
Genomic Characterization of the Titan-like Cell Producing Naganishia tulchinskyi, the First Novel Eukaryote Isolated from the International Space Station

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation of Microorganisms
2.2. Whole Genome Sequencing Analyses
2.3. Phylogenetic Analyses
2.4. Morphological Characterization under Simulated Microgravity Conditions
2.4.1. Growth Conditions for Phenotypic Analyses under Different Conditions
2.4.2. Confocal Microscopy
2.4.3. Scanning Electron Microscopy
2.4.4. Transmission Electron Microscopy
2.4.5. Statistical Analyses
2.5. Survey of Genes and Proteins Involved in Resistance
2.6. De Novo Genome Annotation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functions | GenBank# | Enzyme, Protein, Other Biomolecules | Naganishia tulchinskyi | Naganishia vishniacii | Papiliotrema laurentii | S. cerevisiae | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| IF6SW-B1T | 1F7SW-B1 | 1F1SW-F1 | IIF5SW-F1 | CBS 10616 | IF7SW-B5 | IF7SW-F4 | S288C | ||||
| Pigment Biosynthesis | AAY33922 | Geranylgeranyl diphosphate synthase | * | ** | ** | ** | ** | * | ** | ** | [39] |
| ACI43098 | Cytochrome P450 reductase | ** | ** | ** | ** | ** | ** | ** | ** | [40] | |
| AHW57996 | Farnesyl pyrophosphate synthase | ** | ** | ** | ** | ** | ** | ** | ** | [41] | |
| BAA33979 | Isopentenyl-diphosphate delta isomerase | ** | ** | ** | ** | ** | ** | ** | ** | [42] | |
| NP_009555 | Alpha, alpha-trehalase NTH2 | ** | ** | ** | ** | ** | ** | ** | ** | [43] | |
| NP_010284 | Alpha, alpha-trehalase NTH1 | ** | ** | ** | ** | ** | ** | ** | ** | [44] | |
| NP_015351 | Alpha, alpha-trehalase ATH1 | ** | [43] | ||||||||
| NP_594975 | Putative trehalose-phosphate synthase Tps2 | ** | ** | * | ** | ** | ** | ** | ** | [45] | |
| AAO47570 | Phytoene-beta carotene synthase | ** | ** | ** | ** | * | * | * | [46] | ||
| Radio- resistance | RAD51 | Recombinase RAD51 | ** | ** | ** | ** | ** | ** | ** | ** | [47] |
| RAD54 | DNA repair and recombination protein RAD54-like protein | ** | ** | ** | ** | ** | ** | ** | ** | [47] | |
| RDH54 | DNA-dependent ATPase RDH54 | ** | ** | ** | ** | ** | ** | ** | ** | [47] | |
| CAA54491 | RAD9 | * | * | [48] | |||||||
| CAA54492 | RAD9 | * | [48] | ||||||||
| Microgravity tolerance | 1HT3_A | Chain A, Proteinase K | ** | ** | ** | ** | ** | ** | ** | ** | [49] |
| 1IC6_A | Chain A, Proteinase K | ** | ** | ** | ** | ** | ** | ** | ** | [50] | |
| 4LHK_A | Chain A, Flocculin | ** | [51] | ||||||||
| 4LHK_B | Chain B, Flocculin | ** | [51] | ||||||||
| 4LHN_A | Chain A, Flocculation protein FLO1 | ** | [51] | ||||||||
| P26306 | DNA repair protein rad9 | * | * | [52] | |||||||
| P38844 | Daughter-specific expression protein 2 | ** | [53] | ||||||||
| P40077 | Daughter-specific expression protein 1 | ** | [54] | ||||||||
| P48013 | DNA repair protein rad9 | * | [48] | ||||||||
| Q59Y31 | Flocculation protein 1 | ** | [55] | ||||||||
| Name | Origin * | # of Sequences | Date of Access | Data Set |
|---|---|---|---|---|
| Naganishia tulchinskyi (n = 4) | This study | Assembly | ||
| Naganishia cerealis | UniProt | 3 | 4/5/21 | Protein sequences |
| Naganishia diffluens | UniProt | 7 | 4/5/21 | Protein sequences |
| Naganishia liquefaciens | UniProt | 4 | 4/5/21 | Protein sequences |
| Naganishia sp. | UniProt | 3 | 4/5/21 | Protein sequences |
| Cryptococcus neoformans var grubii | UniProt | 65 | 4/5/21 | Protein sequences |
| Cryptococcus neoformans var neoformans | UniProt | 370 | 4/5/21 | Protein sequences |
| Cryptococcus neoformans var grubii | ESTdb | 69 | 3/31/21 | Transcriptome |
| Cryptococcus neoformans var neoformans | ESTdb | 74,724 | 3/31/21 | Transcriptome |
| Cryptococcus vishniacci (Naganishia vishniacii) | ESTdb | 37 | 3/31/21 | Transcriptome |
| Papiliotrema laurentii (n = 2) | This study | Assembly |
2.7. Functional Annotation
2.8. Data Availability
3. Results and Discussion
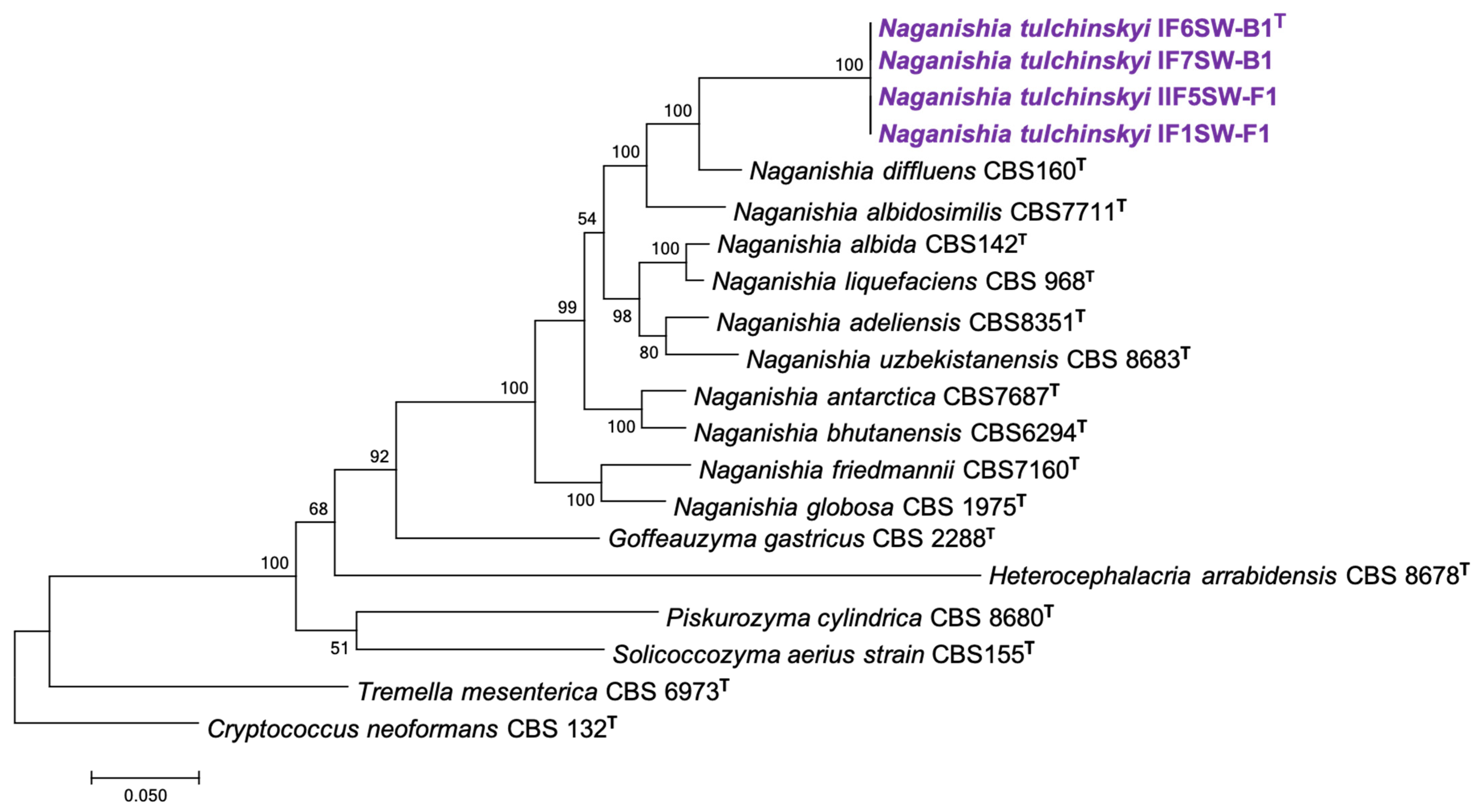
3.1. Phylogenetic Analyses of Novel ISS Strains
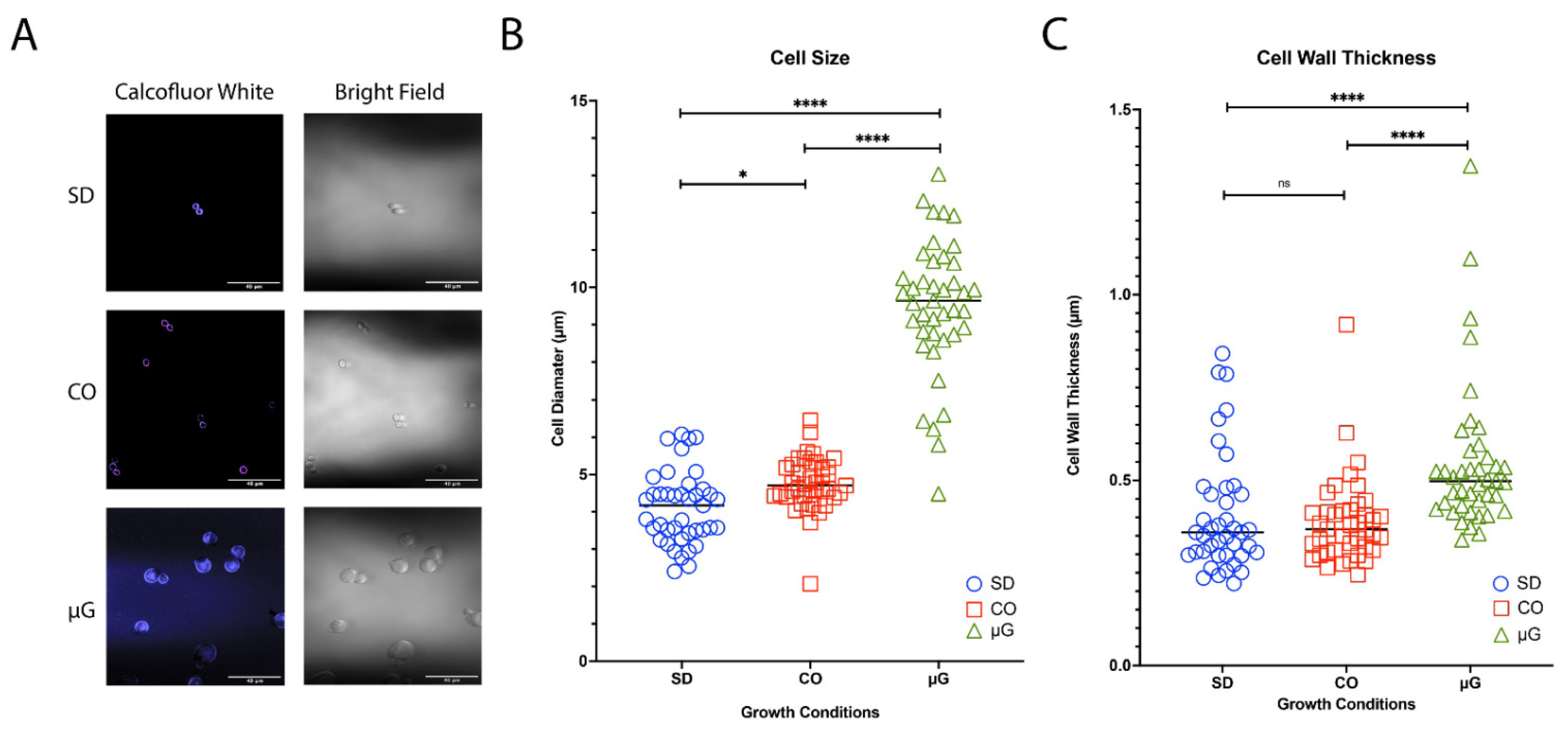
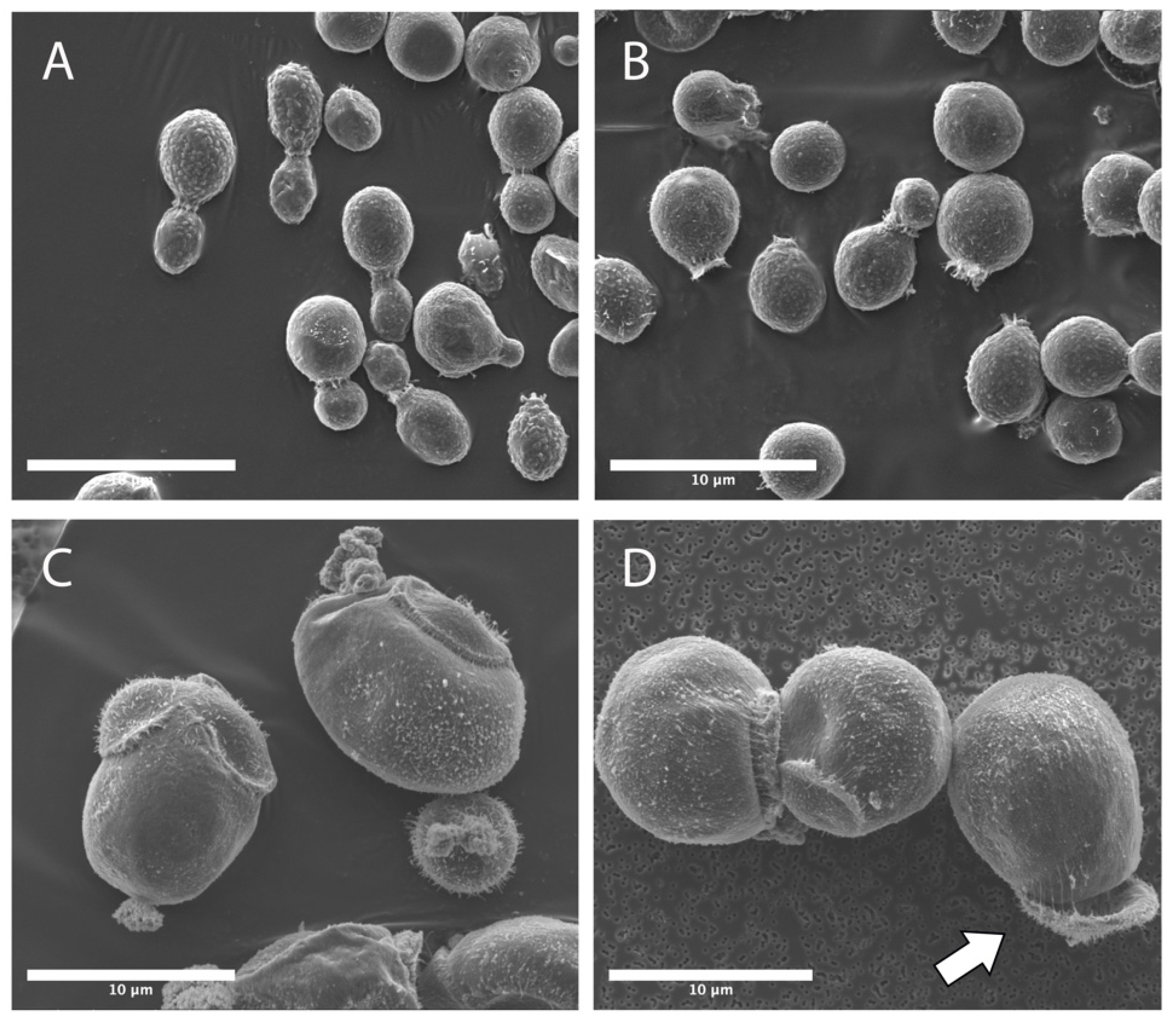
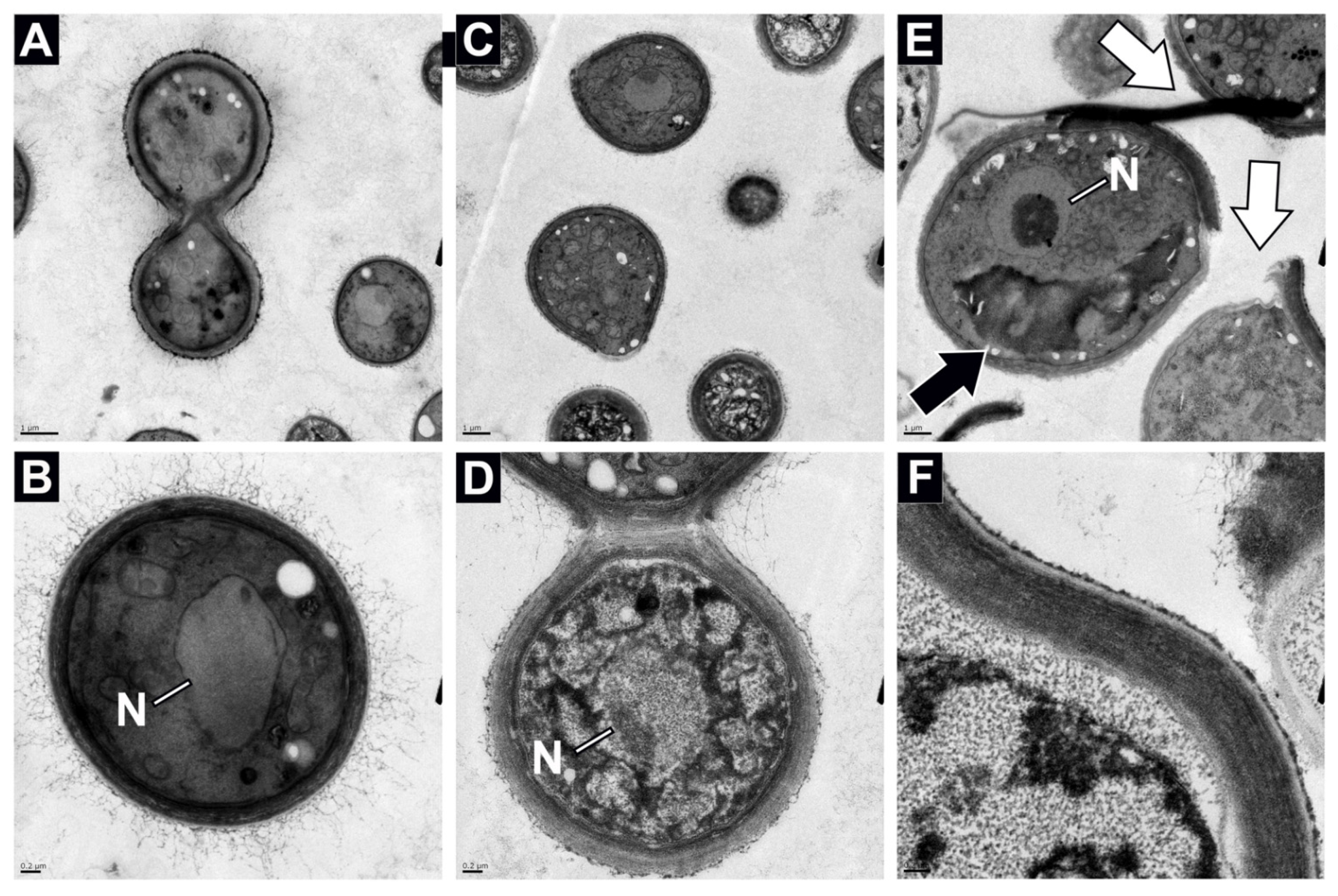
3.2. Titan-like Cell Characterization of Novel ISS Strains
3.3. Genomic Analyses of the Novel ISS Strain
3.4. Taxonomy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fotedar, R.; Kolecka, A.; Boekhout, T.; Fell, J.W.; Anand, A.; Al Malaki, A.; Zeyara, A.; Al Marri, M. Naganishia qatarensis sp. nov., a novel basidiomycetous yeast species from a hypersaline marine environment in Qatar. Int. J. Syst. Evol. Microbiol. 2018, 68, 2924–2929. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Z.; Wang, Q.M.; Theelen, B.; Groenewald, M.; Bai, F.Y.; Boekhout, T. Phylogeny of tremellomycetous yeasts and related dimorphic and filamentous basidiomycetes reconstructed from multiple gene sequence analyses. Stud. Mycol. 2015, 81, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fell, J.W.; Boekhout, T.; Fonseca, A.; Scorzetti, G.; Statzell-Tallman, A. Biodiversity and systematics of basidiomycetous yeasts as determined by large-subunit rDNA D1/D2 domain sequence analysis. Int. J. Syst. Evol. Microbiol. 2000, 50, 1351–1371. [Google Scholar] [CrossRef] [PubMed]
- Scorzetti, G.; Fell, J.W.; Fonseca, A.; Statzell-Tallman, A. Systematics of basidiomycetous yeasts: A comparison of large subunit D1/D2 and internal transcribed spacer rDNA regions. FEMS Yeast Res. 2002, 2, 495–517. [Google Scholar] [CrossRef]
- Sielaff, A.C.; Urbaniak, C.; Mohan, G.B.M.; Stepanov, V.G.; Tran, Q.; Wood, J.M.; Minich, J.; McDonald, D.; Mayer, T.; Knight, R.; et al. Characterization of the total and viable bacterial and fungal communities associated with the International Space Station surfaces. Microbiome 2019, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Bijlani, S.; Singh, N.K.; Mason, C.E.; Wang, C.C.C.; Venkateswaran, K. Draft Genome Sequences of Tremellomycetes Strains Isolated from the International Space Station. Microbiol. Resour. Announc. 2020, 9, e00504-20. [Google Scholar] [CrossRef]
- Goto, S.; Sugiyama, J.; Iizuka, H. A Taxonomic Study of Antarctic Yeasts. Mycologia 1969, 61, 748–774. [Google Scholar] [CrossRef]
- Vishniac, H.S.; Kurtzman, C.P. Cryptococcus antarcticus sp. nov. and Cryptococcus albidosimilis sp. nov., Basidioblastomycetes from Antarctic Soils. Int. J. Syst. Evol. Microbiol. 1992, 42, 547–553. [Google Scholar] [CrossRef] [Green Version]
- Vishniac, H.S. Cryptococcus friedmannii, a New Species of Yeast from the Antarctic. Mycologia 1985, 77, 149–153. [Google Scholar] [CrossRef]
- Vishniac, H.S.; Hempeling, W.P. Cryptococcus vishniacii sp. nov., an Antarctic Yeast. Int. J. Syst. Evol. Microbiol. 1979, 29, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Turchetti, B.; Selbmann, L.; Blanchette, R.A.; Di Mauro, S.; Marchegiani, E.; Zucconi, L.; Arenz, B.E.; Buzzini, P. Cryptococcus vaughanmartiniae sp. nov. and Cryptococcus onofrii sp. nov.: Two new species isolated from worldwide cold environments. Extremophiles 2015, 19, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Boekhout, T.; Bandoni, R.J.; Fell, J.W.; Kwon-Chung, K.J. Chapter 74—Discussion of teleomorphic and anamorphic genera of heterobasidiomycetous yeasts. In The Yeasts, 4th ed.; Kurtzman, C.P., Fell, J.W., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; pp. 609–625. [Google Scholar]
- Fonseca, Á.; Boekhout, T.; Fell, J.W. Chapter 138—Cryptococcus Vuillemin (1901). In The Yeasts, 4th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: London, UK, 2011; pp. 1661–1737. [Google Scholar]
- Khan, Z.U.; Ahmad, S.; Hagen, F.; Fell, J.W.; Kowshik, T.; Chandy, R.; Boekhout, T. Cryptococcus randhawai sp. nov., a novel anamorphic basidiomycetous yeast isolated from tree trunk hollow of Ficus religiosa (peepal tree) from New Delhi, India. Antonie Leeuwenhoek 2010, 97, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Turchetti, B.; Buzzini, P.; Goretti, M.; Branda, E.; Diolaiuti, G.; D’Agata, C.; Smiraglia, C.; Vaughan-Martini, A. Psychrophilic yeasts in glacial environments of Alpine glaciers. FEMS Microbiol. Ecol. 2008, 63, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passoth, V.; Andersson, A.C.; Olstorpe, M.; Theelen, B.; Boekhout, T.; Schnürer, J. Cryptococcus cerealis sp. nov. a psychrophilic yeast species isolated from fermented cereals. Antonie Leeuwenhoek 2009, 96, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Furman-Kuklinska, K.; Naumnik, B.; Mysliwiec, M. Fungaemia due to Cryptococcus laurentii as a complication of immunosuppressive therapy—A case report. Adv. Med. Sci. 2009, 54, 116–119. [Google Scholar] [CrossRef] [Green Version]
- Rimek, D.; Haase, G.; Lück, A.; Casper, J.; Podbielski, A. First Report of a Case of Meningitis Caused by Cryptococcus adeliensis in a Patient with Acute Myeloid Leukemia. J. Clin. Microbiol. 2004, 42, 481–483. [Google Scholar] [CrossRef] [Green Version]
- Gerstein, A.C.; Fu, M.S.; Mukaremera, L.; Li, Z.; Ormerod, K.L.; Fraser, J.A.; Berman, J.; Nielsen, K. Polyploid titan cells produce haploid and aneuploid progeny to promote stress adaptation. mBio 2015, 6, e01340-15. [Google Scholar] [CrossRef] [Green Version]
- Dambuza, I.M.; Drake, T.; Chapuis, A.; Zhou, X.; Correia, J.; Taylor-Smith, L.; LeGrave, N.; Rasmussen, T.; Fisher, M.C.; Bicanic, T.; et al. The Cryptococcus neoformans Titan cell is an inducible and regulated morphotype underlying pathogenesis. PLoS Pathog. 2018, 14, e1006978. [Google Scholar] [CrossRef] [Green Version]
- Dyląg, M.; Colon-Reyes, R.J.; Kozubowski, L. Titan cell formation is unique to Cryptococcus species complex. Virulence 2020, 11, 719–729. [Google Scholar] [CrossRef]
- Checinska, A.; Probst, A.J.; Vaishampayan, P.; White, J.R.; Kumar, D.; Stepanov, V.G.; Fox, G.E.; Nilsson, H.R.; Pierson, D.L.; Perry, J.; et al. Microbiomes of the dust particles collected from the International Space Station and Spacecraft Assembly Facilities. Microbiome 2015, 3, 50. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hommel, B.; Mukaremera, L.; Cordero, R.J.B.; Coelho, C.; Desjardins, C.A.; Sturny-Leclère, A.; Janbon, G.; Perfect, J.R.; Fraser, J.A.; Casadevall, A.; et al. Titan cells formation in Cryptococcus neoformans is finely tuned by environmental conditions and modulated by positive and negative genetic regulators. PLoS Pathog. 2018, 14, e1006982. [Google Scholar] [CrossRef] [Green Version]
- Greetham, D.; Wimalasena, T.; Kerruish, D.W.; Brindley, S.; Ibbett, R.N.; Linforth, R.L.; Tucker, G.; Phister, T.G.; Smart, K.A. Development of a phenotypic assay for characterisation of ethanologenic yeast strain sensitivity to inhibitors released from lignocellulosic feedstocks. J. Ind. Microbiol. Biotechnol. 2014, 41, 931–945. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Nizovoy, P.; Bellora, N.; Haridas, S.; Sun, H.; Daum, C.; Barry, K.; Grigoriev, I.V.; Libkind, D.; Connell, L.B.; Moline, M. Unique genomic traits for cold adaptation in Naganishia vishniacii, a polyextremophile yeast isolated from Antarctica. FEMS Yeast Res. 2021, 21, foaa056. [Google Scholar] [CrossRef]
- Bellora, N.; Moliné, M.; David-Palma, M.; Coelho, M.A.; Hittinger, C.T.; Sampaio, J.P.; Gonçalves, P.; Libkind, D. Comparative genomics provides new insights into the diversity, physiology, and sexuality of the only industrially exploited tremellomycete: Phaffia rhodozyma. BMC Genom. 2016, 17, 901. [Google Scholar] [CrossRef] [Green Version]
- Cantarel, B.L.; Korf, I.; Robb, S.M.; Parra, G.; Ross, E.; Moore, B.; Holt, C.; Alvarado, A.S.; Yandell, M. MAKER: An easy-to-use annotation pipeline designed for emerging model organism genomes. Genome Res. 2008, 18, 188–196. [Google Scholar] [CrossRef] [Green Version]
- UniProt. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Smit, R.H.A. RepeatModeler. Available online: http://www.repeatmasker.org/RepeatModeler/ (accessed on 5 December 2021).
- Chen, M.M.; Lin, H.; Chiang, L.M.; Childers, C.P.; Poelchau, M.F. The GFF3toolkit: QC and Merge Pipeline for Genome Annotation. Methods Mol. Biol. 2019, 1858, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Korf, I. Gene finding in novel genomes. BMC Bioinform. 2004, 5, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanke, M.; Keller, O.; Gunduz, I.; Hayes, A.; Waack, S.; Morgenstern, B. AUGUSTUS: Ab initio prediction of alternative transcripts. Nucleic Acids Res. 2006, 34, W435–W439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seppey, M.; Manni, M.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness. Methods Mol. Biol. 2019, 1962, 227–245. [Google Scholar] [CrossRef] [PubMed]
- Card, D. Maker_Genome_Annotation.md. Available online: https://gist.github.com/darencard/bb1001ac1532dd4225b030cf0cd61ce2 (accessed on 5 December 2021).
- Lodato, P.; Alcaíno, J.; Barahona, S.; Niklitschek, M.; Carmona, M.; Wozniak, A.; Baeza, M.; Jiménez, A.; Cifuentes, V. Expression of the carotenoid biosynthesis genes in Xanthophyllomyces dendrorhous. Biol. Res. 2007, 40, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Alcaino, J.; Barahona, S.; Carmona, M.; Lozano, C.; Marcoleta, A.; Niklitschek, M.; Sepulveda, D.; Baeza, M.; Cifuentes, V. Cloning of the cytochrome p450 reductase (crtR) gene and its involvement in the astaxanthin biosynthesis of Xanthophyllomyces dendrorhous. BMC Microbiol. 2008, 8, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcaíno, J.; Romero, I.; Niklitschek, M.; Sepúlveda, D.; Rojas, M.C.; Baeza, M.; Cifuentes, V. Functional characterization of the Xanthophyllomyces dendrorhous farnesyl pyrophosphate synthase and geranylgeranyl pyrophosphate synthase encoding genes that are involved in the synthesis of isoprenoid precursors. PLoS ONE 2014, 9, e96626. [Google Scholar]
- Kajiwara, S.; Fraser, P.D.; Kondo, K.; Misawa, N. Expression of an exogenous isopentenyl diphosphate isomerase gene enhances isoprenoid biosynthesis in Escherichia coli. Biochem. J. 1997, 324, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Goffeau, A.; Barrell, B.G.; Bussey, H.; Davis, R.W.; Dujon, B.; Feldmann, H.; Galibert, F.; Hoheisel, J.D.; Jacq, C.; Johnston, M.; et al. Life with 6000 genes. Science 1996, 274, 546–567. [Google Scholar] [CrossRef] [Green Version]
- Jacq, C.; Alt-Mörbe, J.; Andre, B.; Arnold, W.; Bahr, A.; Ballesta, J.P.; Bargues, M.; Baron, L.; Becker, A.; Biteau, N.; et al. The nucleotide sequence of Saccharomyces cerevisiae chromosome IV. Nature 1997, 387, 75–78. [Google Scholar] [CrossRef]
- Wood, V.; Gwilliam, R.; Rajandream, M.A.; Lyne, M.; Lyne, R.; Stewart, A.; Sgouros, J.; Peat, N.; Hayles, J.; Baker, S.; et al. The genome sequence of Schizosaccharomyces pombe. Nature 2002, 415, 871–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodato, P.; Alcaino, J.; Barahona, S.; Retamales, P.; Cifuentes, V. Alternative splicing of transcripts from crtI and crtYB genes of Xanthophyllomyces dendrorhous. Appl. Environ. Microbiol. 2003, 69, 4676–4682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, K.-W.; Yang, D.-H.; Kim, M.-K.; Seo, H.S.; Lim, S.; Bahn, Y.-S. Unraveling Fungal Radiation Resistance Regulatory Networks through the Genome-Wide Transcriptome and Genetic Analyses of Cryptococcus neoformans. mBio 2016, 7, e01483-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lieberman, H.B.; Hopkins, K.M. Schizosaccharomyces malidevorans and Sz. octosporus homologues of Sz. pombe rad9, a gene that mediates radioresistance and cell-cycle progression. Gene 1994, 150, 281–286. [Google Scholar] [CrossRef]
- Gourinath, S.; Degenhardt, M.; Eschenburg, S.; Moore, K.; Delucas, L.J.; Betzel, C.; Singh, T.P. Mercury induced modifications in the stereochemistry of the active site through Cys-73 in a serine protease—Crystal structure of the complex of a partially modified proteinase K with mercury at 1.8 A resolution. Indian J. Biochem. Biophys. 2001, 38, 298–302. [Google Scholar]
- Betzel, C.; Gourinath, S.; Kumar, P.; Kaur, P.; Perbandt, M.; Eschenburg, S.; Singh, T.P. Structure of a serine protease proteinase K from Tritirachium album limber at 0.98 A resolution. Biochemistry 2001, 40, 3080–3088. [Google Scholar] [CrossRef]
- Ielasi, F.S.; Goyal, P.; Sleutel, M.; Wohlkonig, A.; Willaert, R.G. The mannose-specific lectin domains of Flo1p from Saccharomyces cerevisiae and Lg-Flo1p from S. pastorianus: Crystallization and preliminary X-ray diffraction analysis of the adhesin-carbohydrate complexes. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2013, 69, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Murray, J.M.; Carr, A.M.; Lehmann, A.R.; Watts, F.Z. Cloning and characterisation of the rad9 DNA repair gene from Schizosaccharomyces pombe. Nucleic Acids Res. 1991, 19, 3525–3531. [Google Scholar] [CrossRef] [Green Version]
- Johnston, M.; Andrews, S.; Brinkman, R.; Cooper, J.; Ding, H.; Dover, J.; Du, Z.; Favello, A.; Fulton, L.; Gattung, S. Complete nucleotide sequence of Saccharomyces cerevisiae chromosome VIII. Science 1994, 265, 2077–2082. [Google Scholar] [CrossRef]
- Diederich, B.; Wilkinson, J.F.; Magnin, T.; Najafi, S.M.A.; Errington, J.; Yudkin, M. Role of interactions between SpoIIAA and SpoIIAB in regulating cell-specific transcription factor sF of B. subtilis. Genes Dev. 1994, 8, 2653–2663. [Google Scholar] [CrossRef] [Green Version]
- Jones, T.; Federspiel, N.A.; Chibana, H.; Dungan, J.; Kalman, S.; Magee, B.B.; Newport, G.; Thorstenson, Y.R.; Agabian, N.; Magee, P.T.; et al. The diploid genome sequence of Candida albicans. Proc. Natl. Acad. Sci. USA 2004, 101, 7329–7334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.K.; Wood, J.M.; Karouia, F.; Venkateswaran, K. Succession and persistence of microbial communities and antimicrobial resistance genes associated with International Space Station environmental surfaces. Microbiome 2018, 6, 214. [Google Scholar] [CrossRef] [PubMed]
- Shanks, R.M.; Stella, N.A.; Kalivoda, E.J.; Doe, M.R.; O’Dee, D.M.; Lathrop, K.L.; Guo, F.L.; Nau, G.J. A Serratia marcescens OxyR homolog mediates surface attachment and biofilm formation. J. Bacteriol. 2007, 189, 7262–7272. [Google Scholar] [CrossRef] [Green Version]
- Schembri, M.A.; Blom, J.; Krogfelt, K.A.; Klemm, P. Capsule and fimbria interaction in Klebsiella pneumoniae. Infect. Immun. 2005, 73, 4626–4633. [Google Scholar] [CrossRef] [Green Version]
- Li, S.C.; Kane, P.M. The yeast lysosome-like vacuole: Endpoint and crossroads. Biochim. Biophys. Acta 2009, 1793, 650–663. [Google Scholar] [CrossRef] [Green Version]
- Thumm, M. Structure and function of the yeast vacuole and its role in autophagy. Microsc. Res. Tech. 2000, 51, 563–572. [Google Scholar] [CrossRef]
- Weisman, L.S. Yeast vacuole inheritance and dynamics. Annu. Rev. Genet. 2003, 37, 435–460. [Google Scholar] [CrossRef]
- Okagaki, L.H.; Strain, A.K.; Nielsen, J.N.; Charlier, C.; Baltes, N.J.; Chrétien, F.; Heitman, J.; Dromer, F.; Nielsen, K. Cryptococcal cell morphology affects host cell interactions and pathogenicity. PLoS Pathog. 2010, 6, e1000953. [Google Scholar] [CrossRef]
- Zaragoza, O.; Nielsen, K. Titan cells in Cryptococcus neoformans: Cells with a giant impact. Curr. Opin. Microbiol. 2013, 16, 409–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crabtree, J.N.; Okagaki, L.H.; Wiesner, D.L.; Strain, A.K.; Nielsen, J.N.; Nielsen, K. Titan cell production enhances the virulence of Cryptococcus neoformans. Infect. Immun. 2012, 80, 3776–3785. [Google Scholar] [CrossRef] [Green Version]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. InterProScan: Protein domains identifier. Nucleic Acids Res. 2005, 33, W116–W120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaragoza, O.; García-Rodas, R.; Nosanchuk, J.D.; Cuenca-Estrella, M.; Rodríguez-Tudela, J.L.; Casadevall, A. Fungal Cell Gigantism during Mammalian Infection. PLOS Pathog. 2010, 6, e1000945. [Google Scholar] [CrossRef]
- Han, Y.-W.; Kajitani, R.; Morimoto, H.; Palihati, M.; Kurokawa, Y.; Ryusui, R.; Argunhan, B.; Tsubouchi, H.; Abe, F.; Kajiwara, S.; et al. Draft Genome Sequence of Naganishia liquefaciens Strain N6, Isolated from the Japan Trench. Microbiol. Resour. Announc. 2020, 9, e00827-20. [Google Scholar] [CrossRef] [PubMed]
- Vajpeyi, S.; Chandran, K. Draft Genome Sequence of the Oleaginous Yeast Cryptococcus albidus var. albidus. Genome Announc. 2016, 4, e00390-16. [Google Scholar] [CrossRef] [Green Version]
- Giaever, G.; Chu, A.M.; Ni, L.; Connelly, C.; Riles, L.; Véronneau, S.; Dow, S.; Lucau-Danila, A.; Anderson, K.; André, B.; et al. Functional profiling of the Saccharomyces cerevisiae genome. Nature 2002, 418, 387–391. [Google Scholar] [CrossRef]
- Nichols, H.L.; Zhang, N.; Wen, X. Proteomics and genomics of microgravity. Physiol. Genom. 2006, 26, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheehan, K.B.; McInnerney, K.; Purevdorj-Gage, B.; Altenburg, S.D.; Hyman, L.E. Yeast genomic expression patterns in response to low-shear modeled microgravity. BMC Genom. 2007, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Seel, W.; Baust, D.; Sons, D.; Albers, M.; Etzbach, L.; Fuss, J.; Lipski, A. Carotenoids are used as regulators for membrane fluidity by Staphylococcus xylosus. Sci. Rep. 2020, 10, 330. [Google Scholar] [CrossRef] [PubMed]
- Dieser, M.; Greenwood, M.; Foreman, C.M. Carotenoid Pigmentation in Antarctic Heterotrophic Bacteria as a Strategy to Withstand Environmental Stresses. Arct. Antarct. Alp. Res. 2010, 42, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Slade, D.; Radman, M. Oxidative Stress Resistance in Deinococcus radiodurans. Microbiol. Mol. Biol. Rev. 2011, 75, 133–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulino-Lima, I.G.; Janot-Pacheco, E.; Galante, D.; Cockell, C.; Olsson-Francis, K.; Brucato, J.R.; Baratta, G.A.; Strazzulla, G.; Merrigan, T.; McCullough, R.; et al. Survival of Deinococcus radiodurans against laboratory-simulated solar wind charged particles. Astrobiology 2011, 11, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Stahl, W. Carotenoids and UV protection. Photochem. Photobiol. Sci. 2004, 3, 749–752. [Google Scholar] [CrossRef]
- Rapoport, A.; Guzhova, I.; Bernetti, L.; Buzzini, P.; Kieliszek, M.; Kot, A.M. Carotenoids and Some Other Pigments from Fungi and Yeasts. Metabolites 2021, 11, 92. [Google Scholar] [CrossRef]
- Miao, L.; Chi, S.; Wu, M.; Liu, Z.; Li, Y. Deregulation of phytoene-β-carotene synthase results in derepression of astaxanthin synthesis at high glucose concentration in Phaffia rhodozyma astaxanthin-overproducing strain MK19. BMC Microbiol. 2019, 19, 133. [Google Scholar] [CrossRef]





| Carbon Source | Naganishia tulchinskyi (This Study) | N. vaughanmartiniaea | N. onofriia | N. saitoib | N. friedmanniib | N. qatarensisc | N. cerealisd | N. randhawaee | N. globosab | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IF6SW-B1 | IF7SW-B1 | IF1SW-F1 | IIF5SW-F1 | ||||||||||
| Growth at: | |||||||||||||
| 4 °C | + | + | + | + | + | + | NT | + | NT | NT | NT | NT | |
| 25 °C | + | + | + | + | + | + | + | - | + | + | + | + | |
| 30 °C | + | + | + | + | - | - | + | - | + | + | + | + | |
| 35 °C | - | - | - | - | - | - | - | - | + | - | - | - | |
| Differential characteristics among various Naganishia species | |||||||||||||
| D-Arabinose | + | + | + | + | + | + | - | - | + | + | w | - | |
| D-Ribose | + | + | + | + | + | v | v | - | + | v | - | - | |
| L-Sorbose | + | + | + | + | v | v | - | - | + | + | - | - | |
| D-Mannitol | + | + | + | - | NT | NT | NT | - | + | + | + | + | |
| α-D-Glucose | + | + | + | + | + | + | + | + | - | NT | NT | NT | |
| L-Arabinose | + | + | + | + | + | + | + | + | + | w | w | - | |
| Salicin | - | + | + | + | + | + | + | + | - | NT | NT | NT | |
| D-Trehalose | + | + | - | + | + | + | + | + | - | NT | NT | NT | |
| Maltose | + | + | + | - | + | + | + | + | - | NT | NT | NT | |
| D-Xylose | + | + | - | + | + | + | + | + | - | NT | NT | NT | |
| Sucrose | + | + | - | + | + | + | + | v | - | NT | NT | NT | |
| Cellobiose | - | - | - | - | + | + | + | + | - | NT | NT | NT | |
| Arbutin | - | - | - | - | + | + | NT | NT | - | NT | NT | NT | |
| D-Galactose | - | - | - | - | + | + | -/w | - | + | + | + | w | |
| Glycerol | - | - | - | - | v | + | -/s | - | + | - | - | - | |
| Hexadecane | - | - | - | - | - | - | NT | NT | NT | NT | NT | NT | |
| L-Malic acid | - | - | - | - | - | - | + | v | NT | NT | NT | NT | |
| L-Rhamnose | - | - | - | - | + | + | + | - | + | + | + | + | |
| Lactose | - | - | - | - | v | + | v | - | + | + | + | v | |
| Mannitol | - | - | - | - | + | + | + | - | + | - | + | - | |
| Melezitose | - | - | - | - | + | + | + | + | - | NT | NT | NT | |
| Methyl a-glucoside | - | - | - | - | + | + | + | v | - | NT | NT | NT | |
| Myo-inositol | - | - | - | - | v | + | + | - | + | + | + | + | |
| Raffinose | - | - | - | - | -/w | + | -/w | - | + | w | + | w | |
| Xylitol | - | - | - | - | + | + | - | - | + | + | + | - | |
| Citrate | - | - | - | - | - | - | + | v | - | - | - | + | |
| Melibiose | - | - | - | - | - | - | - | - | + | - | - | - | |
| Meso erythritol | - | - | - | - | - | - | - | - | w | - | - | - | |
| Glucono D-lactone | - | - | - | - | - | v | NT | NT | + | + | - | NT | |
| N-acetyl D-glucosamine | - | - | - | - | - | - | NT | NT | - | NT | NT | NT | |
| Common characteristics among N. tulchinskyi strains | |||||||||||||
| D-Cellobiose | + | + | + | + | |||||||||
| Succinic Acid Mono-Methyl Ester plus D-Xylose | + | + | + | + | |||||||||
| Palatinose | + | + | + | + | |||||||||
| Inulin | - | - | - | - | |||||||||
| Differential characteristics among N. tulchinskyi strains | |||||||||||||
| Gentiobiose | + | + | + | - | |||||||||
| D-Mannitol | + | + | + | - | |||||||||
| Maltose | + | + | + | - | |||||||||
| Acetic Acid | + | + | - | + | |||||||||
| Maltotriose | + | + | - | + | |||||||||
| Turanose | + | + | - | + | |||||||||
| D- Glucuronic Acid plus D-Xylose | + | + | - | + | |||||||||
| 1,2- Propanediol plus D-Xylose | + | + | - | + | |||||||||
| D-Trehalose | + | + | - | + | |||||||||
| D-Xylose | + | + | - | + | |||||||||
| Sucrose | + | + | - | + | |||||||||
| D-Melezitose | - | + | + | + | |||||||||
| Dextrin | - | + | + | + | |||||||||
| Propionic Acid | + | + | - | - | |||||||||
| D-Raffinose | - | - | + | - | |||||||||
| D-Sorbitol | - | - | + | - | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bijlani, S.; Parker, C.; Singh, N.K.; Sierra, M.A.; Foox, J.; Wang, C.C.C.; Mason, C.E.; Venkateswaran, K. Genomic Characterization of the Titan-like Cell Producing Naganishia tulchinskyi, the First Novel Eukaryote Isolated from the International Space Station. J. Fungi 2022, 8, 165. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8020165
Bijlani S, Parker C, Singh NK, Sierra MA, Foox J, Wang CCC, Mason CE, Venkateswaran K. Genomic Characterization of the Titan-like Cell Producing Naganishia tulchinskyi, the First Novel Eukaryote Isolated from the International Space Station. Journal of Fungi. 2022; 8(2):165. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8020165
Chicago/Turabian StyleBijlani, Swati, Ceth Parker, Nitin K. Singh, Maria A. Sierra, Jonathan Foox, Clay C. C. Wang, Christopher E. Mason, and Kasthuri Venkateswaran. 2022. "Genomic Characterization of the Titan-like Cell Producing Naganishia tulchinskyi, the First Novel Eukaryote Isolated from the International Space Station" Journal of Fungi 8, no. 2: 165. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8020165