
Taxonomy, Phylogenetic and Ancestral Area Reconstruction in Phyllachora, with Four Novel Species from Northwestern China

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collecting, Morphological Study, and Depositing Specimens
2.2. DNA Isolation, Amplification, and Sequencing
2.3. Sequence Alignment and Phylogenetic Analyses
2.4. Reconstruction of Ancestral State
2.5. Calibration Procedure
3. Results
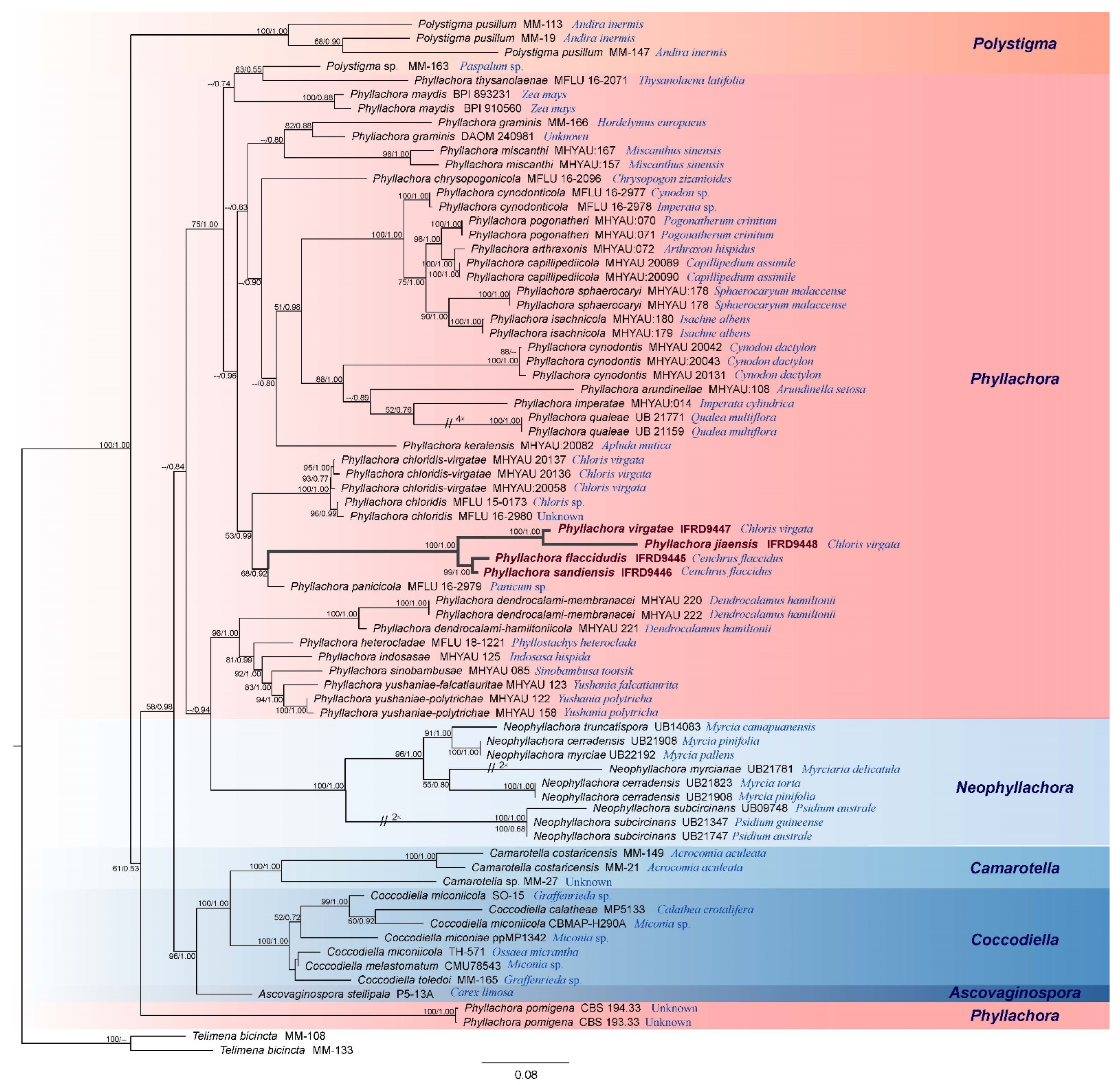
3.1. Molecular Phylogenetic Results
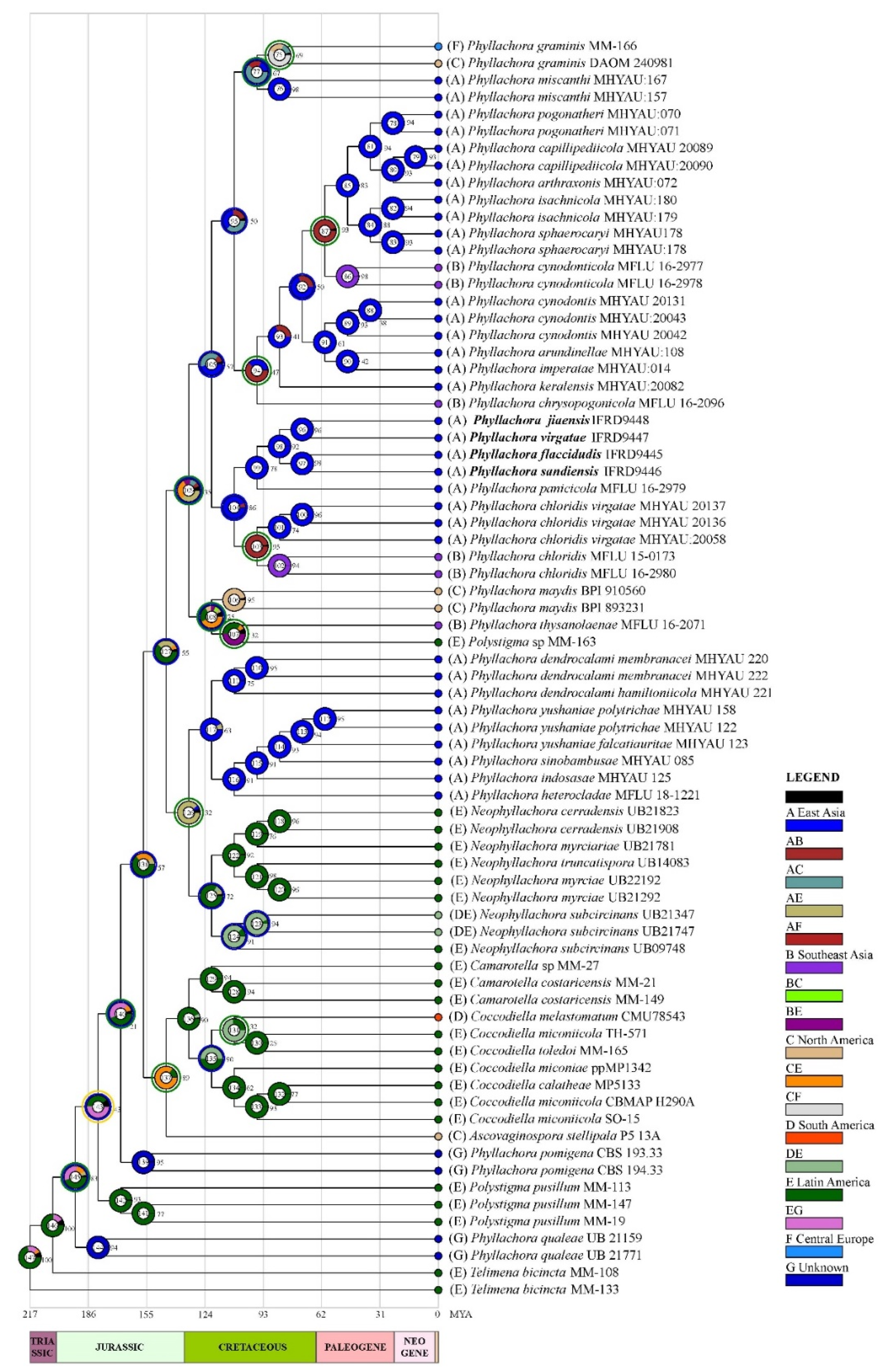
3.2. Ancestral Area Reconstruction Analysis for Phyllachora
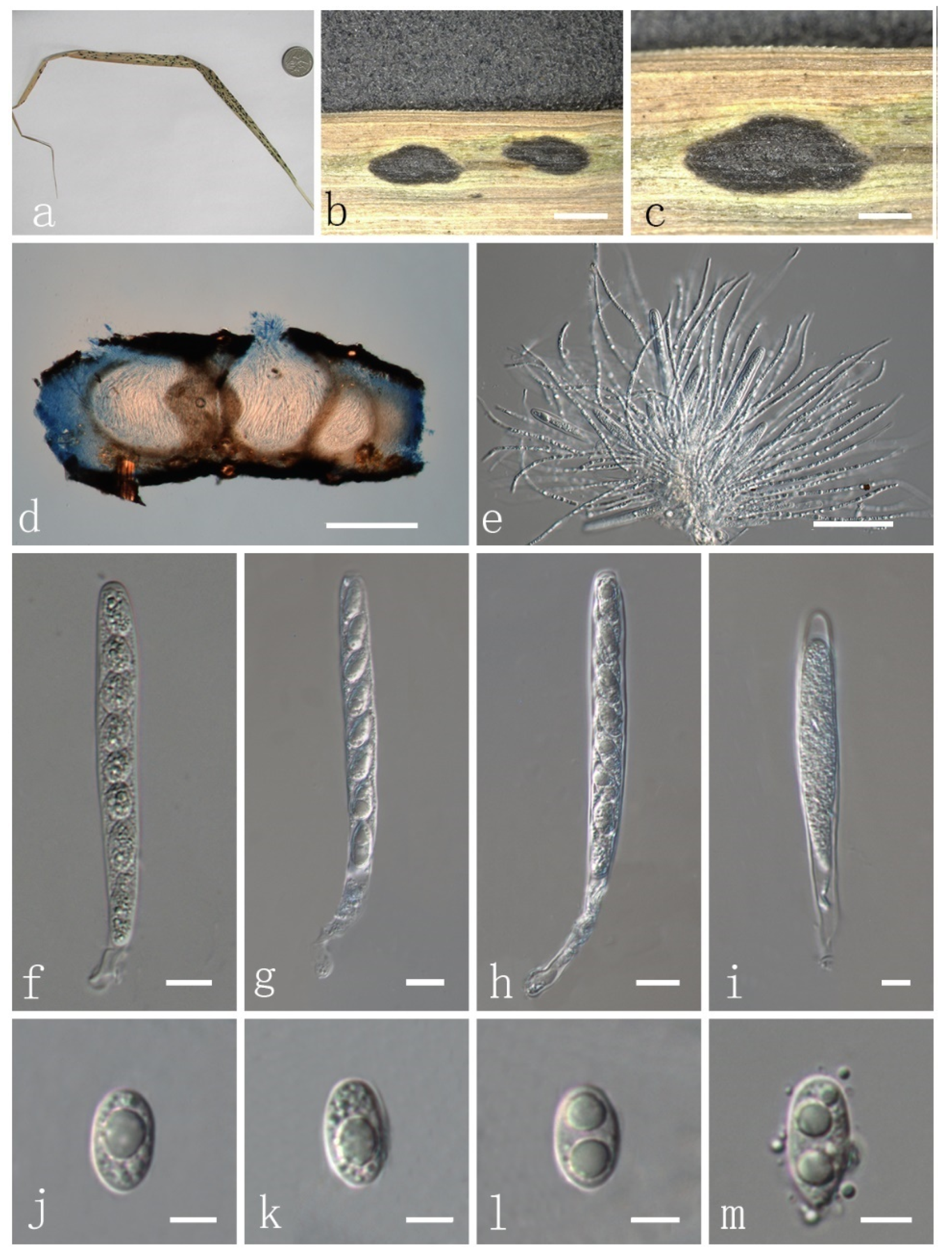
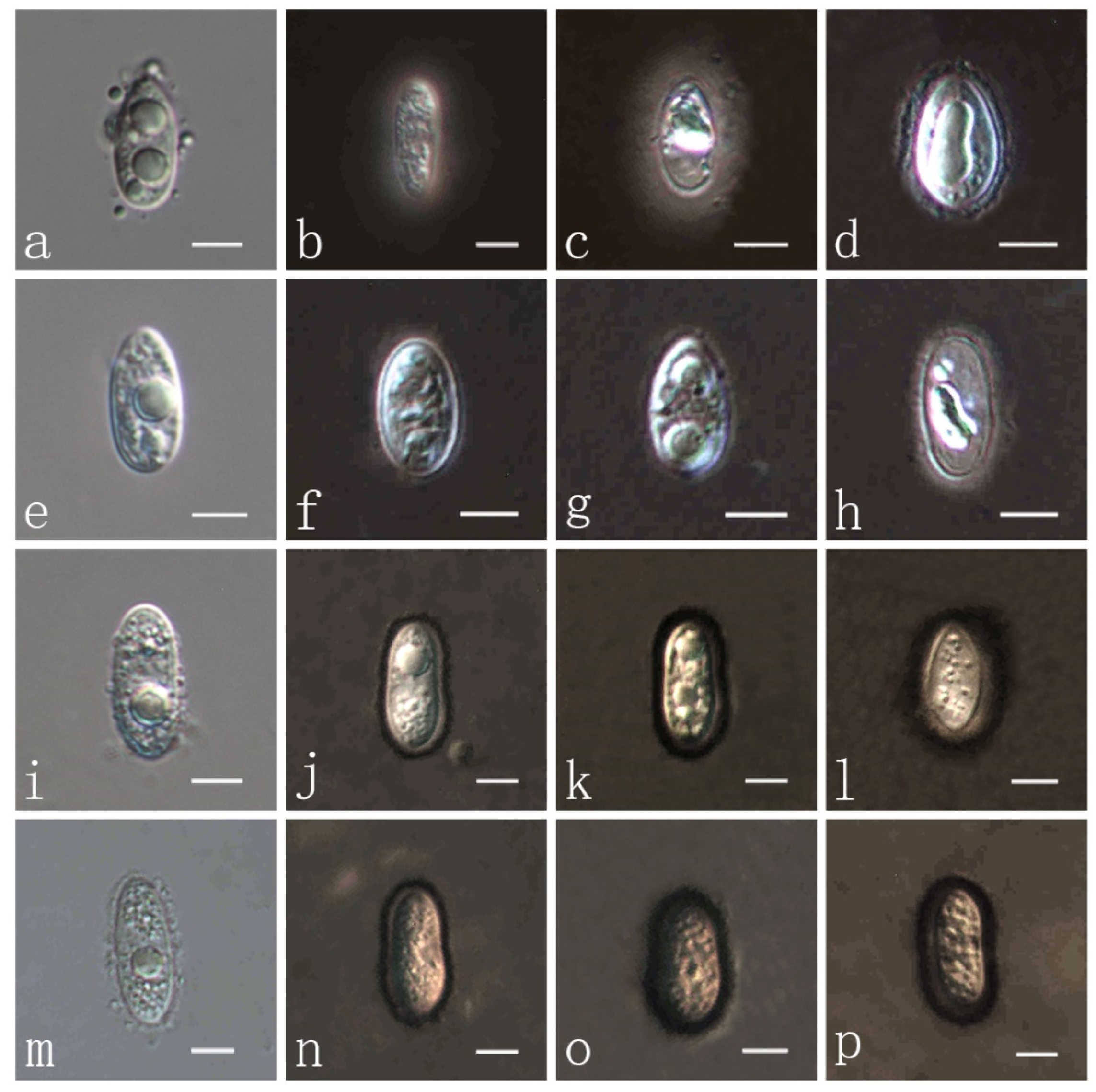
3.3. Taxonomy of Fungi
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barr, M.E. The Ascomycete Connection. Mycologia 1983, 75, 1–13. [Google Scholar] [CrossRef]
- Hyde, K.D.; Cannon, P.F.; Barr, M.E. Phaeochoraceae, a new family from palms. Syst. Ascomycetum. 1997, 15, 117–120. [Google Scholar]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of Fungi, 10th ed.; CABI: London, UK, 2008; p. 527. [Google Scholar]
- Maharachchikumbura, S.; Hyde, K.D.; Jones, E.; Mckenzie, E.; Wijayawardene, N.N. Families of Sordariomycetes. Fungal Divers. 2016, 79, 1–317. [Google Scholar] [CrossRef]
- Cannon, P.F. A Revision of Phyllachora and Some Similar Genera on the Host Family Leguminosae; CABI: Wallingford, UK, 1991; pp. 1–302. [Google Scholar]
- Pearce, C.A.; Reddell, P.; Hyde, K.D. A revision of Phyllachora (Ascomycotina) on hosts in the angiosperm family Asclepiadaceae, including P. gloriana sp. nov. on Tylophora benthamii from Australia. Fungal Divers. 1999, 3, 123–138. Available online: https://www.fungaldiversity.org/fdp/sfdp/FD_3_123-138.pdf (accessed on 13 May 2022).
- Piepenbring, M.; Hofmann, T.A.; Kirschner, R.; Mangelsdorff, R.; Trampe, T. Diversity patterns of Neotropical plant parasitic microfungi. Ecotropica 2011, 17, 27–40. [Google Scholar]
- Mardones, M.; Trampe-Jaschik, T.; Oster, S.; Elliott, M.; Piepenbring, M. Phylogeny of the order Phyllachorales (Ascomycota; Sordariomycetes): Among and within order relationships based on five molecular loci. Persoonia 2017, 39, 74–90. [Google Scholar] [CrossRef]
- Sutton, B.C.; Hodges, C.S. Hawaiian Forest Fungi. III. A New Species, Gloeocoryneum hawaiiense, on Acacia koa. Mycologia 1983, 75, 280–284. [Google Scholar] [CrossRef]
- Calatayud, V.; Navarro-Rosinés, P.; Calvo, E. Lichenochora Mediterraneae (phyllacorales, Ascomycota) a new Lichenicolous Fungus from Spain. Lichenologist 2000, 32, 225–231. [Google Scholar] [CrossRef]
- Pearce, C.A.; Reddell, P.; Hyde, K.D. Revision of the Phyllachoraceae (Ascomycota) on hosts in the angiosperm family, proteaceae. Aust. Syst. Bot. 2001, 14, 283–328. [Google Scholar] [CrossRef]
- Mardones, M.; Trampe-Jaschik, T.; Piepenbring, M. Phylogenetics and taxonomy of Telimenaceae (Phyllachorales) from Central America. Mycol. Prog. 2020, 19, 1587–1599. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T. Outline of Fungi and fungus-like taxa 2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Parbery, D.G. Studies on graminicolous species of Phyllachora Nke. in Fckl. V. A taxonomic monograph. Aust. J. Bot. 1967, 15, 271–375. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Kirk, P.M.; Sutton, B.C.; Pegler, D.N. Ainsworth and Bisby’s Dictionary of the Fungi, 8th ed.; CABI: Oxford, UK, 1995; p. 616. [Google Scholar]
- Konta, S.; Maharachchikumbura, S.S.N.; Senanayake, I.C.; McKenzie, E.H.C.; Stadler, M.; Boonmee, S.; Phookamsak, R.; Jayawardena, R.S.; Senwanna, C.; Hyde, K.D.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Theissen, F.; Sydow, H. Die Dothideales. Annales Mycologici 1915, 13, 147–746. [Google Scholar]
- Santos, M.D.M.D.; Fonseca-Boiteux, M.E.; Boiteux, L.S.; Câmara, P.E.A.S.; Dianese, J.C. ITS-phylogeny and taxonomy of Phyllachora species on native Myrtacae from the Brazilian Cerrado. Mycologia 2016, 108, 1141–1164. [Google Scholar] [CrossRef]
- Clements, F.E.; Shear, C.L. The Genera of Fungi, 2nd ed.; Wilson Co.: New York, NY, USA, 1931. [Google Scholar]
- Yang, C.L.; Xu, X.L.; Liu, Y.G.; Hyde, K.D.; Mckenzie, E. A new species of Phyllachora (Phyllachoraceae, Phyllachorales) on Phyllostachys heteroclada from Sichuan, China. Phytotaxa 2019, 392, 186–196. [Google Scholar] [CrossRef]
- McCoy, A.G.; Romberg, M.K.; Zaworsk, E.R.; Robertson, A.E.; Phibbs, A.; Hudelson, B.D.; Smith, D.L.; Beiriger, R.L.; Raid, R.N.; Byrne, J.M.; et al. First Report of Tar Spot on Corn (Zea mays) Caused by Phyllachora maydis in Florida, Iowa, Michigan, and Wisconsin. Plant Dis. 2018, 102, 6. [Google Scholar] [CrossRef]
- Lana, F.D.; Plewa, D.E.; Phillippi, E.S.; Garzonio, D.; Hesterman, R.; Kleczewski, N.M.; Paul, P.A. First report of tar spot of maize (Zea mays), caused by Phyllachora maydis, in Ohio. Plant Dis. 2019, 103, 1780. [Google Scholar] [CrossRef]
- Mottaleb, K.A.; Loladze, A.; Sonder, K.; Kruseman, G.; Vicente, F.S. Threats of Tar Spot Complex disease of maize in the United States of America and its global consequences. Mitig Adapt Strateg Glob Chang 2019, 24, 281–300. [Google Scholar] [CrossRef] [Green Version]
- Telenko, D.E.P.; Ross, T.J.; Shim, S.; Wang, Q.; Singh, R. Draft Genome Sequence Resource for Phyllachora maydis—An Obligate Pathogen That Causes Tar Spot of Corn with Recent Economic Impacts in the United States. Mol. Plant Microbe Interact. 2020, 33, 884–887. [Google Scholar] [CrossRef] [Green Version]
- Cannon, P.F. Proposal to merge the Phyllachorales with the Diaporthales, with a new family structure. Syst. Ascomycetum 1988, 7, 23–43. [Google Scholar]
- Zhang, Z.Y.; Zhang, T. Flora Fungorum Sinicorum, Vol. 46, Phyllachora; Science Press: Beijing, China, 2014; p. 530. (In Chinese) [Google Scholar]
- Tamakaew, N.; Maharachchikumbura, S.; Hyde, K.D.; Cheewangkoon, R. Tar spot fungi from Thailand. Mycosphere 2017, 8, 1054–1058. [Google Scholar] [CrossRef]
- Wu, H.X.; Li, Y.M.; Ariyawansa, H.A.; Li, W.J.; Hyde, K.D. A new species of Microthyrium from Yunnan, China. Phytotaxa 2014, 176, 213–218. [Google Scholar] [CrossRef] [Green Version]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, S.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Michael, A., David, H., Eds.; Academic Press: London, UK, 1990; pp. 315–322. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Li, X.; Wu, S.; Wang, C.; Feng, Y.; Zhao, C.; Chen, Z.-Q.; Yu, J.-F.; Luo, R.; Promputtha, I.; Sun, D.-F. Two new species of Phyllachora (Phyllachoraceae; Phyllachorales) on bamboo from China. Phytotaxa 2019, 425, 78–86. [Google Scholar] [CrossRef]
- Dayarathne, M.; Maharachchikumbura, S.; Jones, E.B.G.; Goonasekara, I.D.; Bulgakov, T.S.; Al-Sadi, A.M.; Lumyong, S.; McKenzie, E.H.C. Neophyllachora gen. nov. (Phyllachorales), three new species of Phyllachora from Poaceae and resurrection of Polystigmataceae (Xylariales). Mycosphere 2017, 8, 1598–1625. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinformatics 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. raxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2021, 12, 373–377. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest v2. Program distributed by the author. Bioinformatics 2004, 24, 533–581. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Jeewon, R.; Hyde, K.D. Phylogenetic investigations of Sordariaceae based on multiple gene sequences and morphology. Mycol. Res. 2006, 110, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Guo, X.Y.; Hyde, K.D. Morphological and molecular characterization of a new anamorphic genus Cheirosporium; from freshwater in China. Persoonia 2008, 20, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.K.; Phookamsak, R.; Doilom, M.; Wikee, S.; Li, Y.M.; Ariyawansha, H.; Boonmee, S.; Chomnunti, P.; Dai, D.-Q.; Bhat, J.D.; et al. Towards a natural classification of Botryosphaeriales. Fungal Divers. 2012, 57, 149–210. [Google Scholar] [CrossRef]
- Wu, H.X.; Schoch, C.L.; Boonmee, S.; Bahkali, A.H.; Chomnunti, P.; Hyde, K.D. A reappraisal of Microthyriaceae. Fungal Divers. 2011, 51, 189–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Harris, A.J.; Blair, C.; He, X.J. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Mol. Phylogenet. Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef]
- Hongsanan, S.; Maharachchikumbura, S.S.; Hyde, K.D.; Samarakoon, M.C.; Jeewon, R.; Zhao, Q.; Al-Sadi, A.; Bahkali, A.H. An updated phylogeny of Sordariomycetes based on phylogenetic and molecular clock evidence. Fungal Divers. 2017, 84, 25–41. [Google Scholar] [CrossRef]
- Liu, N.; Guo, L. Two new records of Phyllachora (Phyllachorales) from China. Mycotaxon 2009, 107, 303–306. [Google Scholar] [CrossRef]
- Lan, J.Q.; Lu, H.J.; Wu, J. Three new records of Phyllachora in China. Mycosystema 2012, 31, 639–641. [Google Scholar]
- Parbery, D.G. Studies on Graminicolous species, of Phyllachora Nke. in Fckl. VI. Additions and corrections to part V. Aust. J. Bot. 1971, 19, 207–235. [Google Scholar] [CrossRef]
- Wu, J. Additional three new Chinese records of Phyllachora. Mycosystema 2012, 31, 642–644. [Google Scholar]
- Teng, X.Q. Additional three new records of Phyllachora in China. Mycosystema 2011, 30, 782–784. [Google Scholar]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 12 April 2022).
- Silva-Hanlin, D.M.W.; Halin, R.T. The order Phyllachorales: Taxonomic review. Mycoscience 1998, 39, 97–104. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Humphries, C.J.; Escudero, M.; Martín-Bravo, S. Vicariance Biogeography. In Reference Module in Life Sciences, 3rd ed.; Roitberg, B., Ed.; Elsevier: Cambridge, UK, 2017; pp. 1–13. [Google Scholar]
- Feng, J.G.; Fan, F.C.; Li, F. Origin, Evolution and Distribution of the Gramineae. Chinese J. Bot. 1996, 13, 9–13. (In Chinese) [Google Scholar]
- Tzvelev, N.N. The system of grasses (Poaceae) and their evolution. Bot. Rev. 1989, 55, 141–203. [Google Scholar] [CrossRef]
- Schneider, J.V.; Bissiengou, P.; Amaral, M.; Tahir, A.; Fay, M.F.; Thines, M.; Sosef, M.S.; Zizka, G.; Chatrou, L.W. Phylogenetics, ancestral state reconstruction, and a new infrafamilial classification of the pantropical Ochnaceae (Medusagynaceae, Ochnaceae s.str. Quiinaceae) based on five dna regions. Mol. Phylogenet. Evol. 2014, 78, 199–214. [Google Scholar] [CrossRef]
- Ho, L.; Dinh, V.; Nguyen, C.V. Multi-task learning improves ancestral state reconstruction. Theor. Popul. Biol. 2019, 126, 33–39. [Google Scholar] [CrossRef]
- Gautam, A.K.; Verma, R.K.; Avasthi, S.; Sushma; Bohra, Y.; Devadatha, B.; Niranjan, M.; Suwannarach, N. Current Insight into Traditional and Modern Methods in Fungal Diversity Estimates. J. Fungi 2022, 8, 226. [Google Scholar] [CrossRef]
- Li, J.; Han, L.H.; Liu, X.B.; Zhao, Z.W.; Yang, Z.L. The saprotrophic Pleurotus ostreatus species complex: Late Eocene origin in East Asia; multiple dispersal, and complex speciation. IMA Fungus 2020, 11, e10. [Google Scholar] [CrossRef]
- Quan, Y.; Muggia, L.; Moreno, L.F.; Wang, M.; Hoog, S.D. A re-evaluation of the Chaetothyriales using criteria of comparative biology. Fungal Divers. 2020, 103, 47–85. [Google Scholar] [CrossRef]
- Thiyagaraja, V.; Lücking, R.; Ertz, D.; Karunarathna, S.C.; Wanasinghe, D.N.; Lumyong, S.; Hyde, K. The Evolution of Life Modes in Stictidaceae, with Three Novel Taxa. J. Fungi 2021, 7, 105. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Location | Source | Host Family | GenBank Accession Numbers | Reference | ||
|---|---|---|---|---|---|---|---|
| LSU | SSU | ITS | |||||
| Ascovaginospora stellipalaT | North America (northern Wisconsin) | P5-13A | Cyperaceae | U85088 | U85087 | - | [33] |
| Camarotella costaricensis | Latin America (Panama) | MM-21 | Arecaceae | KX430490 | KX451851 | KX451900 | [8] |
| Camarotella costaricensis | Latin America (Panama) | MM-149 | Arecaceae | KX430484 | KX451863 | KX451913 | [8] |
| Camarotella sp. | Latin America (Panama) | MM-27 | Arecaceae | KX430492 | KX451852 | KX451901 | [8] |
| Coccodiella calatheaeT | Latin America (Panama) | MP5133 | Marantaceae | MF460370 | MF460376 | MF460366 | [8] |
| Coccodiella melastomatum | North America (Venezuela) | CMU78543 | Melastomataceae | - | U78543 | - | [8] |
| Coccodiella miconiae | Latin America (Panama) | ppMP1342 | Melastomataceae | KX430506 | KX451871 | MF460365 | [8] |
| Coccodiella miconiicola | Latin America (Panama) | TH-571 | Melastomataceae | KX430512 | KX451880 | - | [8] |
| Coccodiella miconiicola | Latin America (Panama) | CBMAP-H290A | Melastomataceae | MF460373 | MF460379 | MF460368 | [32] |
| Coccodiella miconiicola | Latin America (Ecuador) | SO-15 | Melastomataceae | MF460374 | MF460380 | MF460369 | [32] |
| Coccodiella toledoi | Latin America (Ecuador) | MM-165 | Melastomataceae | KX430488 | KX451865 | KX451917 | [8] |
| Neophyllachora cerradensis | Latin America (Brazil) | UB21823 | Myrtaceae | - | - | KC683470 | [18] |
| Neophyllachora cerradensisT | Latin America (Brazil) | UB21908 | Myrtaceae | - | - | KC683471 | [18] |
| Neophyllachora myrciae | Latin America (Brazil) | UB21292 | Myrtaceae | - | - | KC683463 | [18] |
| Neophyllachora myrciae | Latin America (Brazil) | UB22192 | Myrtaceae | - | - | KC683476 | [18] |
| Neophyllachora myrciariaeT | Latin America (Brazil) | UB21781 | Myrtaceae | - | - | KC683469 | [18] |
| Neophyllachora subcircinans | Latin America (Brazil) | UB09748 | Myrtaceae | - | - | KC683441 | [18] |
| Neophyllachora subcircinans | Latin America (Brazil), South America (Paraguay) | UB21347 | Myrtaceae | - | - | KC683466 | [18] |
| Neophyllachora subcircinans | Latin America (Brazil), South America (Paraguay) | UB21747 | Myrtaceae | - | KC902622 | KC683467 | [18] |
| Neophyllachora truncatispora | Latin America (Brazil) | UB14083 | Myrtaceae | - | KC902614 | KC683448 | [18] |
| Phyllachora arthraxonis | East Asia (China) | MHYAU:072 | Poaceae | MG269803 | - | MG269749 | [20] |
| Phyllachora arundinellae | East Asia (China) | MHYAU:108 | Poaceae | MG269815 | - | MG269761 | [20] |
| Phyllachora capillipediicola | East Asia (China) | MHYAU 20089 | Poaceae | MG356698 | - | KY498084 | [32] |
| Phyllachora capillipediicola | East Asia (China) | MHYAU:20090 | Poaceae | MG356699 | - | KY498115 | [20] |
| Phyllachora chloridisT | Southeast Asia (Thailand) | MFLU 15-0173 | Poaceae | MF197499 | MF197505 | KY594026 | [33] |
| Phyllachora chloridis | Southeast Asia (Thailand) | MFLU 16-2980 | Poaceae | MF197500 | MF197506 | KY594027 | [33] |
| Phyllachora chloridis-virgatae | East Asia (China) | MHYAU 20136 | Poaceae | MG356685 | - | KY498122 | [32] |
| Phyllachora chloridis-virgatae | East Asia (China) | MHYAU:20058 | Poaceae | MG356683 | - | KY498102 | [32] |
| Phyllachora chloridis-virgatae | East Asia (China) | MHYAU 20137 | Poaceae | MG356686 | - | KY498092 | [32] |
| Phyllachora chrysopogonicolaT | Southeast Asia (Thailand) | MFLU 16-2096 | Poaceae | MF372146 | - | MF372145 | [20] |
| Phyllachora cynodonticolaT | Southeast Asia (Thailand) | MFLU 16-2977 | Poaceae | MF197501 | MF197507 | KY594024 | [33] |
| Phyllachora cynodonticola | Southeast Asia (Thailand) | MFLU 16-2978 | Poaceae | MF197502 | MF197508 | KY594025 | [33] |
| Phyllachora cynodontis | East Asia (China) | MHYAU 20042 | Poaceae | KY498080 | - | KY471328 | [32] |
| Phyllachora cynodontis | East Asia (China) | MHYAU:20043 | Poaceae | KY498081 | - | KY471329 | [20] |
| Phyllachora cynodontis | East Asia (China) | MHYAU 20131 | Poaceae | KY498079 | - | KY471327 | [32] |
| Phyllachora dendrocalami-hamiltoniicola | East Asia (China) | MHYAU 221 | Poaceae | MK614118 | - | - | [32] |
| Phyllachora dendrocalami-membranacei | East Asia (China) | MHYAU 220 | Poaceae | MK614117 | - | MK614102 | [32] |
| Phyllachora dendrocalami-membranacei | East Asia (China) | MHYAU 222 | Poaceae | MK614119 | - | MK614103 | [32] |
| Phyllachora flaccidudisT | East Asia (China) | IFRD9445 | Poaceae | ON072101 | ON072097 | ON075524 | This study |
| Phyllachora graminis | North America (Canada) | DAOM 240981 | Poaceae | - | - | HQ317550 | [33] |
| Phyllachora graminis | Central Europe (Germany) | MM-166 | Poaceae | - | KX451869 | KX451920 | [8] |
| Phyllachora heterocladaeT | East Asia (China) | MFLU 18-1221 | Poaceae | MK296472 | MK296468 | MK305902 | [20] |
| Phyllachora imperatae | East Asia (China) | MHYAU:014 | Poaceae | MG269800 | - | MG269746 | [20] |
| Phyllachora indosasae | East Asia (China) | MHYAU 125 | Poaceae | MG195662 | - | MG195637 | [32] |
| Phyllachora isachnicolaT | East Asia (China) | MHYAU:179 | Poaceae | MH018563 | - | MH018561 | [20] |
| Phyllachora isachnicola | East Asia (China) | MHYAU:180 | Poaceae | MH018564 | - | MH018562 | [20] |
| Phyllachora jiaensisT | East Asia (China) | IFRD9448 | Poaceae | ON075440 | ON072100 | ON075527 | This study |
| Phyllachora keralensis | East Asia (China) | MHYAU:20082 | Poaceae | MG269792 | - | KY498106 | [20] |
| Phyllachora maydis | North America (USA) | BPI 893231 | Poaceae | - | - | KU184459 | [33] |
| Phyllachora maydis | North America (Wisconsin) | BPI 910560 | Poaceae | - | - | MG881846 | [20] |
| Phyllachora miscanthi | East Asia (China) | MHYAU:167 | Poaceae | MG195669 | - | MG195644 | [20] |
| Phyllachora miscanthi | East Asia (China) | MHYAU:157 | Poaceae | MG195668 | - | MG195643 | [20] |
| Phyllachora panicicolaT | East Asia (China) | MFLU 16-2979 | Poaceae | MF197503 | MF197504 | KY594028 | [33] |
| Phyllachora pogonatheri | East Asia (China) | MHYAU:071 | Poaceae | MG269802 | - | MG269748 | [20] |
| Phyllachora pogonatheri | East Asia (China) | MHYAU:070 | Poaceae | MG269801 | - | MG269747 | [20] |
| Phyllachora pomigena | unknown | CBS 194.33 | Unknown | MH866861 | - | MH855410 | [20] |
| Phyllachora pomigena | unknown | CBS 193.33 | Unknown | MH866860 | - | MH855409 | [20] |
| Phyllachora qualeae | unknown | UB 21159 | Vochysiaceae | - | - | KU682781 | [33] |
| Phyllachora qualeae | unknown | UB 21771 | Vochysiaceae | - | - | KU682780 | [33] |
| Phyllachora sandiensisT | East Asia (China) | IFRD9446 | Poaceae | ON075528 | ON072098 | ON075525 | This study |
| Phyllachora sinobambusae | East Asia (China) | MHYAU 085 | Poaceae | MG195655 | - | MG195630 | [32] |
| Phyllachora sphaerocaryiT | East Asia (China) | MHYAU 178 | Poaceae | MK614114 | - | MK614100 | [32] |
| Phyllachora sphaerocaryi | East Asia (China) | MHYAU:178 | Poaceae | - | - | MH018560 | [20] |
| Phyllachora thysanolaenaeT | Southeast Asia (Thailand) | MFLU 16-2071 | Poaceae | - | MF372147 | - | [20] |
| Phyllachora virgataesT | East Asia (China) | IFRD9447 | Poaceae | ON075439 | ON072099 | ON075526 | This study |
| Phyllachora yushaniae-falcatiauritae | East Asia (China) | MHYAU 123 | Poaceae | MG195656 | - | MG195631 | [32] |
| Phyllachora yushaniae-polytrichae | East Asia (China) | MHYAU 122 | Poaceae | MG195657 | MH992455 | MG195632 | [32] |
| Phyllachora yushaniae-polytrichae | East Asia (China) | MHYAU 158 | Poaceae | MG195658 | - | MG195633 | [32] |
| Polystigma pusillum | Latin America (Costa Rica) | MM-113 | Fabaceae | KX430474 | KX451858 | KX451907 | [8] |
| Polystigma pusillum | Latin America (Costa Rica) | MM-147 | Fabaceae | KX430483 | KX451862 | - | [8] |
| Polystigma pusillum | Latin America (Panama) | MM-19 | Fabaceae | KX430489 | KX451850 | KX451899 | [8] |
| Polystigma sp. | Latin America (Ecuador) | MM-163 | Poaceae | KX430487 | KX451864 | KX451916 | [8] |
| Telimena bicincta | Latin America (Costa Rica) | MM-108 | Picramniaceae | KX430473 | KX451857 | KX451906 | [8] |
| Telimena bicincta | Latin America (Costa Rica) | MM-133 | Picramniaceae | KX430478 | KX451861 | KX451910 | [8] |
| Fungal Taxa | Hosts | Color of the Stromata | Asci (μm) | Ascospores (μm) | References | ||
|---|---|---|---|---|---|---|---|
| Size | No. of Septa | Shape | |||||
| P. africana (P. oblongospora) | Eremopogon delavayi, Leea elata, Chloris sp. | Black | 100–140 × 9–12.5 | 10–17 × 5–9 | Aseptate | ovoid | [44] |
| P. sphaerosperma (P. cenchricola) | Cenchrus echinatus | Black | 65–100 × 10–13 | 8–11 × 7–9 | Aseptate | nearly spherical | [14] |
| P. centothecae | Centotheca lappacea | Bright | 46.3 × 9.0, pedicel 11.6 × 2.6 | 7.7–9.0 × 4.6–5.1 | Aseptate | oval | [45] |
| P. coorgiana | Coix lachryma-jobi | Bright | 59–100 ×18–26 | 10.3–20.6 × 7.2–12.9 | Aseptate | ellipsoid or ovoid, rarely subglobose | [45] |
| P. chloridis | Chloris sp. | Bright | 50–72 × 6–8 | 8–12 × 3.5–4.8 | Aseptate | fusiform to oval | [33] |
| P. cynodontis | Chloris sp. | Unknown | 45–50 × 12–15 with a stipe 20–25 long, sometimes short | 8–15 × 5–6 | 0–1 | ovoid | [14,46] |
| P. digitariicola | Digitaria sanguinalis | Bright | 62–103 × 12–15 | 10–18 × 6–8 | Aseptate | ellipsoid, rounded at both ends, rarely subglobose or oval | [45] |
| P. eriochloae var. colombiensis | Gramineae | Unknown | 37–71 × 102–13 | 4–13.9 × 5.4–7.2 | Aseptate | ovate, rarely subglobose, rarely irregularly | [47] |
| P. eriochloae var. eriochloae | Eragrostis sp. | Bright | 43.7–69.4 × 10.3–12.9 | 7.7–12.9 × 5.1–5.1 | Aseptate | ovoid or tear-like | [47] |
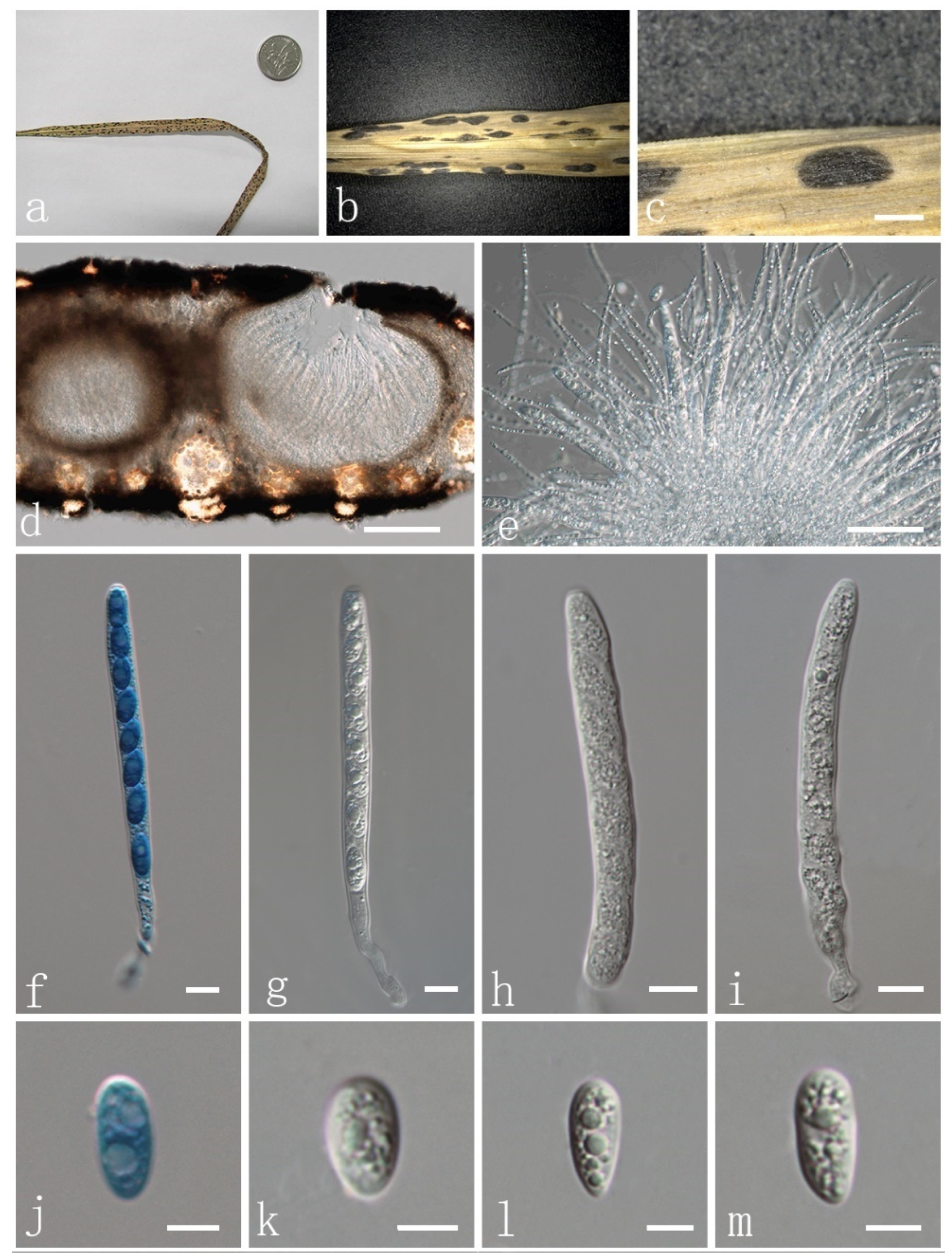
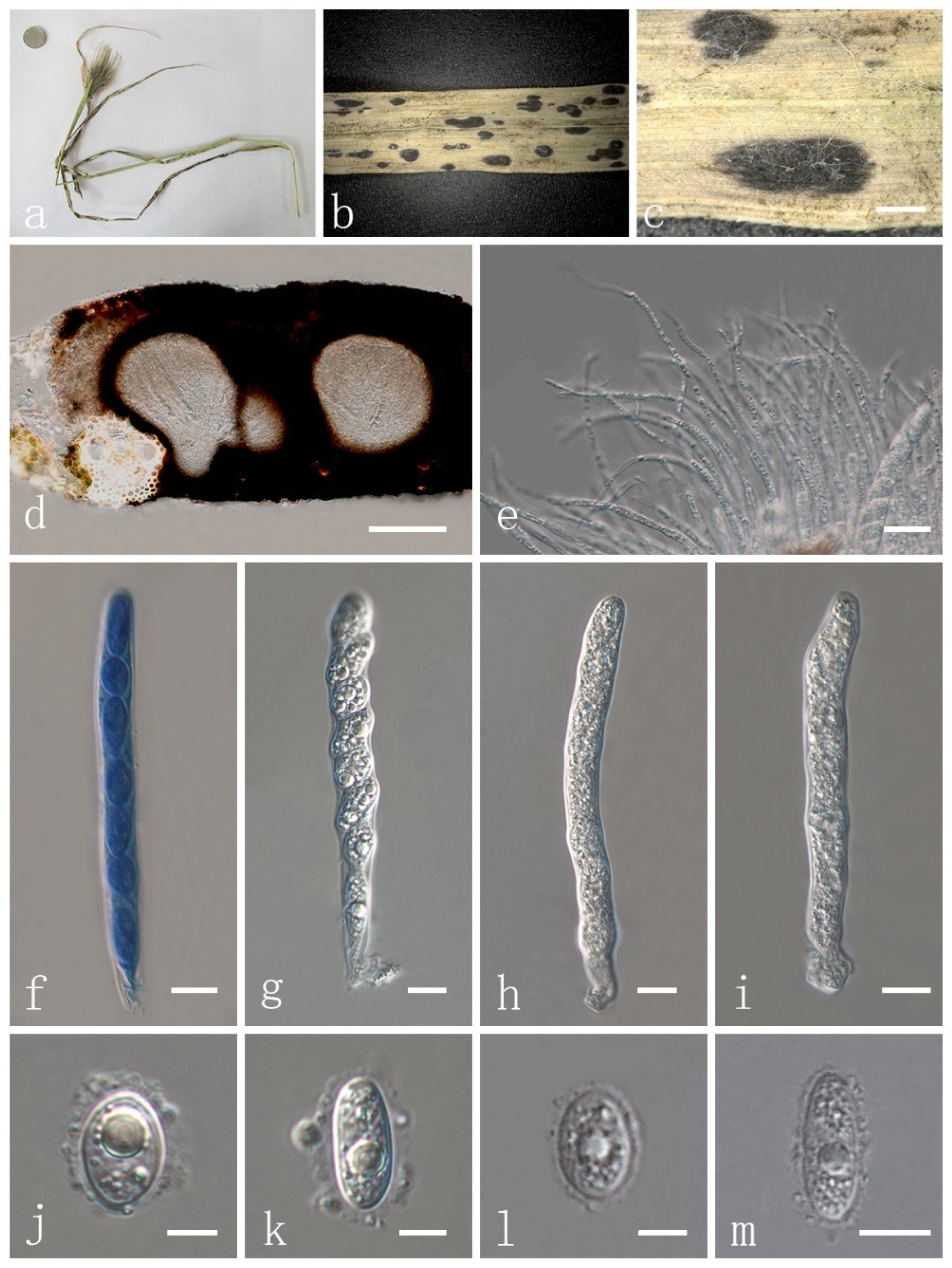
| P. flaccidudis | Cenchrus flaccidus | Black | 82–110 × 7–10 | 11–13 × 4–7 | Aseptate | drop shape, oval to ellipse, rounded at the ends | This study |
| P. graminis | Chloris sp., Elymus sp., Agvopyron sp., Arrenathevum sp., Asperella sp., Agrostis sp., Brachyelytvum sp., Bvomus sp., Cinna sp. | Dark brown to black | 60–70× 8–10 | 7–14× 4–7 | Aseptate | oval to ovoid or ovoid with obtuse end flattened or blunted | [14] |
| P. graminis var. cynodonticola | Cynodon dactylon | Bright | 82–87 × 7.7–8.1, with short peduncle 26 × 2.1 | 7.5–14 × 5.1–6.5 | Aseptate | usually oblique, rarely irregularly biseriate, ellipsoid or subglobose | [47] |
| P. jiaensis | Chloris virgata | Black | 77–114 × 8–12 | 9–17 × 8–9 | Aseptate | oval to ellipse, rounded at the ends | This study |
| P. koondrookensis | Chloris truncatae | Black | 75-87 × 12-16 | 14–16.5 × 5–5.5 | Aseptate | uniseriatae vel inordinatae, anguste ellipsoideae usque and oblongae | [14] |
| P. minutissima | Pennisetum flaccidum, Chloris sp., Panicum sp., Pennisetum sp., Pseudoechinochlaena sp., Setaria sp. | Black | 51.2–54.0 × 11.6–14.9 | 15.7–19.1 × 6.3–8.2 | Aseptate | ovoid or ovate acuminately | [14,48] |
| P. platyelliptica | Themeda giguntia | Bright | 64.1–92.1 × 10.1–12.1 | 13.6–16.5 × 3.8–6.5 | Aseptate | narrow-ellipsoid | [48] |
| P. panicicola | Panicum sp. | Bright | 110–130 × 10–14 | 14–16 × 6–8 | Aseptate | ellipsoidal, rounded at the ends | [33] |
| P. sandiensis | Cenchrus flaccidus | Black | 92–126 × 7–10 | 10–14 × 6–7 | Aseptate | drop shape, oval to ellipse | This study |
| P. virgatae | Chloris virgata | Bright | 84–120 × 7–11 | 10–15 × 6–9 | Aseptate | oval to ellipse, rounded at the ends | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.-C.; Wu, H.-X.; Li, Y.; Li, X.-H.; Song, J.-Y.; Suwannarach, N.; Wijayawardene, N.N. Taxonomy, Phylogenetic and Ancestral Area Reconstruction in Phyllachora, with Four Novel Species from Northwestern China. J. Fungi 2022, 8, 520. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050520
Li J-C, Wu H-X, Li Y, Li X-H, Song J-Y, Suwannarach N, Wijayawardene NN. Taxonomy, Phylogenetic and Ancestral Area Reconstruction in Phyllachora, with Four Novel Species from Northwestern China. Journal of Fungi. 2022; 8(5):520. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050520
Chicago/Turabian StyleLi, Jin-Chen, Hai-Xia Wu, Yuying Li, Xin-Hao Li, Jia-Yu Song, Nakarin Suwannarach, and Nalin N. Wijayawardene. 2022. "Taxonomy, Phylogenetic and Ancestral Area Reconstruction in Phyllachora, with Four Novel Species from Northwestern China" Journal of Fungi 8, no. 5: 520. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050520