
Differential Expression of Amanitin Biosynthetic Genes and Novel Cyclic Peptides in Amanita molliuscula


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample
2.2. Phylogenetic Tree Construction
2.3. Genome Sequencing and Assembly
2.4. Transcriptome Sequencing and Analysis
2.5. Venn Diagram Construction
2.6. Visualization of the Genome
2.7. LC-MS and LC-MS/MS Analyses for Novel Cyclic Peptides
2.8. Cloning of MSDIN and POPB Genes in A. molliuscula
3. Results
3.1. Phylogenetic Position of A. molliuscula
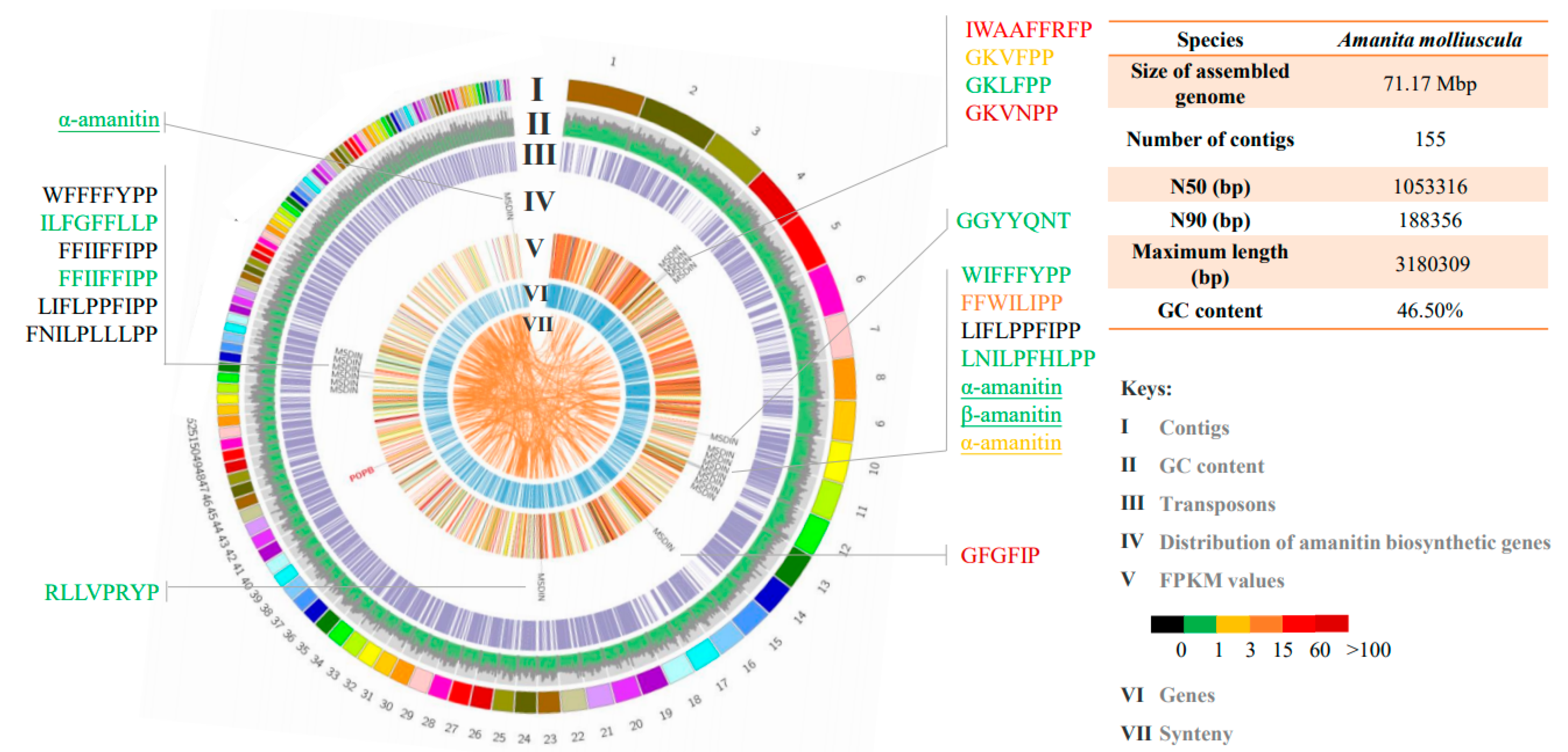
3.2. Genome and Transcriptome of A. molliuscula
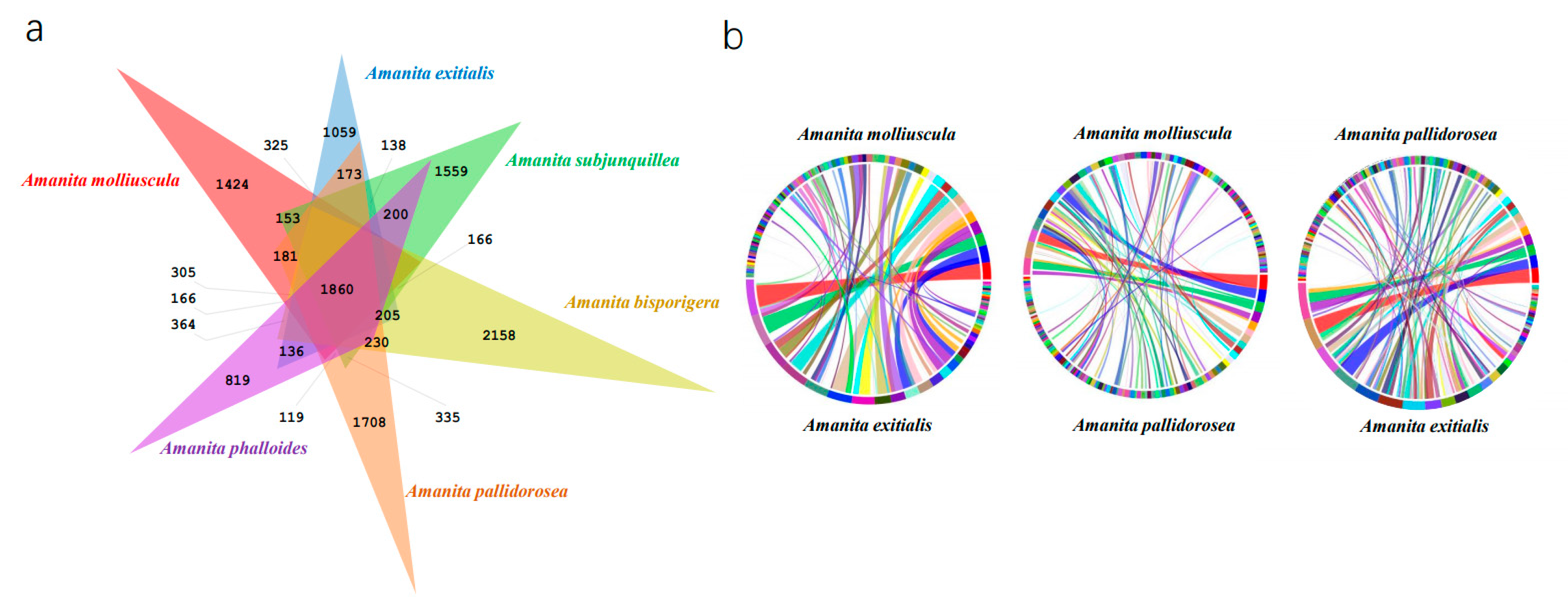
3.3. Orthology and Synteny Analysis of A. molliuscula
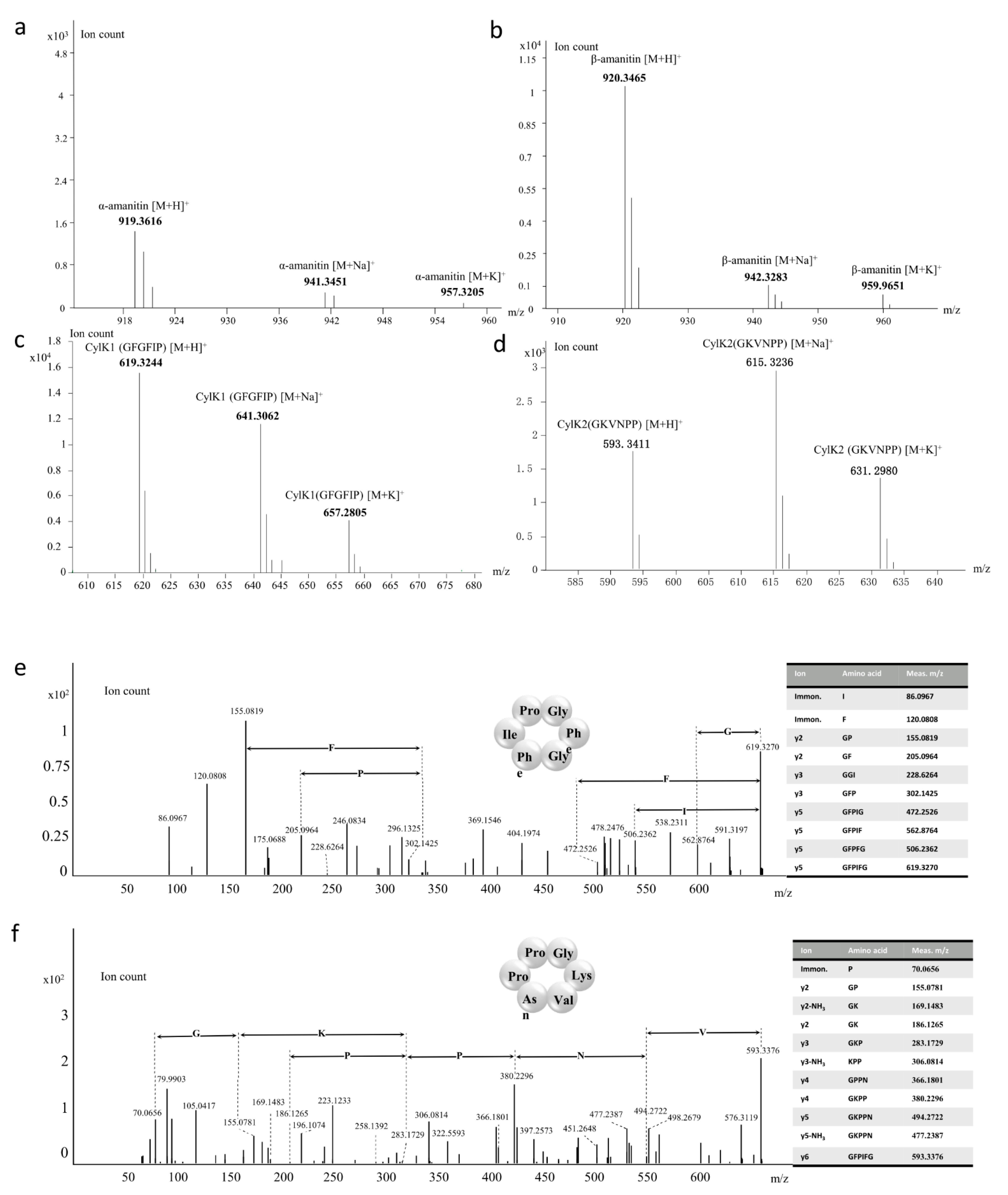
3.4. Amatoxins and Novel Cyclic Peptides from A. molliuscula
3.5. Structures of AmPOPB, AmAMA1, AmAMA2, AmAMA3, AmCylK1 and AmCylK2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wieland, T. Peptides of Poisonous Amanita Mushrooms; Springer: Berlin/Heidelberg, Germany, 1986. [Google Scholar]
- Li, H.J.; Zhang, H.S.; Zhang, Y.Z.; Zhang, K.P.; Sun, C.Y. Mushroom poisoning outbreaks—China, 2019. China CDC Wkly. 2020, 2, 19–24. [Google Scholar] [CrossRef]
- Li, H.J.; Zhang, H.S.; Zhang, Y.Z.; Zhou, J.; Yin, Y.; He, Q.; Jiang, S.F.; Ma, P.B.; Zhang, Y.T.; Wen, K.; et al. Mushroom poisoning outbreaks—China, 2020. China CDC Wkly. 2021, 3, 41–45. [Google Scholar] [CrossRef]
- Chen, Z.H.; Yang, Z.L.; Bau, T.; Li, T.H. Poisonous Mushrooms: Recognition and Poisoning Treatment; Science Press: Beijing, China, 2016. [Google Scholar]
- Walton, J.D. The Cyclic Peptide Toxins of Amanita and Other Poisonous Mushrooms; Springer International Publishing AG: Cham, Switzerland, 2018. [Google Scholar]
- Greenleaf, A.L.B.; Leslie, M.; Jiamachello, P.F.; Coulter, D.E. α-Amanitin-resistant D. melanogaster with an altered RNA polymerase II. Cell 1979, 18, 613–622. [Google Scholar] [CrossRef]
- Bushnell, D.A.; Patrick, C.; Kornberg, R.D. Structural basis of transcription: α-Amanitin-RNA polymerase II cocrystal at 2.8 Å resolution. Proc. Natl. Acad. Sci. USA 2002, 99, 1218–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kume, K.; Ikeda, M.; Miura, S.; Ito, K.; Sato, K.A.; Ohmori, Y.; Endo, F.; Katagiri, H.; Ishida, K.; Ito, C.; et al. α-Amanitin restrains cancer relapse from drug-tolerant cell subpopulations via TAF15. Sci. Rep. 2016, 6, 25895. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, X.; Han, C.; Wan, G.; Huang, X.; Ivan, C.; Jiang, D.; Rodriguez-Aguayo, C.; Lopez-Berestein, G.; Rao, P.H.; et al. TP53 loss creates therapeutic vulnerability in colorectal cancer. Nature 2015, 520, 697–701. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Hong, S.Y.; Sgambelluri, R.M.; Angelos, E.; Li, X.; Walton, J.D. Peptide macrocyclization catalyzed by a prolyl oligopeptidase involved in α-amanitin biosynthesis. Chem. Biol. 2014, 21, 1610–1617. [Google Scholar] [CrossRef] [Green Version]
- Sgambelluri, R.M.; Smith, M.O.; Walton, J.D. Versatility of prolyl oligopeptidase B in peptide macrocyclization. ACS. Synth. Biol. 2018, 7, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Hallen, H.E.; Luo, H.; Scott-Craig, J.S.; Walton, J.D. Gene family encoding the major toxins of lethal Amanita mushrooms. Proc. Natl. Acad. Sci. USA 2007, 104, 19097–19101. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Cai, Q.; Lüli, Y.J.; Li, X.; Sinha, R.; Hallen-Adams, H.E.; Yang, Z.L. The MSDIN family in amanitin-producing mushrooms and evolution of the prolyl oligopeptidase genes. IMA Fungus 2018, 9, 225–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lüli, Y.J.; Cai, Q.; Chen, Z.H.; Sun, H.; Zhu, X.T.; Li, X.; Yang, Z.L.; Luo, H. Genome of lethal Lepiota venenata and insights into the evolution of toxin-biosynthetic genes. BMC Genom. 2019, 20, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Hallen-Adams, H.E.; Scott-Craig, J.S.; Walton, J.D. Ribosomal biosynthesis of α-amanitin in Galerina marginata. Fungal Genet. Biol. 2012, 49, 123–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Q.; Tulloss, R.E.; Tang, L.P.; Tolgor, B.; Zhang, P.; Chen, Z.H.; Yang, Z.L. Multi-locus phylogeny of lethal amanitas: Implications for species diversity and historical biogeography. BMC Evol. Biol. 2014, 14, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Deng, W.Q.; Li, T.H.; Song, B.; Shen, Y.H. Illumina-based de novo transcriptome sequencing and analysis of Amanita exitialis basidiocarps. Gene 2013, 532, 63–71. [Google Scholar] [CrossRef]
- Pulman, J.A.; Childs, K.L.; Sgambelluri, R.M.; Walton, J.D. Expansion and diversification of the MSDIN family of cyclic peptide genes in the poisonous agarics Amanita phalloides and A. bisporigera. BMC Genom. 2016, 17, 1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, S.W.; Li, X.C.; Lüli, Y.J.; Li, X.; Chen, Z.H.; Yuan, P.C.; Yang, Z.L.; Li, G.H.; Luo, H. Novel cyclic peptides from lethal Amanita mushrooms through a genome-guided approach. J. Fungi 2021, 7, 204. [Google Scholar] [CrossRef]
- Cai, Q.; Cui, Y.Y.; Yang, Z.L. Lethal Amanita species in China. Mycologia 2016, 5, 993–1009. [Google Scholar]
- Kazutaka, K.; Daron, M.S. A simple method to control over-alignment in the MAFFT multiple sequence alignment program. Bioinformatics 2016, 32, 1933–1942. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A. MrModeltest v2.2; Program distributed by the author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Seppey, M.; Manni, M.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness. Methods Mol. Biol. 2019, 1962, 227–245. [Google Scholar] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Jia, G. Fastp: An ultra-fast all-in-one fastq preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Landmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map (SAM) format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Smyth, G.K.; Wei, S. The subread aligner: Fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 2013, 10, e108. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Emms, D.; Kelly, S. OrthoFinder: Solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 2015, 16, 157. [Google Scholar] [CrossRef] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Soderlund, C.; Bomhoff, M.; Nelson, W.M. SyMAP v3.4: A turnkey synteny system with application to plant genomes. Nucleic Acids Res. 2011, 39, e68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallen, H.E.; Watling, R.; Adams, G.C. Taxonomy and toxicity of Conocybe lactea and related species. Mycol. Res. 2003, 107, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Enjalbert, F.; Gallion, C.; Jehl, F.; Monteil, H.; Faulstich, H. Simultaneous assay for amatoxins and phallotoxins in Amanita phalloides by high-performance liquid chromatography. J. Chromatogr. 1992, 598, 227–236. [Google Scholar] [CrossRef]
- Cui, Y.Y.; Cai, Q.; Tang, L.P.; Liu, J.W.; Yang, Z.L. The family Amanitaceae: Molecular phylogeny, higher-rank taxonomy and the species in China. Fungal Divers. 2018, 91, 5–230. [Google Scholar] [CrossRef]
- Yang, Z.L. Atlas of the Chinese Species of Amanitaceae; Science Press: Beijing, China, 2015. [Google Scholar]
- Li, P.; Deng, W.Q.; Li, T.H. The molecular diversity of toxin gene families in lethal Amanita mushrooms. Toxicon 2014, 83, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.H.; Zou, J.P.; Deng, W.Q.; Li, T.H.; Jiang, Z.D. Molecular cloning and the expression pattern of AePOPB involved in the α-amanitin biosynthesis in Amanita exitialis fruiting bodies. Gene 2018, 662, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Lutz, C.; Simon, W.; Werner-Simon, S.; Pahl, A.; Muller, C. Total synthesis of α- and β-amanitin. Angew. Chem. Int. Ed. 2020, 59, 11390–11393. [Google Scholar] [CrossRef] [PubMed]
- Matinkhoo, K.; Pryyma, A.; Todorovic, M.; Patrick, B.O.; Perrin, D.M. Synthesis of the death-cap mushroom toxin α-amanitin. J. Am. Chem. Soc. 2018, 140, 6513–6517. [Google Scholar] [CrossRef]
- Siegert, M.J.; Knittel, C.H.; Sussmuth, R.D. A convergent total synthesis of the death cap toxin α-amanitin. Angew. Chem. Int. Ed. 2019, 59, 5500–5504. [Google Scholar] [CrossRef]
- May, J.P.; Perrin, D.M. Tryptathionine bridges in peptide synthesis. Biopolymers 2010, 88, 714–724. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides and Proteins | FPKM Values | Notes | |||
|---|---|---|---|---|---|
| Amanitin biosynthetic genes | MSDINATRLP | IWGIGCNP | CVGDDVTTLLTRGEALC | 0.24 | α-amanitin 3 copies |
| MSDINATRLA | IWGIGCNP | CVGDDVTALLTRGEALC | 0.15 | α-amanitin | |
| MSDINATRLA | IWGIGCNP | CVGDDVTALLTRGEALC | 2.70 | α-amanitin | |
| MSDINATRLA | IWGIGCDP | CVGDDVTALLTRGEALC | 0.78 | β-amanitin | |
| MSNINALRLP | GFGFIP | YASGDVDYTLTRGESLS | 357.26 | CylK1 | |
| MSDINATRFP | GKVNPP | YVGDDVDDIIIRGEKLC | 113.45 | CylK2 | |
| MSNINASRLP | IWAAFFRFP | CVGDEVDGILRSGESLC | 38.94 | ||
| MTDINATRLP | FFWILIPP | CVDDVDNTVHSGDNLC | 4.63 | ||
| MSDINATRFP | GKVFPP | YVGDDVDDIIIRGEK | 2.07 | ||
| MSNINATRFP | GKLFPP | YVGDDVDDIIIRGDKLC | 0.93 | ||
| MSDINASRLP | RLLVPRYP | CIDEDAEAILRSGECL | 0.90 | ||
| MTDINATRLP | ILFGFFLLP | CVDGVDNTLHSGENLC | 0.66 | ||
| MSDINSIHLP | GGYYQNT | FVGDDVEGILNRGERLC | 0.61 | ||
| MTDINATRLP | LNILPFHLPP | CVDDVDNTLHSGENLC | 0.29 | ||
| MTDINATRLP | FFIIFFIPP | CVDDVDNTLHSGENLC | 0.25 | 2 copies | |
| MTDINATRLP | FFIIFFIPP | CVDDVDNTLHSGENLC | 0.00 | ||
| MTDINATRLP | WIFFFYPP | CVDDVDNTLHSGENLC | 0.09 | ||
| MTDINATRLP | LIFLPPFIPP | CVDDVDNTLHSGENLC | 0.00 | 2 copies | |
| MTDINATRLP | LIFLPPFIPP | CVDDVDNTLHSGENLC | 0.00 | ||
| MTDINATRLP | WFFFFYPP | CVDDVDNTLHSGENLC | 0.00 | ||
| MTDINATRLP | FNILPLLLPP | CVDDVDNTLHSGENLC | 0.00 | ||
| POPB | 178.43 | ||||
| Housekeeping genes | POPA | 6.99 | |||
| rbp2 | 46.96 | ||||
| Cyclic Peptides | Elemental Composition | Theoretical (m/z) | Measured (m/z) | δ (ppm) |
|---|---|---|---|---|
| IWGIGCNP (α-amanitin) | C39H54N10O14S + H+ | 919.3614 | 919.3616 | 0.2175 |
| IWGIGCDP (β-amanitin) | C39H53N9O15S + H+ | 920.3455 | 920.3465 | 1.0865 |
| GFGFIP (CylK1) | C33H42N6O6 + H+ | 619.3239 | 619.3244 | 0.8073 |
| GKVNPP (CylK2) | C27H44N8O7 + H+ | 593.3406 | 593.3411 | 0.8427 |
| IWAAFFRFP | C61H77N13O9 + H+ | 1136.604 | N/A | N/A |
| GGYYQNT | C35H45N9O12 + H+ | 784.326 | N/A | N/A |
| RLLVPRYP | C48H78N14O9 + H+ | 995.6149 | N/A | N/A |
| LNILPFHLPP | C58H87N13O11 + H+ | 1142.6721 | N/A | N/A |
| LIFLPPFIPP | C62H90N10O10 + H+ | 1135.6914 | N/A | N/A |
| FFWILIPP | C57H75N9O8 + H+ | 1014.5811 | N/A | N/A |
| FFIIFFIPP | C64H83N9O9 + H+ | 1122.6387 | N/A | N/A |
| WIFFFYPP | C63H71N9O9 + H+ | 1098.5448 | N/A | N/A |
| GKVFPP | C32H47N7O6 + H+ | 626.3661 | N/A | N/A |
| GKLFPP | C33H49N7O6 + H+ | 640.3817 | N/A | N/A |
| ILFGFFLLP | C58H81N9O9 + H+ | 1048.623 | N/A | N/A |
| WFFFFYPP | C66H69N9O9 + H+ | 1132.5291 | N/A | N/A |
| FNILPLLLPP | C58H91N11O11 + H+ | 1118.6972 | N/A | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lüli, Y.; Zhou, S.; Li, X.; Chen, Z.; Yang, Z.; Luo, H. Differential Expression of Amanitin Biosynthetic Genes and Novel Cyclic Peptides in Amanita molliuscula. J. Fungi 2021, 7, 384. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7050384
Lüli Y, Zhou S, Li X, Chen Z, Yang Z, Luo H. Differential Expression of Amanitin Biosynthetic Genes and Novel Cyclic Peptides in Amanita molliuscula. Journal of Fungi. 2021; 7(5):384. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7050384
Chicago/Turabian StyleLüli, Yunjiao, Shengwen Zhou, Xuan Li, Zuohong Chen, Zhuliang Yang, and Hong Luo. 2021. "Differential Expression of Amanitin Biosynthetic Genes and Novel Cyclic Peptides in Amanita molliuscula" Journal of Fungi 7, no. 5: 384. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7050384