
In Vitro Activity of Novel Antifungal Olorofim against Filamentous Fungi and Comparison to Eight Other Antifungal Agents

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. CLSI Reference Method MIC Results for Olorofim, Azoles, Echinocandins and Amphotericin B against Molds
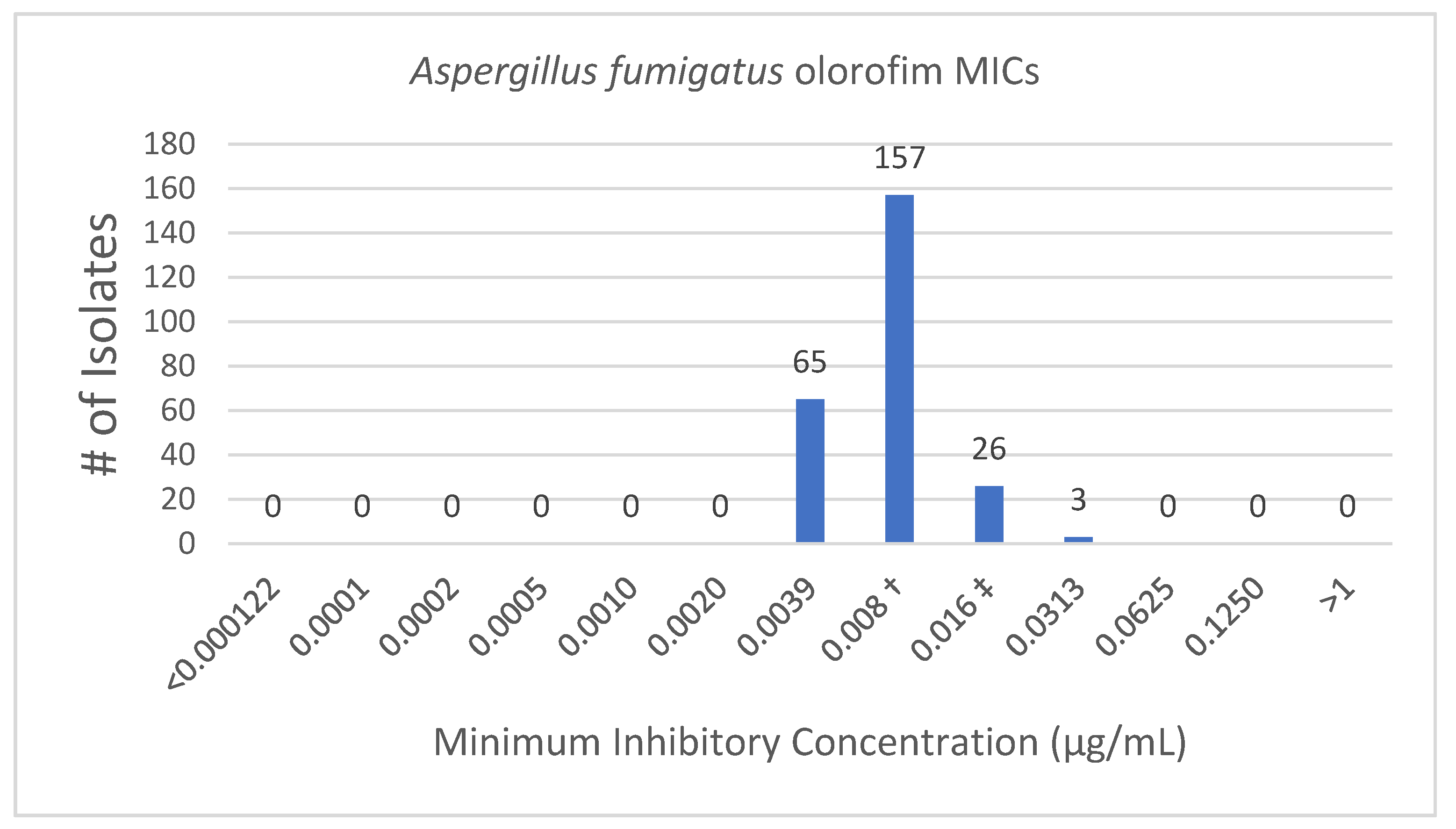
3.1.1. Olorofim
3.1.2. Azoles
3.1.3. Echinocandins
3.1.4. Amphotericin B
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Pfaller, M.A.; Pappas, P.G.; Wingard, J.R. Invasive Fungal Pathogens: Current Epidemiological Trends. Clin. Infect. Dis. 2006, 43, S3–S14. [Google Scholar] [CrossRef]
- Rauseo, A.M.; Coler-Reilly, A.; Larson, L.; Spec, A. Hope on the Horizon: Novel Fungal Treatments in Development. Open Forum Infect Dis. 2020, 7, ofaa016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallabhaneni, S.; Mody, R.K.; Walker, T.; Chiller, T. The Global Burden of Fungal Diseases. Infect Dis. Clin. N. Am. 2016, 30, 1–11. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [Green Version]
- Kosmidis, C.; Denning, D.W. The clinical spectrum of pulmonary aspergillosis. Thorax 2015, 70, 270–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verweij, P.E.; Mellado, E.; Melchers, W.J. Multiple-triazole-resistant aspergillosis. N. Engl. J. Med. 2007, 356, 1481–1483. [Google Scholar] [CrossRef] [Green Version]
- Lestrade, P.P.A.; Meis, J.F.; Melchers, W.J.G.; Verweij, P.E. Triazole resistance in Aspergillus fumigatus: Recent insights and challenges for patient management. Clin. Microbiol. Infect. 2019, 25, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Enoch, D.A.; Yang, H.; Aliyu, S.H.; Micallef, C. The Changing Epidemiology of Invasive Fungal Infections. Methods Mol. Biol. 2017, 1508, 17–65. [Google Scholar] [PubMed]
- Safdar, A.; Ma, J.; Saliba, F.; Dupont, B.; Wingard, J.R.; Hachem, R.Y.; Mattiuzzi, G.N.; Chandrasekar, P.H.; Kontoyiannis, D.P.; Rolston, K.V.; et al. Drug-induced nephrotoxicity caused by amphotericin B lipid complex and liposomal amphotericin B: A review and meta-analysis. Medicine (Baltimore) 2010, 89, 236–244. [Google Scholar] [CrossRef]
- Townsend, R.; Dietz, A.; Hale, C.; Akhtar, S.; Kowalski, D.; Lademacher, C.; Lasseter, K.; Pearlman, H.; Rammelsberg, D.; Schmitt-Hoffman, A.; et al. Pharmacokinetic Evaluation of CYP3A4-Mediated Drug-Drug Interactions of Isavuconazole with Rifampin, Ketoconazole, Midazolam, and Ethinyl Estradiol/Norethindrone in Healthy Adults. Clin. Pharmacol. Drug. Dev. 2017, 6, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Verweij, P.E.; Chowdhary, A.; Melchers, W.J.; Meis, J.F. Azole Resistance in Aspergillus fumigatus: Can We Retain the Clinical Use of Mold-Active Antifungal Azoles? Clin. Infect. Dis. 2016, 62, 362–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquierdo, A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet Infect. Dis. 2017, 17, e383–e392. [Google Scholar] [CrossRef]
- Chowdhary, A.; Kathuria, S.; Xu, J.; Meis, J.F. Emergence of azole-resistant aspergillus fumigatus strains due to agricultural azole use creates an increasing threat to human health. PLoS Pathog. 2013, 9, e1003633. [Google Scholar] [CrossRef]
- Resendiz Sharpe, A.; Lagrou, K.; Meis, J.F.; Chowdhary, A.; Lockhart, S.R.; Verweij, P.E. Triazole resistance surveillance in Aspergillus fumigatus. Med. Mycol. 2018, 56 (Suppl. S1), 83–92. [Google Scholar] [CrossRef] [Green Version]
- Hurst, S.F.; Berkow, E.L.; Stevenson, K.L.; Litvintseva, A.P.; Lockhart, S.R. Isolation of azole-resistant Aspergillus fumigatus from the environment in the south-eastern USA. J. Antimicrob. Chemother. 2017, 72, 2443–2446. [Google Scholar] [CrossRef]
- Wiederhold, N.P. Antifungal resistance: Current trends and future strategies to combat. Infect. Drug Resist. 2017, 10, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dannaoui, E. Antifungal resistance in mucorales. Int. J. Antimicrob. Agents 2017, 50, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Buil, J.B.; Rijs, A.J.M.M.; Meis, J.F.; Birch, M.; Law, D.; Melchers, W.J.G.; Verweij, P.E. In vitro activity of the novel antifungal compound F901318 against difficult-to-treat Aspergillus isolates. J. Antimicrob. Chemother. 2017, 72, 2548–2552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, J.D.; Sibley, G.E.M.; Beckmann, N.; Dobb, K.S.; Slater, M.J.; McEntee, L.; du Pré, S.; Livermore, J.; Bromley, M.J.; Wiederhold, N.P.; et al. F901318 represents a novel class of antifungal drug that inhibits dihydroorotate dehydrogenase. Proc. Natl. Acad. Sci. USA 2016, 113, 12809–12814. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, K.M.; Astvad, K.M.T.; Hare, R.K.; Arendrup, M.C. EUCAST Determination of Olorofim (F901318) Susceptibility of Mold Species, Method Validation, and MICs. Antimicrob. Agents Chemother. 2018, 62, e00487-18. [Google Scholar] [CrossRef] [Green Version]
- Wiederhold, N.P.; Najvar, L.K.; Jaramillo, R.; Olivo, M.; Birch, M.; Law, D.; Rex, J.H.; Catano, G.; Patterson, T.F. The Orotomide Olorofim Is Efficacious in an Experimental Model of Central Nervous System Coccidioidomycosis. Antimicrob. Agents Chemother. 2018, 62, e00999-18. [Google Scholar] [CrossRef] [Green Version]
- du Pré, S.; Beckmann, N.; Almeida, M.C.; Sibley, G.E.M.; Law, D.; Brand, A.C.; Birch, M.; Read, N.D.; Oliver, J.D. Effect of the Novel Antifungal Drug F901318 (Olorofim) on Growth and Viability of Aspergillus fumigatus. Antimicrob. Agents Chemother. 2018, 62, e00231-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivero-Menendez, O.; Cuenca-Estrella, M.; Alastruey-Izquierdo, A. In vitro activity of olorofim (F901318) against clinical isolates of cryptic species of Aspergillus by EUCAST and CLSI methodologies. J. Antimicrob. Chemother. 2019, 74, 1586–1590. [Google Scholar] [CrossRef] [Green Version]
- Seyedmousavi, S.; Chang, Y.C.; Law, D.; Birch, M.; Rex, J.H.; Kwon-Chung, K.J. Efficacy of Olorofim (F901318) against Aspergillus fumigatus, A. nidulans, and A. tanneri in Murine Models of Profound Neutropenia and Chronic Granulomatous Disease. Antimicrob. Agents Chemother. 2019, 63, e00129-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, W.; Eadie, K.; Konings, M.; Rijnders, B.; Fahal, A.H.; Oliver, J.D.; Birch, M.; Verbon, A.; van de Sande, W. Madurella mycetomatis, the main causative agent of eumycetoma, is highly susceptible to olorofim. J. Antimicrob. Chemother. 2020, 75, 936–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, C.; Law, D.; Birch, M.; Halliday, C.; Sorrell, T.C.; Rex, J.; Slavin, M.; Chen, S.C. In vitro activity of the novel antifungal compound F901318 against Australian Scedosporium and Lomentospora fungi. Med. Mycol. 2018, 56, 1050–1054. [Google Scholar] [CrossRef]
- Talbot, J.J.; Frisvad, J.C.; Meis, J.F.; Hagen, F.; Verweij, P.E.; Hibbs, D.E.; Lai, F.; Groundwater, P.W.; Samson, R.A.; Kidd, S.E.; et al. cyp51A Mutations, Extrolite Profiles, and Antifungal Susceptibility in Clinical and Environmental Isolates of the Aspergillus viridinutans Species Complex. Antimicrob. Agents Chemother. 2019, 63, e00632-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lackner, M.; Birch, M.; Naschberger, V.; Grässle, D.; Beckmann, N.; Warn, P.; Gould, J.; Law, D.; Lass-Flörl, C.; Binder, U. Dihydroorotate dehydrogenase inhibitor olorofim exhibits promising activity against all clinically relevant species within Aspergillus section Terrei. J. Antimicrob. Chemother. 2018, 73, 3068–3073. [Google Scholar] [CrossRef]
- Kennedy, T.; Graham, A.; Steiner, J. An open-label study in healthy volunteers to evaluate the potential for cytochrome P450 3A4 inhibition by F901318 using oral midazolam as a probe. In Proceedings of the ECCMID, Vienna, Austria, 22–25 April 2017. [Google Scholar]
- Guinea, J.; Verweij, P.E.; Meletiadis, J.; Mouton, J.W.; Barchiesi, F.; Arendrup, M.C. How to: EUCAST recommendations on the screening procedure E.Def 10.1 for the detection of azole resistance in Aspergillus fumigatus isolates using four-well azole-containing agar plates. Clin. Microbiol. Infect. 2019, 25, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Institute CaLS. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, M38, 3rd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Berkow, E.L.; Nunnally, N.S.; Bandea, A.; Kuykendall, R.; Beer, K.; Lockhart, S.R. Detection of TR34/L98H CYP51A Mutation through Passive Surveillance for Azole-Resistant Aspergillus fumigatus in the United States from 2015 to 2017. Antimicrob. Agents Chemother. 2018, 62, e02240-17. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.P.; Kirby, J.E. Verification of an Automated, Digital Dispensing Platform for At-Will Broth Microdilution-Based Antimicrobial Susceptibility Testing. J. Clin. Microbiol. 2016, 54, 2288–2293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arendrup, M.C.; Meletiadis, J.; Mouton, J.W.; Guinea, J.; Cuenca-Estrella, M.; Lagrou, K.; Howard, S.J. Subcommittee on Antifungal Susceptibility Testing (AFST) of the ESCMID European Committee for Antimicrobial Susceptibility Testing (EUCAST). EUCAST technical note on isavuconazole breakpoints for Aspergillus, itraconazole breakpoints for Candida and updates for the antifungal susceptibility testing method documents. Clin. Microbiol. Infect. 2016, 22, 571. [Google Scholar] [PubMed] [Green Version]
- Buil, J.B.; Oliver, J.; Law, D.; Tehupeiory-Kooreman, M.; Rex, J.H.; Hokken, M.W.J.; Melchers, W.J.G.; Birch, M.; Verweij, P.E. Molecular Mechanism and frequency of olorofim resistance in Aspergillus fumigatus. In Proceedings of the Trends in Medical Mycology bi-annual meeting of the European Confederation of Medical Mycology (ECMM), Nice, France, 11–14 October 2019. [Google Scholar]
- Wiederhold, N.P.; Patterson, H.P.; Birch, M.; Law, D.; Rex, J. Evaluation of the In Vitro Activity of Olorofim against Fusarium Species. In Proceedings of the Trends in Medical Mycology bi-annual meeting of the European Confederation of Medical Mycology (ECMM), Nice, France, 11–14 October 2019. [Google Scholar]
- Vehreschild, J.J.; Birtel, A.; Vehreschild, M.J.; Liss, B.; Farowski, F.; Kochanek, M.; Sieniawski, M.; Steinbach, A.; Wahlers, K.; Fätkenheuer, G.; et al. Mucormycosis treated with posaconazole: Review of 96 case reports. Crit. Rev. Microbiol. 2013, 39, 310–324. [Google Scholar] [CrossRef]
- Marty, F.M.; Ostrosky-Zeichner, L.; Cornely, O.A.; Mullane, K.M.; Perfect, J.R.; Thompson, G.R., 3rd; Alangaden, G.J.; Brown, J.M.; Fredricks, D.N.; Heinz, W.J.; et al. VITAL and FungiScope Mucormycosis Investigators. Isavuconazole treatment for mucormycosis: A single-arm open-label trial and case-control analysis. Lancet Infect. Dis. 2016, 16, 828–837. [Google Scholar] [CrossRef]
- Skiada, A.; Lass-Floerl, C.; Klimko, N.; Ibrahim, A.; Roilides, E.; Petrikkos, G. Challenges in the diagnosis and treatment of mucormycosis. Med. Mycol. 2018, 56 (Suppl. S1), 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Quality Control or Reference Strain | Species Tested |
|---|---|
| Aspergillus fumigatus ATCC MYA-3626 | Fusarium spp., Phialemonium curvatum, Phaeoacremonium parasiticum, Sarocladium kiliense, Ramularia sp., Metarrhizium anisopliae and Pleurostomophora richardsiae |
| Candida krusei ATCC 6258 | Rhizopus spp. |
| Azole susceptible A. fumigatus B7698 Azole resistant A. fumigatus B7815 (CDC internal controls) | A. fumigatus |
| Species | Olorofim | Voriconazole | Anidulafungin | Caspofungin | Itraconazole | Isavuconazole | Posaconazole | Micafungin | Amphotericin B |
|---|---|---|---|---|---|---|---|---|---|
| A. fumigatus WT (n = 246) | |||||||||
| MIC 50 | 0.008 | ||||||||
| Range | 0.004–0.03 | ||||||||
| A. fumigatus TR34/L98H (n = 5) | |||||||||
| MIC 50 | 0.008 | 2 | 16 | ||||||
| Range | 0.008–0.008 | 0.3–2 | 4–>16 | ||||||
| R. microsporus (n = 3) | |||||||||
| MIC 50 | >2 | 4 | 4 | 16 | 0.06 | 0.5 | 0.5 | >8 | 0.06 |
| Range | 2–>2 | 0.5–4 | 0.25–>16 | 0.25–>16 | 0.06–0.125 | 0.125–1 | 0.06–0.125 | 0.25–>8 | 0.016–0.125 |
| R. oryzae (n = 16) | |||||||||
| MIC 50 | >2 | 4 | >16 | >16 | 0.125 | 4 | 0.3 | >8 | 1.5 |
| Range | >2–>2 | 1–8 | 8–>16 | 16–>16 | 0.03–>16 | 0.25–>8 | 0.03–>16 | 4–>8 | 0.3–4 |
| F. chlamydosporum (n = 1) | |||||||||
| MIC 50 | >2 | 2 | >16 | >16 | >16 | >8 | >16 | >8 | 2 |
| Range | |||||||||
| F. dimerum (n = 2) | |||||||||
| MIC 50 | >2 | 4 | >16 | >16 | >16 | >8 | >16 | >8 | 4 |
| Range | 2–>2 | 4–4 | 8–>16 | 16–>16 | >16–>16 | 8–>8 | >16–>16 | 2–>8 | 3–4 |
| F. moniliforme (n = 1) | |||||||||
| MIC 50 | 0.03 | 2 | 0.06 | 0.125 | >16 | >8 | >16 | 0.02 | 0.5 |
| Range | |||||||||
| F. oxysporum (n = 5) | |||||||||
| MIC 50 | 2 | 16 | 16 | 16 | >16 | >8 | >16 | 8 | 24 |
| Range | 0.12–>2 | 4–>16 | 0.125–>16 | 0.125–>16 | >16–>16 | 8–>8 | 2–>16 | 0.06–>8 | 0.8–>32 |
| F. solani (n = 11) | |||||||||
| MIC 50 | >2 | 8 | 8 | 8 | >16 | >8 | >16 | 4 | >32 |
| Range | 2–>2 | 1–>16 | 1–>16 | 2–>16 | >16–>16 | >8–>8 | >16–>16 | 0.125–>8 | 1.5–>32 |
| F. verticilloides (n = 1) | |||||||||
| MIC 50 | 0.5 | 1 | >16 | >16 | 0.3 | 1 | 0.06 | >8 | >32 |
| Range | |||||||||
| M. anisopliae (n = 1) | |||||||||
| MIC 50 | 0.5 | 1 | 0.125 | 0.06 | >16 | 8 | >16 | <0.008 | >32 |
| Range | |||||||||
| P. parasiticum (n = 1) | |||||||||
| MIC 50 | 2 | 1 | 8 | >16 | >16 | 8 | 0.5 | >8 | 0.3 |
| Range | |||||||||
| P. curvatum (n = 1) | |||||||||
| MIC 50 | 2 | 1 | 8 | >16 | 0.5 | 8 | 0.125 | >8 | 0.125 |
| Range | |||||||||
| P. richardsiae (n = 1) | |||||||||
| MIC 50 | 0.06 | 0.4 | |||||||
| Range | |||||||||
| Ramularia species (n = 1) | |||||||||
| MIC 50 | 0.015 | 1 | <0.008 | <0.008 | 0.5 | 1 | 0.3 | <0.008 | 16 |
| Range | |||||||||
| S. kiliense (n = 1) | |||||||||
| MIC 50 | 0.5 | 2 | 8 | 0.5 | >16 | >8 | >16 | 4 | 12 |
| Range |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgacopoulos, O.; Nunnally, N.S.; Ransom, E.M.; Law, D.; Birch, M.; Lockhart, S.R.; Berkow, E.L. In Vitro Activity of Novel Antifungal Olorofim against Filamentous Fungi and Comparison to Eight Other Antifungal Agents. J. Fungi 2021, 7, 378. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7050378
Georgacopoulos O, Nunnally NS, Ransom EM, Law D, Birch M, Lockhart SR, Berkow EL. In Vitro Activity of Novel Antifungal Olorofim against Filamentous Fungi and Comparison to Eight Other Antifungal Agents. Journal of Fungi. 2021; 7(5):378. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7050378
Chicago/Turabian StyleGeorgacopoulos, Ourania, Natalie S. Nunnally, Eric M. Ransom, Derek Law, Mike Birch, Shawn R. Lockhart, and Elizabeth L. Berkow. 2021. "In Vitro Activity of Novel Antifungal Olorofim against Filamentous Fungi and Comparison to Eight Other Antifungal Agents" Journal of Fungi 7, no. 5: 378. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7050378