
The PPIP5K Family Member Asp1 Controls Inorganic Polyphosphate Metabolism in S. pombe


1
Eukaryotische Mikrobiologie, Institut für Funktionelle Genomforschung der Mikroorganismen, Heinrich Heine University, 40225 Düsseldorf, Germany
2
Department of Biomedical Sciences, Faculty of Health Sciences, Universidad Cardenal Herrera, CEU Universities, 46115 Valencia, Spain
3
Medical Research Council Laboratory for Molecular Cell Biology, University College London, London WC1E 6BT, UK
*
Authors to whom correspondence should be addressed.
J. Fungi 2021, 7(8), 626; https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080626
Submission received: 20 July 2021
/
Revised: 30 July 2021
/
Accepted: 30 July 2021
/
Published: 31 July 2021
(This article belongs to the Special Issue Inorganic Polyphosphate Metabolism in Yeasts)
Abstract
:Inorganic polyphosphate (polyP) which is ubiquitously present in both prokaryotic and eukaryotic cells, consists of up to hundreds of orthophosphate residues linked by phosphoanhydride bonds. The biological role of this polymer is manifold and diverse and in fungi ranges from cell cycle control, phosphate homeostasis and virulence to post-translational protein modification. Control of polyP metabolism has been studied extensively in the budding yeast Saccharomyces cerevisiae. In this yeast, a specific class of inositol pyrophosphates (IPPs), named IP7, made by the IP6K family member Kcs1 regulate polyP synthesis by associating with the SPX domains of the vacuolar transporter chaperone (VTC) complex. To assess if this type of regulation was evolutionarily conserved, we determined the elements regulating polyP generation in the distantly related fission yeast Schizosaccharomyces pombe. Here, the VTC machinery is also essential for polyP generation. However, and in contrast to S. cerevisiae, a different IPP class generated by the bifunctional PPIP5K family member Asp1 control polyP metabolism. The analysis of Asp1 variant S. pombe strains revealed that cellular polyP levels directly correlate with Asp1-made IP8 levels, demonstrating a dose-dependent regulation. Thus, while the mechanism of polyP synthesis in yeasts is conserved, the IPP player regulating polyP metabolism is diverse.
1. Introduction
Inorganic polyphosphate (polyP), the linear polymer of phosphate residues, is ubiquitously present in living organisms. Although polyP is one of the simplest of biological polymers, it regulates a pleiotropy of biological processes. Intrinsic to its polymeric charged nature is its ability to regulate phosphate and cation homeostasis. Additionally, polyP controls bacterial virulence [1], energy metabolism [2], protein folding [3], cell cycle progression [4], and signal transduction via lysine-polyphosphorylation [5]. This list of polyP-modulated functions is not exhaustive, and we recommend the interested readers the following polyP reviews [6,7,8,9].
The enzymology of synthesis of polyP is known for bacteria and for a number of unicellular eukaryotes, while in metazoans, the synthesis of polyP remains an open question [10]. Bacteria have two types of enzymes capable of synthesizing polyP, named polyphosphatekinases PPK1 and PPK2 [11]. In the social amoeba Dictyostelium discoideum, polyP is synthesized via a PPK1 homologous gene that was acquired by horizontal gene transfer [12]. In fungi such as the budding yeast Saccharomyces cerevisiae and protists such as Trypanosoma and Leishmania, the vacuole transporter chaperone (VTC) complex is responsible for polyP synthesis [13,14]. This complex is best characterized in S. cerevisiae were the VTC subunit four (Vtc4) possesses the catalytic domain synthesizing polyP [15]. Consequently, yeast strains with a VTC4 deletion have no or extremely low levels of polyP [16].
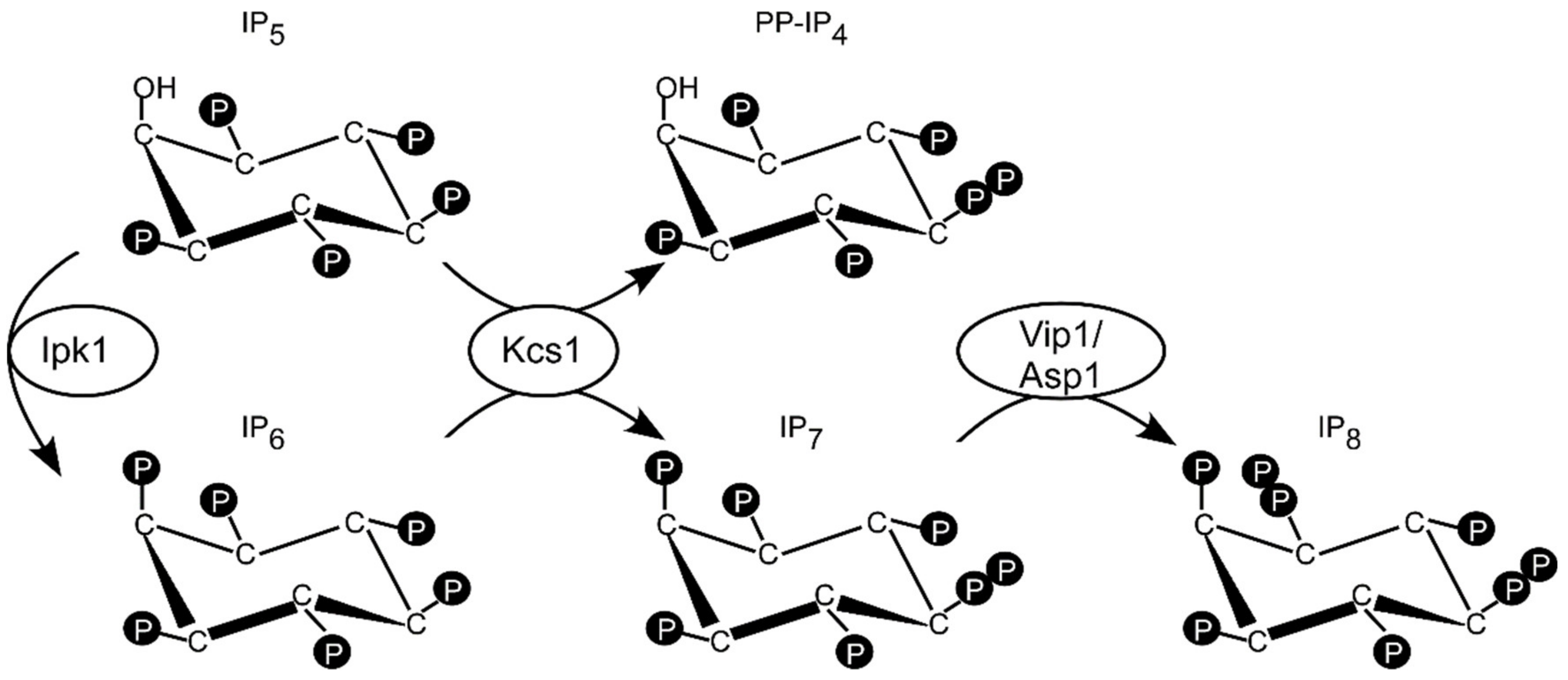
In S. cerevisiae polyP synthesis is regulated by inositol pyrophosphates (IPPs also known as PP-IP) that are a class of cellular messengers possessing one or more phosphoanhydride (pyro) moiety/ies. IPP synthesis is mediated by a cascade of conserved enzymes, starting with phospholipase C hydrolyzing PI(4,5)P2 to IP3 and diacylglycerol [17]. IP3 is the substrate of the Arg82 inositol polyphosphate multikinase generating IP4 and IP5 [18,19]. Phosphorylation of IP5 by the Ipk1 protein results in inositol hexakisphosphate IP6 (phytic acid) [20]. The best-studied IPPs are 5-IP7 (IP7) and 1,5-IP8 (IP8). IP7 is made by the Kcs1 enzyme family from IP6 (Figure 1) [21,22]. Kcs1 can also synthesize PP-IP4 from inositol pentakisphosphate IP5 [23] (Figure 1). IP8 is made by the PPIP5K family by adding a diphosphate group to position 1 of 5-IP7 [24,25,26,27] (Figure 1). PPIP5K members, which are named Vip1 in the budding yeast S. cerevisiae and Asp1 in the fission yeast Schizosaccharomyces pombe, are bifunctional enzymes with a N-terminal kinase domain and a C-terminal pyrophosphatase domain that converts IP8 back to IP7 [28] (Figure 1).
The S. cerevisiae Ksc1 enzyme is essential for intracellular polyP generation, as a kcs1∆ (deletion of the KCS1 gene) strain was identified in a genetic screening as possessing undetectable polyP levels [29]. Interestingly, strains with an ipk1Δ null allele, which are also unable to generate IP7 (Figure 1) have increased levels of polyP which led to the suggestion that IP7 does not control polyP synthesis. Instead, for PP-IP4, the other inositol pyrophosphate synthesized by Kcs1 is responsible for polyP synthesis (Figure 1) [29]. A subsequent study characterizing the ipk1Δkcs1Δ double mutant strain confirmed that polyP synthesis depends only on the fine control of Kcs1-synthesized IPPs [30]. Conversely, the S. cerevisiae vip1Δ strain possesses normal polyP levels, demonstrating that IP8 plays no role in the regulation of polyP synthesis [30].
IPPs activate Vtc4 by binding to its N-terminal SPX regulatory domain. The crystal structure of this domain, termed after SYG1/Pho81/XPR1 proteins, reveals a basic surface coordinating the inositol polyphosphates [31]. Extensive binding studies suggested that IP8 had a binding affinity for Vtc4 higher than IP7, However, further genetic studies and biochemical assays on purified vacuoles conclusively demonstrated that the IP7 synthesized by the S. cerevisiae Kcs1 is the physiological ligand activating Vtc4 polyP synthesis [32]. The SPX domain is present in several budding yeast and plant proteins involved in various aspects of phosphate homeostatic control. However, among human proteins only a single protein, the phosphate exported XPR1 [33], possesses this domain. In the fission yeast S. pombe, six proteins have a SPX domain including the Pi transmembrane transporter Pet1 and the VTC complex components Vtc2 and Vtc4 (Pombase at www.pombase.org).
To investigate if the Kcs1-controlled regulation of polyP synthesis is evolutionarily conserved, we analyzed modulation of polyP synthesis in another extensively studied yeast model, the fission yeast Schizosaccharomyces pombe. S. pombe evolutionary roots are considered to go back to early ascomycete lineage and the two yeasts diverged approximately 350 million years ago (reviewed in [34]). Thus, conservation or non-conservation of the control of a biological process between the two yeasts will point to a common or a more diverse control of the specific process in question, respectively.
2. Materials and Methods
2.1. Strains and Media
S. cerevisiae strains used are isogenic to BY4741 (MATa, his3Δ1; leu2Δ0; met15Δ0; ura3Δ0) and have been described [30].
S. pombe strains used in this study: vtc4Δ, asp1Δ, asp1D333A, asp1H397A, asp1I808D, and asp11−364 were generated by PCR-based gene targeting using the kanamycin resistance (Kanr) cassette as described previously [35]. The genotypes of the strains are listed in Table 1. Yeast cultures were grown in rich medium (YE5S) or as indicated in the text.
Plasmids harboring asp1365−920 and asp1365–920/H397A are derivatives of pJR2-3XL and expressed under the control of the thiamine-repressible nmt1+ promoter [35,36,37]. To de-repress the nmt1+ promoter, transformed yeast cells were grown under plasmid-selective conditions in thiamine-less minimal medium (MM) before harvesting.
2.2. Poly-P Extraction
Logarithmically growing cultures from the different S. cerevisiae or S. pombe strains (OD595 = 10 unit) were centrifuged at 1000× g for 2 min. The cell pellet was washed in water and resuspended in 250 μL of LETS Buffer (0.1 m LiCl, 10 mm EDTA, 10 mm Tris, pH 8.0, and 0.5% SDS) and mixed with 250 μL of phenol buffered at pH4.8. After adding glass beads the samples were vortexed for 5 min at 4 °C, followed by centrifugation at 15,000× g for 5 min at 4 °C. The polyP containing water phase was transferred to a new tube followed by chloroform extraction. The water phase was transferred to a new tube and RNA and polyP were precipitated by adding 2.5 volumes of ethanol and incubating at −20 °C for >16 h. Samples were spun down 15,000× g for 10 min at 4 °C, the RNA/polyP pellet was suspended in 100μL of ddH2O. RNA concentration was calculated by reading the absorbance at 260 nm.
2.3. PAGE Analysis of Extracted polyP
Extracted polyP (20 μg of RNA) were resolved on a 24 by 16 by 0.1 cm gel using a 30% non-denaturing polyacrylamide/bis acrylamide (19:1) in Tris/Borate/EDTA (TBE) buffer. Before loading the samples, gels were pre-run for 30 min at 200 V. Samples were resolved at 5 mA 600 V overnight at 4 °C, until the Orange-G front reached 10 cm from the gel’s bottom. Gels were stained with toluidine blue as previously described [38].
2.4. Phosphate Overplus Experiment
The S. pombe strain was grown in complete Pombe Glutamate Medium (PMG) [39]. Exponentially growing cells (OD595 = 50–70 units) were centrifuged at 1000× g for 2 min washed twice with phosphate-free PMG and divided into five identical cultures (8–10 mL each) in phosphate-free-PMG. After 2 h incubation at 30 °C, buffered potassium phosphate (pH 7.0) was added to a final concentration of 15.5 mM followed by further incubation for 2 h at 30 °C. Cells were harvested at the indicated times, washed with phosphate-free PMG, and stored at −20 °C until polyP extraction.
2.5. Quantification of polyP by Malachite Green
PolyP was measured as phosphate after enzymatic digestion of the polymer. RNA/polyP (2–5 μg) were incubated with recombinant Ppx1 and Ddp1 in reaction buffer (20 mM HEPEs pH 6.8; 6 mM MgSO4; 1 mM DTT; 100 mM NaCl) for 1 h at 37 °C. Phosphate present in 5–10% of digested RNA/polyP, and undigested RNA/polyP (1 μL) were assayed using the malachite green assay. Samples and phosphate standards were distributed in a 96 well plate and the volume adjusted to 100 μL with ddH2O. Then 100 μL of freshly mixed Molybtate (175 mM (NH4)2MoO4; 2 M H2SO4)/Malachite (0.15 malachite green; 1.4 g polyvinyl alcohol (100,000 MW) in 400 mL H2O) solution (4/3) was added to each well. Absorbance was measured at 650 nm after a 10 min incubation at room temperature. Potassium phosphate standard calibration curve was used to determine the concentration of polyP release phosphate after subtracting the amount of phosphate present in undigested samples.
2.6. Asp1 Phosphatase In Vitro Assay
The in vitro phosphatase assays were performed using 4 µg of bacterially generated and purified GST-Asp1365−920, GST-Asp1365−920/H397A or GST-Ddp1. Before running the assay, IP7 was generated via an in vitro kinase assay using the Asp1 kinase as previously described [28]. The GST-tagged proteins were incubated with either IP7 or polyP (type 15) in the reaction buffer (150mM Hepes pH 6.8, 250 mM NaCl, 30 mM MgSO4, and 5 mM DTT). All reactions were incubated for 18 h and the resulting inositol polyphosphates resolved on a 35.5% PAGE and stained with Toluidine Blue.
3. Results
3.1. The S. pombe PPIP5K Family Member Asp1 Controls polyP Generation
The metabolic relationship between inositol pyrophosphates and polyP has been primarily characterized in budding yeast [30]. To determine if polyP regulation by IPPs was a conserved feature, we compared the polyP levels of S. cerevisiae wild type, vtc4Δ, and vip1Δ strains with the respective S. pombe strains wild type, vtc4∆, and asp1Δ. Kcs1 is an essential protein in S. pombe as our tetrad analysis of a diploid, heterogenous kcs1+/ksc1∆ strain gave no viable ksc1∆ spores. Thus, a S. pombe kcs1Δ strain could not be included in our analysis. A SPX-containing homolog of the S. cerevisiae Vtc4 protein existsin S. pombe but has not been investigated (Pombase). To determine if the protein encoded by the S. pombe vtc4+ gene was required for polyP generation, we generated a vtc4∆ strain for the analysis.
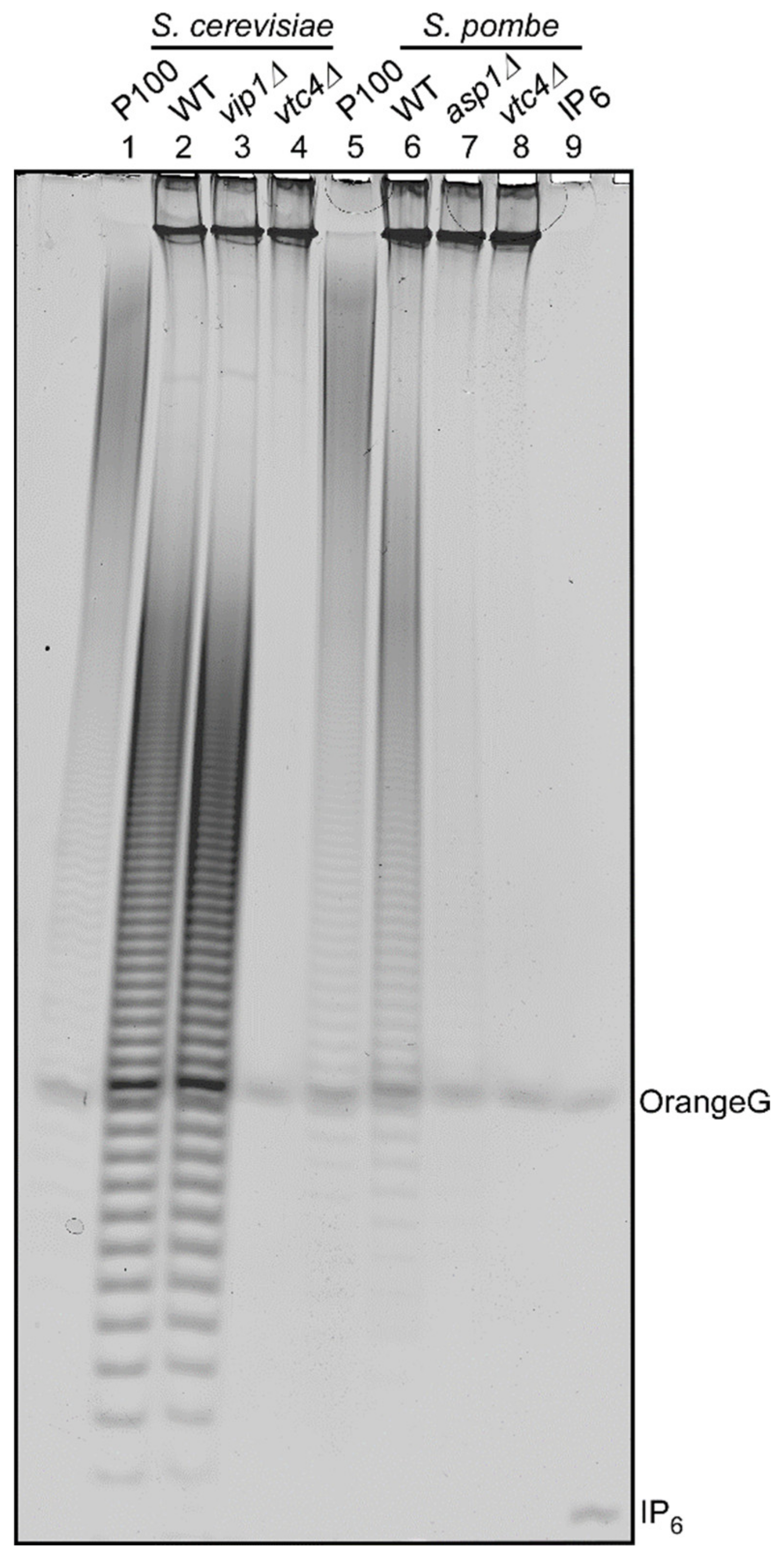
S. pombe and S cerevisiae strains were grown in rich media YE5S or YPD, respectively, and polyP was extracted from logarithmically growing cultures followed by PAGE analysis of phenol extracted polyP. As published previously, S. cerevisiae wild-type and vip1∆ strains have comparable amounts of polyP (Figure 2, lanes 2 and 3) but polyP is undetectable in the vtc4∆ strain (Figure 2, lane 4). Likewise, an S. pombe wild-type strain has polyP (Figure 2, lane 6), while the vtc4∆ strain does not (Figure 2, lane 8). Unexpectantly, and in contrast to the S. cerevisiae vip1∆ strain, deletion of the asp1+ gene encoding the S. pombe PPIP5K family member resulted in severely reduced polyP levels (Figure 2, lane 7). As Asp1 is solely responsible for the generation of IP8 in this yeast [28], we conclude that polyP generation in S. pombe requires IP8.
3.2. Asp1-Made IP8 Modulates polyP in a Dose-Dependent Manner
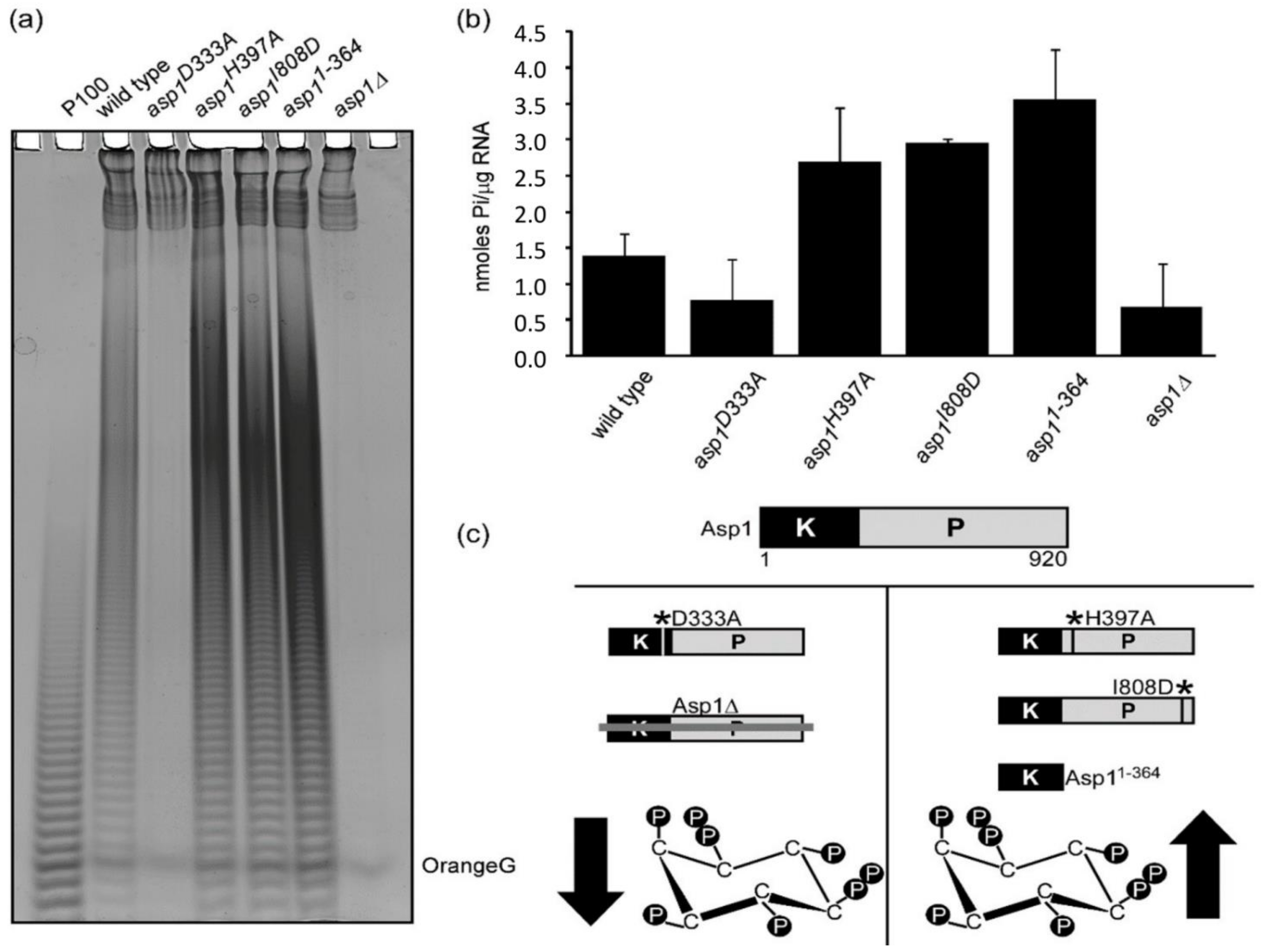
As a member of the PPIP5K bi-functional enzyme family, Asp1 has a N-terminal kinase domain and a C-terminal pyrophosphatase domain [24]. As the former generates IP8, which is used by the latter as a substrate, intracellular IP8 levels are regulated by the two opposing Asp1 enzymatic activities. This has allowed the generation of Asp1-variant strains, that have altered intracellular IP8 compared to a wild-type strain. All Asp1-variants are expressed in comparable amounts via the native asp1+ promoter as the wild-type asp1+ gene was replaced by the asp1 variant version(s) in the genome [28,35,40]. asp1D333A and asp1∆ strains cannot synthesize IP8 (Figure 3c), while strains asp1H397A, asp1I808D and asp11−364 all have approximately two-fold higher IP8 levels than the wild-type stain, due to a non-functional or absent pyrophosphatase domain (Figure 3c). Interestingly, strains unable to generate IP8 had massively reduced levels of polyP (Figure 3a and quantification in 3b) while strains with higher than wild-type levels of IP8 showed a higher than wild-type polyP levels (Figure 3b,c).
Furthermore, wild-type yeast transformants expressing the Asp1 pyrophosphatase domain Asp1365−920 on a plasmid, also had reduced polyP levels (Figure 4a). Yeast transformants with high-copy, plasmid-borne expression of asp1365−920 exhibit in severely reduced intracellular IP8 compared to control transformants [28]. Reduction in polyP levels was not observed when an enzymatically inactive version of the Asp1 pyrophosphatase asp1365−920H397A was expressed (Figure 4a). Plasmid-borne expression of this mutant Asp1 pyrophosphatase variant does not alter intracellular IP8 levels [28]. Thus, our analysis demonstrates an interrelation between IP8 and polyP in the fission yeast S. pombe and points to a dose-dependent IP8 regulation of polyP generation.
3.3. The Asp1 Phosphatase Cannot Metabolize polyP In Vitro
We have shown previously that the DIPP enzyme family homologue Ddp1 from S. cerevisiae which hydrolyzes IPPs also has polyP endopolyphosphatase activity [30]. To determine, if this might also be the case for the Asp1 pyrophosphatase, we generated and purified recombinant, bacterially-made GST-Asp1365−920, GST Asp1365−920H397A, and GST-Ddp1 proteins and tested them in an in vitro phosphatase assay using either Asp1-made IP7 or PolyP as substrate. As shown in Figure 4b (left panels), Ddp1 and Asp1365−920 have phosphatase activity using IP7 as the substrate. However, while Ddp1 also has endopolyphosphatase activity, Asp1365−920 does not (Figure 4b, right panels). We conclude that the reduction in intracellular polyP upon high-copy expression of the Asp1 pyrophosphatase, is not caused by Asp1365−920 polyP hydrolysis but by the reduction of IP8.
3.4. Polyphosphate Overplus Mechanism Is Conserved in S. pombe
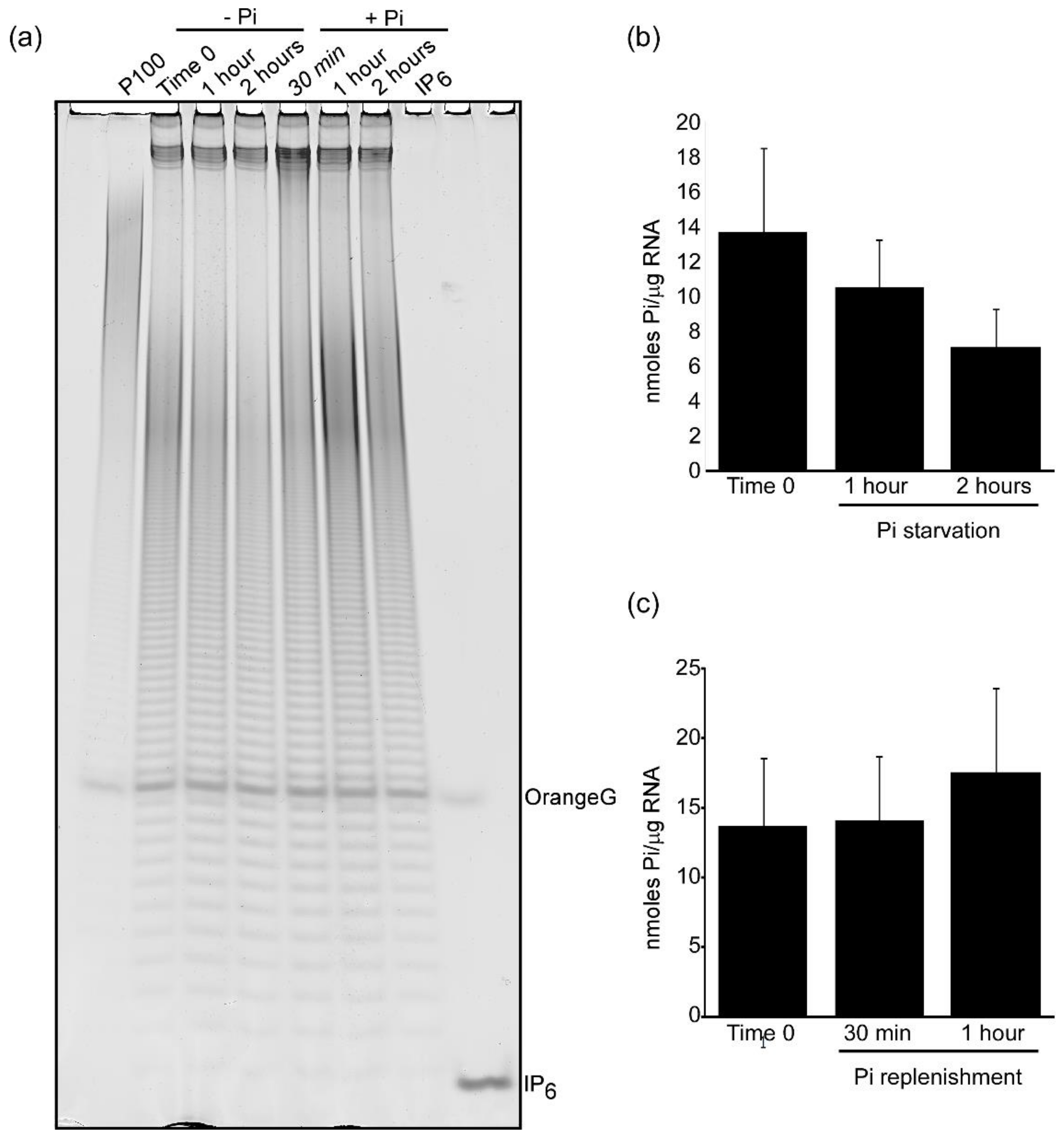
S. cerevisiae cells metabolically respond to phosphate starvation utilizing vacuole stored polyP that is rapidly degraded to supply cellular biochemistry with phosphates. The resupply of phosphate to phosphate starved yeast results in a rapid increase in polyP levels far exceeding its normal amount (overplus) [41,42,43]. To study if in S. pombe polyP follows the same metabolic fate, a wild type S. pombe strain was pre-grown in phosphate-containing minimal medium Pombe Glutamate Medium (PMG). Logarithmically growing yeast cultures were then shifted to phosphate-free PMG medium for 2 h followed by phosphate addition. PAGE analysis of polyP extracted at different time points revealed that shifting S. pombe cells into phosphate-free medium caused rapid polyP hydrolysis, resulting in a ~50% polyP decrease (Figure 5a left panel, quantification in Figure 5b). This is in contrast to the >80% decrease recorded for S. cerevisiae using a comparable experimental setup [30]. This shallow reduction in polyP observed in fission yeast shifted to phosphate-free condition is agreement with a previous report in which polyP depletion was observed after six hours incubation in phosphate-free medium [44]. Resupply of phosphate induced a rapid synthesis of polyP. One hour after phosphate-resupply, the polyP cellular level exceeded the polyP normally present in exponentially growing yeast (Figure 5a, right panels and Figure 5c). These data demonstrate that polyP in S. pombe follows a metabolic fate similar to the one recorded in other microorganisms.
4. Discussion
The dual-functionality PPIP5K enzymes generate and fine-tune IP8 cellular levels via their N-terminus kinase domain synthesizing IP8 and their-C-terminal phosphatase domain that degrades IP8 to IP7. IP8 modulated biological processes are manifold and in the fission yeast S. pombe we have shown that these high energy molecules, generated by Asp1, control genome stability by modulating mitotic machineries, adapt the yeast life cycle due to environmental signals and regulate the integrity of the cytoskeleton and mitochondria [28,35,37]. Recently, the Schwer lab has identified Asp1 as a regulator of phosphate homeostasis controlling RNA 3’processing/transcription termination of S. pombe phosphate acquisition genes [45]. Our present analysis shows that Asp1 generated IP8 also controls polyP synthesis in a dose-dependent manner; thus, pinpointing to a central role of IP8 in phosphate homeostasis in this yeast.
Phosphate homeostasis is one of the fundamental biological events regulated by IPPs [46,47]. In budding yeast, this relationship has been investigated from a signaling perspective trough the activation of the PHO pathway [48] as well as metabolically trough the accumulation of polyP inside the vacuole [29,30]. Pioneering works carried out in S. cerevisiae elucidated the molecular mechanism regulating polyP vacuolar accumulation: a combination of structural, genetic, and biochemical approaches has emphasized a specific role for the Kcs1 generated 5-IP7 in regulating Vtc4 polyP-synthesizing activity, and consequently the accumulation of the polymer inside the budding yeast vacuole [31,32]. Consequently S. cerevisiae kcs1Δ strains have undetectable [29] or very low polyP levels [30]. Conversely, S. cerevisiae vip1Δ strains unable to synthesize IP8 and thus accumulating 5-IP7 have normal [30] or even higher than WT levels of polyp [32].
Intriguingly, our present work uncovers that the IPPs selectivity regulating cellular polyP levels is not conserved among species. While the members of the Vtc4 family in both budding and fission yeast are responsible for polyP synthesis, different enzymes in the two species control polyP levels. The S. pombe asp1Δ strain possesses IPPs deficiency similar to the budding yeast vip1Δ mutant, as it is devoid of IP8 and contains higher level of 5-IP7 [28]. However, and in stark contrast to the findings for the S. cerevisiae vip1 strain, the S. pombe asp1Δ strain has very low levels of polyP. Analysis of several strains expressing Asp1 variants with deficient kinase or phosphatase function, clearly demonstrated that in S. pombe IP8 modulates polyP cellular level, and it is doing so in a dose-dependent manner.
How is this distinct IPP regulation of polyP levels in fission and budding yeasts achieved? Crystal structure analysis of the SPX regulatory domain revealed a basic surface where negative charged molecules such as IP6 or IPPs can bind [31]. Binding studies carried out on several SPX domains provided an affinity scale whereby IP8 > IP7 > IP6 [31,32] substantially reflecting the decreasing charging nature of the inositol phosphate tested. These structural and binding studies did not reveal a selective binding pocket that may explain SPX IPP discrimination. Therefore, if the differential IPP regulation of polyP cellular level between budding and fission yeast is due to a differential IPP activation of Vtc4, additional structural elements must exist. Potential candidates include another protein or small molecule cofactor that confer selectivity to the SPX domain. A better characterization of the fission yeast VTC complex is necessary to address this important issue. We cannot exclude at present that the different IPPs regulation of polyP level between fission and budding yeast is independent from SPX/Vtc4 paradigm.
The dose-dependent regulation of polyP generation by intracellular IP8 levels was manifested by an increase in intracellular IP8 levels above wild-type levels leading to a polyP increase above wild-type levels. At present it is unclear how this modulation is achieved on a molecular level. However, it is important to note that Asp1-made IP8 also controls microtubule dynamics [37,40]. The S. pombe interphase microtubule cytoskeleton controls the nucleus-proximal localization of vacuoles as well as their fusion/fission dynamics [49,50]. As microtubule stability is increased or decreased with increasing or decreasing IP8 levels, respectively, it is possible that vacuolar membrane dynamics and thus the membrane bound VTC complexes functionality is influenced by the IP8-modulated microtubule dynamics.
Author Contributions
Author contribution is as follows: Conceptualization A.S., U.F. and M.P.-O.; Methodology E.W., M.P.-O., U.F. and A.S.; Validation, E.W., M.P.-O. and A.S.; Formal Analysis, M.P.-O.; Investigation, E.W., M.P.-O. and A.S.; Resources, U.F., M.P.-O., E.W. and A.S.; Writing—Original Draft Preparation, U.F. and A.S.; Writing—Review & Editing, U.F. and A.S.; Visualization, M.P.-O. and U.F.; Supervision, U.F. and A.S.; Project Administration, U.F. and A.S.; Funding Acquisition, A.S. and U.F. All authors have read and agreed to the published version of the manuscript.
Funding
A.S. is supported by Medical Research Council grant MR/T028904/1. This work was supported by the Deutsche Forschungsgemeinschaft project FOR1334 to U.F.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rashid, M.H.; Rumbaugh, K.; Passador, L.; Davies, D.G.; Hamood, A.N.; Iglewski, B.H.; Kornberg, A. Polyphosphate kinase is essential for biofilm development, quorum sensing, and virulence of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2000, 97, 9636–9641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solesio, M.E.; Xie, L.; McIntyre, B.; Ellenberger, M.; Mitaishvili, E.; Bhadra-Lobo, S.; Bettcher, L.F.; Bazil, J.N.; Raftery, D.; Jakob, U.; et al. Depletion of mitochondrial inorganic polyphosphate (polyP) in mammalian cells causes metabolic shift from oxidative phosphorylation to glycolysis. Biochem. J. 2021, 478, 1631–1646. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.; Wholey, W.-Y.; Wagner, N.O.; Cremers, C.M.; Mueller-Schickert, A.; Hock, N.T.; Krieger, A.G.; Smith, E.M.; Bender, R.A.; Bardwell, J.C.; et al. Polyphosphate is a primordial chaperone. Mol. Cell 2014, 53, 689–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bru, S.; Láinez, J.M.M.; Hernández-Ortega, S.; Quandt, E.; Torres-Torronteras, J.; Marti, R.; Canadell, D.; Ariño, J.; Sharma, S.; Jiménez, J.; et al. Polyphosphate is involved in cell cycle progression and genomic stability in Saccharomyces cerevisiae. Mol. Microbiol. 2016, 101, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, C.; Livermore, T.; Saiardi, A. Protein polyphosphorylation of lysine residues by inorganic polyphosphate. Mol. Cell 2015, 58, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Rao, N.N.; Gómez-García, M.R.; Kornberg, A. Inorganic polyphosphate: Essential for growth and survival. Annu. Rev. Biochem. 2009, 78, 605–647. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, J.; Bru, S.; Ribeiro, M.; Clotet, J. Polyphosphate: Popping up from oblivion. Curr. Genet. 2017, 63, 15–18. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Jakob, U. Inorganic polyphosphate, a multifunctional polyanionic protein scaffold. J. Biol. Chem. 2019, 294, 2180–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, S.N.J.; Docampo, R. Polyphosphate and its diverse functions in host cells and pathogens. PLoS Pathog. 2013, 9, e1003230. [Google Scholar] [CrossRef]
- Desfougères, Y.; Saiardi, A.; Azevedo, C. Inorganic polyphosphate in mammals: Where’s Wally? Biochem. Soc. Trans. 2020, 48, 95–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Ishige, K.; Kornberg, A. A polyphosphate kinase (PPK2) widely conserved in bacteria. Proc. Natl. Acad. Sci. USA 2002, 99, 16678–16683. [Google Scholar] [CrossRef] [Green Version]
- Livermore, T.M.; Chubb, J.R.; Saiardi, A. Developmental accumulation of inorganic polyphosphate affects germination and energetic metabolism in Dictyostelium discoideum. Proc. Natl. Acad. Sci. USA 2016, 113, 996–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, C.O.; Ruiz, F.A.; Vieira, M.; Hill, J.; Docampo, R. An acidocalcisomal exopolyphosphatase from Leishmania major with high affinity for short chain polyphosphate. J. Biol. Chem. 2002, 277, 50899–50906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulakovskaya, T.V.; Andreeva, N.A.; Ledova, L.A.; Ryazanova, L.P.; Trilisenko, L.V.; Eldarov, M.A. Enzymes of polyphosphate metabolism in yeast: Properties, functions, practical significance. Biochemistry 2021, 86, S96–S108. [Google Scholar] [CrossRef] [PubMed]
- Hothorn, M.; Neumann, H.; Lenherr, E.D.; Wehner, M.; Rybin, V.; Hassa, P.O.; Uttenweiler, A.; Reinhardt, M.; Schmidt, A.; Seiler, J.; et al. Catalytic core of a membrane-associated eukaryotic polyphosphate polymerase. Science 2009, 324, 513–516. [Google Scholar] [CrossRef]
- Tomashevsky, A.; Kulakovskaya, E.; Trilisenko, L.; Kulakovskiy, I.; Kulakovskaya, T.; Fedorov, A.; Eldarov, M. VTC4 polyphosphate polymerase knockout increases stress resistance of saccharomyces cerevisiae cells. Biology 2021, 10, 487. [Google Scholar] [CrossRef]
- Berridge, M.J. Inositol trisphosphate and calcium signalling. Nat. Cell Biol. 1993, 361, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Odom, A.R.; Stahlberg, A.; Wente, S.R.; York, J.D. A role for nuclear inositol 1,4,5-trisphosphate kinase in transcriptional control. Science 2000, 287, 2026–2029. [Google Scholar] [CrossRef] [PubMed]
- Saiardi, A.; Nagata, E.; Luo, H.R.; Sawa, A.; Luo, X.; Snowman, A.M.; Snyder, S.H. Mammalian inositol polyphosphate multikinase synthesizes inositol 1,4,5-trisphosphate and an inositol pyrophosphate. Proc. Natl. Acad. Sci. USA 2001, 98, 2306–2311. [Google Scholar] [CrossRef] [Green Version]
- York, J.D.; Odom, A.R.; Murphy, R.; Ives, E.B.; Wente, S.R. A phospholipase C-dependent inositol polyphosphate kinase pathway required for efficient messenger RNA export. Science 1999, 285, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Draškovič, P.; Saiardi, A.; Bhandari, R.; Burton, A.; Ilc, G.; Kovačevič, M.; Snyder, S.H.; Podobnik, M. Inositol hexakisphosphate kinase products contain diphosphate and triphosphate groups. Chem. Biol. 2008, 15, 274–286. [Google Scholar] [CrossRef] [Green Version]
- Saiardi, A.; Erdjument-Bromage, H.; Snowman, A.M.; Tempst, P.; Snyder, S.H. Synthesis of diphosphoinositol pentakisphosphate by a newly identified family of higher inositol polyphosphate kinases. Curr. Biol. 1999, 9, 1323–1326. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Lev, S.; Saiardi, A.; Desmarini, D.; Sorrell, T.; Djordjevic, J.T. Identification of a major IP5 kinase in cryptococcus neoformans confirms that PP-IP5/IP7, not IP6, is essential for virulence. Sci. Rep. 2016, 6, 23927. [Google Scholar] [CrossRef]
- Fridy, P.; Otto, J.C.; Dollins, D.E.; York, J.D. Cloning and characterization of two human VIP1-like inositol hexakisphosphate and diphosphoinositol pentakisphosphate kinases. J. Biol. Chem. 2007, 282, 30754–30762. [Google Scholar] [CrossRef] [Green Version]
- Mulugu, S.; Bai, W.; Fridy, P.C.; Bastidas, R.J.; Otto, J.C.; Dollins, D.E.; Haystead, T.A.; Ribeiro, A.A.; York, J.D. A conserved family of enzymes that phosphorylate inositol hexakisphosphate. Science 2007, 316, 106–109. [Google Scholar] [CrossRef]
- Wang, H.; Falck, J.; Hall, T.M.T.; Shears, S. Structural basis for an inositol pyrophosphate kinase surmounting phosphate crowding. Nat. Chem. Biol. 2011, 8, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Fridy, P.; Ribeiro, A.A.; Choi, J.H.; Barma, D.K.; Vogel, G.; Falck, J.; Shears, S.; York, J.D.; Mayr, G.W. Structural analysis and detection of biological inositol pyrophosphates reveal that the family of VIP/diphosphoinositol pentakisphosphate kinases are 1/3-kinases. J. Biol. Chem. 2009, 284, 1863–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual-Ortiz, M.; Saiardi, A.; Walla, E.; Jakopec, V.; Künzel, N.A.; Span, I.; Vangala, A.; Fleig, U. Asp1 bifunctional activity modulates spindle function via controlling cellular inositol pyrophosphate levels in schizosaccharomyces pombe. Mol. Cell. Biol. 2018, 38, e00047-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auesukaree, C.; Tochio, H.; Shirakawa, M.; Kaneko, Y.; Harashima, S. Plc1p, Arg82p, and Kcs1p, enzymes involved in inositol pyrophosphate synthesis, are essential for phosphate regulation and polyphosphate accumulation in saccharomyces cerevisiae. J. Biol. Chem. 2005, 280, 25127–25133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonetti, A.; Szijgyarto, Z.; Bosch, D.; Loss, O.; Azevedo, C.; Saiardi, A. Identification of an evolutionarily conserved family of inorganic polyphosphate endopolyphosphatases. J. Biol. Chem. 2011, 286, 31966–31974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wild, R.; Gerasimaite, R.; Jung, J.-Y.; Truffault, V.; Pavlovic, I.; Schmidt, A.; Saiardi, A.; Jessen, H.J.; Poirier, Y.; Hothorn, M.; et al. Control of eukaryotic phosphate homeostasis by inositol polyphosphate sensor domains. Science 2016, 352, 986–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerasimaite, R.; Pavlovic, I.; Capolicchio, S.; Hofer, A.; Schmidt, A.; Jessen, H.J.; Mayer, A. Inositol pyrophosphate specificity of the SPX-dependent polyphosphate polymerase VTC. ACS Chem. Biol. 2017, 12, 648–653. [Google Scholar] [CrossRef]
- Azevedo, C.; Saiardi, A. Correction: The new world of inorganic polyphosphates. Biochem. Soc. Trans. 2016, 44, 1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, C.S.; Wood, V.; Fantes, P.A. An ancient yeast for young geneticists: A primer on the schizosaccharomyces pombe model system. Genetica 2015, 201, 403–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pöhlmann, J.; Fleig, U. Asp1, a conserved 1/3 inositol polyphosphate kinase, regulates the dimorphic switch in schizosaccharomyces pombe. Mol. Cell. Biol. 2010, 30, 4535–4547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribas, J.C.; Moreno, M.B.; Durán, A. A family of multifunctional thiamine-repressible expression vectors for fission yeast. Yeast 2000, 16, 861–872. [Google Scholar] [CrossRef]
- Topolski, B.; Jakopec, V.; Künzel, N.A.; Fleig, U. Inositol pyrophosphate kinase Asp1 modulates chromosome segregation fidelity and spindle function in schizosaccharomyces pombe. Mol. Cell. Biol. 2016, 36, 3128–3140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Losito, O.; Szijgyarto, Z.; Resnick, A.C.; Saiardi, A. Inositol pyrophosphates and their unique metabolic complexity: Analysis by gel electrophoresis. PLoS ONE 2009, 4, e5580. [Google Scholar] [CrossRef] [PubMed]
- Rai, S.K.; Atwood-Moore, A.; Levin, H.L. High-frequency lithium acetate transformation of schizosaccharomyces pombe. Breast Cancer 2018, 1721, 167–177. [Google Scholar] [CrossRef]
- Pöhlmann, J.; Risse, C.; Seidel, C.; Pohlmann, T.; Jakopec, V.; Walla, E.; Ramrath, P.; Takeshita, N.; Baumann, S.; Feldbrügge, M.; et al. The Vip1 inositol polyphosphate kinase family regulates polarized growth and modulates the microtubule cytoskeleton in fungi. PLoS Genet. 2014, 10, e1004586. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, N.; DeRisi, J.; Brown, P.O. New components of a system for phosphate accumulation and polyphosphate metabolism in saccharomyces cerevisiae revealed by genomic expression analysis. Mol. Biol. Cell 2000, 11, 4309–4321. [Google Scholar] [CrossRef] [Green Version]
- Harold, F.M. Inorganic polyphosphates in biology: Structure, metabolism, and function. Bacteriol. Rev. 1966, 30, 772–794. [Google Scholar] [CrossRef]
- Dawes, E.A.; Senior, P.J. The role and regulation of energy reserve polymers in micro-organisms. Adv. Microb. Physiol. 1973, 10, 135–266. [Google Scholar] [CrossRef]
- Muller, J. Synthesis and degradation of polyphosphate in the fission yeast schizosaccharomyces pombe: Mutations in phosphatase genes do not affect polyphosphate metabolism. FEMS Microbiol. Lett. 1992, 92, 151–156. [Google Scholar] [CrossRef]
- Sanchez, A.; Garg, A.; Shuman, S.; Schwer, B. Inositol pyrophosphates impact phosphate homeostasis via modulation of RNA 3′ processing and transcription termination. Nucleic Acids Res. 2019, 47, 8452–8469. [Google Scholar] [CrossRef]
- Wilson, M.; Livermore, T.; Saiardi, A. Inositol pyrophosphates: Between signalling and metabolism. Biochem. J. 2013, 452, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, C.; Saiardi, A. Eukaryotic phosphate homeostasis: The inositol pyrophosphate perspective. Trends Biochem. Sci. 2017, 42, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-S.; Mulugu, S.; York, J.D.; O’Shea, E.K. Regulation of a Cyclin-CDK-CDK inhibitor complex by inositol pyrophosphates. Science 2007, 316, 109–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadwick, W.L.; Biswas, S.K.; Bianco, S.; Chan, Y.-H.M. Non-random distribution of vacuoles in schizosaccharomyces pombe. Phys. Biol. 2020, 17, 065004. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, D.; Pollard, P.; Win, T.Z.; Hyams, J.S. Myosin V-mediated vacuole distribution and fusion in fission yeast. Curr. Biol. 2001, 11, 1124–1127. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Schematic representation of the inositol pyrophosphates metabolic pathway in S. pombe and S. cerevisiae. Inositol hexakisphosphate IP6 synthesized by Ipk1 is further phosphorylated to IP7 by the Kcs1 enzyme, that is also metabolizing IP5 to PP-IP4. IP8 is synthesized by Asp1 (S. pombe) or Vip1 (S. cerevisiae).
Figure 1.
Schematic representation of the inositol pyrophosphates metabolic pathway in S. pombe and S. cerevisiae. Inositol hexakisphosphate IP6 synthesized by Ipk1 is further phosphorylated to IP7 by the Kcs1 enzyme, that is also metabolizing IP5 to PP-IP4. IP8 is synthesized by Asp1 (S. pombe) or Vip1 (S. cerevisiae).

Figure 2.
Synthesis of polyP in S. pombe depends on Asp1 function. Phenol-extracted PolyP from exponentially growing yeast cultures and equal amounts of RNA (20 μg) were resolved on 30% PAGE and visualized by toluidine staining. The extracts of the following strains were analyzed: (from left to right) S. cerevisiae wild-type (WT), vip1Δ, and vtc4Δ and S. pombe wild-type (WT), asp1Δ, and vtc4Δ. The figure shows a representative analysis from 3 independent experiments with virtually identical results.
Figure 2.
Synthesis of polyP in S. pombe depends on Asp1 function. Phenol-extracted PolyP from exponentially growing yeast cultures and equal amounts of RNA (20 μg) were resolved on 30% PAGE and visualized by toluidine staining. The extracts of the following strains were analyzed: (from left to right) S. cerevisiae wild-type (WT), vip1Δ, and vtc4Δ and S. pombe wild-type (WT), asp1Δ, and vtc4Δ. The figure shows a representative analysis from 3 independent experiments with virtually identical results.

Figure 3.
Asp1 generated IP8 modulate polyP synthesis in a dose dependent manner. (a) Asp1 generated IP8 regulate cellular polyP levels. S. pombe polyP detected as a dark smear by PAGE analysis of lysates of the wild-type, asp1D333A, asp1H397A, asp1I808D, asp11−364, and asp1Δ strains. (b) quantification of polyP levels shown in (a) by Malachite Green assay. (c) schematic representation of the correlation between Asp1 pyrophosphatase activity and IP8 levels in S. pombe. All Asp1 variants are expressed endogenously instead of the WT Asp1. asp1Δ and asp1D333A strains have no IP8, while strains asp1H397A, asp1I808D, and asp11−364 generate more IP8 than the wild-type strain. * denotes postion of mutation in Asp1 variant.
Figure 3.
Asp1 generated IP8 modulate polyP synthesis in a dose dependent manner. (a) Asp1 generated IP8 regulate cellular polyP levels. S. pombe polyP detected as a dark smear by PAGE analysis of lysates of the wild-type, asp1D333A, asp1H397A, asp1I808D, asp11−364, and asp1Δ strains. (b) quantification of polyP levels shown in (a) by Malachite Green assay. (c) schematic representation of the correlation between Asp1 pyrophosphatase activity and IP8 levels in S. pombe. All Asp1 variants are expressed endogenously instead of the WT Asp1. asp1Δ and asp1D333A strains have no IP8, while strains asp1H397A, asp1I808D, and asp11−364 generate more IP8 than the wild-type strain. * denotes postion of mutation in Asp1 variant.

Figure 4.
Asp1 phosphatase cannot hydrolyze polyP. (a) top panel: polyP analysis (detected as a dark smear by PAGE analysis of lysates) of a wild-type S. pombe strain transformed with either a control plasmid or plasmids expressing asp1365−920 or asp1365-920/H397A under nmt1+ promoter de-repressed conditions. Bottom panel: quantification of polyP levels using a Malachite Green assay. (b) the in vitro phosphatase activity of the proteins Asp1365−920, Asp1365−920/H397A, or S. cerevisiae Ddp1 were determined using IP7 (left panel) or polyP (right panel) as substrate. (c) Western blot analysis of GST-tagged proteins Ddp1, Asp1365−920, and Asp1365-920/H397A expressed and purified from bacteria.
Figure 4.
Asp1 phosphatase cannot hydrolyze polyP. (a) top panel: polyP analysis (detected as a dark smear by PAGE analysis of lysates) of a wild-type S. pombe strain transformed with either a control plasmid or plasmids expressing asp1365−920 or asp1365-920/H397A under nmt1+ promoter de-repressed conditions. Bottom panel: quantification of polyP levels using a Malachite Green assay. (b) the in vitro phosphatase activity of the proteins Asp1365−920, Asp1365−920/H397A, or S. cerevisiae Ddp1 were determined using IP7 (left panel) or polyP (right panel) as substrate. (c) Western blot analysis of GST-tagged proteins Ddp1, Asp1365−920, and Asp1365-920/H397A expressed and purified from bacteria.

Figure 5.
polyP metabolism in response to phosphate starvation is conserved in S. pombe. (a) polyP profile of S. pombe cells during phosphate starvation, -Pi (left) and phosphate resupply conditions, +Pi (right). (b) quantification by Malachite Green assay of polyP levels in phosphate-starving cells. (c) quantification of polyP levels by Malachite Green assay after phosphate-resupply. The gel shows a representative analysis from four independent experiments quantified as average +/− SD in the graphs.
Figure 5.
polyP metabolism in response to phosphate starvation is conserved in S. pombe. (a) polyP profile of S. pombe cells during phosphate starvation, -Pi (left) and phosphate resupply conditions, +Pi (right). (b) quantification by Malachite Green assay of polyP levels in phosphate-starving cells. (c) quantification of polyP levels by Malachite Green assay after phosphate-resupply. The gel shows a representative analysis from four independent experiments quantified as average +/− SD in the graphs.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
S. pombe strains used in this study.
| Name | Genotype | Source |
|---|---|---|
| UFY605 | his3-D1, ade6-M210, leu1-32, ura4-D18, h− | K. Gould |
| UFY1156 | asp1Δ::kanR his3-D1 ade6-M216 leu1-32 ura4-D18, h− | U. Fleig |
| UFY1511 | asp1D333A::kanR his3-D1 ade6-M210 leu1-32 ura4-D18, h+ | U. Fleig |
| UFY1579 | asp1H397A::kanR his3-D1 ade6-M210 leu1-32 ura4-D18, h+ | U. Fleig |
| UFY2294 | asp11−364::kanR his3-D1 ade6-M21x leu1-32 ura4-D18, h+ | U. Fleig |
| UFY2553 | asp1I808D::kanR his3-D1 ade6-M216 leu1-32 ura4-D18, h− | U. Fleig |
| UFY3033 | vtc4Δ::kanRhis3-D1 ade6-M210 leu1-32 ura4-D18, h- | U. Fleig |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pascual-Ortiz, M.; Walla, E.; Fleig, U.; Saiardi, A. The PPIP5K Family Member Asp1 Controls Inorganic Polyphosphate Metabolism in S. pombe. J. Fungi 2021, 7, 626. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080626
AMA Style
Pascual-Ortiz M, Walla E, Fleig U, Saiardi A. The PPIP5K Family Member Asp1 Controls Inorganic Polyphosphate Metabolism in S. pombe. Journal of Fungi. 2021; 7(8):626. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080626
Chicago/Turabian StylePascual-Ortiz, Marina, Eva Walla, Ursula Fleig, and Adolfo Saiardi. 2021. "The PPIP5K Family Member Asp1 Controls Inorganic Polyphosphate Metabolism in S. pombe" Journal of Fungi 7, no. 8: 626. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080626
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.