
Histidine Kinase Sln1 and cAMP/PKA Signaling Pathways Antagonistically Regulate Sporisorium scitamineum Mating and Virulence via Transcription Factor Prf1

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Construction of Strains
2.3. Nucleic Acid Related Manipulation
2.4. Sporidia Staining and Epifluorescence Microscopy
2.5. SsHog1 Phosphorylation Assays
2.6. Yeast Two-Hybrid (Y2H) Assays
2.7. Assessment of Pathogenicity and Relative Fungal Biomass
2.8. Statistical Analysis
3. Results
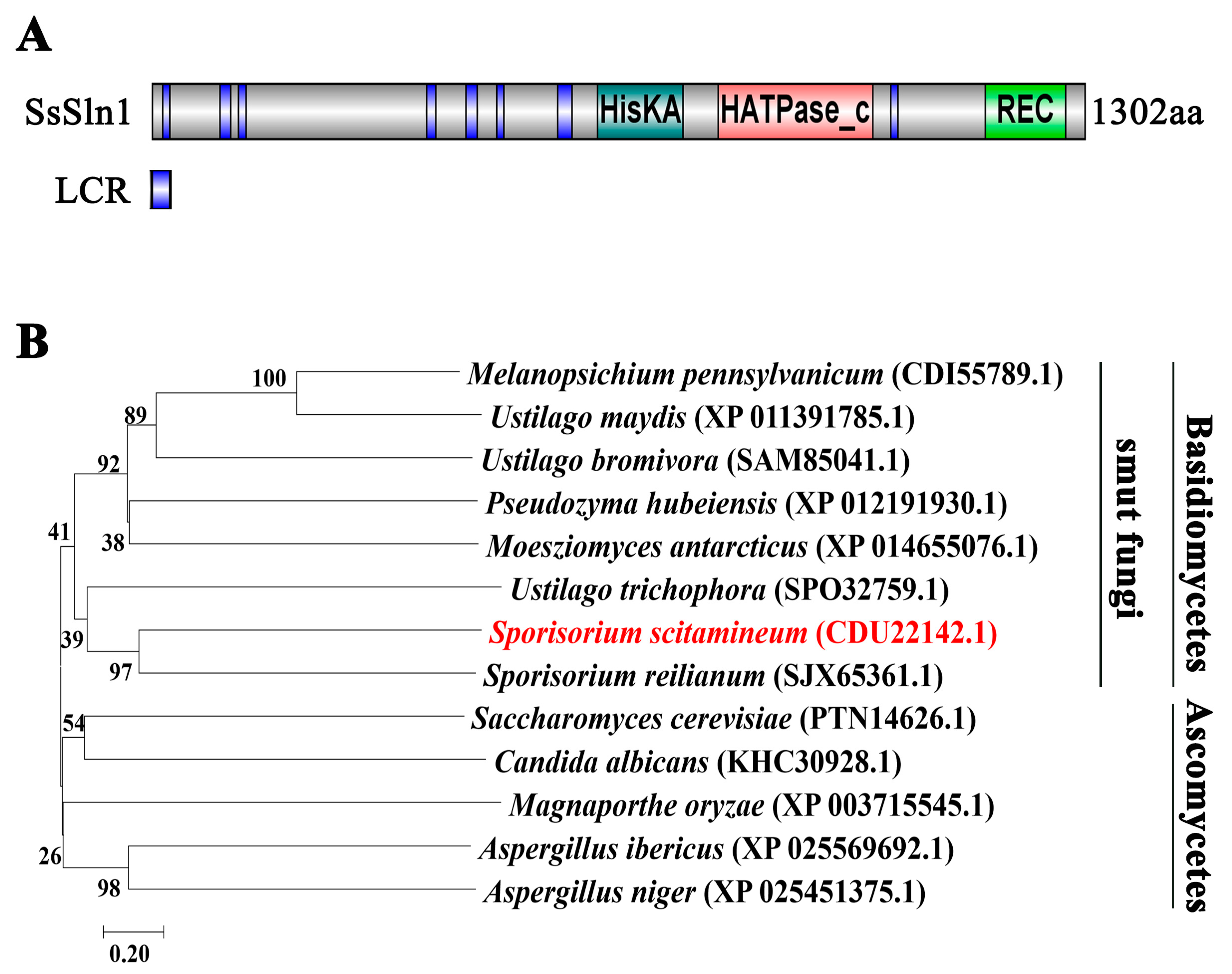
3.1. Identification of SsSln1 Protein in S. scitamineum
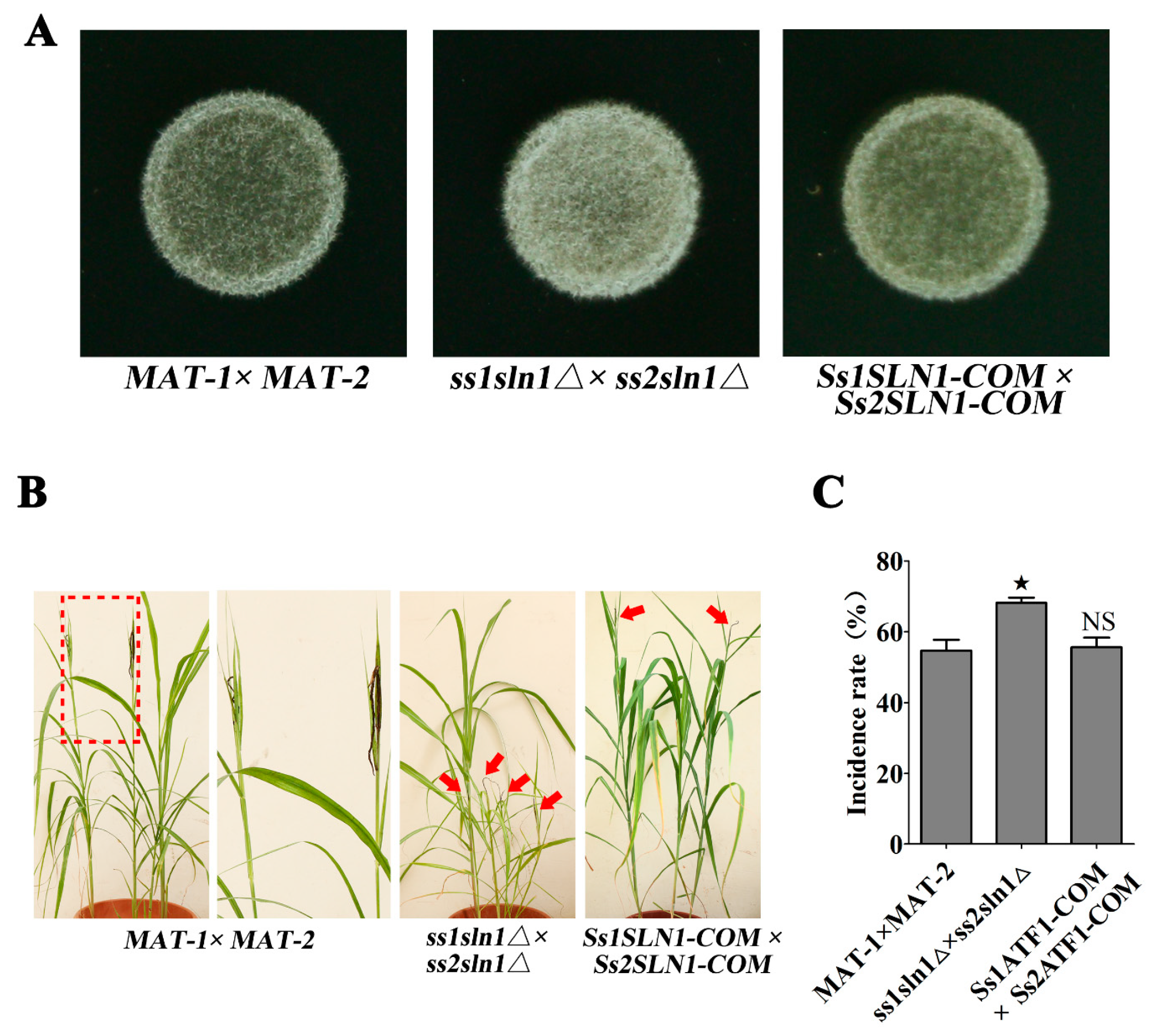
3.2. Deletion of SsSLN1 Gene Promotes Mating and Pathogenicity of S. scitamineum
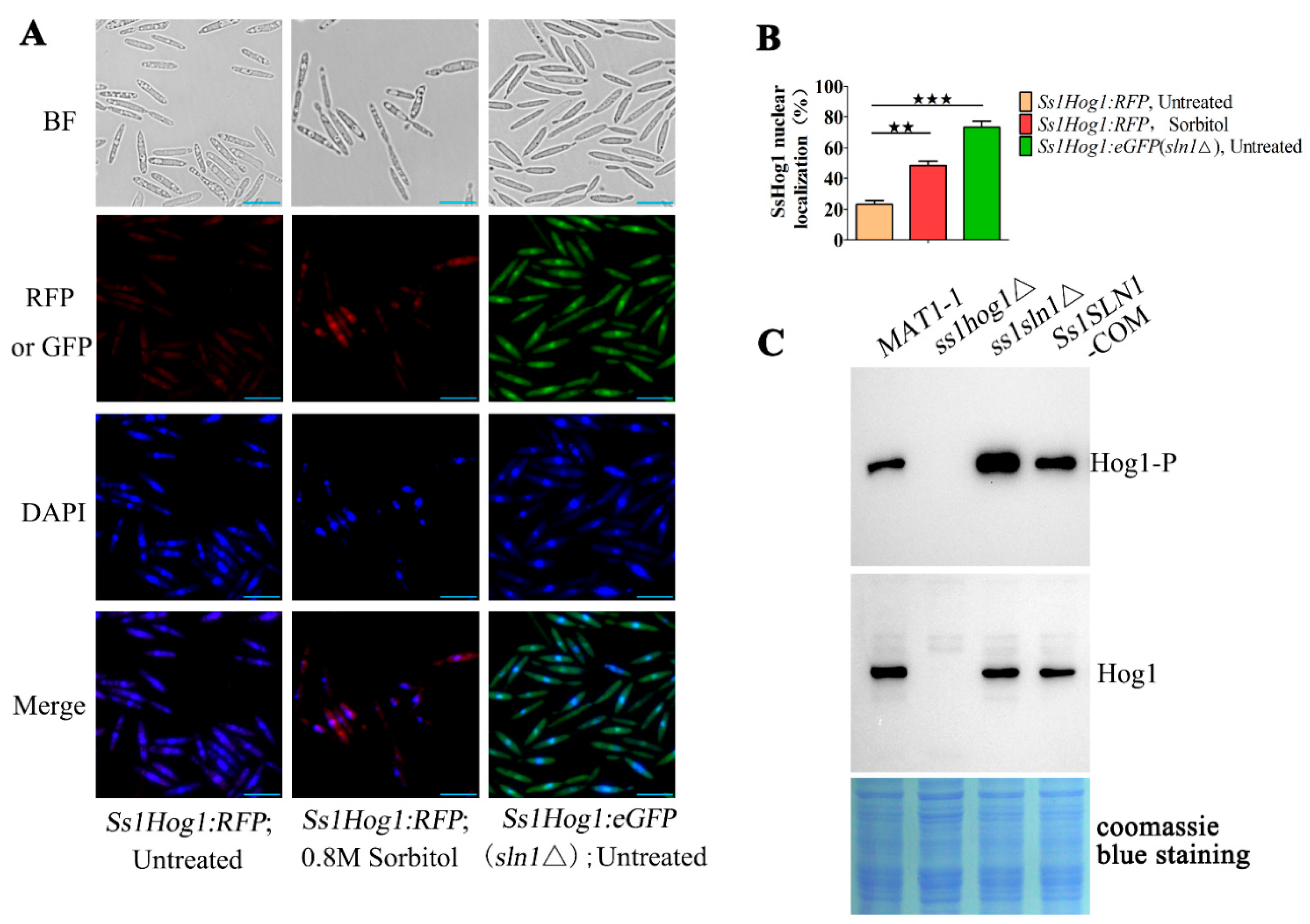
3.3. SsSln1 Negatively Regulates Phosphorylation and Nuclear Localization of SsHog1
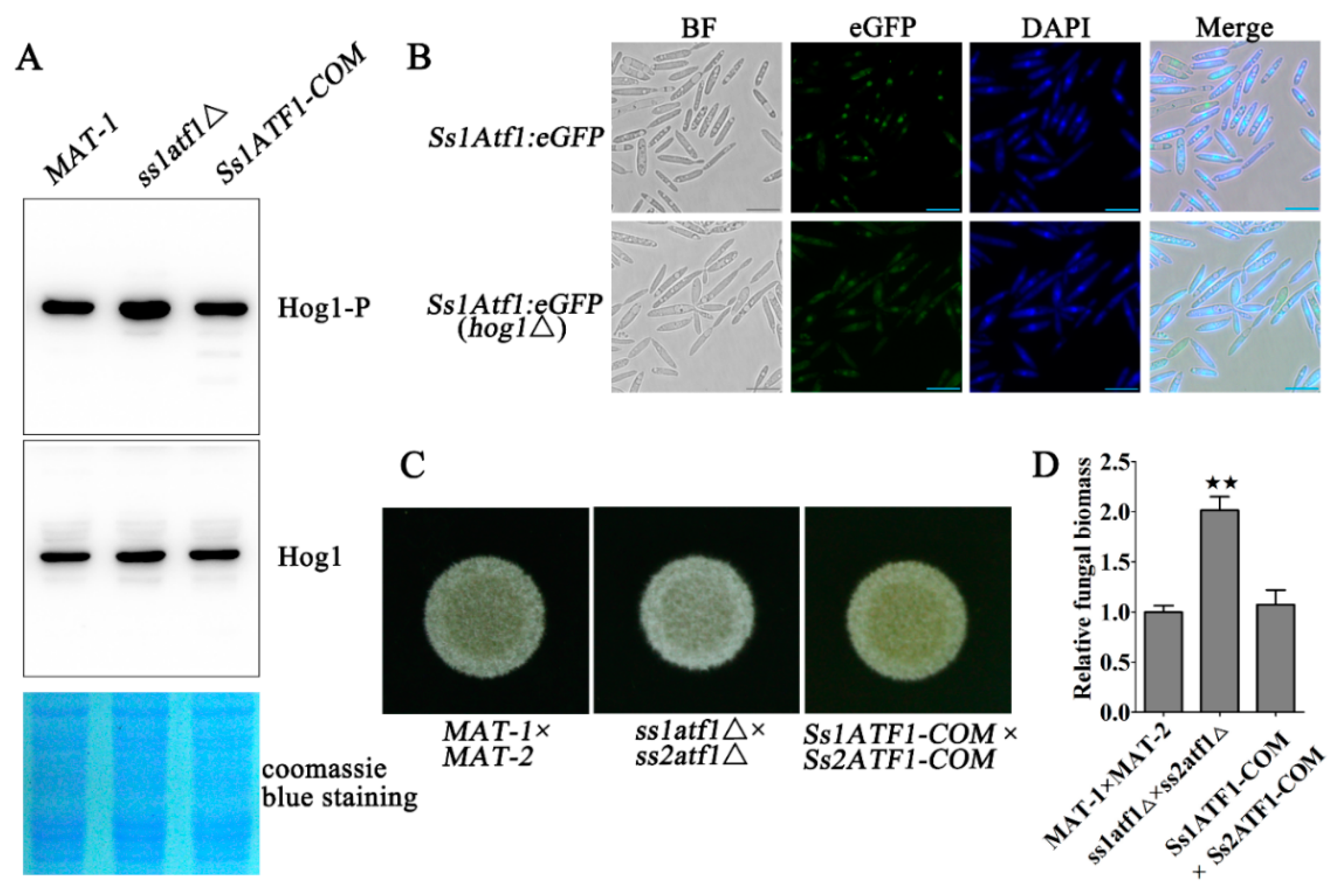
3.4. Phosphorylation of SsHog1 Is Necessary for Mating and Virulence of S. scitamineum
3.5. Phosphorylation Level of SsHog1 Positively Mediates Mating and Virulence of S. scitamineum
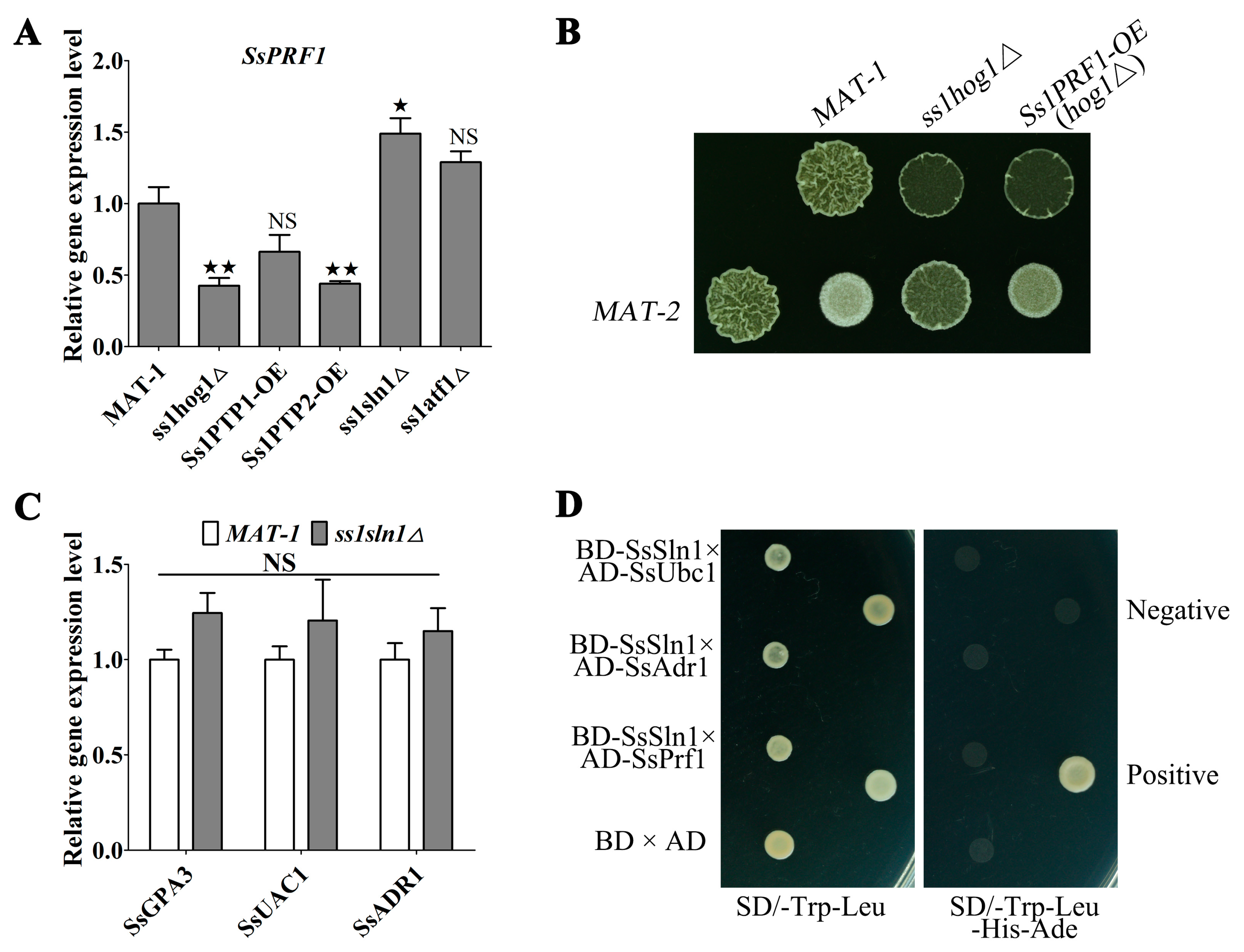
3.6. SsSln1 Negatively Regulates the Transcription of SsPRF1 by SsHog1 Phosphorylation, but Not through the cAMP/PKA Signaling Pathway
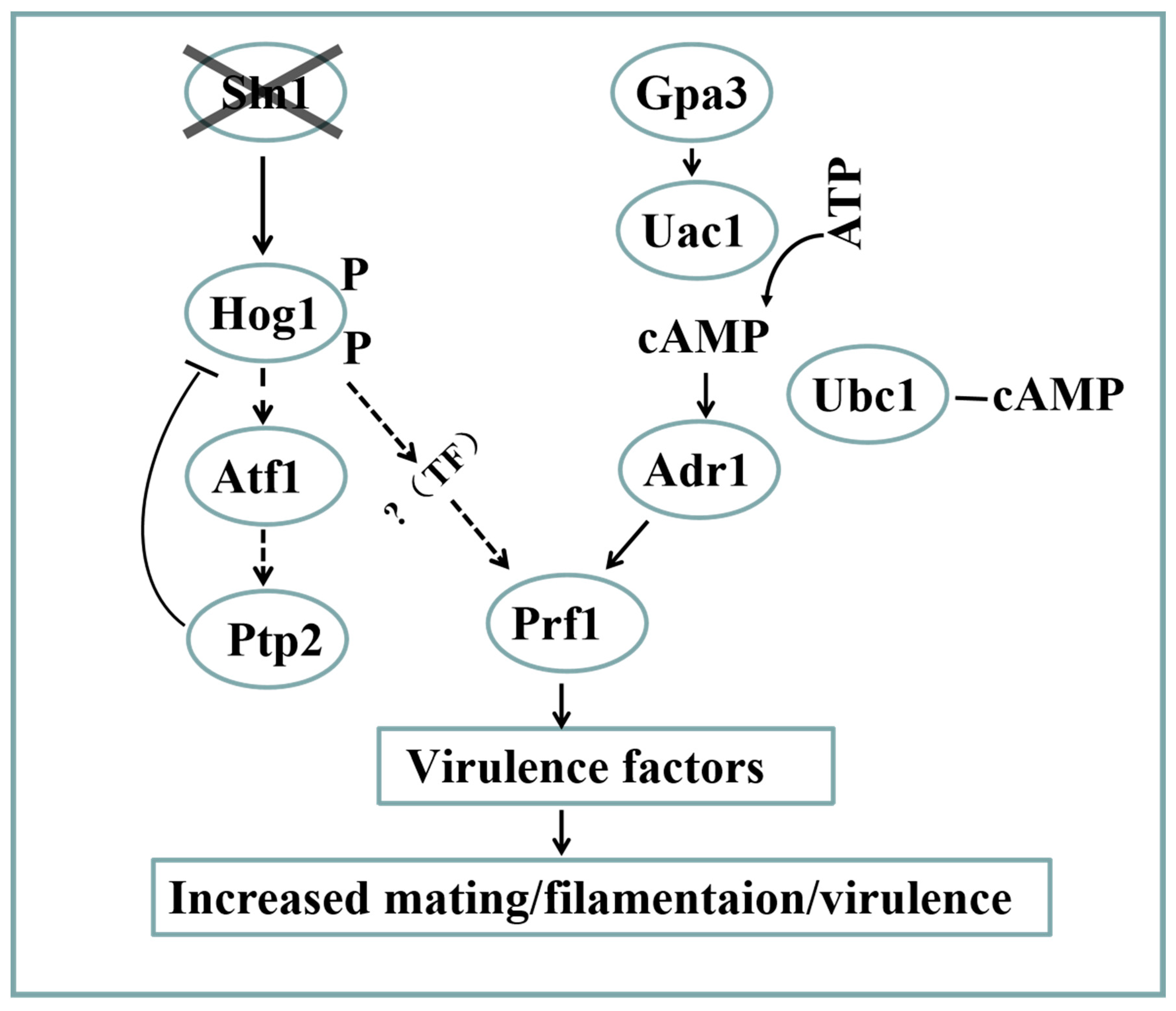
3.7. Cross-Talk between the SsSln1 and cAMP/PKA Pathways Antagonistically Regulates Mating and Virulence by SsPRF1 Transcription
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taniguti, L.M.; Schaker, P.D.; Benevenuto, J.; Peters, L.P.; Carvalho, G.; Palhares, A.; Quecine, M.C.; Nunes, F.R.; Kmit, M.C.; Wai, A.; et al. Complete Genome Sequence of Sporisorium scitamineum and Biotrophic Interaction Transcriptome with Sugarcane. PLoS ONE 2015, 10, e0129318. [Google Scholar] [CrossRef]
- Yan, M.; Zhu, G.; Lin, S.; Xian, X.; Chang, C.; Xi, P.; Shen, W.; Huang, W.; Cai, E.; Jiang, Z.; et al. The mating-type locus b of the sugarcane smut Sporisorium scitamineum is essential for mating, filamentous growth and pathogenicity. Fungal Genet. Biol. 2016, 86, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Saito, H. Histidine phosphorylation and two-component signaling in eukaryotic cells. Chem. Rev. 2001, 101, 2497–2509. [Google Scholar] [CrossRef]
- Catlett, N.L.; Yoder, O.C.; Turgeon, B.G. Whole-genome analysis of two-component signal transduction genes in fungal pathogens. Eukaryot. Cell 2003, 2, 1151–1161. [Google Scholar] [CrossRef] [Green Version]
- Motoyama, T.; Ochiai, N.; Morita, M.; Iida, Y.; Usami, R.; Kudo, T. Involvement of putative response regulator genes of the rice blast fungus Magnaporthe oryzae in osmotic stress response, fungicide action, and pathogenicity. Curr. Genet. 2008, 54, 185–195. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, K.; Zhang, X.; Song, W.; Zhao, Q.; Dong, Y.; Guo, M.; Zheng, X.; Zhang, Z. A two-component histidine kinase, MoSLN1, is required for cell wall integrity and pathogenicity of the rice blast fungus, Magnaporthe oryzae. Curr. Genet. 2010, 56, 517–528. [Google Scholar] [CrossRef]
- Posas, F.; Wurgler-Murphy, S.M.; Maeda, T.; Witten, E.A.; Thai, T.C.; Saito, H. Yeast HOG1 MAP kinase cascade is regulated by a multistep phosphorelay mechanism in the SLN1-YPD1-SSK1 “two-component” osmosensor. Cell 1996, 86, 865–875. [Google Scholar] [CrossRef] [Green Version]
- Hohmann, S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef] [Green Version]
- Alex, L.A.; Borkovich, K.A.; Simon, M.I. Hyphal development in Neurospora crassa: Involvement of a two-component histidine kinase. Proc. Natl. Acad. Sci. USA 1996, 93, 3416–3421. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, M.M.; Enderlin, C.S.; Selitrennikoff, C.P. The osmotic-1 locus of Neurospora crassa encodes a putative histidine kinase similar to osmosensors of bacteria and yeast. Curr. Microbiol. 1997, 34, 340–347. [Google Scholar] [CrossRef]
- Furukawa, K.; Katsuno, Y.; Urao, T.; Yabe, T.; Yamada-Okabe, T.; Yamada-Okabe, H.; Yamagata, Y.; Abe, K.; Nakajima, T. Isolation and functional analysis of a gene, tcsB, encoding a transmembrane hybrid-type histidine kinase from Aspergillus nidulans. Appl. Environ. Microbiol. 2002, 68, 5304–5310. [Google Scholar] [CrossRef] [Green Version]
- Fassler, J.S.; West, A.H. Histidine phosphotransfer proteins in fungal two-component signal transduction pathways. Eukaryot. Cell 2013, 12, 1052–1060. [Google Scholar] [CrossRef] [Green Version]
- Day, A.M.; Smith, D.A.; Ikeh, M.A.; Haider, M.; Herrero-de-Dios, C.M.; Brown, A.J.; Morgan, B.A.; Erwig, L.P.; MacCallum, D.M.; Quinn, J. Blocking two-component signalling enhances Candida albicans virulence and reveals adaptive mechanisms that counteract sustained SAPK activation. PLoS Pathog. 2017, 13, e1006131. [Google Scholar] [CrossRef] [PubMed]
- Yamada-Okabe, T.; Mio, T.; Ono, N.; Kashima, Y.; Matsui, M.; Arisawa, M.; Yamada-Okabe, H. Roles of three histidine kinase genes in hyphal development and virulence of the pathogenic fungus Candida albicans. J. Bacteriol. 1999, 181, 7243–7247. [Google Scholar] [CrossRef] [Green Version]
- Ryder, L.S.; Dagdas, Y.F.; Kershaw, M.J.; Venkataraman, C.; Madzvamuse, A.; Yan, X.; Cruz-Mireles, N.; Soanes, D.M.; Oses-Ruiz, M.; Styles, V.; et al. A sensor kinase controls turgor-driven plant infection by the rice blast fungus. Nature 2019, 574, 423–427. [Google Scholar] [CrossRef] [PubMed]
- Posas, F.; Saito, H. Osmotic activation of the HOG MAPK pathway via Ste11p MAPKKK: Scaffold role of Pbs2p MAPKK. Science 1997, 276, 1702–1705. [Google Scholar] [CrossRef] [PubMed]
- Posas, F.; Saito, H. Activation of the yeast SSK2 MAP kinase kinase kinase by the SSK1 two-component response regulator. EMBO J. 1998, 17, 1385–1394. [Google Scholar] [CrossRef] [Green Version]
- Tatebayashi, K.; Yamamoto, K.; Tanaka, K.; Tomida, T.; Maruoka, T.; Kasukawa, E.; Saito, H. Adaptor functions of Cdc42, Ste50, and Sho1 in the yeast osmoregulatory HOG MAPK pathway. EMBO J. 2006, 25, 3033–3044. [Google Scholar] [CrossRef]
- Maeda, T.; Takekawa, M.; Saito, H. Activation of yeast PBS2 MAPKK by MAPKKKs or by binding of an SH3-containing osmosensor. Science 1995, 269, 554–558. [Google Scholar] [CrossRef]
- Van Wuytswinkel, O.; Reiser, V.; Siderius, M.; Kelders, M.C.; Ammerer, G.; Ruis, H.; Mager, W.H. Response of Saccharomyces cerevisiae to severe osmotic stress: Evidence for a novel activation mechanism of the HOG MAP kinase pathway. Mol. Microbiol. 2000, 37, 382–397. [Google Scholar] [CrossRef]
- O’Rourke, S.M.; Herskowitz, I. Unique and redundant roles for HOG MAPK pathway components as revealed by whole-genome expression analysis. Mol. Biol. Cell 2004, 15, 532–542. [Google Scholar] [CrossRef]
- Horie, T.; Tatebayashi, K.; Yamada, R.; Saito, H. Phosphorylated Ssk1 prevents unphosphorylated Ssk1 from activating the Ssk2 mitogen-activated protein kinase kinase kinase in the yeast high-osmolarity glycerol osmoregulatory pathway. Mol. Cell. Biol. 2008, 28, 5172–5183. [Google Scholar] [CrossRef] [Green Version]
- Zarrinpar, A.; Bhattacharyya, R.P.; Nittler, M.P.; Lim, W.A. Sho1 and Pbs2 act as coscaffolds linking components in the yeast high osmolarity MAP kinase pathway. Mol. Cell 2004, 14, 825–832. [Google Scholar] [CrossRef]
- Hohmann, S. Control of high osmolarity signalling in the yeast Saccharomyces cerevisiae. FEBS Lett. 2009, 583, 4025–4029. [Google Scholar] [CrossRef] [Green Version]
- Ferrigno, P.; Posas, F.; Koepp, D.; Saito, H.; Silver, P.A. Regulated nucleo/cytoplasmic exchange of HOG1 MAPK requires the importin beta homologs NMD5 and XPO1. EMBO J. 1998, 17, 5606–5614. [Google Scholar] [CrossRef]
- Reiser, V.; Ruis, H.; Ammerer, G. Kinase activity-dependent nuclear export opposes stress-induced nuclear accumulation and retention of Hog1 mitogen-activated protein kinase in the budding yeast Saccharomyces cerevisiae. Mol. Biol. Cell 1999, 10, 1147–1161. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Mondal, A.K. Evidence that C-terminal non-kinase domain of Pbs2p has a role in high osmolarity-induced nuclear localization of Hog1p. Biochem. Biophys. Res. Commun. 2005, 328, 906–913. [Google Scholar] [CrossRef]
- Hersen, P.; McClean, M.N.; Mahadevan, L.; Ramanathan, S. Signal processing by the HOG MAP kinase pathway. Proc. Natl. Acad. Sci. USA 2008, 105, 7165–7170. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhou, Q.; Guo, Z.; Liu, P.; Shen, L.; Chai, N.; Qian, B.; Cai, Y.; Wang, W.; Yin, Z.; et al. A self-balancing circuit centered on MoOsm1 kinase governs adaptive responses to host-derived ROS in Magnaporthe oryzae. eLife 2020, 9, e61605. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.T.; Byun, H.J.; Jung, K.W.; Hong, J.; Cheong, E.; Bahn, Y.S. Distinct and redundant roles of protein tyrosine phosphatases Ptp1 and Ptp2 in governing the differentiation and pathogenicity of Cryptococcus neoformans. Eukaryot. Cell 2014, 13, 796–812. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Harashima, T.; Heitman, J. Signal transduction cascades regulating pseudohyphal differentiation of Saccharomyces cerevisiae. Curr. Opin. Microbiol. 2000, 3, 567–572. [Google Scholar] [CrossRef]
- Thevelein, J.M.; Cauwenberg, L.; Colombo, S.; De Winde, J.H.; Donation, M.; Dumortier, F.; Kraakman, L.; Lemaire, K.; Ma, P.; Nauwelaers, D.; et al. Nutrient-induced signal transduction through the protein kinase A pathway and its role in the control of metabolism, stress resistance, and growth in yeast. Enzym. Microb. Technol. 2000, 26, 819–825. [Google Scholar] [CrossRef]
- Hogan, D.A.; Sundstrom, P. The Ras/cAMP/PKA signaling pathway and virulence in Candida albicans. Future Microbiol. 2009, 4, 1263–1270. [Google Scholar] [CrossRef]
- Choi, J.; Jung, W.H.; Kronstad, J.W. The cAMP/protein kinase A signaling pathway in pathogenic basidiomycete fungi: Connections with iron homeostasis. J. Microbiol. 2015, 53, 579–587. [Google Scholar] [CrossRef] [Green Version]
- Marroquin-Guzman, M.; Wilson, R.A. GATA-Dependent Glutaminolysis Drives Appressorium Formation in Magnaporthe oryzae by Suppressing TOR Inhibition of cAMP/PKA Signaling. PLoS Pathog. 2015, 11, e1004851. [Google Scholar] [CrossRef] [PubMed]
- Caza, M.; Kronstad, J.W. The cAMP/Protein Kinase a Pathway Regulates Virulence and Adaptation to Host Conditions in Cryptococcus neoformans. Front. Cell. Infect. Microbiol. 2019, 9, 212. [Google Scholar] [CrossRef]
- Martínez-Soto, D.; Ortiz-Castellanos, L.; Robledo-Briones, M.; León-Ramírez, C.G. Molecular Mechanisms Involved in the Multicellular Growth of Ustilaginomycetes. Microorganisms 2020, 8, 1072. [Google Scholar] [CrossRef]
- Chang, C.; Cai, E.; Deng, Y.Z.; Mei, D.; Qiu, S.; Chen, B.; Zhang, L.H.; Jiang, Z. cAMP/PKA signalling pathway regulates redox homeostasis essential for Sporisorium scitamineum mating/filamentation and virulence. Environ. Microbiol. 2019, 21, 959–971. [Google Scholar] [CrossRef]
- Zhu, G.; Deng, Y.; Cai, E.; Yan, M.; Cui, G.; Wang, Z.; Zou, C.; Zhang, B.; Xi, P.; Chang, C.; et al. Identification and Functional Analysis of the Pheromone Response Factor Gene of Sporisorium scitamineum. Front. Microbiol. 2019, 10, 2115. [Google Scholar] [CrossRef]
- Garrido, E.; Pérez-Martín, J. The crk1 gene encodes an Ime2-related protein that is required for morphogenesis in the plant pathogen Ustilago maydis. Mol. Microbiol. 2003, 47, 729–743. [Google Scholar] [CrossRef]
- Cai, E.; Li, L.; Deng, Y.; Sun, S.; Jia, H.; Wu, R.; Zhang, L.; Jiang, Z.; Chang, C. MAP kinase Hog1 mediates a cytochrome P450 oxidoreductase to promote the Sporisorium scitamineum cell survival under oxidative stress. Environ. Microbiol. 2021, 23, 3306–3317. [Google Scholar] [CrossRef]
- Cai, E.; Mei, D.; Zhang, X.; Sun, X.; Li, L.; Wu, R.; Deng, Y.; Jiang, Z.; Chang, C. A gene knockout method based on protoplast transformation with two PCR fragments in Sporisorium scitamineum. Mycosystema 2020, 39, 2314–2317. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sun, S.; Deng, Y.; Cai, E.; Yan, M.; Li, L.; Chen, B.; Chang, C.; Jiang, Z. The Farnesyltransferase β-Subunit Ram1 Regulates Sporisorium scitamineum Mating, Pathogenicity and Cell Wall Integrity. Front. Microbiol. 2019, 10, 976. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderone, R.A.; Fonzi, W.A. Virulence factors of Candida albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef]
- Lipa, P.; Janczarek, M. Phosphorylation systems in symbiotic nitrogen-fixing bacteria and their role in bacterial adaptation to various environmental stresses. PeerJ 2020, 8, e8466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, T.; Wurgler-Murphy, S.M.; Saito, H. A two-component system that regulates an osmosensing MAP kinase cascade in yeast. Nature 1994, 369, 242–245. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Monge, R.; Navarro-García, F.; Molero, G.; Diez-Orejas, R.; Gustin, M.; Pla, J.; Sánchez, M.; Nombela, C. Role of the mitogen-activated protein kinase Hog1p in morphogenesis and virulence of Candida albicans. J. Bacteriol. 1999, 181, 3058–3068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahn, Y.S.; Kojima, K.; Cox, G.M.; Heitman, J. Specialization of the HOG pathway and its impact on differentiation and virulence of Cryptococcus neoformans. Mol. Biol. Cell 2005, 16, 2285–2300. [Google Scholar] [CrossRef] [Green Version]
- Ko, Y.J.; Yu, Y.M.; Kim, G.B.; Lee, G.W.; Maeng, P.J.; Kim, S.; Floyd, A.; Heitman, J.; Bahn, Y.S. Remodeling of global transcription patterns of Cryptococcus neoformans genes mediated by the stress-activated HOG signaling pathways. Eukaryot. Cell 2009, 8, 1197–1217. [Google Scholar] [CrossRef] [Green Version]
- Winkelströter, L.K.; Bom, V.L.; de Castro, P.A.; Ramalho, L.N.; Goldman, M.H.; Brown, N.A.; Rajendran, R.; Ramage, G.; Bovier, E.; Dos Reis, T.F.; et al. High osmolarity glycerol response PtcB phosphatase is important for Aspergillus fumigatus virulence. Mol. Microbiol. 2015, 96, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Bruder Nascimento, A.C.; Dos Reis, T.F.; de Castro, P.A.; Hori, J.I.; Bom, V.L.; de Assis, L.J.; Ramalho, L.N.; Rocha, M.C.; Malavazi, I.; Brown, N.A.; et al. Mitogen activated protein kinases SakA(HOG1) and MpkC collaborate for Aspergillus fumigatus virulence. Mol. Microbiol. 2016, 100, 841–859. [Google Scholar] [CrossRef] [Green Version]
- Dixon, K.P.; Xu, J.R.; Smirnoff, N.; Talbot, N.J. Independent signaling pathways regulate cellular turgor during hyperosmotic stress and appressorium-mediated plant infection by Magnaporthe grisea. Plant Cell 1999, 11, 2045–2058. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Guo, W.; Chen, Y.; Dong, S.; Zhang, X.; Zhang, H.; Song, W.; Wang, W.; Wang, Q.; Lv, R.; et al. The basic leucine zipper transcription factor Moatf1 mediates oxidative stress responses and is necessary for full virulence of the rice blast fungus Magnaporthe oryzae. Mol. Plant Microbe Interact. 2010, 23, 1053–1068. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.Z.; Zhang, B.; Chang, C.; Wang, Y.; Lu, S.; Sun, S.; Zhang, X.; Chen, B.; Jiang, Z. The MAP Kinase SsKpp2 Is Required for Mating/Filamentation in Sporisorium scitamineum. Front. Microbiol. 2018, 9, 2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Accession Number for Protein | Resistance Marker and Strain Background | Reference or Source |
|---|---|---|---|
| MAT-1 | a1, b1 | [2] | |
| MAT-2 | a2, b2 | [2] | |
| ss1sln1Δ | CDU22142.1 | Hygromycin, MAT-1 | This study |
| ss2sln1Δ | CDU22142.1 | Hygromycin, MAT-2 | This study |
| Ss1SLN1-COM | CDU22142.1 | Zeocin, ss1sln1Δ | This study |
| Ss2SLN1-COM | CDU22142.1 | Zeocin, ss2sln1Δ | This study |
| ss1atf1Δ | CDU21933.1 | Hygromycin, MAT-1 | This study |
| ss2atf1Δ | CDU21933.1 | Hygromycin, MAT-2 | This study |
| Ss1ATF1-COM | CDU21933.1 | Zeocin, ss1atf1Δ | This study |
| Ss2ATF1-COM | CDU21933.1 | Zeocin, ss2atf1Δ | This study |
| ss1hog1Δ | CDU23149.1 | Hygromycin, MAT-1 | [41] |
| Ss1PTP1-OE | CDU22358.1 | Zeocin, MAT-1 | This study |
| Ss2PTP1-OE | CDU22358.1 | Zeocin, MAT-2 | This study |
| Ss1PTP2-OE | CDU23562.1 | Zeocin, MAT-1 | This study |
| Ss2PTP2-OE | CDU23562.1 | Zeocin, MAT-2 | This study |
| Ss1PRF1-OE(hog1Δ) | CDU22680.1 | Zeocin, ss1hog1Δ | This study |
| Ss1Hog1:RFP | CDU23149.1 | Hygromycin, MAT-1 | This study |
| Ss1Hog1:eGFP(sln1Δ) | CDU23149.1 | Zeocin, ss1sln1Δ | This study |
| Ss1Atf1:eGFP | CDU21933.1 | Zeocin, MAT-1 | This study |
| Ss1Atf1:eGFP(hog1Δ) | CDU21933.1 | Zeocin, ss1hog1Δ | This study |
| ss1sln1Δgpa3Δ | CDU22378.1 | Zeocin, ss1sln1Δ | This study |
| ss1sln1Δuac1Δ | CDU25762.1 | Zeocin, ss1sln1Δ | This study |
| ss1sln1Δadr1Δ | CDU22361.1 | Zeocin, ss1sln1Δ | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, E.; Sun, S.; Deng, Y.; Huang, P.; Sun, X.; Wang, Y.; Chang, C.; Jiang, Z. Histidine Kinase Sln1 and cAMP/PKA Signaling Pathways Antagonistically Regulate Sporisorium scitamineum Mating and Virulence via Transcription Factor Prf1. J. Fungi 2021, 7, 610. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080610
Cai E, Sun S, Deng Y, Huang P, Sun X, Wang Y, Chang C, Jiang Z. Histidine Kinase Sln1 and cAMP/PKA Signaling Pathways Antagonistically Regulate Sporisorium scitamineum Mating and Virulence via Transcription Factor Prf1. Journal of Fungi. 2021; 7(8):610. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080610
Chicago/Turabian StyleCai, Enping, Shuquan Sun, Yizhen Deng, Peishen Huang, Xian Sun, Yuting Wang, Changqing Chang, and Zide Jiang. 2021. "Histidine Kinase Sln1 and cAMP/PKA Signaling Pathways Antagonistically Regulate Sporisorium scitamineum Mating and Virulence via Transcription Factor Prf1" Journal of Fungi 7, no. 8: 610. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080610