
H2O2 Induces Major Phosphorylation Changes in Critical Regulators of Signal Transduction, Gene Expression, Metabolism and Developmental Networks in Aspergillus nidulans


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media, and Growth Conditions
2.2. Protein Extraction
2.3. Protein Digestion and Phosphopeptide Enrichment
2.4. Liquid Chromatography Coupled to Tandem Mass Spectrometry (LC-MS/MS) Analysis
2.5. Database Searching
2.6. Phosphopeptide and Protein Identification Criteria
2.7. Bioinformatics Analyses
3. Results and Discussion
3.1. About One Fifth of A. nidulans Predicted Proteins Are Phosphorylated during Growth
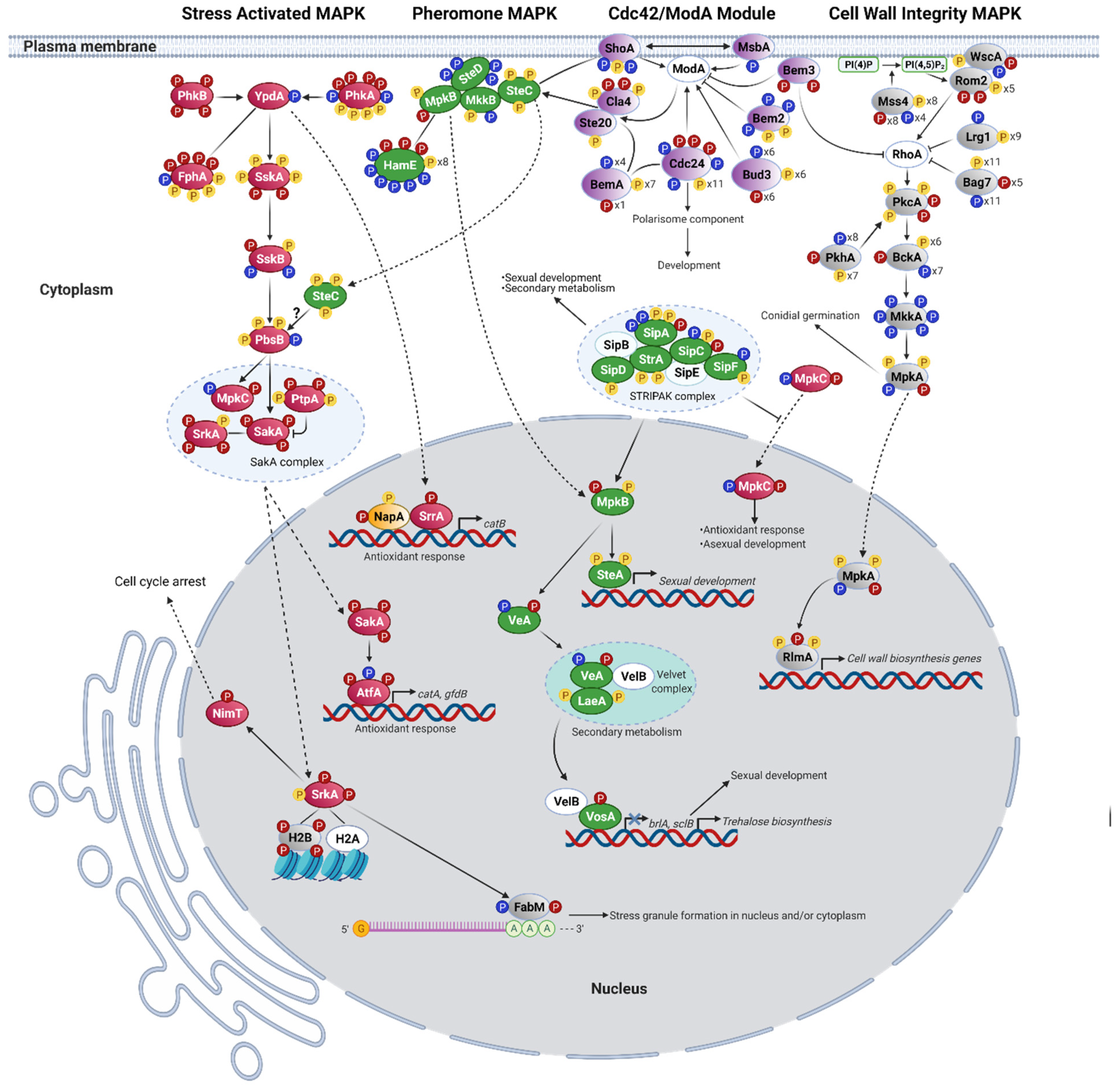
3.2. H2O2 Induces the Activation of the SakA/MpkC Stress MAPK Pathway
3.3. H2O2 in the Activation of the MpkB Pheromone MAPK Pathway
3.4. H2O2 in the Regulation of RhoA and the Activation of the MpkA Cell Wall Integrity Pathway (CWI)
3.5. Msb2-Sho1 and the Cdc42 Module as Integrators of MAPK Signaling in Response to H2O2
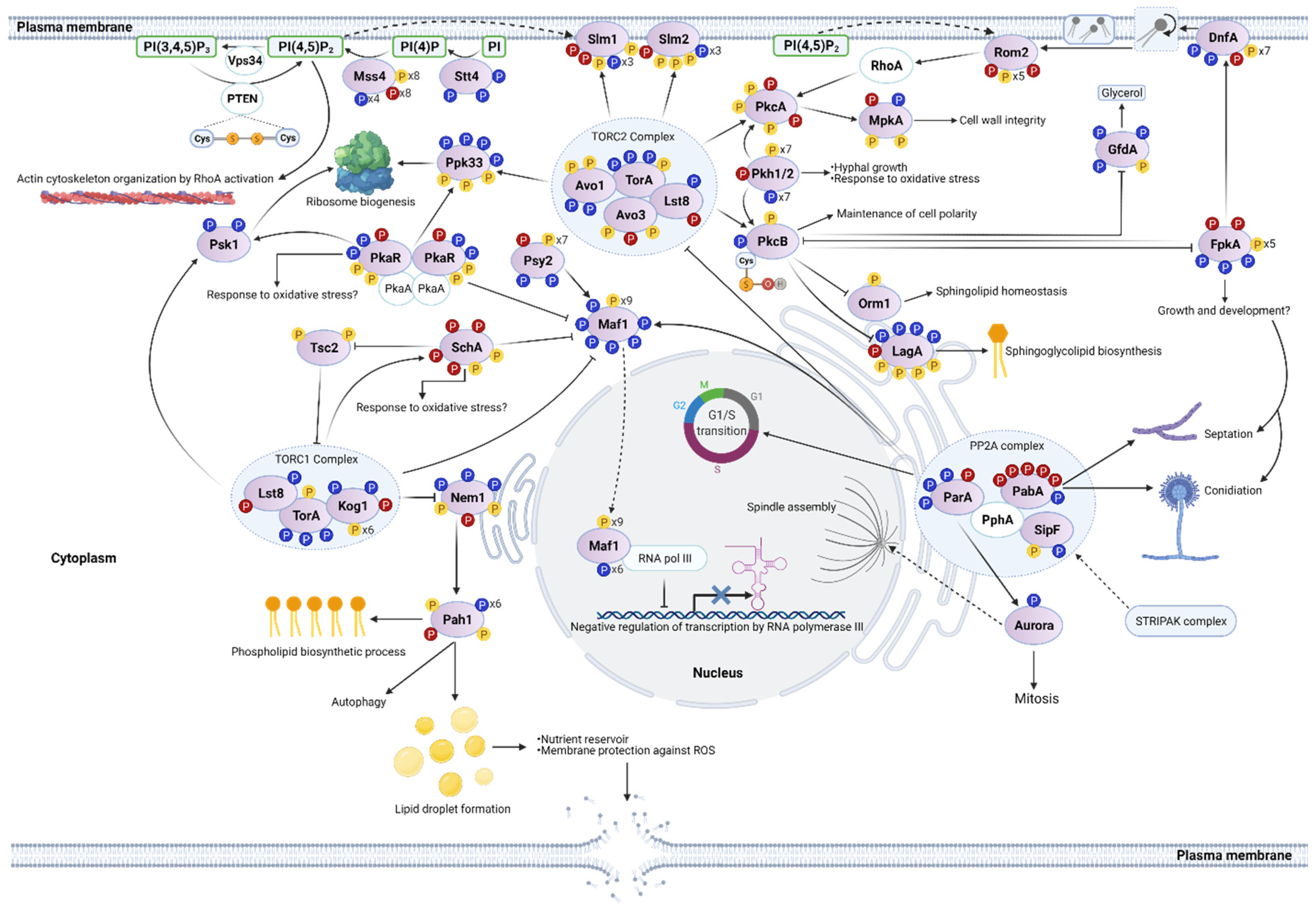
3.6. H2O2 in Phosphoinositide and TOR Signaling
3.7. The Phosphorylation of Multiple Proteins Involved in Transcriptional Regulation Is Affected by H2O2
3.8. H2O2 in the Regulation of Nitrogen Assimilation and the Reprogramming of Metabolism
3.9. The Phosphorylation of Proteosome-Mediated Proteolysis Components Is Affected by H2O2
3.10. H2O2 in the Regulation of Gluconeogenesis and the Pentose Phosphate Pathway (PPP)
3.11. H2O2 in the Phosphorylation of Proteins Involved in ROS Metabolism
4. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguirre, J.; Lambeth, J.D. Nox enzymes from fungus to fly to fish and what they tell us about Nox function in mammals. Free Radic. Biol. Med. 2010, 49, 1342–1353. [Google Scholar] [CrossRef] [Green Version]
- Rhee, S.G. Redox signaling: Hydrogen peroxide as intracellular messenger. Exp. Mol. Med. 1999, 31, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Hansberg, W.; Salas-Lizana, R.; Dominguez, L. Fungal catalases: Function, phylogenetic origin and structure. Arch. Biochem. Biophys. 2012, 525, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Harman, D. Aging—A Theory Based on Free-Radical and Radiation-Chemistry. J. Gerontol. 1956, 11, 298–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demple, B. Study of redox-regulated transcription factors in prokaryotes. Methods 1997, 11, 267–278. [Google Scholar] [CrossRef]
- Rosner, J.L.; Storz, G. Regulation of bacterial responses to oxidative stress. Curr. Top. Cell Regul. 1997, 35, 163–177. [Google Scholar] [CrossRef]
- Shirasu, K.; Dixon, R.A.; Lamb, C. Signal transduction in plant immunity. Curr. Opin. Immunol. 1996, 8, 3–7. [Google Scholar] [CrossRef]
- Sundaresan, M.; Yu, Z.X.; Ferrans, V.J.; Irani, K.; Finkel, T. Requirement for Generation of H2O2 for Platelet-Derived Growth-Factor Signal-Transduction. Science 1995, 270, 296–299. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, J.; Rios-Momberg, M.; Hewitt, D.; Hansberg, W. Reactive oxygen species and development in microbial eukaryotes. Trends Microbiol. 2005, 13, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Hansberg, W.; Aguirre, J. Hyperoxidant states cause microbial cell differentiation by cell isolation from dioxygen. J. Theor. Biol. 1990, 142, 201–221. [Google Scholar] [CrossRef]
- Hansberg, W.; Degroot, H.; Sies, H. Reactive Oxygen Species Associated with Cell-Differentiation in Neurospora crassa. Free Radic. Bio. Med. 1993, 14, 287–293. [Google Scholar] [CrossRef]
- Aguirre, J. Oxidación de Glutamina Sintetasa, Glutamato Deshidrogenasa Biosintética y Otras Proteínas en Neurospora crassa: Relación con la Conidiación. Ph.D. Thesis, Universidad Nacional Autónoma de México, México City, Mexico, 1988. [Google Scholar]
- Aguirre, J.; Hansberg, W. Oxidation of Neurospora crassa glutamine synthetase. J. Bacteriol. 1986, 166, 1040–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguirre, J.; Rodriguez, R.; Hansberg, W. Oxidation of Neurospora crassa NADP-specific glutamate-dehydrogenase by activated oxygen species. J. Bacteriol. 1989, 171, 6243–6250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansberg, W.; Aguirre, J.; Ríos-Momberg, M.; Rangel, P.; Peraza, L.; de Oca, Y.M.; Cano-Domínguez, N. Cell differentiation as a response to oxidative stress. In Stress in Yeast and Filamentous Fungi; Avery, S., Strattford, M., Valerius, O., van West, P., Eds.; Academic Press: London, UK, 2008; pp. 235–257. [Google Scholar]
- Cano-Dominguez, N.; Alvarez-Delfin, K.; Hansberg, W.; Aguirre, J. NADPH oxidases NOX-1 and NOX-2 require the regulatory subunit NOR-1 to control cell differentiation and growth in Neurospora crassa. Eukaryot. Cell 2008, 7, 1352–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cano-Dominguez, N.; Bowman, B.; Peraza-Reyes, L.; Aguirre, J. Neurospora crassa NADPH Oxidase NOX-1 Is Localized in the Vacuolar System and the Plasma Membrane. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Lara-Ortiz, T.; Riveros-Rosas, H.; Aguirre, J. Reactive oxygen species generated by microbial NADPH oxidase NoxA regulate sexual development in Aspergillus nidulans. Mol. Microbiol. 2003, 50, 1241–1255. [Google Scholar] [CrossRef] [Green Version]
- Malagnac, F.; Lalucque, H.; Lepere, G.; Silar, P. Two NADPH oxidase isoforms are required for sexual reproduction and ascospore germination in the filamentous fungus Podospora anserina. Fungal Genet. Biol. 2004, 41, 982–997. [Google Scholar] [CrossRef] [PubMed]
- Dirschnabel, D.E.; Nowrousian, M.; Cano-Dominguez, N.; Aguirre, J.; Teichert, I.; Kuck, U. New Insights Into the Roles of NADPH Oxidases in Sexual Development and Ascospore Germination in Sordaria macrospora. Genetics 2014, 196, 729–744. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Bazan, V.; Pardo, J.P.; Aguirre, J. DnmA and FisA Mediate Mitochondria and Peroxisome Fission, and Regulate Mitochondrial Function, ROS Production and Development in Aspergillus nidulans. Front. Microbiol. 2020, 11, 837. [Google Scholar] [CrossRef]
- Jaimes-Arroyo, R.; Lara-Rojas, F.; Bayram, O.; Valerius, O.; Braus, G.H.; Aguirre, J. The SrkA Kinase Is Part of the SakA Mitogen-Activated Protein Kinase Interactome and Regulates Stress Responses and Development in Aspergillus nidulans. Eukaryot. Cell 2015, 14, 495–510. [Google Scholar] [CrossRef] [Green Version]
- Garrido-Bazan, V.; Jaimes-Arroyo, R.; Sanchez, O.; Lara-Rojas, F.; Aguirre, J. SakA and MpkC Stress MAPKs Show Opposite and Common Functions during Stress Responses and Development in Aspergillus nidulans. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, L.; Aguirre, J. Multiple catalase genes are differentially regulated in Aspergillus nidulans. J. Bacteriol. 2001, 183, 1434–1440. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, L.; Sanchez, O.; Shiozaki, K.; Aguirre, J. SakA MAP kinase is involved in stress signal transduction, sexual development and spore viability in Aspergillus nidulans. Mol. Microbiol. 2002, 45, 1153–1163. [Google Scholar] [CrossRef]
- Mendoza-Martínez, A.E.; Cano-Domínguez, N.; Aguirre, J. Yap1 homologs mediate more than the redox regulation of the antioxidant response in filamentous fungi. Fungal Biol. 2019, 1–10. [Google Scholar] [CrossRef]
- Mendoza-Martinez, A.E.; Lara-Rojas, F.; Sanchez, O.; Aguirre, J. NapA Mediates a Redox Regulation of the Antioxidant Response, Carbon Utilization and Development in Aspergillus nidulans. Front. Microbiol. 2017, 8, 516. [Google Scholar] [CrossRef]
- Vargas-Perez, I.; Sanchez, O.; Kawasaki, L.; Georgellis, D.; Aguirre, J. Response regulators SrrA and SskA are central components of a phosphorelay system involved in stress signal transduction and asexual sporulation in Aspergillus nidulans. Eukaryot. Cell 2007, 6, 1570–1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Autreaux, B.; Toledano, M.B. ROS as signalling molecules: Mechanisms that generate specificity in ROS homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef]
- Ostman, A.; Frijhoff, J.; Sandin, A.; Bohmer, F.D. Regulation of protein tyrosine phosphatases by reversible oxidation. J. Biochem. 2011, 150, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonks, N.K. Protein tyrosine phosphatases: From genes, to function, to disease. Nat. Rev. Mol. Cell Biol. 2006, 7, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Byrne, D.P.; Shrestha, S.; Galler, M.; Cao, M.; Daly, L.A.; Campbell, A.E.; Eyers, C.E.; Veal, E.A.; Kannan, N.; Eyers, P.A. Aurora A regulation by reversible cysteine oxidation reveals evolutionarily conserved redox control of Ser/Thr protein kinase activity. Sci. Signal. 2020, 13. [Google Scholar] [CrossRef]
- Lara-Rojas, F.; Sanchez, O.; Kawasaki, L.; Aguirre, J. Aspergillus nidulans transcription factor AtfA interacts with the MAPK SakA to regulate general stress responses, development and spore functions. Mol. Microbiol. 2011, 80, 436–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conibear, A.C. Deciphering protein post-translational modifications using chemical biology tools. Nat. Rev. Chem. 2020, 4, 674–695. [Google Scholar] [CrossRef]
- Needham, E.J.; Parker, B.L.; Burykin, T.; James, D.E.; Humphrey, S.J. Illuminating the dark phosphoproteome. Sci. Signal. 2019, 12. [Google Scholar] [CrossRef] [Green Version]
- Chelius, C.L.; Ribeiro, L.F.C.; Huso, W.; Kumar, J.; Lincoln, S.; Tran, B.; Goo, Y.A.; Srivastava, R.; Harris, S.D.; Marten, M.R. Phosphoproteomic and transcriptomic analyses reveal multiple functions for Aspergillus nidulans MpkA independent of cell wall stress. Fungal Genet. Biol. 2019, 125, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, L.F.C.; Chelius, C.; Boppidi, K.R.; Naik, N.S.; Hossain, S.; Ramsey, J.J.J.; Kumar, J.; Ribeiro, L.F.; Ostermeier, M.; Tran, B.; et al. Comprehensive Analysis of Aspergillus nidulans PKA Phosphorylome Identifies a Novel Mode of CreA Regulation. MBIO 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Ramsubramaniam, N.; Harris, S.D.; Marten, M.R. The phosphoproteome of Aspergillus nidulans reveals functional association with cellular processes involved in morphology and secretion. Proteomics 2014, 14, 2454–2459. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.W.; Käfer, E. Improved protocols for Aspergillus minimal medium: Trace element and minimal medium stock solution. Fungal Genet. Newsl. 2001, 48, 20–21. [Google Scholar] [CrossRef] [Green Version]
- Jami, M.S.; Barreiro, C.; Garcia-Estrada, C.; Martin, J.F. Proteome analysis of the penicillin producer Penicillium chrysogenum: Characterization of protein changes during the industrial strain improvement. Mol. Cell Proteom. 2010, 9, 1182–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, A.; Nesvizhskii, A.I.; Kolker, E.; Aebersold, R. Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 2002, 74, 5383–5392. [Google Scholar] [CrossRef]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef] [PubMed]
- Zuniga-Leon, E.; Carrasco-Navarro, U.; Fierro, F. NeVOmics: An Enrichment Tool for Gene Ontology and Functional Network Analysis and Visualization of Data from OMICs Technologies. Genes 2018, 9, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. Data, information, knowledge and principle: Back to metabolism in KEGG. Nucleic Acids Res. 2014, 42, D199–D205. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.; Grant, C.E.; Noble, W.S.; Bailey, T.L. MoMo: Discovery of statistically significant post-translational modification motifs. Bioinformatics 2019, 35, 2774–2782. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Sicheritz-Ponten, T.; Gupta, R.; Gammeltoft, S.; Brunak, S. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar] [CrossRef]
- Horn, H.; Schoof, E.M.; Kim, J.; Robin, X.; Miller, M.L.; Diella, F.; Palma, A.; Cesareni, G.; Jensen, L.J.; Linding, R. KinomeXplorer: An integrated platform for kinome biology studies. Nat. Methods 2014, 11, 603–604. [Google Scholar] [CrossRef]
- Skromne, I.; Sanchez, O.; Aguirre, J. Starvation Stress Modulates the Expression of the Aspergillus nidulans brlA Regulatory Gene. Microbiol-Sgm 1995, 141, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Navarro, R.E.; Stringer, M.A.; Hansberg, W.; Timberlake, W.E.; Aguirre, J. catA, a new Aspergillus nidulans gene encoding a developmentally regulated catalase. Curr. Genet. 1996, 29, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, L. Las Catalasas y la Respuesta Antioxidante Durante el Crecimiento y la Diferenciacion en Aspergillus nidulans; Universidad Nacional Autónoma de México: México city, Mexico, 1998. [Google Scholar]
- Forman, H.J.; Bernardo, A.; Davies, K.J.A. What is the concentration of hydrogen peroxide in blood and plasma? Arch. Biochem. Biophys. 2016, 603, 48–53. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Cerqueira, G.C.; Arnaud, M.B.; Inglis, D.O.; Skrzypek, M.S.; Binkley, G.; Simison, M.; Miyasato, S.R.; Binkley, J.; Orvis, J.; Shah, P.; et al. The Aspergillus Genome Database: Multispecies curation and incorporation of RNA-Seq data to improve structural gene annotations. Nucleic Acids Res. 2014, 42, D705–D710. [Google Scholar] [CrossRef] [Green Version]
- Gene Ontology, C. The Gene Ontology resource: Enriching a GOld mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef] [PubMed]
- Mohanta, T.K.; Mohanta, N.; Parida, P.; Panda, S.K.; Ponpandian, L.N.; Bae, H. Genome-Wide Identification of Mitogen-Activated Protein Kinase Gene Family across Fungal Lineage Shows Presence of Novel and Diverse Activation Loop Motifs. PLoS ONE 2016, 11, e0149861. [Google Scholar] [CrossRef] [Green Version]
- Bilsland, E.; Molin, C.; Swaminathan, S.; Ramne, A.; Sunnerhagen, P. Rck1 and Rck2 MAPKAP kinases and the HOG pathway are required for oxidative stress resistance. Mol. Microbiol. 2004, 53, 1743–1756. [Google Scholar] [CrossRef]
- Buck, V.; Quinn, J.; Soto Pino, T.; Martin, H.; Saldanha, J.; Makino, K.; Morgan, B.A.; Millar, J.B. Peroxide sensors for the fission yeast stress-activated mitogen-activated protein kinase pathway. Mol. Biol. Cell 2001, 12, 407–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.Z.; Armant, O.; Fischer, R. Fungi use the SakA (HogA) pathway for phytochrome-dependent light signalling. Nat. Microbiol. 2016, 1. [Google Scholar] [CrossRef]
- Nguyen, A.; Chen, P.; Cai, H. Role of CaMKII in hydrogen peroxide activation of ERK1/2, p38 MAPK, HSP27 and actin reorganization in endothelial cells. FEBS Lett. 2004, 572, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Aviles, S.; Lambea, E.; Moldon, A.; Grande, M.; Fajardo, A.; Rodriguez-Gabriel, M.A.; Hidalgo, E.; Aligue, R. Activation of Srk1 by the mitogen-activated protein kinase Sty1/Spc1 precedes its dissociation from the kinase and signals its degradation. Mol. Biol. Cell 2008, 19, 1670–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, S.H.; Cheung, W.L.; Hsu, J.Y.; Diaz, R.L.; Smith, M.M.; Allis, C.D. Sterile 20 kinase phosphorylates histone H2B at serine 10 during hydrogen peroxide-induced apoptosis in S. cerevisiae. Cell 2005, 120, 25–36. [Google Scholar] [CrossRef] [Green Version]
- Marhoul, J.F.; Adams, T.H. Aspergillus fabM encodes an essential product that is related to poly(A)-binding proteins and activates development when overexpressed. Genetics 1996, 144, 1463–1470. [Google Scholar] [CrossRef]
- Soukup, A.A.; Fischer, G.J.; Luo, J.; Keller, N.P. The Aspergillus nidulans Pbp1 homolog is required for normal sexual development and secondary metabolism. Fungal Genet. Biol. 2017, 100, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Alberti, S.; Dormann, D. Liquid-Liquid Phase Separation in Disease. Annu. Rev. Genet. 2019, 53, 171–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangus, D.A.; Smith, M.M.; McSweeney, J.M.; Jacobson, A. Identification of factors regulating poly(A) tail synthesis and maturation. Mol. Cell. Biol. 2004, 24, 4196–4206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.S.; Kato, M.; Wu, X.; Litsios, A.; Sutter, B.M.; Wang, Y.; Hsu, C.H.; Wood, N.E.; Lemoff, A.; Mirzaei, H.; et al. Yeast Ataxin-2 Forms an Intracellular Condensate Required for the Inhibition of TORC1 Signaling during Respiratory Growth. Cell 2019, 177, 697–710.e17. [Google Scholar] [CrossRef] [Green Version]
- Holmes, K.J.; Klass, D.M.; Guiney, E.L.; Cyert, M.S. Whi3, an S. cerevisiae RNA-Binding Protein, Is a Component of Stress Granules That Regulates Levels of Its Target mRNAs. PLoS ONE 2013, 8, e84060. [Google Scholar] [CrossRef] [Green Version]
- Gerbich, T.M.; McLaughlin, G.A.; Cassidy, K.; Gerber, S.; Adalsteinsson, D.; Gladfelter, A.S. Phosphoregulation provides specificity to biomolecular condensates in the cell cycle and cell polarity. J. Cell Biol. 2020, 219. [Google Scholar] [CrossRef] [PubMed]
- Kato, M.; Yang, Y.S.; Sutter, B.M.; Wang, Y.; McKnight, S.L.; Tu, B.P. Redox State Controls Phase Separation of the Yeast Ataxin-2 Protein via Reversible Oxidation of Its Methionine-Rich Low-Complexity Domain. Cell 2019, 177, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Salat-Canela, C.; Paulo, E.; Sanchez-Mir, L.; Carmona, M.; Ayte, J.; Oliva, B.; Hidalgo, E. Deciphering the role of the signal- and Sty1 kinase-dependent phosphorylation of the stress-responsive transcription factor Atf1 on gene activation. J. Biol. Chem. 2017, 292, 13635–13644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santo, P.D.; Blanchard, B.; Hoffman, C.S. The Schizosaccharomyces pombe pyp1 protein tyrosine phosphatase negatively regulates nutrient monitoring pathways. J. Cell Sci. 1996, 109 Pt 7, 1919–1925. [Google Scholar] [CrossRef]
- Di, Y.; Holmes, E.J.; Butt, A.; Dawson, K.; Mironov, A.; Kotiadis, V.N.; Gourlay, C.W.; Jones, N.; Wilkinson, C.R. H(2)O(2) stress-specific regulation of S. pombe MAPK Sty1 by mitochondrial protein phosphatase Ptc4. EMBO J. 2012, 31, 563–575. [Google Scholar] [CrossRef] [Green Version]
- Visintin, R.; Craig, K.; Hwang, E.S.; Prinz, S.; Tyers, M.; Amon, A. The phosphatase Cdc14 triggers mitotic exit by reversal of Cdk-dependent phosphorylation. Mol. Cell 1998, 2, 709–718. [Google Scholar] [CrossRef]
- de Assis, L.J.; Ries, L.N.; Savoldi, M.; Dinamarco, T.M.; Goldman, G.H.; Brown, N.A. Multiple Phosphatases Regulate Carbon Source-Dependent Germination and Primary Metabolism in Aspergillus nidulans. G3 2015, 5, 857–872. [Google Scholar] [CrossRef] [Green Version]
- Klionsky, D.J. Monitoring autophagy in yeast: The Pho8Delta60 assay. Methods Mol. Biol. 2007, 390, 363–371. [Google Scholar] [CrossRef]
- Schinke, J.; Gulko, M.K.; Christmann, M.; Valerius, O.; Stumpf, S.K.; Stirz, M.; Braus, G.H. The DenA/DEN1 Interacting Phosphatase DipA Controls Septa Positioning and Phosphorylation-Dependent Stability of Cytoplasmatic DenA/DEN1 during Fungal Development. PLoS Genet. 2016, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salogiannis, J.; Christensen, J.R.; Songster, L.D.; Aguilar-Maldonado, A.; Shukla, N.; Reck-Peterson, S.L. PxdA interacts with the DipA phosphatase to regulate peroxisome hitchhiking on early endosomes. Mol. Biol. Cell 2021, 32, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y. Regulation of the cell cycle by protein phosphatase 2A in Saccharomyces cerevisiae. Microbiol. Mol. Biol. R. 2006, 70, 440. [Google Scholar] [CrossRef] [Green Version]
- Bayram, O.; Bayram, O.S.; Ahmed, Y.L.; Maruyama, J.I.; Valerius, O.; Rizzoli, S.O.; Ficner, R.; Irniger, S.; Braus, G.H. The Aspergillus nidulans MAPK Module AnSte11-Ste50-Ste7-Fus3 Controls Development and Secondary Metabolism. PLoS Genet. 2012, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frawley, D.; Karahoda, B.; Sarikaya Bayram, O.; Bayram, O. The HamE scaffold positively regulates MpkB phosphorylation to promote development and secondary metabolism in Aspergillus nidulans. Sci. Rep. 2018, 8, 16588. [Google Scholar] [CrossRef] [Green Version]
- Frawley, D.; Stroe, M.C.; Oakley, B.R.; Heinekamp, T.; Strassburger, M.; Fleming, A.B.; Brakhage, A.A.; Bayram, O. The Pheromone Module SteC-MkkB-MpkB-SteD-HamE Regulates Development, Stress Responses and Secondary Metabolism in Aspergillus fumigatus. Front. Microbiol. 2020, 11, 811. [Google Scholar] [CrossRef]
- Seo, J.A.; Han, K.H.; Yu, J.H. The gprA and gprB genes encode putative G protein-coupled receptors required for self-fertilization in Aspergillus nidulans. Mol. Microbiol. 2004, 53, 1611–1623. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Keller, N.P. Genetic involvement of a cAMP-dependent protein kinase in a g protein signaling pathway regulating morphological and chemical transitions in Aspergillus nidulans. Genetics 2001, 157, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Wieser, J.; Adams, T.H. The Aspergillus flbA RGS domain protein antagonizes G protein signaling to block proliferation and allow development. EMBO J. 1996, 15, 5184–5190. [Google Scholar] [CrossRef] [PubMed]
- Lafon, A.; Seo, J.A.; Han, K.H.; Yu, J.H.; d’Enfert, C. The heterotrimeric G-protein GanB(alpha)-SfaD(beta)-GpgA(gamma) is a carbon source sensor involved in early cAMP-dependent germination in Aspergillus nidulans. Genetics 2005, 171, 71–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gartner, A.; Nasmyth, K.; Ammerer, G. Signal transduction in Saccharomyces cerevisiae requires tyrosine and threonine phosphorylation of FUS3 and KSS1. Genes Dev. 1992, 6, 1280–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuck, U.; Radchenko, D.; Teichert, I. STRIPAK, a highly conserved signaling complex, controls multiple eukaryotic cellular and developmental processes and is linked with human diseases. Biol. Chem. 2019, 400, 1005–1022. [Google Scholar] [CrossRef]
- Elramli, N.; Karahoda, B.; Sarikaya-Bayram, O.; Frawley, D.; Ulas, M.; Oakley, C.E.; Oakley, B.R.; Seiler, S.; Bayram, O. Assembly of a heptameric STRIPAK complex is required for coordination of light-dependent multicellular fungal development with secondary metabolism in Aspergillus nidulans. PLoS Genet. 2019, 15. [Google Scholar] [CrossRef] [Green Version]
- Marquez-Fernandez, O.; Trigos, A.; Ramos-Balderas, J.L.; Viniegra-Gonzalez, G.; Deising, H.B.; Aguirre, J. Phosphopantetheinyl transferase CfwA/NpgA is required for Aspergillus nidulans secondary metabolism and asexual development. Eukaryot. Cell 2007, 6, 710–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soid-Raggi, G.; Sanchez, O.; Aguirre, J. TmpA, a member of a novel family of putative membrane flavoproteins, regulates asexual development in Aspergillus nidulans. Mol. Microbiol. 2006, 59, 854–869. [Google Scholar] [CrossRef]
- Soid-Raggi, G.; Sanchez, O.; Ramos-Balderas, J.L.; Aguirre, J. The Adenylate-Forming Enzymes AfeA and TmpB Are Involved in Aspergillus nidulans Self-Communication during Asexual Development. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Bok, J.W.; Keller, N.P. LaeA, a regulator of secondary metabolism in Aspergillus spp. Eukaryot. Cell 2004, 3, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarikaya-Bayram, O.; Palmer, J.M.; Keller, N.; Braus, G.H.; Bayram, O. One Juliet and four Romeos: VeA and its methyltransferases. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rauscher, S.; Pacher, S.; Hedtke, M.; Kniemeyer, O.; Fischer, R. A phosphorylation code of the Aspergillus nidulans global regulator VelvetA (VeA) determines specific functions. Mol. Microbiol. 2016, 99, 909–924. [Google Scholar] [CrossRef] [PubMed]
- Baidya, S.; Duran, R.M.; Lohmar, J.M.; Harris-Coward, P.Y.; Cary, J.W.; Hong, S.Y.; Roze, L.V.; Linz, J.E.; Calvo, A.M. VeA Is Associated with the Response to Oxidative Stress in the Aflatoxin Producer Aspergillus flavus. Eukaryot. Cell 2014, 13, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Reverberi, M.; Zjalic, S.; Ricelli, A.; Punelli, F.; Camera, E.; Fabbri, C.; Picardo, M.; Fanelli, C.; Fabbri, A.A. Modulation of antioxidant defense in Aspergillus parasiticus is involved in aflatoxin biosynthesis: A role for the ApyapA gene. Eukaryot. Cell 2008, 7, 988–1000. [Google Scholar] [CrossRef] [Green Version]
- Roze, L.V.; Chanda, A.; Wee, J.; Awad, D.; Linz, J.E. Stress-related transcription factor AtfB integrates secondary metabolism with oxidative stress response in aspergilli. J. Biol. Chem. 2011, 286, 35137–35148. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.W.; Yu, J.H.; Kelkar, H.S.; Fernandes, M.; Nesbitt, T.C.; Keller, N.P.; Adams, T.H.; Leonard, T.J. Twenty-five coregulated transcripts define a sterigmatocystin gene cluster in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 1996, 93, 1418–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, M.R.; Nielsen, J.B.; Klitgaard, A.; Petersen, L.M.; Zachariasen, M.; Hansen, T.J.; Blicher, L.H.; Gotfredsen, C.H.; Larsen, T.O.; Nielsen, K.F.; et al. Accurate prediction of secondary metabolite gene clusters in filamentous fungi. Proc. Natl. Acad. Sci. USA 2013, 110, E99–E107. [Google Scholar] [CrossRef] [Green Version]
- Matias, A.C.; Pedroso, N.; Teodoro, N.; Marinho, H.S.; Antunes, F.; Nogueira, J.M.F.; Herrero, E.; Cyrne, L. Down-regulation of fatty acid synthase increases the resistance of Saccharomyces cerevisiae cells to H(2)O(2). Free Radic. Bio. Med. 2007, 43, 1458–1465. [Google Scholar] [CrossRef]
- Nielsen, M.L.; Nielsen, J.B.; Rank, C.; Klejnstrup, M.L.; Holm, D.K.; Brogaard, K.H.; Hansen, B.G.; Frisvad, J.C.; Larsen, T.O.; Mortensen, U.H. A genome-wide polyketide synthase deletion library uncovers novel genetic links to polyketides and meroterpenoids in Aspergillus nidulans. FEMS Microbiol. Lett. 2011, 321, 157–166. [Google Scholar] [CrossRef]
- Adams, T.H.; Boylan, M.T.; Timberlake, W.E. brlA is necessary and sufficient to direct conidiophore development in Aspergillus nidulans. Cell 1988, 54, 353–362. [Google Scholar] [CrossRef]
- Clutterbuck, A.J. A mutational analysis of conidial development in Aspergillus nidulans. Genetics 1969, 63, 317–327. [Google Scholar] [CrossRef]
- Pascon, R.C.; Miller, B.L. Morphogenesis in Aspergillus nidulans requires Dopey (DopA), a member of a novel family of leucine zipper-like proteins conserved from yeast to humans. Mol. Microbiol. 2000, 36, 1250–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veith, D.; Scherr, N.; Efimov, V.P.; Fischer, R. Role of the spindle-pole-body protein ApsB and the cortex protein ApsA in microtubule organization and nuclear migration in Aspergillus nidulans. J. Cell Sci. 2005, 118, 3705–3716. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.Y.; Mead, M.E.; Kim, S.C.; Rokas, A.; Yu, J.H. WetA bridges cellular and chemical development in Aspergillus flavus. PLoS ONE 2017, 12, e0179571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayram, O.; Sari, F.; Braus, G.H.; Irniger, S. The protein kinase ImeB is required for light-mediated inhibition of sexual development and for mycotoxin production in Aspergillus nidulans. Mol. Microbiol. 2009, 71, 1278–1295. [Google Scholar] [CrossRef] [PubMed]
- Garrido, E.; Voss, U.; Muller, P.; Castillo-Lluva, S.; Kahmann, R.; Perez-Martin, J. The induction of sexual development and virulence in the smut fungus Ustilago maydis depends on Crk1, a novel MAPK protein. Gene Dev. 2004, 18, 3117–3130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimi, A.; Fujioka, T.; Mizutani, O.; Marui, J.; Hagiwara, D.; Abe, K. Mitogen-activated protein kinases MpkA and MpkB independently affect micafungin sensitivity in Aspergillus nidulans. Biosci. Biotechnol. Biochem. 2015, 79, 836–844. [Google Scholar] [CrossRef] [Green Version]
- Levin, D.E. Regulation of cell wall biogenesis in Saccharomyces cerevisiae: The cell wall integrity signaling pathway. Genetics 2011, 189, 1145–1175. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, T.; Mizutani, O.; Furukawa, K.; Sato, N.; Yoshimi, A.; Yamagata, Y.; Nakajima, T.; Abe, K. MpkA-Dependent and -independent cell wall integrity signaling in Aspergillus nidulans. Eukaryot. Cell 2007, 6, 1497–1510. [Google Scholar] [CrossRef] [Green Version]
- Lucena, R.; Alcaide-Gavilan, M.; Schubert, K.; He, M.; Domnauer, M.G.; Marquer, C.; Klose, C.; Surma, M.A.; Kellogg, D.R. Cell Size and Growth Rate Are Modulated by TORC2-Dependent Signals. Curr. Biol. 2018, 28, 196. [Google Scholar] [CrossRef] [Green Version]
- Samantaray, S.; Neubauer, M.; Helmschrott, C.; Wagener, J. Role of the guanine nucleotide exchange factor Rom2 in cell wall integrity maintenance of Aspergillus fumigatus. Eukaryot. Cell 2013, 12, 288–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desrivieres, S.; Cooke, F.T.; Parker, P.J.; Hall, M.N. MSS4, a phosphatidylinositol-4-phosphate 5-kinase required for organization of the actin cytoskeleton in Saccharomyces cerevisiae. J. Biol. Chem. 1998, 273, 15787–15793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahs, A.; Ischebeck, T.; Heilig, Y.; Stenzel, I.; Hempel, F.; Seiler, S.; Heilmann, I. The Essential Phosphoinositide Kinase MSS-4 Is Required for Polar Hyphal Morphogenesis, Localizing to Sites of Growth and Cell Fusion in Neurospora crassa. PLoS ONE 2012, 7, e51454. [Google Scholar] [CrossRef]
- De Souza, C.P.; Hashmi, S.B.; Osmani, A.H.; Andrews, P.; Ringelberg, C.S.; Dunlap, J.C.; Osmani, S.A. Functional analysis of the Aspergillus nidulans kinome. PLoS ONE 2013, 8, e58008. [Google Scholar] [CrossRef] [Green Version]
- Colabardini, A.C.; Brown, N.A.; Savoldi, M.; Goldman, M.H.; Goldman, G.H. Functional characterization of Aspergillus nidulans ypkA, a homologue of the mammalian kinase SGK. PLoS ONE 2013, 8, e57630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inagaki, M.; Schmelzle, T.; Yamaguchi, K.; Irie, K.; Hall, M.N.; Matsumoto, K. PDK1 homologs activate the Pkc1-mitogen-activated protein kinase pathway in yeast. Mol. Cell. Biol. 1999, 19, 8344–8352. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Rubio, G.; Sellers-Moya, A.; Martin, H.; Molina, M. Differential Role of Threonine and Tyrosine Phosphorylation in the Activation and Activity of the Yeast MAPK Slt2. Int. J. Mol. Sci. 2021, 22, 1110. [Google Scholar] [CrossRef]
- Boyce, K.J.; Andrianopoulos, A. Ste20-related kinases: Effectors of signaling and morphogenesis in fungi. Trends Microbiol. 2011, 19, 400–410. [Google Scholar] [CrossRef]
- Marker, R.; Blank-Landeshammer, B.; Beier-Rosberger, A.; Sickmann, A.; Kuck, U. Phosphoproteomic analysis of STRIPAK mutants identifies a conserved serine phosphorylation site in PAK kinase CLA4 to be important in fungal sexual development and polarized growth. Mol. Microbiol. 2020, 113, 1053–1069. [Google Scholar] [CrossRef]
- Riquelme, M.; Aguirre, J.; Bartnicki-Garcia, S.; Braus, G.H.; Feldbrugge, M.; Fleig, U.; Hansberg, W.; Herrera-Estrella, A.; Kamper, J.; Kuck, U.; et al. Fungal Morphogenesis, from the Polarized Growth of Hyphae to Complex Reproduction and Infection Structures. Microbiol. Mol. Biol. Rev. 2018, 82. [Google Scholar] [CrossRef] [Green Version]
- Virag, A.; Lee, M.P.; Si, H.; Harris, S.D. Regulation of hyphal morphogenesis by cdc42 and rac1 homologues in Aspergillus nidulans. Mol. Microbiol. 2007, 66, 1579–1596. [Google Scholar] [CrossRef] [Green Version]
- Tatebayashi, K.; Yamamoto, K.; Tanaka, K.; Tomida, T.; Maruoka, T.; Kasukawa, E.; Saito, H. Adaptor functions of Cdc42, Ste50, and Sho1 in the yeast osmoregulatory HOG MAPK pathway. EMBO J. 2006, 25, 3033–3044. [Google Scholar] [CrossRef]
- Silva, L.P.; Frawley, D.; Assis, L.J.; Tierney, C.; Fleming, A.B.; Bayram, O.; Goldman, G.H. Putative Membrane Receptors Contribute to Activation and Efficient Signaling of Mitogen-Activated Protein Kinase Cascades during Adaptation of Aspergillus fumigatus to Different Stressors and Carbon Sources. mSphere 2020, 5. [Google Scholar] [CrossRef]
- Gulli, M.P.; Jaquenoud, M.; Shimada, Y.; Niederhauser, G.; Wiget, P.; Peter, M. Phosphorylation of the Cdc42 exchange factor Cdc24 by the PAK-like kinase Cla4 may regulate polarized growth in yeast. Mol. Cell 2000, 6, 1155–1167. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.E.; Kang, P.J.; Park, H.O. Regulation of Cdc42 for polarized growth in budding yeast. Microb. Cell 2020, 7, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Knaus, M.; Pelli-Gulli, M.P.; Van Drogen, F.; Springer, S.; Jaquenoud, M.; Peter, M. Phosphorylation of Bem2p and Bem3p may contribute to local activation of Cdc42p at bud emergence. EMBO J. 2007, 26, 4501–4513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, I.; Eu, S.; Koch, D.; Bleimling, N.; Goody, R.S.; Muller, M.P. Structure of the tandem PX-PH domains of Bem3 from Saccharomyces cerevisiae. Acta Cryst. Sect. F Struct. Biol. Commun. 2018, 74, 315–321. [Google Scholar] [CrossRef] [PubMed]
- De Craene, J.O.; Bertazzi, D.L.; Bar, S.; Friant, S. Phosphoinositides, Major Actors in Membrane Trafficking and Lipid Signaling Pathways. Int. J. Mol. Sci. 2017, 18, 634. [Google Scholar] [CrossRef]
- Mitra, P.; Zhang, Y.; Rameh, L.E.; Ivshina, M.P.; McCollum, D.; Nunnari, J.J.; Hendricks, G.M.; Kerr, M.L.; Field, S.J.; Cantley, L.C.; et al. A novel phosphatidylinositol(3,4,5)P3 pathway in fission yeast. J. Cell Biol. 2004, 166, 205–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balla, T.; Gulyas, G.; Kim, Y.J.; Pemberton, J. Phosphoinositides and Calcium Signaling. A Marriage Arranged in Er-Pm Contact Sites. Curr. Opin. Physiol. 2020, 17, 149–157. [Google Scholar] [CrossRef]
- Pohlmann, J.; Risse, C.; Seidel, C.; Pohlmann, T.; Jakopec, V.; Walla, E.; Ramrath, P.; Takeshita, N.; Baumann, S.; Feldbrugge, M.; et al. The Vip1 inositol polyphosphate kinase family regulates polarized growth and modulates the microtubule cytoskeleton in fungi. PLoS Genet. 2014, 10, e1004586. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.; Lee, S.R.; Yang, K.S.; Ahn, Y.; Kim, Y.J.; Stadtman, E.R.; Rhee, S.G. Reversible oxidation and inactivation of the tumor suppressor PTEN in cells stimulated with peptide growth factors. Proc. Natl. Acad. Sci. USA 2004, 101, 16419–16424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.R.; Yang, K.S.; Kwon, J.; Lee, C.; Jeong, W.; Rhee, S.G. Reversible inactivation of the tumor suppressor PTEN by H2O2. J. Biol. Chem. 2002, 277, 20336–20342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Park, J.; Han, S.J.; Yang, S.Y.; Yoon, H.J.; Park, I.; Woo, H.A.; Lee, S.R. Redox regulation of tumor suppressor PTEN in cell signaling. Redox Biol. 2020, 34, 101553. [Google Scholar] [CrossRef]
- Lanze, C.E.; Gandra, R.M.; Foderaro, J.E.; Swenson, K.A.; Douglas, L.M.; Konopka, J.B. Plasma Membrane MCC/Eisosome Domains Promote Stress Resistance in Fungi. Microbiol. Mol. Biol. Rev. 2020, 84. [Google Scholar] [CrossRef]
- Li, L.; Naseem, S.; Sharma, S.; Konopka, J.B. Flavodoxin-Like Proteins Protect Candida albicans from Oxidative Stress and Promote Virulence. PLoS Pathog. 2015, 11, e1005147. [Google Scholar] [CrossRef] [Green Version]
- Roelants, F.M.; Leskoske, K.L.; Marshall, M.N.M.; Locke, M.N.; Thorner, J. The TORC2-Dependent Signaling Network in the Yeast Saccharomyces cerevisiae. Biomolecules 2017, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Gagliardi, P.A.; Puliafito, A.; Primo, L. PDK1: At the crossroad of cancer signaling pathways. Semin. Cancer Biol. 2018, 48, 27–35. [Google Scholar] [CrossRef]
- Chen, E.J.; Kaiser, C.A. LST8 negatively regulates amino acid biosynthesis as a component of the TOR pathway. J. Cell Biol. 2003, 161, 333–347. [Google Scholar] [CrossRef] [Green Version]
- Fillinger, S.; Chaveroche, M.K.; Shimizu, K.; Keller, N.; d’Enfert, C. cAMP and ras signalling independently control spore germination in the filamentous fungus Aspergillus nidulans. Mol. Microbiol. 2002, 44, 1001–1016. [Google Scholar] [CrossRef]
- de Castro, P.A.; dos Reis, T.F.; Dolan, S.K.; Manfiolli, A.O.; Brown, N.A.; Jones, G.W.; Doyle, S.; Riano-Pachon, D.M.; Squina, F.M.; Caldana, C.; et al. The Aspergillus fumigatus SchA(SCH9) kinase modulates SakA(HOG1) MAP kinase activity and it is essential for virulence. Mol. Microbiol. 2016, 102, 642–671. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.A.; Terasawa, M.; Mostofa, M.G.; Ushimaru, T. The TORC1-Nem1/Spo7-Pah1/lipin axis regulates microautophagy induction in budding yeast. Biochem. Biophys. Res. Commun. 2018, 504, 505–512. [Google Scholar] [CrossRef]
- Park, Y.; Han, G.S.; Mileykovskaya, E.; Garrett, T.A.; Carman, G.M. Altered Lipid Synthesis by Lack of Yeast Pah1 Phosphatidate Phosphatase Reduces Chronological Life Span. J. Biol. Chem. 2015, 290, 25382–25394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeyo, O.; Horn, P.J.; Lee, S.K.; Binns, D.D.; Chandrahas, A.; Chapman, K.D.; Goodman, J.M. The yeast lipin orthologue Pah1p is important for biogenesis of lipid droplets. J. Cell Biol. 2011, 192, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarc, E.; Petan, T. Lipid Droplets and the Management of Cellular Stress. Yale J. Biol. Med. 2019, 92, 435–452. [Google Scholar]
- Boisnard, S.; Lagniel, G.; Garmendia-Torres, C.; Molin, M.; Boy-Marcotte, E.; Jacquet, M.; Toledano, M.B.; Labarre, J.; Chedin, S. H2O2 activates the nuclear localization of Msn2 and Maf1 through thioredoxins in Saccharomyces cerevisiae. Eukaryot. Cell 2009, 8, 1429–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Moir, R.D.; Willis, I.M. Regulation of RNA Polymerase III Transcription Involves SCH9-dependent and SCH9-independent Branches of the Target of Rapamycin (TOR) Pathway. J. Biol. Chem. 2009, 284, 12604–12608. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Panepinto, J.C.; Fortwendel, J.R.; Fox, L.; Oliver, B.G.; Askew, D.S.; Rhodes, J.C. Deletion of the regulatory subunit of protein kinase a in Aspergillus fumigatus alters morphology, sensitivity to oxidative damage, and virulence. Infect. Immun. 2006, 74, 4865–4874. [Google Scholar] [CrossRef] [Green Version]
- Santhanam, A.; Hartley, A.; Duvel, K.; Broach, J.R.; Garrett, S. PP2A phosphatase activity is required for stress and Tor kinase regulation of yeast stress response factor Msn2p. Eukaryot. Cell 2004, 3, 1261–1271. [Google Scholar] [CrossRef] [Green Version]
- Touati, S.A.; Hofbauer, L.; Jones, A.W.; Snijders, A.P.; Kelly, G.; Uhlmann, F. Cdc14 and PP2A Phosphatases Cooperate to Shape Phosphoproteome Dynamics during Mitotic Exit. Cell Rep. 2019, 29, 2105. [Google Scholar] [CrossRef]
- Zhong, G.W.; Jiang, P.; Qiao, W.R.; Zhang, Y.W.; Wei, W.F.; Lu, L. Protein phosphatase 2A (PP2A) regulatory subunits ParA and PabA orchestrate septation and conidiation and are essential for PP2A activity in Aspergillus nidulans. Eukaryot. Cell 2014, 13, 1494–1506. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.K.; Andres, D.A. mTORC2 Is Required for Rit-Mediated Oxidative Stress Resistance. PLoS ONE 2014, 9, e115602. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Morigasaki, S.; Tatebe, H.; Tamanoi, F.; Shiozaki, K. Fission yeast TOR complex 2 activates the AGC-family Gad8 kinase essential for stress resistance and cell cycle control. Cell Cycle 2008, 7, 358–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatebe, H.; Murayama, S.; Yonekura, T.; Hatano, T.; Richter, D.; Furuya, T.; Kataoka, S.; Furuita, K.; Kojima, C.; Shiozaki, K. Substrate specificity of TOR complex 2 is determined by a ubiquitin-fold domain of the Sin1 subunit. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, T.; Cohen, P. Activation of serum- and glucocorticoid-regulated protein kinase by agonists that activate phosphatidylinositide 3-kinase is mediated by 3-phosphoinositide-dependent protein kinase-1 (PDK1) and PDK2. Biochem. J. 1999, 339, 319–328. [Google Scholar] [CrossRef]
- Xie, X.; Zhang, D.; Zhao, B.; Lu, M.K.; You, M.; Condorelli, G.; Wang, C.Y.; Guan, K.L. IkappaB kinase epsilon and TANK-binding kinase 1 activate AKT by direct phosphorylation. Proc. Natl. Acad. Sci. USA 2011, 108, 6474–6479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourgoint, C.; Rispal, D.; Berti, M.; Filipuzzi, I.; Helliwell, S.B.; Prouteau, M.; Loewith, R. Target of rapamycin complex 2-dependent phosphorylation of the coat protein Pan1 by Akl1 controls endocytosis dynamics in Saccharomyces cerevisiae. J. Biol. Chem. 2018, 293, 12043–12053. [Google Scholar] [CrossRef] [Green Version]
- Hatakeyama, R.; Kono, K.; Yoshida, S. Ypk1 and Ypk2 kinases maintain Rho1 at the plasma membrane by flippase-dependent lipid remodeling after membrane stresses. J. Cell Sci. 2017, 130, 1169–1178. [Google Scholar] [CrossRef] [Green Version]
- Conrad, M.; Schothorst, J.; Kankipati, H.N.; Van Zeebroeck, G.; Rubio-Texeira, M.; Thevelein, J.M. Nutrient sensing and signaling in the yeast Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2014, 38, 254–299. [Google Scholar] [CrossRef] [Green Version]
- Cardenas, M.E.; Cutler, N.S.; Lorenz, M.C.; Di Como, C.J.; Heitman, J. The TOR signaling cascade regulates gene expression in response to nutrients. Gene Dev. 1999, 13, 3271–3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crampton, N.; Kodiha, M.; Shrivastava, S.; Umar, R.; Stochaj, U. Oxidative stress inhibits nuclear protein export by multiple mechanisms that target FG nucleoporins and Crm1. Mol. Biol. Cell 2009, 20, 5106–5116. [Google Scholar] [CrossRef]
- Kodiha, M.; Stochaj, U. Nuclear transport: A switch for the oxidative stress-signaling circuit? J. Signal. Transduct. 2012, 2012, 208650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kodiha, M.; Tran, D.; Qian, C.; Morogan, A.; Presley, J.F.; Brown, C.M.; Stochaj, U. Oxidative stress mislocalizes and retains transport factor importin-alpha and nucleoporins Nup153 and Nup88 in nuclei where they generate high molecular mass complexes. Biochim. Biophys. Acta 2008, 1783, 405–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, E.J.; Wente, S.R. Dynamic nuclear pore complexes: Life on the edge. Cell 2006, 125, 1041–1053. [Google Scholar] [CrossRef] [Green Version]
- Raices, M.; D’Angelo, M.A. Nuclear pore complexes and regulation of gene expression. Curr. Opin. Cell Biol. 2017, 46, 26–32. [Google Scholar] [CrossRef]
- Napetschnig, J.; Blobel, G.; Hoelz, A. Crystal structure of the N-terminal domain of the human protooncogene Nup214/CAN. Proc. Natl. Acad. Sci. USA 2007, 104, 1783–1788. [Google Scholar] [CrossRef] [Green Version]
- De Souza, C.P.; Osmani, A.H.; Hashmi, S.B.; Osmani, S.A. Partial nuclear pore complex disassembly during closed mitosis in Aspergillus nidulans. Curr. Biol. 2004, 14, 1973–1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaunay, A.; Isnard, A.D.; Toledano, M.B. H2O2 sensing through oxidation of the Yap1 transcription factor. EMBO J. 2000, 19, 5157–5166. [Google Scholar] [CrossRef]
- Kos, I.; Patterson, M.J.; Znaidi, S.; Kaloriti, D.; da Silva Dantas, A.; Herrero-de-Dios, C.M.; d’Enfert, C.; Brown, A.J.; Quinn, J. Mechanisms Underlying the Delayed Activation of the Cap1 Transcription Factor in Candida albicans following Combinatorial Oxidative and Cationic Stress Important for Phagocytic Potency. Mbio 2016, 7, e00331. [Google Scholar] [CrossRef] [Green Version]
- He, X.J.; Mulford, K.E.; Fassler, J.S. Oxidative stress function of the Saccharomyces cerevisiae Skn7 receiver domain. Eukaryot. Cell 2009, 8, 768–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aristizabal, M.J.; Dever, K.; Negri, G.L.; Shen, M.; Hawe, N.; Benschop, J.J.; Holstege, F.C.P.; Krogan, N.J.; Sadowski, I.; Kobor, M.S. Regulation of Skn7-dependent, oxidative stress-induced genes by the RNA polymerase II-CTD phosphatase, Fcp1, and Mediator kinase subunit, Cdk8, in yeast. J. Biol. Chem. 2019, 294, 16080–16094. [Google Scholar] [CrossRef] [PubMed]
- Huisinga, K.L.; Pugh, B.F. A genome-wide housekeeping role for TFIID and a highly regulated stress-related role for SAGA in Saccharomyces cerevisiae. Mol. Cell 2004, 13, 573–585. [Google Scholar] [CrossRef]
- Sellam, A.; Askew, C.; Epp, E.; Lavoie, H.; Whiteway, M.; Nantel, A. Genome-wide Mapping of the Coactivator Ada2p Yields Insight into the Functional Roles of SAGA/ADA Complex in Candida albicans. Mol. Biol. Cell. 2009, 20, 2389–2400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zapater, M.; Sohrmann, M.; Peter, M.; Posas, F.; de Nadal, E. Selective requirement for SAGA in Hog1-mediated gene expression depending on the severity of the external osmostress conditions. Mol. Cell. Biol. 2007, 27, 3900–3910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaffer, M.P.; Jones, A.W.; Flynn, H.R.; Snijders, A.P.; Nurse, P. Quantitative Phosphoproteomics Reveals the Signaling Dynamics of Cell-Cycle Kinases in the Fission Yeast Schizosaccharomyces pombe. Cell Rep. 2018, 24, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Alejandro-Osorio, A.L.; Huebert, D.J.; Porcaro, D.T.; Sonntag, M.E.; Nillasithanukroh, S.; Lwill, J.; Gasch, A.P. The histone deacetylase Rpd3p is required for transient changes in genomic expression in response to stress. Genome Biol. 2009, 10. [Google Scholar] [CrossRef] [Green Version]
- de Nadal, E.; Zapater, M.; Alepuz, P.M.; Sumoy, L.; Mas, G.; Posas, F. The MAPK Hog1 recruits Rpd3 histone deacetylase to activate osmoresponsive genes. Nature 2004, 427, 370–374. [Google Scholar] [CrossRef]
- Todd, R.B. Regulation of Fungal Nitrogen Metabolism. In The Mycota III; Hoffmeister, D., Ed.; Springer International Publishing Switzerland: Cham, Switzerland, 2016; Volume 3, p. 414. [Google Scholar]
- Schinko, T.; Gallmetzer, A.; Amillis, S.; Strauss, J. Pseudo-constitutivity of nitrate-responsive genes in nitrate reductase mutants. Fungal Genet. Biol. 2013, 54, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Gallmetzer, A.; Silvestrini, L.; Schinko, T.; Gesslbauer, B.; Hortschansky, P.; Dattenbock, C.; Muro-Pastor, M.I.; Kungl, A.; Brakhage, A.A.; Scazzocchio, C.; et al. Reversible Oxidation of a Conserved Methionine in the Nuclear Export Sequence Determines Subcellular Distribution and Activity of the Fungal Nitrate Regulator NirA. PLoS Genet. 2015, 11, e1005297. [Google Scholar] [CrossRef] [Green Version]
- Schinko, T.; Berger, H.; Lee, W.; Gallmetzer, A.; Pirker, K.; Pachlinger, R.; Buchner, I.; Reichenauer, T.; Guldener, U.; Strauss, J. Transcriptome analysis of nitrate assimilation in Aspergillus nidulans reveals connections to nitric oxide metabolism. Mol. Microbiol. 2010, 78, 720–738. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.R.; Toone, W.M.; Mata, J.; Lyne, R.; Burns, G.; Kivinen, K.; Brazma, A.; Jones, N.; Bahler, J. Global transcriptional responses of fission yeast to environmental stress. Mol. Biol. Cell 2003, 14, 214–229. [Google Scholar] [CrossRef] [Green Version]
- Sylvain, M.A.; Liang, X.B.; Hellauer, K.; Turcotte, B. Yeast zinc cluster proteins Dal81 and Uga3 cooperate by targeting common coactivators for transcriptional activation of gamma-aminobutyrate responsive genes. Genetics 2011, 188, 523–534. [Google Scholar] [CrossRef] [Green Version]
- Galanopoulou, K.; Scazzocchio, C.; Galinou, M.E.; Liu, W.; Borbolis, F.; Karachaliou, M.; Oestreicher, N.; Hatzinikolaou, D.G.; Diallinas, G.; Amillis, S. Purine utilization proteins in the Eurotiales: Cellular compartmentalization, phylogenetic conservation and divergence. Fungal Genet. Biol. 2014, 69, 96–108. [Google Scholar] [CrossRef]
- Todd, R.B.; Fraser, J.A.; Wong, K.H.; Davis, M.A.; Hynes, M.J. Nuclear accumulation of the GATA factor AreA in response to complete nitrogen starvation by regulation of nuclear export. Eukaryot. Cell 2005, 4, 1646–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polotnianka, R.; Monahan, B.J.; Hynes, M.J.; Davis, M.A. TamA interacts with LeuB, the homologue of Saccharomyces cerevisiae Leu3p, to regulate gdhA expression in Aspergillus nidulans. Mol. Genet. Genom. 2004, 272, 452–459. [Google Scholar] [CrossRef]
- Macheda, M.L.; Hynes, M.J.; Davis, M.A. The Aspergillus nidulans gltA gene encoding glutamate synthase is required for ammonium assimilation in the absence of NADP-glutamate dehydrogenase. Curr. Genet. 1999, 34, 467–471. [Google Scholar] [CrossRef]
- Hemmings, B.A. Reactivation of the phospho form of the NAD-dependent glutamate dehydrogenase by a yeast protein phosphatase. Eur. J. Biochem. 1981, 116, 47–50. [Google Scholar] [CrossRef] [PubMed]
- Uno, I.; Matsumoto, K.; Adachi, K.; Ishikawa, T. Regulation of NAD-dependent glutamate dehydrogenase by protein kinases in Saccharomyces cerevisiae. J. Biol. Chem. 1984, 259, 1288–1293. [Google Scholar] [CrossRef]
- Pajares, M.; Jimenez-Moreno, N.; Dias, I.H.K.; Debelec, B.; Vucetic, M.; Fladmark, K.E.; Basaga, H.; Ribaric, S.; Milisav, I.; Cuadrado, A. Redox control of protein degradation. Redox Biol. 2015, 6, 409–420. [Google Scholar] [CrossRef] [Green Version]
- Natorff, R.; Piotrowska, M.; Paszewski, A. The Aspergillus nidulans sulphur regulatory gene sconB encodes a protein with WD40 repeats and an F-box. Mol. Gen. Genet. 1998, 257, 255–263. [Google Scholar] [CrossRef]
- Lauinger, L.; Flick, K.; Kaiser, P. Cdc48/Shp1 participates in dissociation of protein complexes to regulate their activity. Curr. Genet. 2021. [Google Scholar] [CrossRef]
- Sienko, M.; Natorff, R.; Skoneczny, M.; Kruszewska, J.; Paszewki, A.; Brzywczy, J. Regulatory mutations affecting sulfur metabolism induce environmental stress response in Aspergillus nidulans. Fungal Genet. Biol. 2014, 65, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.Z.; Gao, J.; Igbalajobi, O.; Skoneczny, M.; Sienko, M.; Maciejewska, A.M.; Brzywczy, J.; Fischer, R. The sulfur metabolism regulator MetR is a global regulator controlling phytochrome-dependent light responses in Aspergillus nidulans. Sci. Bull. 2021, 66, 592–602. [Google Scholar] [CrossRef]
- Boase, N.A.; Kelly, J.M. A role for creD, a carbon catabolite repression gene from Aspergillus nidulans, in ubiquitination. Mol. Microbiol. 2004, 53, 929–940. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Koepp, D.M. Hect E3 ubiquitin ligase Tom1 controls Dia2 degradation during the cell cycle. Mol. Biol. Cell 2012, 23, 4203–4211. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, I.; Keusch, J.J.; Challa, K.; Iesmantavicius, V.; Gasser, S.M.; Gut, H. The Sir4 H-BRCT domain interacts with phospho-proteins to sequester and repress yeast heterochromatin. EMBO J. 2019, 38. [Google Scholar] [CrossRef]
- Suresh, H.G.; Pascoe, N.; Andrews, B. The structure and function of deubiquitinases: Lessons from budding yeast. Open Biol. 2020, 10. [Google Scholar] [CrossRef]
- Huang, O.W.; Ma, X.L.; Yin, J.P.; Flinders, J.; Maurer, T.; Kayagaki, N.; Phung, Q.; Bosanac, I.; Arnott, D.; Dixit, V.M.; et al. Phosphorylation-dependent activity of the deubiquitinase DUBA. Nat. Struct. Mol. Biol. 2012, 19, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Harting, R.; Bayram, O.; Laubinger, K.; Valerius, O.; Braus, G.H. Interplay of the fungal sumoylation network for control of multicellular development. Mol. Microbiol. 2013, 90, 1125–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kors, S.; Geijtenbeek, K.; Reits, E.; Schipper-Krom, S. Regulation of Proteasome Activity by (Post-)transcriptional Mechanisms. Front. Mol. Biosci. 2019, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvaro, C.G.; O’Donnell, A.F.; Prosser, D.C.; Augustine, A.A.; Goldman, A.; Brodsky, J.L.; Cyert, M.S.; Wendland, B.; Thorner, J. Specific alpha-arrestins negatively regulate Saccharomyces cerevisiae pheromone response by down-modulating the G-protein-coupled receptor Ste2. Mol. Cell. Biol. 2014, 34, 2660–2681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.J.; He, L.Q.; Yao, K. The Antioxidative Function of Alpha-Ketoglutarate and Its Applications. Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef]
- Carey, B.W.; Finley, L.W.; Cross, J.R.; Allis, C.D.; Thompson, C.B. Intracellular alpha-ketoglutarate maintains the pluripotency of embryonic stem cells. Nature 2015, 518, 413–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuehne, A.; Emmert, H.; Soehle, J.; Winnefeld, M.; Fischer, F.; Wenck, H.; Gallinat, S.; Terstegen, L.; Lucius, R.; Hildebrand, J.; et al. Acute Activation of Oxidative Pentose Phosphate Pathway as First-Line Response to Oxidative Stress in Human Skin Cells. Mol. Cell 2015, 59, 359–371. [Google Scholar] [CrossRef] [Green Version]
- Izawa, S.; Maeda, K.; Miki, T.; Mano, J.; Inoue, Y.; Kimura, A. Importance of glucose-6-phosphate dehydrogenase in the adaptive response to hydrogen peroxide in Saccharomyces cerevisiae. Biochem. J. 1998, 330, 811–817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dal Santo, S.; Stampfl, H.; Krasensky, J.; Kempa, S.; Gibon, Y.; Petutschnig, E.; Rozhon, W.; Heuck, A.; Clausen, T.; Jonak, C. Stress-Induced GSK3 Regulates the Redox Stress Response by Phosphorylating Glucose-6-Phosphate Dehydrogenase in Arabidopsis. Plant. Cell 2012, 24, 3380–3392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, M.I.; Jalil, S.U.; Ansari, S.A.; Hasanuzzaman, M. GABA shunt: A key-player in mitigation of ROS during stress. Plant. Growth Regul. 2021, 94, 131–149. [Google Scholar] [CrossRef]
- Bouche, N.; Fait, A.; Bouchez, D.; Moller, S.G.; Fromm, H. Mitochondrial succinic-semialdehyde dehydrogenase of the gamma-aminobutyrate shunt is required to restrict levels of reactive oxygen intermediates in plants. Proc. Natl. Acad. Sci. USA 2003, 100, 6843–6848. [Google Scholar] [CrossRef] [Green Version]
- Márquez, D.; Escalera-Fanjul, X.; El Hafidi, M.; Aguirre López, B.; Riego-Ruiz, L.; González, A. Alanine represses GABA utilization and induces alanine transaminase required for mitochondrial function in Saccharomyces cerevisiae. Front. Microbiol. 2021, in press. [Google Scholar] [CrossRef]
- Zhou, Y.B.; Liu, C.; Tang, D.Y.; Yan, L.; Wang, D.; Yang, Y.Z.; Gui, J.S.; Zhao, X.Y.; Li, L.G.; Tang, X.D.; et al. The Receptor-Like Cytoplasmic Kinase STRK1 Phosphorylates and Activates CatC, Thereby Regulating H2O2 Homeostasis and Improving Salt Tolerance in Rice. Plant. Cell 2018, 30, 1100–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, S.G. Overview on Peroxiredoxin. Mol. Cells 2016, 39, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Becerril, A. Evaluación de NoxR y RacA como Posibles Reguladores de la Actividad de la NADPH Oxidasa NoxA de Aspergillus nidulans; Universidad Nacional Autónoma de México: México city, Mexico, 2010. [Google Scholar]
- Takemoto, D.; Tanaka, A.; Scott, B. A p67(Phox)-like regulator is recruited to control hyphal branching in a fungal-grass mutualistic symbiosis. Plant. Cell 2006, 18, 2807–2821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belambri, S.A.; Rolas, L.; Raad, H.; Hurtado-Nedelec, M.; Dang, P.M.; El-Benna, J. NADPH oxidase activation in neutrophils: Role of the phosphorylation of its subunits. Eur. J. Clin. Investig. 2018, 48 (Suppl. 2), e12951. [Google Scholar] [CrossRef] [Green Version]
- Dang, P.M.; Morel, F.; Gougerot-Pocidalo, M.A.; El Benna, J. Phosphorylation of the NADPH oxidase component p67(PHOX) by ERK2 and P38MAPK: Selectivity of phosphorylated sites and existence of an intramolecular regulatory domain in the tetratricopeptide-rich region. Biochemistry 2003, 42, 4520–4526. [Google Scholar] [CrossRef]
- Elliott, N.A.; Volkert, M.R. Stress induction and mitochondrial localization of Oxr1 proteins in yeast and humans. Mol. Cell. Biol. 2004, 24, 3180–3187. [Google Scholar] [CrossRef] [Green Version]
- Jaramillo-Gutierrez, G.; Molina-Cruz, A.; Kumar, S.; Barillas-Mury, C. The Anopheles gambiae oxidation resistance 1 (OXR1) gene regulates expression of enzymes that detoxify reactive oxygen species. PLoS ONE 2010, 5, e11168. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Lin, X.; Segers, F.; Suganthan, R.; Hildrestrand, G.A.; Rinholm, J.E.; Aas, P.A.; Sousa, M.M.L.; Holm, S.; Bolstad, N.; et al. OXR1A, a Coactivator of PRMT5 Regulating Histone Arginine Methylation. Cell Rep. 2020, 30, 4165–4178.e67. [Google Scholar] [CrossRef]
- Oliver, P.L.; Finelli, M.J.; Edwards, B.; Bitoun, E.; Butts, D.L.; Becker, E.B.E.; Cheeseman, M.T.; Davies, B.; Davies, K.E. Oxr1 Is Essential for Protection against Oxidative Stress-Induced Neurodegeneration. PLoS Genet. 2011, 7. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Protein | Phosphosites without H2O2 | Phosphosites with H2O2 |
|---|---|---|
| AN9008/FphA | S15, S74, S77, T79, S80 | S74, S77, T79, S80, S98, S100, S104, T105 |
| AN3102/PhkA | S94, S95, S452, T479, S480, T481 | S89, S90, S92, S94, S95, S96, T479, S480 |
| AN3101/PhkB | S396, T818, S819, S1454 | |
| AN2005/YpdA | S3 | |
| AN7697/SskA | T506, T508, S559, S792, S794 | T506, S508, S558, S559, S789, S792 |
| AN1180/SskB | S90, T92, T1175 | S90, T1173 |
| AN0931/PbsB | S174, S177, S179, S210 | S174, S179, S210 |
| AN1017/SakA/Hog1 | T171, Y173, T176 | |
| AN4668/MpkC | T176 | Y173 |
| AN6982/PtpA | S88, S408 | S88, S408, S412, S413 |
| AN4483/SrkA | S451 | S31, T412, T413, S451 |
| AN4000/FabM | S207 | T368, S376 |
| AN2911/AtfA | T159 | S132, S136 |
| AN3688/SrrA | T244 | |
| AN3469/H2B | S11, T12, S137, S138 | |
| AN3941/NimT | S256 | |
| AN5744 (Putative 14-3-3 like protein; ortholog of S. cerevisiae Bmh2) | S211 | |
| AN4182/NimX/Cdc28 | Y15 | T14,Y15 |
| AN3648/NimE | S136, S138, S142 | S136, S138, S142 |
| AN2412/CmkA | S376 | |
| AN8827/CmkC | S34, S475, S553 | S34, S140, S475 |
| Protein | Phosphosites without H2O2 | Phosphosites with H2O2 |
|---|---|---|
| AN2269/SteC | S197, S389, S535 | S197, S389, S535 |
| AN7252/SteD | S112, S243, S311, Y319 | |
| AN2701/HamE | S425, S901, S966, S973, S1037, S1039, S1040, S1043, T1111, S1112, S1115, S1503, S1536, T1538 | S425, S973, T1001, S1004, T1005, S1037, S1039, S1040, T1111, S1112, S1115, |
| AN3422/Ste7/MkkB | S359, S383 | S383 |
| AN3719/MpkB | Y184 | T182, Y184 |
| AN6190/SipA | S7, S16, S334, S335 | S334, S335, S337 |
| AN6611/SipC | S85, T415 | T84, S85 |
| AN4632/SipD | S482 | S482 |
| AN4085/SipF | T614, S615 | S615 |
| AN8071/StrA | S248, S3798 | S248, S379 |
| AN6190/SipA | S7, S16, S334, S335 | S334, S335, S337 |
| AN6611/SipC | S85, T415 | T84, S85 |
| AN4632/SipD | S482 | S482 |
| AN4085/SipF | T614, S615 | S615 |
| AN0807/LaeA | S39, S70 | S39, S70 |
| AN1052/VeA | S243 | S374 |
| AN2290/SteA | S550, T552 | S550, T552 |
| AN1959/VosA | T247 | |
| AN6243/ImeB | T211, Y212, S214, T215 | T45, S48, T55, T210, T211 |
| AN3090/GanA | S288 |
| Protein | Phosphosites without H2O2 | Phosphosites with H2O2 |
|---|---|---|
| AN4674 (Membrane protein Wsc4, putative; similar to of S. cerevisiae Wsc4 | T267, S283 | S283, S286 |
| AN4719/Similar to Rom1/2 Rho guanine nucleotide exchange factor; similar to of S. cerevisiae Rom2 | S277, S280, S281, S796, S797 | T42, T195, S277, S280, T281, S796, S797 |
| AN7576 (Predicted Rho GTPase activating protein, similar to S. cerevisiae Lrg1) | S83, S655, S679, S680, S764, S1163, T1164, S1165, T1166, S1168, S1170 | S83, T92, T93 S655, S679, S680, S764, S1163, T1164, S1165, T1166, S1168 |
| AN11994 (Predicted Rho GTPase activating protein, similar to of S. cerevisiae Bag7) | S306, S313, S315, S316, S318, S335, S337, T340, S341, S344, S348, T352, T366, S372, S394, S474, S627, T729, S731, S740, S741, T824 | S313, S315, S316, S318, S323, S335, S337, S364, T366, S372, S378, T389, T729, S735, S740, S741, |
| AN0106/PkcA | S586, S588, S594 | S586, S588, S590, S591, S594 |
| AN3110/PkhA (ortholog of S. cerevisiae Pkh2) | T134, S136, S150, S201, S203, S424, S425, T429, S617, S619, S755, S756, S759, S760, T764 | T134, S136, T207, S424, S425, S759, S760, T764 |
| AN4887/BckA | S763, T767, S955, S979, S983, S1003, S1014, S1033, T1037, S1059, S1061, T1067, S1234 | S719, S763, S979, S983, S1003, S1014, S1234 |
| AN4189/MkkA | S117, S121, T144, S149, S169, T367 | |
| AN5666/MpkA | T100, Y102, T105 | Y98, T100, Y102 |
| AN2984/RlmA | S317, S337 | T315, S317, S337 |
| Protein | Phosphosites without H2O2 | Phosphosites with H2O2 |
|---|---|---|
| AN7698/ShoA | S138, T207, S215 | S138 |
| AN7041/MsbA | S788 | |
| AN5592 (Predicted guanine nucleotide exchange factor ortholog of S. cerevisiae Cdc24) | S565, T579, S582, S627, S636, S666, S713, S715, S716, S760, S764, T766, T799, S800 | S525, S565, T579, S582, S584, S627, S636, S666, S670, S713, S715, S716, S760, S764 |
| AN0113/Bud3 | S229, T232, T480, S792, S818, S838, T1071, S1250, T1251, S1252, S1257, S1269 | S229, T232, T794, S818, T820, S838, T840, S1108, S1164, T1251, S1252, S1302 |
| AN3746 (Rho GTPase activator Bem2, putative) | S50, S160, S161, S236, S238 | S160, S161 |
| AN7030/BemA | T108, S115, S467, S470, S471, S476, S527, S528, T530, S533, T534 | S467, S476, S478, S527, S528, T530, S533, T534 |
| AN2067/Ste20 | S44 | S44 |
| AN8836/Cla4 | S295, S309 | T64, T66, S295, S309 |
| AN5787 (GTPase activator activity, role in negative regulation of Rho protein signal transduction, similar to of S. cerevisiae Bem3) | S475, S616 |
| Protein | Phosphosites without H2O2 | Phosphosites with H2O2 |
|---|---|---|
| Phosphatidylinositol kinases | ||
| AN2766/MssA (Putative phosphatidylinositol-4-phosphate 5-kinase; similar to of S. cerevisiae Mss4) | S65, S68, S90, S191, S196, S198, S208, S209, S258, S262, S270, S271 | S68, T181, S182, S196, S198, T200, S208, S209, S246, S247, S258, S262, T264, S271, T279, S284 |
| AN4278/Stt4/SstA | Y788, S790, T792 | |
| AN10791/Lsb6 | T64, T65, S84, S467, T671, S672, S675 | T64, T65, S84, S467, T671, S672, S675 |
| AN5797/VlpA (inositol polyphosphate kinase) | S94, S672 | S672 |
| Phosphatidylinositol transfer protein | ||
| AN3709 (CRAL/TRIO domain protein, similar to S. cerevisiae CSR1) | T168, S174 | |
| Phosphatidylinositol kinase regulator | ||
| AN5857/Vac7 | T422, S424, S350, Y463, T464, S465, S466, S575, S589 | S292, S350, S455, T459, T462, Y463, T464, S465, S466, T570, S575, S589, S795 |
| phosphatidylinositol phosphatases | ||
| AN7745 (Similar to S. cerevisiae INP52) | S17, S957 | S957 |
| AN8053/INP52 | S346, S350, S359 | |
| AN0925 (Phosphoric ester hydrolase, similar to S. cerevisiae Sac1) | S257, S258, S259 | S255, S257, S258, S259 |
| Phospholipases | ||
| AN10413 (Phosphatidylinositol-3-phosphate binding, phospholipase D activity) | S135, S167, S189, S204, T205, S208, S224, S294, S544, Y555, S581, S582, S590, S591, T593, T599, S1336 | S39, S47, S135, S189, S204, T205, S208, S294, S581, S582, T586, S590, S591, T593, T599, T603, S1336, S1338, S1461 |
| AN2947/PlcA (Putative 1-phosphatidylinositol-4,5-bisphosphate phosphodiesterase, phospholipase C) | S258, S262 | S258, S262, S263 |
| Known effectors | ||
| AN5705 (Ortholog of S. cerevisiae Vps74) | S29, S32, S36 | S25, S29, S32 |
| AN2496 (Similar to S. cerevisiae Efr3) | T254, T794, S795, S850, S854, S924, S1053, S1146, T1148, Y244, S245, | S245, T248, T254, T794, S795, S850, S854, S924, S1053, S1146, T1148 |
| AN0127 (Phosphatidylinositol-3,5-bisphosphate binding, phosphatidylinositol-3-phosphate binding, phosphatidylinositol-4-phosphate binding, ubiquitin binding activity, ortholog of S. cerevisiae ATG18) | S283, S285, S286, T296, S322 | S283, S286, S288, T296, T321, S322 |
| AN0576/Vps15 | S209 | S209 |
| AN6351/ATG20 | T255, S256, S260, S261, S504 | S260, S261, T503, S504, S505 |
| AN4416/PepA | T210, S148, T149, S195 | S148, T149, S195 |
| AN2224/SogB | S523, S525, S527, S547 | S523, S525, S527, S547 |
| AN4666 (Phosphatidylinositol-3,5-bisphosphate binding, ortholog of S. cerevisiae Hsv2) | S270 | S270 |
| AN11104 (Protein with a predicted role in actin assembly; similar to S. cerevisiae Las17) | S164, T315, S321, S355 | S280 |
| Protein | Phosphosites without H2O2 | Phosphosites with H2O2 |
|---|---|---|
| AN1060/kdmA | T246, S249 | |
| AN0986 (PHD finger domain protein | S73, S87, S358, S424 | S358, S424 |
| AN2857/Pho23 | S349, S467 | S349 |
| AN3668 (PHD finger domain protein, putative) | S509, S510, S707, S708, S711, S755, S756 | S509, S510, S755, S756 |
| AN4694 (Rpd3L complex) | S471 | T468, S471 |
| AN5640/Nmy1 | T285, S287, S313 | T285, S287, S304, S313 |
| AN6675/NTO1 | S11 | S6, S11 |
| AN7300 (Similar to S. cerevisiae Rco1) | S260, T439, S441, T443 | |
| AN8211 | S25, T298, S1297, S1301, S1520, T1623, S1640 | T298, S402, S1297, S1301, S1520, T1623, S1640 |
| AN8939 (PHD finger protein) | S199 | |
| Cti6/AN1777 | T48, S52, S180, T182, T448, S458, S463 | S180, S217, T448, T450, S453, S458, T460, S461 |
| AN0044 | S143, S396 | |
| AN7422 (Ubiquitin carboxyl-terminal hydrolase) | S350 | S350 |
| AN4637 | S60 | |
| AN5167 (Phospholipid binding, zinc ion binding activity) | S346, S349, S407, S410 | S349, S617, S620, S621 |
| AN5891 (PHD finger and SET domain protein, putative) | S186, S188 | |
| AN6136 | T401 | |
| AN9421 (RING finger domain protein, putative) | S199 | |
| AN4497 (MIZ zinc finger domain protein) | S133 | S133 |
| AN5516 (Meiosis specific protein Hop1, putative) | S398, T302, S304 | |
| AN10822/SizA | S36, S304 |
| Protein | Phosphosites without H2O2 | Phosphosites with H2O2 |
|---|---|---|
| AN7037/Vps36 | S269 | |
| AN7750/PSY2 Subunit of protein phosphatase PP4 complex | S803, S804, T805, S806, S807, T809, S820, S822 | S803, S804, T805, S806, S807, S808, T809, S822 |
| AN7783/ROM2 similar | S520, T522, S525, S733, S735 | S520, T522, S525, S636, Y639, S735 |
| AN0560/EXO84 | T157, S158, T159, S418 | T157, S158, T159 |
| AN0084/YRB1 | T54, S55, S57, S97 | S97 |
| AN5102/SPT16 | S587 | |
| AN5485/Nup2 | T762, S764, S895, S1005, T1269, S1270 | T1269, S1270 |
| AN3674/Meu6 | S173, T155, S161, T289, S290, S372, S373, S442, T457, S458, S462, S470, S493 | S173, S372, S373, S493, T457, S458, S462, S470 |
| AN4601/ATG26 | S125, T461, S485, S490, S507, S509, T510, S596, T597, T598 | S490, S509, T510 |
| AN6304/Sin1 | T73, T75, S337, S338, S342 | S337, S338, S342 |
| AN3438/SEC7 | S114, T134, S136, S166, S208, S211, S213, S214, S237, S487, S490, S491, S910, T912, S1033, S1035, S1348 | S164, S166, S207, S208, S211, S237, S283, S487, S490, S491, S910, S1035, S1348, S1349 |
| AN3424/OSH3 | S49, T54, S68, S180, S391, S402 | S180, S391, S402 |
| AN2749/SKG3 | S16, S17, S43, S478, S468, S718 | S468 |
| AN5829/NUM1 | S230, S234, S657, S892, S935, S1167, S1177, S1178, S1179 | S378, T656, S657, T799, S806, S833, S935, S1179 |
| Protein | Phosphosites without H2O2 | Phosphosites with H2O2 |
|---|---|---|
| AN6351/ATG20 Ortholog(s) have phosphatidylinositol-3-phosphate binding activity and role in autophagy of mitochondrion | T255, S256, S260, S261, S504 | S260, S261, T503, S504, S505 |
| AN3584/SNX4 Ortholog(s) have phosphatidylinositol-3-phosphate binding activity | S17 | |
| AN7030/BemA | T108, S115, S467, S470, S471, S476, S527, S528, T530, S533, T534 | S467, S476, S478, S527, S528, T530, S533, T534 |
| AN10918/MVP1 calcium ion binding, phosphatidylinositol binding activity | S236, S247, S271, S274, T276 | S236, S247, S271, S274, T276, S353 |
| AN2224/SogB Endosomal phosphatidylinositol-3-phosphate binding retromer complex subunit | S523, S525, S527, S547 | S523, S525, S527 |
| AN5787/Bem3 | S475, S616 | |
| AN4551/AN11912 (Phosphatidylinositol binding, SNAP receptor activity) | S15 | S15 |
| AN10350 phosphatidylinositol binding activity and role in cell communication | S298, S302, S817, S818 | S298, S302, S322, S817, S818 |
| AN6351/ATG20 Ortholog(s) have phosphatidylinositol-3-phosphate binding activity and role in autophagy of mitochondrion | T255, S256, S260, S261, S504 | S260, S261, T503, S504, S505 |
| Protein | Phosphosites without H2O2 | Phosphosites with H2O2 |
|---|---|---|
| AN5982/TorA | S1905, S2285, S2288, S2308 | S1905 |
| AN6304/Sin1/Avo1 | T73, T75, S337, S338, S342 | S337, S338, S342 |
| AN4639/RAPTOR/KOG1 | T13, S14, S19, S20, S51, S960, S987, S990 | T13, S14, S19, S51, S960, S985, S987, |
| AN10756/AN5958/TSC11/Rictor/Avo3 | S104, S105 | S104, S105, Y108 |
| AN1335/LST8 | S267 | S273 |
| AN7681/Maf1 | S82, S83, T147, T151, S154, S156, S233, T235, S237, T242, T255, S259, S261, T260, S305 | S82, S83, T147, T151, S156, T255, S259, S261, S305 |
| AN5973/PkcB/Gad8 | T450, S633 | S633 |
| AN3110/Ksg1/Pkh1/2 | T134, S136, S201, S203, S424, S425, T429, S617, S619, S755, S756, S759, S760, T764 | T134, S136, S207, S424, S425, S759, S760, T764 |
| AN7750/Psy2 | S716, S803, S804, T805, S806, S807, T809, S820, S822 | S803, S804, T805, S806, S807, S808, T809, S822 |
| AN4238/SchA | S240, S769 | S227, S233, S240, S359, S360, S769 |
| AN4171/Slm2 | S31, S33, T472, T481, T482, S494 | S31, S33, S468, T472 |
| AN5671/Slm1 | S75, S79, S135, S612, S656 | T73, S75, S132, S135 |
| AN0802/PAH1 | S112, S117, T118, S147, T188, S190, S222, S224 | S112, S182, S224 |
| AN6590/TSC2 | S459, S462 | S459, S779 |
| AN4987/PkaR | T37, S38, T119, S120 | T37, S38, S53 |
| AN4880/Psk1/YPK3 | S46, S389 | |
| AN7537/Ppk33/YPK2 | S403, T404, S406, S414, T415, S418, S541 | S414, S418, S541 |
| AN1933/ORM1 | S6 | S6 |
| AN2464/LagA/LAG1 | S7, S8, S12, S30, S387, T401, S403, T404 | S8, S12, S30, S387, S400 |
| AN1545/PabA/CDC55 | S154 | S417, S419, T421, S422, S423 |
| AN9467/ParA/RTS1 | S585, T586, T587 | S147, S585, T586, T587 |
| AN4085/TPD3 | T614, S615 | S615 |
| AN0144/FpkA/Nrc2/FPK1 | S102, S175, S255, S258, S437, T455, S457, T461 | T253, S255, S256, S258, S437, T455, S457 |
| AN8672/DnfA/DNF1 | S78, S79, S255, T258, T259, S261, S316, S320, S323 | S50, S79, S255, T259, S261, S316, S320, S323 |
| AN0351/GfdA/GPD1 | S272, S277, S279, S281, T282 | S279 |
| AN1343/NEM1 | S27, S153, S157, S309, S383 | S153, S158, S309 |
| AN10545/WHI2 | S103, S106, S107, S111 | S107, S109, S111 |
| AN5815/Aurora | S80 | |
| AN0182/RasA | S177, S180, S182 | S177, S180, S182 |
| AN2130/Cdc25 | S55, S68, S78, S80, S729, S749 | S55, S68, S80, S331, S713, S715, S721, S731, S733, T736, S749 |
| AN3913/CyaA | S121, S143, S399, S400, S401, S405, S1143, S1153 | S400, S401, S405 |
| AN0829/PdeA | S325 | |
| AN7572/SrrB/Rim15 | S353, T592, S593, S641, S648, S649, S650, S665, S683, S687, S977, S979, S982, S1433, S1437, T1471, S1474, S1819 | T34, S38, S353, S641, S648, S649, S650, S665, S979, S982, S1433, S1436, S1437, S1448, T1467, S1469, T1471, S1474, S1931 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrasco-Navarro, U.; Aguirre, J. H2O2 Induces Major Phosphorylation Changes in Critical Regulators of Signal Transduction, Gene Expression, Metabolism and Developmental Networks in Aspergillus nidulans. J. Fungi 2021, 7, 624. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080624
Carrasco-Navarro U, Aguirre J. H2O2 Induces Major Phosphorylation Changes in Critical Regulators of Signal Transduction, Gene Expression, Metabolism and Developmental Networks in Aspergillus nidulans. Journal of Fungi. 2021; 7(8):624. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080624
Chicago/Turabian StyleCarrasco-Navarro, Ulises, and Jesús Aguirre. 2021. "H2O2 Induces Major Phosphorylation Changes in Critical Regulators of Signal Transduction, Gene Expression, Metabolism and Developmental Networks in Aspergillus nidulans" Journal of Fungi 7, no. 8: 624. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080624