
Effect of Nutritional Factors and Copper on the Regulation of Laccase Enzyme Production in Pleurotus ostreatus

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Fungal Strain and Culture Conditions
2.3. Biomass Production
2.4. Biochemical Analyses
2.4.1. Spectrometric Analyses
2.4.2. Zymogram
2.5. Microscopy Observation
2.6. RNA Isolation and Transcriptional Analyses
2.6.1. Real-Time qPCR
2.6.2. mRNA-Seq Analysis
2.7. Differential Gene Expression Analysis and Gene Annotation
2.8. Statistical Analyses
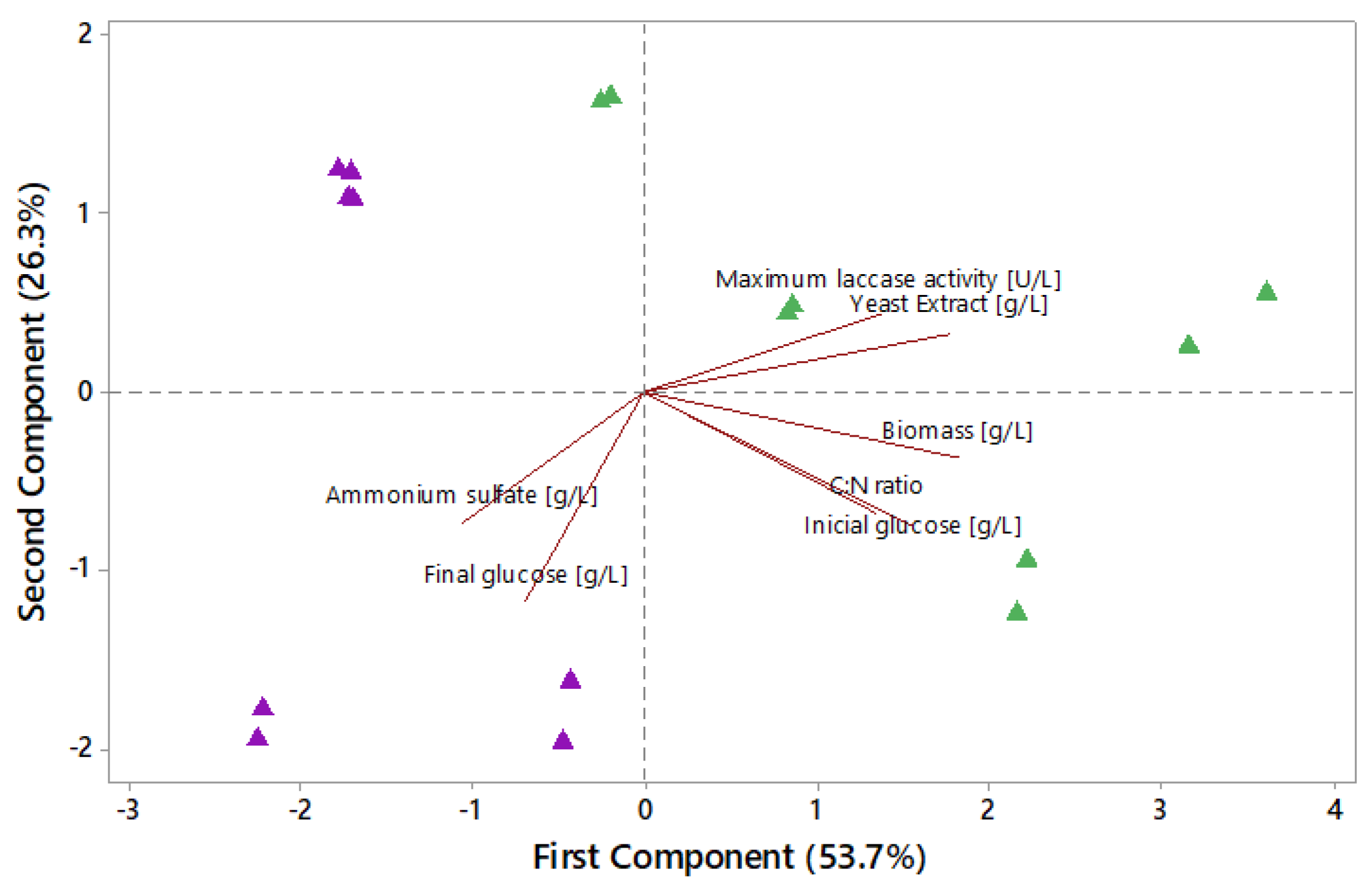
2.8.1. Principal Component Analysis (PCA)
2.8.2. Central Composite Design (CCD)
3. Results
3.1. Effects of Inorganic and Organic Nitrogen Source on Laccase Activity
3.2. Effect of Glucose and Yeast Extract Concentrations on Laccase Activity
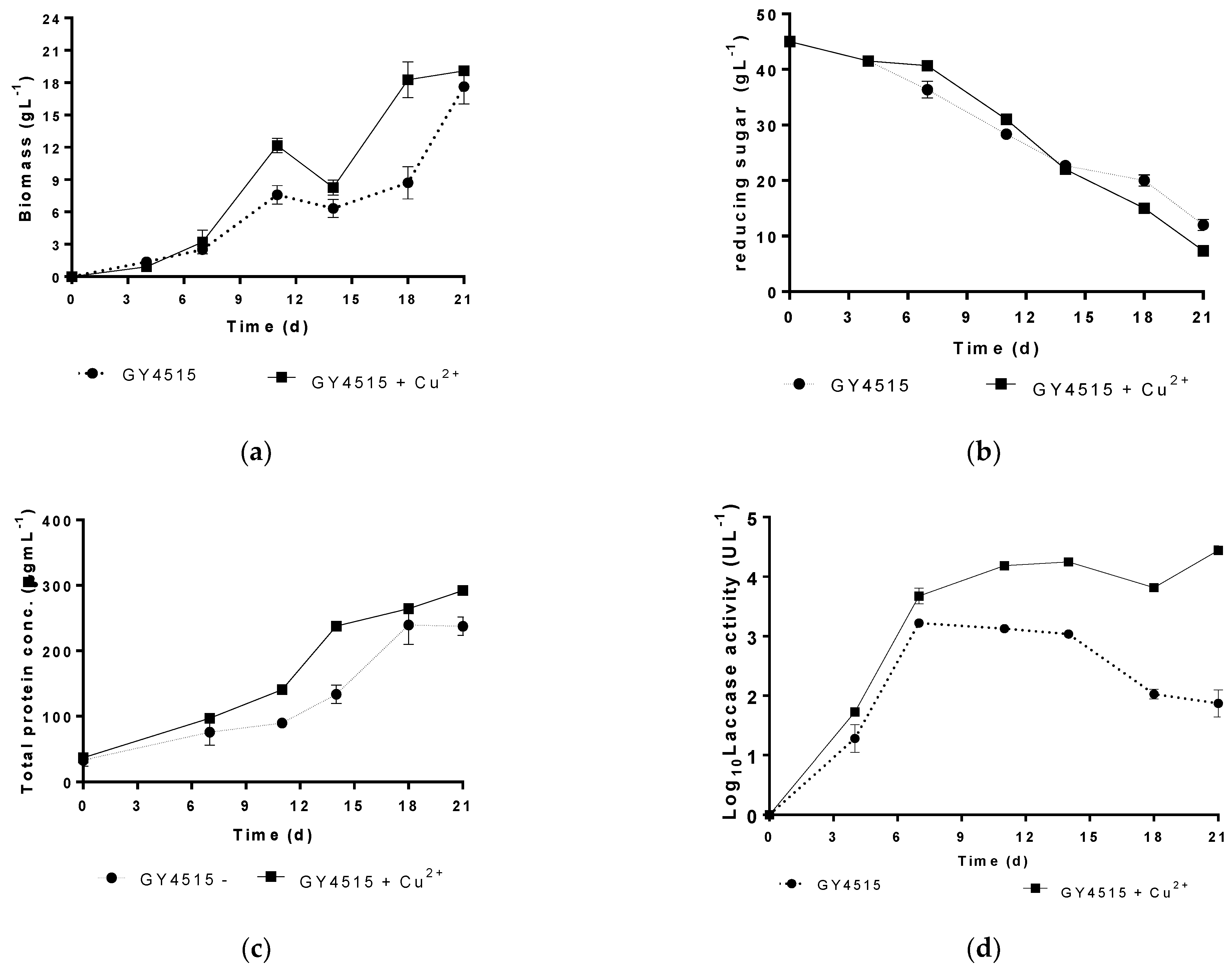
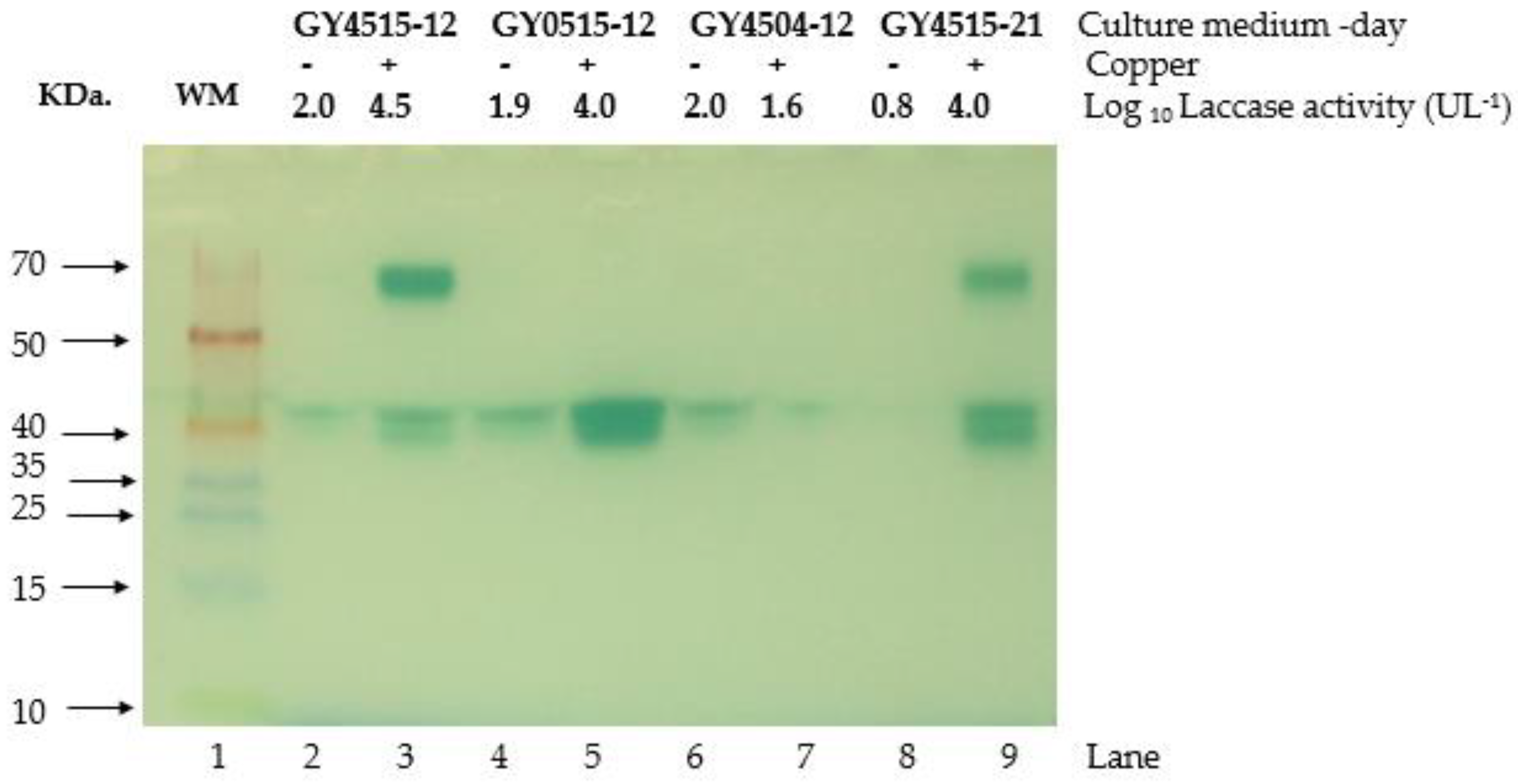
3.3. Characterization of the Growth and Laccase Activity of P. ostreatus under Nutrient Sufficient Conditions with and without Copper Sulfate
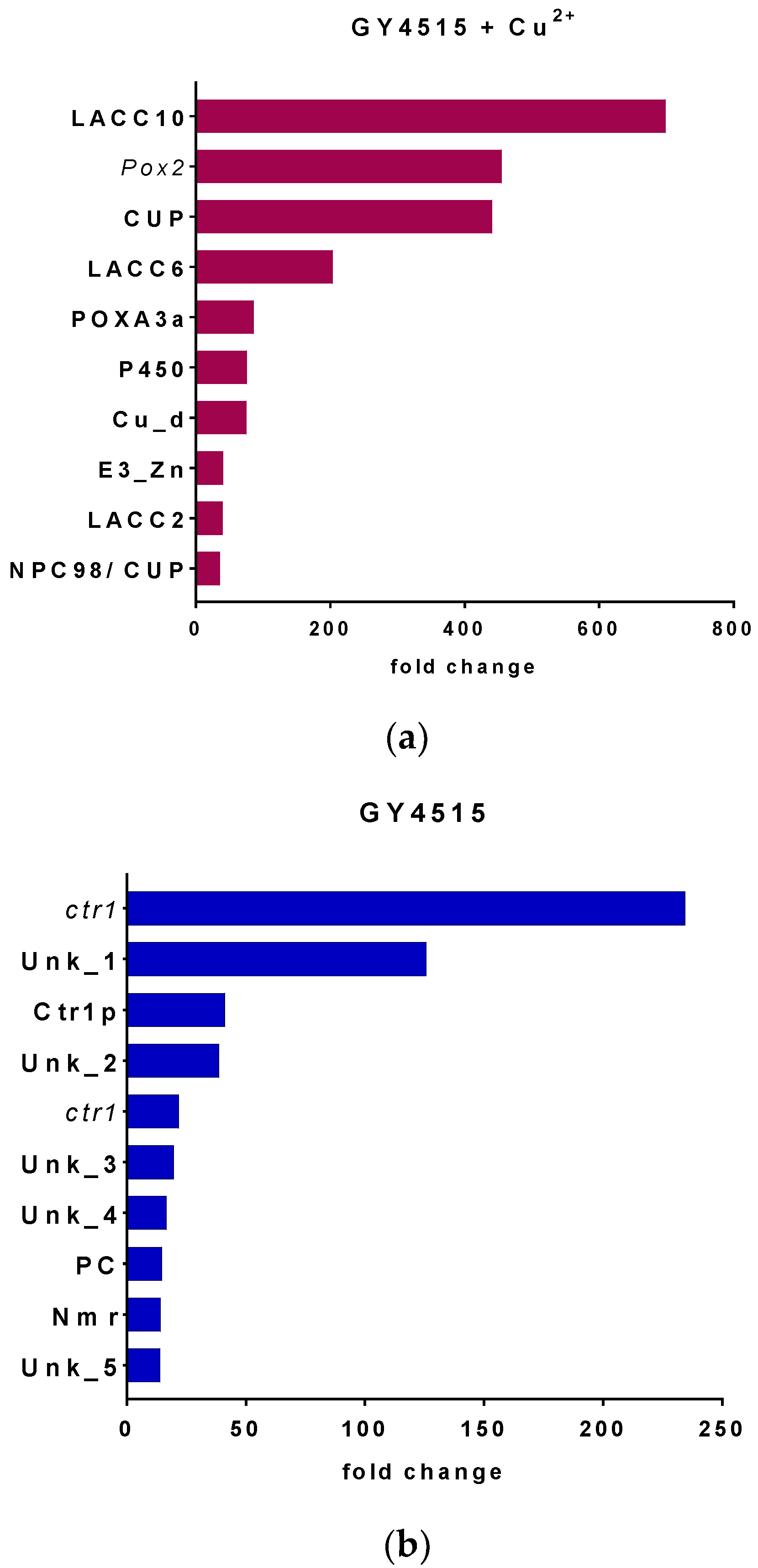
3.4. Transcriptome Analysis of P. ostreatus under Nutrient Sufficient Conditions with and without Copper Sulfate
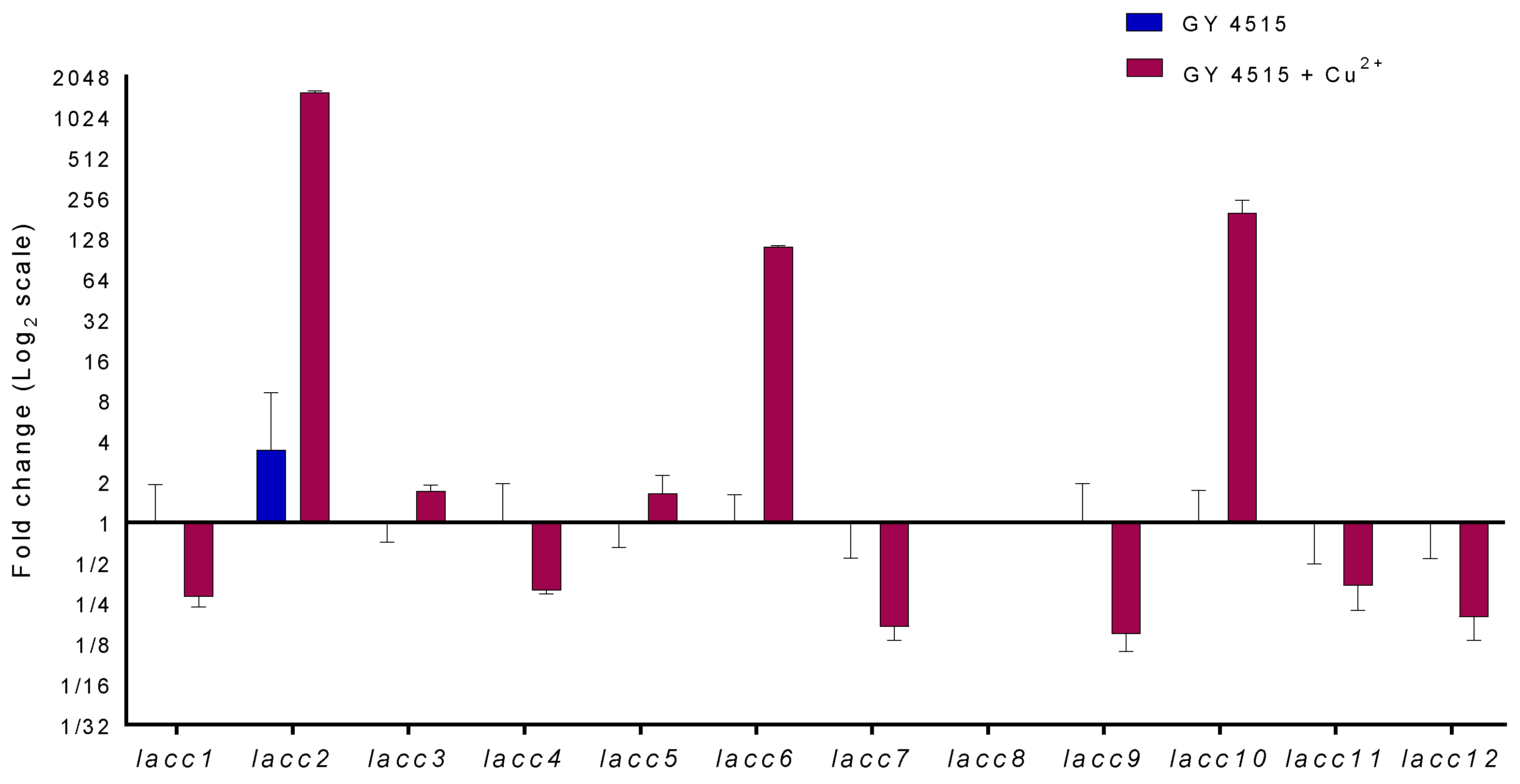
3.5. Analysis of Laccase Gene Transcripts
3.6. Effect Limited-Nutrition Conditions on Laccase Production
3.7. Hyphal Morphology of P. ostreatus under Different Nutrient Conditions with Copper
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giardina, P.; Faraco, V.; Pezzella, C.; Piscitelli, A.; Vanhulle, S.; Sannia, G. Laccases: A never-ending story. Cell. Mol. Life Sci. 2010, 67, 369–385. [Google Scholar] [CrossRef]
- Sirim, D.; Wagner, F.; Wang, L.; Schmid, R.D.; Pleiss, J. The laccase engineering database: A classification and analysis system for laccases and related multicopper oxidases. Database 2011, 2011, bar006. [Google Scholar] [CrossRef] [Green Version]
- Asemoloye, M.D.; Marchisio, M.A.; Gupta, V.K.; Pecoraro, L. Genome-based engineering of ligninolytic enzymes in fungi. Microb. Cell Factories 2021, 20, 20. [Google Scholar] [CrossRef]
- Mate, D.; Alcalde, M. Laccase: A multi-purpose biocatalyst at the forefront of biotechnology. Microb. Biotechnol. 2017, 10, 1457–1467. [Google Scholar] [CrossRef]
- Sun, K.; Li, S.; Si, Y.; Huang, Q. Advances in laccase-triggered anabolism for biotechnology applications. Crit. Rev. Biotechnol. 2021, 41, 969–993. [Google Scholar] [CrossRef]
- Fernández-Fueyo, E.; Ruiz-Dueñas, F.J.; López-Lucendo, M.F.; Pérez-Boada, M.; Rencoret, J.; Gutiérrez, A.; Pisabarro, A.G.; Ramírez, L.; Martínez, A.T. A secretomic view of woody and nonwoody lignocellulose degradation by Pleurotus ostreatus. Biotechnol. Biofuels 2016, 9, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, C. Cultivation of Pleurotus ostreatus and other edible mushrooms. Appl. Microbiol. Biotechnol. 2010, 85, 1321–1337. [Google Scholar] [CrossRef] [PubMed]
- Ohm, R.A.; Riley, R.; Salamov, A.; Min, B.; Choi, I.-G.; Grigoriev, I.V. Genomics of wood-degrading fungi. Fungal Genet. Biol. 2014, 72, 82–90. [Google Scholar] [CrossRef]
- Kues, U.; Ruhl, M. Multiple multi-copper oxidase gene families in basidiomycetes—What for? Curr. Genom. 2011, 12, 72–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanera, R.; Omarini, A.; Santoyo, F.; Pérez, G.; Pisabarro, A.G.; Ramírez, L. Non-additive transcriptional profiles underlie dikaryotic superiority in Pleurotus ostreatus laccase activity. PLoS ONE 2013, 8, e73282. [Google Scholar] [CrossRef] [PubMed]
- Pérez, G.; Pangilinan, J.; Pisabarro, A.G.; Ramírez, L. Telomere organization in the ligninolytic basidiomycete Pleurotus ostreatus. Appl. Environ. Microbiol. 2009, 75, 1427–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piscitelli, A.; Giardina, P.; Lettera, V.; Pezzella, C.; Sannia, G.; Faraco, V. Induction and transcriptional regulation of laccases in fungi. Curr. Genom. 2011, 12, 104–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzella, C.; Autore, F.; Giardina, P.; Piscitelli, A.; Sannia, G.; Faraco, V. The Pleurotus ostreatus laccase multi-gene family: Isolation and heterologous expression of new family members. Curr. Genet. 2009, 55, 45–57. [Google Scholar] [CrossRef]
- Jiao, X.; Li, G.; Wang, Y.; Nie, F.; Cheng, X.; Abdullah, M.; Lin, Y.; Cai, Y. Systematic analysis of the Pleurotus ostreatus laccase gene (PoLac) family and functional characterization of PoLac2 involved in the degradation of cotton-straw lignin. Molecules 2018, 23, 880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanera, R.; Pérez, G.; Omarini, A.; Alfaro, M.; Pisabarro, A.G.; Faraco, V.; Amore, A.; Ramírez, L. Transcriptional and enzymatic profiling of Pleurotus ostreatus laccase genes in submerged and solid-state fermentation cultures. Appl. Environ. Microbiol. 2012, 78, 4037–4045. [Google Scholar] [CrossRef] [Green Version]
- Pezzella, C.; Lettera, V.; Piscitelli, A.; Giardina, P.; Sannia, G. Transcriptional analysis of Pleurotus ostreatus laccase genes. Appl. Microbiol. Biotechnol. 2013, 97, 705–717. [Google Scholar] [CrossRef]
- Giardina, P.; Autore, F.; Faraco, V.; Festa, G.; Palmieri, G.; Piscitelli, A.; Sannia, G. Structural characterization of heterodimeric laccases from Pleurotus ostreatus. Appl. Microbiol. Biotechnol. 2007, 75, 1293–1300. [Google Scholar] [CrossRef]
- Faraco, V.; Ercole, C.; Festa, G.; Giardina, P.; Piscitelli, A.; Sannia, G. Heterologous expression of heterodimeric laccase from Pleurotus ostreatus in Kluyveromyces lactis. Appl. Microbiol. Biotechnol. 2008, 77, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Giardina, P.; Palmieri, G.; Scaloni, A.; Fontanella, B.; Faraco, V.; Cennamo, G.; Sannia, G. Protein and gene structure of a blue laccase from Pleurotus ostreatus. Biochem. J. 1999, 341, 655–663. [Google Scholar] [CrossRef]
- Macellaro, G.; Baratto, M.C.; Piscitelli, A.; Pezzella, C.; de Biani, F.F.; Palmese, A.; Piumi, F.; Record, E.; Basosi, R.; Sannia, G. Effective mutations in a high redox potential laccase from Pleurotus ostreatus. Appl. Microbiol. Biotechnol. 2014, 98, 4949–4961. [Google Scholar] [CrossRef] [PubMed]
- Aza, P.; de Salas, F.; Molpeceres, G.; Rodríguez-Escribano, D.; de la Fuente, I.; Camarero, S. Protein engineering approaches to enhance fungal laccase production in S. cerevisiae. Int. J. Mol. Sci. 2021, 22, 1157. [Google Scholar] [CrossRef]
- Maestre-Reyna, M.; Liu, W.-C.; Jeng, W.-Y.; Lee, C.-C.; Hsu, C.-A.; Wen, T.-N.; Wang, A.H.-J.; Shyur, L.-F. Structural and functional roles of glycosylation in fungal laccase from Lentinus sp. PLoS ONE 2015, 10, e0120601. [Google Scholar] [CrossRef]
- Grandes-Blanco, A.I.; Díaz-Godínez, G.; Téllez-Téllez, M.; Delgado-Macuil, R.J.; Rojas-López, M.; Bibbins-Martínez, M.D. Ligninolytic activity patterns of pleurotus ostreatus obtained by submerged fermentation in presence of 2,6-dimethoxyphenol and remazol brilliant blue R dye. Prep. Biochem. Biotechnol. 2013, 43, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Petris, M.J. The SLC31 (Ctr) copper transporter family. Pflugers Arch.-Eur. J. Physiol. 2004, 447, 752–755. [Google Scholar] [CrossRef]
- Efeyan, A.; Comb, W.C.; Sabatini, D.M. Nutrient-sensing mechanisms and pathways. Nat. Cell Biol. 2015, 517, 302–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, N.A.; Ries, L.N.A.; Goldman, G.H. How nutritional status signalling coordinates metabolism and lignocellulolytic enzyme secretion. Fungal Genet. Biol. 2014, 72, 48–63. [Google Scholar] [CrossRef]
- Adnan, M.; Zheng, W.; Islam, W.; Arif, M.; Abubakar, Y.S.; Wang, Z.; Lu, G. Carbon catabolite repression in filamentous fungi. Int. J. Mol. Sci. 2017, 19, 48. [Google Scholar] [CrossRef] [Green Version]
- Huberman, L.B.; Liu, J.; Qin, L.; Glass, N.L. Regulation of the lignocellulolytic response in filamentous fungi. Fungal Biol. Rev. 2016, 30, 101–111. [Google Scholar] [CrossRef] [Green Version]
- De Assis, L.J.; Silva, L.P.; Bayram, O.; Dowling, P.; Kniemeyer, O.; Krüger, T.; Brakhage, A.A.; Chen, Y.; Dong, L.; Tan, K.; et al. Carbon catabolite repression in filamentous fungi is regulated by phosphorylation of the transcription factor crea. mBio 2021, 12, e03146-20. [Google Scholar] [CrossRef]
- Davis, M.A.; Wong, K.H. Nitrogen metabolism in filamentous fungi. In Cellular and Molecular Biology of Filamentous Fungi; John Wiley & Sons: Hoboken, NJ, USA, 2014; pp. 325–338. [Google Scholar] [CrossRef]
- Tudzynski, B. Nitrogen regulation of fungal secondary metabolism in fungi. Front. Microbiol. 2014, 5, 656. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Qiu, M.; Wang, B.; Yin, W.-B.; Nie, X.; Qin, Q.; Ren, S.; Yang, K.; Zhang, F.; Zhuang, Z.; et al. Functional analysis of the nitrogen metabolite repression regulator gene nmrA in Aspergillus flavus. Front. Microbiol. 2016, 7, 1794. [Google Scholar] [CrossRef] [Green Version]
- Todd, R.B.; Fraser, J.A.; Wong, K.H.; Davis, M.A.; Hynes, M.J. Nuclear accumulation of the GATA factor are A in response to complete nitrogen starvation by regulation of nuclear export. Eukaryot. Cell 2005, 4, 1646–1653. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.H.; Hynes, M.J.; Todd, R.B.; Davis, M.A. Transcriptional control of nmrA by the bZIP transcription factor MeaB reveals a new level of nitrogen regulation in Aspergillus nidulans. Mol. Microbiol. 2007, 66, 534–551. [Google Scholar] [CrossRef]
- Fernandez, J.; Wright, J.D.; Hartline, D.; Quispe, C.F.; Madayiputhiya, N.; Wilson, R.A. Principles of carbon catabolite repression in the rice blast fungus: Tps1, Nmr1-3, and a MATE–family pump regulate glucose metabolism during infection. PLoS Genet. 2012, 8, e1002673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinoco-Valencia, R.; Gómez-Cruz, C.; Galindo, E.; Serrano-Carreón, L. Toward an understanding of the effects of agitation and aeration on growth and laccases production by Pleurotus ostreatus. J. Biotechnol. 2014, 177, 67–73. [Google Scholar] [CrossRef]
- Galhaup, C.; Wagner, H.; Hinterstoisser, B.; Haltrich, D. Increased production of laccase by the wood-degrading basidiomycete Trametes pubescens. Enzym. Microb. Technol. 2002, 30, 529–536. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosaiicyiic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lettera, V.; Piscitelli, A.; Leo, G.; Birolo, L.; Pezzella, C.; Sannia, G. Identification of a new member of Pleurotus ostreatus laccase family from mature fruiting body. Fungal Biol. 2010, 114, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Castanera, R.; López-Varas, L.; Pisabarro, A.G.; Ramírez, L. Validation of reference genes for transcriptional analyses in Pleurotus ostreatus by using reverse transcription-quantitative PCR. Appl. Environ. Microbiol. 2015, 81, 4120–4129. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Larraya, L.M.; Pérez, G.; Peñas, M.M.; Baars, J.J.P.; Mikosch, T.S.P.; Pisabarro, A.G.; Ramírez, L. Molecular karyotype of the white rot fungus Pleurotus ostreatus. Appl. Environ. Microbiol. 1999, 65, 3413–3417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riley, R.; Salamov, A.A.; Brown, D.W.; Nagy, L.G.; Floudas, D.; Held, B.W.; Levasseur, A.; Lombard, V.; Morin, E.; Otillar, R.; et al. Extensive sampling of basidiomycete genomes demonstrates inadequacy of the white-rot/brown-rot paradigm for wood decay fungi. Proc. Natl. Acad. Sci. USA 2014, 111, 9923–9928. [Google Scholar] [CrossRef] [Green Version]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.A.; Otillar, R.; Riley, R.; Salamov, A.A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Morgulis, A.; Coulouris, G.; Raytselis, Y.; Madden, T.L.; Agarwala, R.; Schäffer, A.A. Database indexing for production MegaBLAST searches. Bioinformatics 2008, 24, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Kachlishvili, E.; Penninckx, M.J.; Tsiklauri, N.; Elisashvili, V. Effect of nitrogen source on lignocellulolytic enzyme production by white-rot basidiomycetes under solid-state cultivation. World J. Microbiol. Biotechnol. 2006, 22, 391–397. [Google Scholar] [CrossRef]
- Stajić, M.; Persky, L.; Friesem, D.; Hadar, Y.; Wasser, S.P.; Nevo, E.; Vukojević, J. Effect of different carbon and nitrogen sources on laccase and peroxidases production by selected Pleurotus species. Enzym. Microb. Technol. 2006, 38, 65–73. [Google Scholar] [CrossRef]
- D’Agostini, É.C.; Mantovani, T.R.D.; Valle, J.; Paccola-Meirelles, L.; Colauto, N.; Linde, G.A. Low carbon/nitrogen ratio increases laccase production from basidiomycetes in solid substrate cultivation. Sci. Agric. 2011, 68, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Bao, G.; Huang, S. Optimization of laccase production in the white-rot fungus Pleurotus ostreatus (ACCC 52857) induced through yeast extract and copper. Biotechnol. Biotechnol. Equip. 2016, 30, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Karp, S.G.; Faraco, V.; Amore, A.; Letti, L.A.J.; Soccol, V.T.; Soccol, C.R. Statistical optimization of laccase production and delignification of sugarcane bagasse by Pleurotus ostreatus in solid-state fermentation. BioMed Res. Int. 2015, 2015, 181204. [Google Scholar] [CrossRef] [Green Version]
- Barelle, C.J.; Bohula, E.A.; Kron, S.; Wessels, D.; Soll, D.R.; Schäfer, A.; Brown, A.J.P.; Gow, N.A.R. Asynchronous cell cycle and asymmetric vacuolar inheritance in true hyphae of Candida albicans. Eukaryot. Cell 2003, 2, 398–410. [Google Scholar] [CrossRef] [Green Version]
- Lettera, V.; Del Vecchio, C.; Piscitelli, A.; Sannia, G. Low impact strategies to improve ligninolytic enzyme production in filamentous fungi: The case of laccase in Pleurotus ostreatus. Comptes Rendus Biol. 2011, 334, 781–788. [Google Scholar] [CrossRef]
- Periasamy, R.; Palvannan, T. Optimization of laccase production by Pleurotus ostreatus IMI 395545 using the Taguchi DOE methodology. J. Basic Microbiol. 2010, 50, 548–556. [Google Scholar] [CrossRef]
- Yang, J.; Wang, G.; Ng, T.B.; Lin, J.; Ye, X. Laccase production and differential transcription of laccase genes in Cerrena sp. in response to metal ions, aromatic compounds, and nutrients. Front. Microbiol. 2016, 6, 1558. [Google Scholar] [CrossRef]
- Hazuchová, M.; Chmelová, D.; Ondrejovič, M. The optimization of propagation medium for the increase of laccase production by the white-rot fungus Pleurotus ostreatus. Nova Biotechnol. Chim. 2017, 16, 113–123. [Google Scholar] [CrossRef]
- Baldrian, P.; Gabriel, J. Copper and cadmium increase laccase activity in Pleurotus ostreatus. FEMS Microbiol. Lett. 2002, 206, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Kannaiyan, R.; Kostenko, V.; Martinuzzi, R.J. Nutrient media optimization for simultaneous enhancement of the laccase and peroxidases production by coculture of Dichomitus squalens and Ceriporiopsis subvermispora. Biotechnol. Appl. Biochem. 2015, 62, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Kannaiyan, R.; Mahinpey, N.; Mani, T.; Martinuzzi, R.J.; Kostenko, V. Enhancement of Dichomitus squalens tolerance to copper and copper-associated laccase activity by carbon and nitrogen sources. Biochem. Eng. J. 2012, 67, 140–147. [Google Scholar] [CrossRef]
- Chenthamarakshan, A.; Parambayil, N.; Miziriya, N.; Soumya, P.S.; Lakshmi, M.S.K.; Ramgopal, A.; Dileep, A.; Nambisan, P. Optimization of laccase production from Marasmiellus palmivorus LA1 by Taguchi method of design of experiments. BMC Biotechnol. 2017, 17, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhamare, H.M.; Jadhav, H.P.; Sayyed, R.Z. Statistical optimization for enhanced production of extracellular laccase from Aspergillus sp. HB_RZ4 isolated from bark scrapping. Environ. Sustain. 2018, 1, 159–166. [Google Scholar] [CrossRef]
- Palmieri, G.; Giardina, P.; Bianco, C.; Fontanella, B.; Sannia, G. Copper induction of laccase isoenzymes in the ligninolytic fungus Pleurotus ostreatus. Appl. Environ. Microbiol. 2000, 66, 920–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, G.; Cennamo, G.; Faraco, V.; Amoresano, A.; Sannia, G.; Giardina, P. Atypical laccase isoenzymes from copper supplemented Pleurotus ostreatus cultures. Enzym. Microb. Technol. 2003, 33, 220–230. [Google Scholar] [CrossRef]
- Peñas, M.M.; Azparren, G.; Domínguez, Á.; Sommer, H.; Ramírez, L.; Pisabarro, A.G. Identification and functional characterisation of ctr1, a Pleurotus ostreatus gene coding for a copper transporter. Mol. Genet. Genom. 2005, 274, 402–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (Fw) 1 | Primer Sequence (Rv) 2 |
|---|---|---|
| lacc1 | GGTACATCCTAGCACCCAATG | GACGAGATCAGTTTCCAAGAGG |
| lacc2 | CCCTGGCAGATTGGTATCATG | ATGACAGCGTAAGGGACAAG |
| lacc3 | TCGTTTCCGTCTCGTTTCTC | CTGCGAAGATTTGGATGCTGG |
| lacc4 | CCCCATCCTTTCCATCTTCAC | TAGTTATACACCGAGCTTCCG |
| lacc5 | CGCATTTGCCGCTTTCTT | GGTGACTAGGACTGAGTATCTC |
| lacc6 | GTACAACTACGAAAACCCCG | CAAGGTCAAGATGCCAGT |
| lacc7 | GTTGATAGCCTCCAGATCTTCG | GTAGGATGGCGGAGTTGATG |
| lacc8 | CATTGGCTGTGACTCGAA | GGATCAGAGAATAGCGTTGG |
| lacc9 | CTATCCTTCGGTATGCTGGTG | ATATTGATGTCTGCGCCTCC |
| lacc10 | CCTACTTCCCCTTTGGCTATC | ATGACGAGCAAAGAGTGACC |
| lacc11 | CCTGAATGGTCTGATCTCTGC | CCTATGACTTGGGCTCTTCG |
| lacc12 | GTACTCATTTTCGGCTCCTG | CCACGTAGTCCATCGCAATA |
| sar1 | GGATAGTCTTCCTCGTCGATAG | GGGTGCGTCAATCTTGTTAC |
| gapdh1 | TGGTCCATCGCATAAGGA | ACACGGAAGGACAAACCA |
| actin1 | AGTCGGTGCCTTGGTTAT | ATACCGACCATCACACCT |
| Pep | GATTCCAGAGGACAAGGACGCAA | AAATCTTCCGCGATACGGGTCACT |
| lacc1 | GGTACATCCTAGCACCCAATG | GACGAGATCAGTTTCCAAGAGG |
| Ammonium Sulfate (gL−1) | Yeast Extract (gL−1) | Glucose (gL−1) | C:N | Biomass (gL−1) | Final Glucose (gL−1) | Max. Laccase Activity (UL−1) |
|---|---|---|---|---|---|---|
| 1.0 | 0 | 0.5 | 1:1 | 0.26 ± 0.02 | 0.18 ± 0.03 | 2.85 ± 2.31 |
| 1.0 | 0 | 1.0 | 2:1 | 0.39 ± 0.02 | 0.58 ± 0.02 | 3.40 ± 0.58 |
| 1.0 | 0 | 10.0 | 22:1 | 1.30 ± 0.18 | 6.86 ± 0.97 | 6.35 ± 3.07 |
| 10.0 | 0 | 10.0 | 2:1 | 1.18 ± 0.10 | 6.47 ± 0.51 | 1.74 ± 0.15 |
| 0 | 5.0 | 1.0 | 5:1 | 0.91 ± 0.04 | 0.06 ± 0.02 | 944 ± 71.6 |
| 0 | 5.0 | 10.0 | 14:1 | 3.62 ± 0.01 | 0.12 ± 0.04 | 489 ± 63.2 |
| 0 | 5.0 | 20.0 | 23:1 | 7.65 ± 0.08 | 1.16 ± 0.89 | 104 ± 10.1 |
| 0 | 10.0 | 20.0 | 14:1 | 7.04 ± 0.18 | 0.49 ± 0.13 | 2317 ± 787.7 |
| Culture Name 1 | Factors | Response Variables | ||||
|---|---|---|---|---|---|---|
| Without Copper | With Copper Sulfate 1 mM | |||||
| Glucose (gL−1) | YE (gL−1) | Biomass (gL−1) | Log10 MLA 2 (UL−1) | Biomass (gL−1) | Log10 MLA 2 (UL−1) | |
| GY3008 | 30.0 | 7.50 | 9.25 | 2.69 | 10.4 | 3.82 |
| GY6008 | 60.0 | 7.50 | 15.3 | 3.09 | 15.6 | 2.65 |
| GY3023 | 30.0 | 22.5 | 11.2 | 2.76 | 9.71 | 4.22 |
| GY6023 | 60.0 | 22.5 | 8.68 | 3.12 | 11.0 | 4.02 |
| GY2415 | 23.8 | 15.0 | 12.2 | 3.18 | 10.9 | 4.17 |
| GY6215 | 66.2 | 15.0 | 10.9 | 2.88 | 12.9 | 4.13 |
| GY4504 | 45.0 | 4.39 | 5.29 | 1.99 | 6.13 | 2.22 |
| GY4526 | 45.0 | 25.6 | 10.7 | 3.17 | 15.5 | 3.80 |
| GY4515 | 45.0 | 15.0 | 16.6 | 3.11 | 22.4 | 4.50 |
| GY4515 | 45.0 | 15.0 | 14.5 | 3.06 | 19.8 | 4.47 |
| GY4515 | 45.0 | 15.0 | 16.9 | 3.12 | 19.4 | 4.48 |
| GY4515 | 45.0 | 15.0 | 19.5 | 3.03 | 19.1 | 4.40 |
| GY4515 | 45.0 | 15.0 | 16.5 | 2.99 | 19.0 | 4.41 |
| JGI PC15 V2.0 Transcript Id. | JGI Annotations | Short Name | JGI PC15 V2.0 Transcript Id. | JGI Annotations | Short Name |
|---|---|---|---|---|---|
| GY4515 + Cu2+ | GY4515 | ||||
| 1089723 | Multicopper oxidase, laccase, Lacc10 (PoxC) | Lacc10 | 1095975 1 | Unannotated gene Similar to ctr1 gene for copper transporter, exons 1–3 Pleurotus sp. “Florida” | ctr1 |
| 1105204 1 | Unannotated gen. Similar to Phenol oxidase (pox2) gene | Pox2 | 159791 | Unknown protein | Unk_1 |
| 1097654 | Cupredoxin domin | Cup | 1092022 | Copper transporter, Ctr1p | Ctr1p |
| 1113032 | Multi-copper oxidase Laccase, Lacc6 (PoxA1b) | Lacc6 | 1086646 | Unknown protein | Unk_2 |
| 1067572 | Small subunit of laccase PoxA3a | PoxA3a | 1088435 1 | Unannotated gene Similar to ctr1 gene for copper transporter, exons 1–3 Pleurotus sp. “Florida” | ctr1 |
| 1063469 | Cytochrome P450 CYP2 subfamily | P450 | 1090041 | Unknown protein | Unk_3 |
| 1087630 | Blue (type 1) copper domain | Cu-d | 1090781 | Unknown protein | Unk_4 |
| 1105457 | Predicted E3 ubiquitin ligase/Zinc finger, C3HC4 type | E3_Zn | 1099858 | Polyketide cyclase | PC |
| 1067328 | Multi-copper oxidases, Lacc2 | Lacc2 | 171939 | NmrA-like family | Nmr |
| 1062660 | Nuclear pore complex, Nup98 component (sc Nup145/Nup100/Nup116), and Cupredoxin domain | NPC98/Cup | 1077411 | Unknown protein | Unk_5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durán-Sequeda, D.; Suspes, D.; Maestre, E.; Alfaro, M.; Perez, G.; Ramírez, L.; Pisabarro, A.G.; Sierra, R. Effect of Nutritional Factors and Copper on the Regulation of Laccase Enzyme Production in Pleurotus ostreatus. J. Fungi 2022, 8, 7. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8010007
Durán-Sequeda D, Suspes D, Maestre E, Alfaro M, Perez G, Ramírez L, Pisabarro AG, Sierra R. Effect of Nutritional Factors and Copper on the Regulation of Laccase Enzyme Production in Pleurotus ostreatus. Journal of Fungi. 2022; 8(1):7. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8010007
Chicago/Turabian StyleDurán-Sequeda, Dinary, Daniela Suspes, Estibenson Maestre, Manuel Alfaro, Gumer Perez, Lucía Ramírez, Antonio G. Pisabarro, and Rocío Sierra. 2022. "Effect of Nutritional Factors and Copper on the Regulation of Laccase Enzyme Production in Pleurotus ostreatus" Journal of Fungi 8, no. 1: 7. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8010007