717 | Po. tridactyla | Po. ampla | Prunus sp. | Pr. domestica group (PR) |
| VPRI 22156 | Po. tridactyla | Po. pruni-avium | Pr. laurocerasus (LR) | Pr. laurocerasus (LR) |
| VPRI 22159 | Po. tridactyla | Po. pruni-avium | Pr. padus (PD) | Pr. padus (PD) |
| VPRI 22167 | Po. tridactyla | Po. ampla | Pr. domestica (PR) | Pr. domestica group (PR) |
| VPRI 22232 | Po. tridactyla | Po. ampla | Pr. salicina x persica (PR/AM) | Pr. persica (AM) |
| VPRI 22233 | Po. tridactyla | Po. ampla | Pr. salicina x persica (PR/AM) | Pr. persica (AM) |
| VPRI 41641 | Po. tridactyla | Po. prunina | Pr. armeniaca (PR) | Pr. armeniaca (PR) |
| VPRI 43878 | Podosphaera sp. | Po. ampla | Prunus sp. | Pr. domestica group (PR) |
| VPRI 43879 | Podosphaera sp. | Po. ampla | Pr. cerasifera “elvins” (PR) | Pr. domestica group (PR) |
| VPRI 43880 | Podosphaera sp. | Po. ampla | Pr. cerasifera “elvins” (PR) | Pr. domestica group (PR) |
| VPRI 43881 | Podosphaera sp. | Po. ampla | Pr. armeniaca (PR) | Pr. armeniaca (PR) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smith, R.L.; May, T.W.; Kaur, J.; Sawbridge, T.I.; Mann, R.C.; Pascoe, I.G.; Edwards, J. Re-Evaluation of the Podosphaera tridactyla Species Complex in Australia. J. Fungi 2021, 7, 171. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030171
Smith RL, May TW, Kaur J, Sawbridge TI, Mann RC, Pascoe IG, Edwards J. Re-Evaluation of the Podosphaera tridactyla Species Complex in Australia. Journal of Fungi. 2021; 7(3):171. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030171
Chicago/Turabian StyleSmith, Reannon L., Tom W. May, Jatinder Kaur, Tim I. Sawbridge, Ross C. Mann, Ian G. Pascoe, and Jacqueline Edwards. 2021. "Re-Evaluation of the Podosphaera tridactyla Species Complex in Australia" Journal of Fungi 7, no. 3: 171. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7030171