
Synergistic In Vitro Interaction of Isavuconazole and Isoquercitrin against Candida glabrata

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Drugs
2.3. Medium Preparation
2.4. Microplate Preparation
2.5. Inoculum Preparation and Inoculation of Microplates
2.6. Interpretation of the Results by Fractional Inhibition Concentration Index
2.7. Interpretation of the Results by Response Surface Analysis
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kullberg, B.J.; Arendrup, M.C. Invasive candidiasis. N. Engl. J. Med. 2015, 373, 1445–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yapar, N. Epidemiology and risk factors for invasive candidiasis. Ther. Clin. Risk Manag. 2014, 10, 95–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A.; Diekema, D.J.; Gibbs, D.L.; Newell, V.A.; Ellis, D.; Tullio, V.; Rodloff, A.; Fu, W.; Ling, T.A.; Global Antifungal Surveillance Group. Results from the ARTEMIS DISK global antifungal surveillance study, 1997 to 2007: A 10.5-year analysis of susceptibilities of Candida species to fluconazole and voriconazole as determined by CLSI standardized disk diffusion. J. Clin. Microbiol. 2010, 48, 1366–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montagna, M.T.; Caggiano, G.; Lovero, G.; De Giglio, O.; Coretti, C.; Cuna, T.; Iatta, R.; Giglio, M.; Dalfino, L.; Bruno, F.; et al. Epidemiology of invasive fungal infections in the intensive care unit: Results of a multicenter Italian survey (AURORA Project). Infection 2013, 41, 645–653. [Google Scholar] [CrossRef] [Green Version]
- Vannini, M.; Emery, S.; Lieutier-Colas, F.; Legueult, K.; Mondain, V.; Retur, N.; Gastaud, L.; Pomares, C.; Hasseine, L. Epidemiology of candidemia in NICE area, France: A five-year study of antifungal susceptibility and mortality. J. Mycol. Med. 2022, 32, 101210. [Google Scholar] [CrossRef]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Primers 2018, 4, 18026. [Google Scholar] [CrossRef]
- Cornely, O.A.; Bassetti, M.; Calandra, T.; Garbino, J.; Kullberg, B.J.; Lortholary, O.; Meersseman, W.; Akova, M.; Arendrup, M.C.; Arikan-Akdagli, S.; et al. ESCMID guideline for the diagnosis and management of Candida diseases 2012: Non-neutropenic adult patients. Clin. Microbiol. Infect. 2012, 18 (Suppl. S7), 19–37. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, M.A.; Boyken, L.; Hollis, R.J.; Messer, S.A.; Tendolkar, S.; Diekema, D.J. In vitro susceptibilities of Candida spp. to caspofungin: Four years of global surveillance. J. Clin. Microbiol. 2006, 44, 760–763. [Google Scholar] [CrossRef] [Green Version]
- Orozco, A.S.; Higginbotham, L.M.; Hitchcock, C.A.; Parkinson, T.; Falconer, D.; Ibrahim, A.S.; Ghannoum, M.A.; Filler, S.G. Mechanism of fluconazole resistance in Candida krusei. Antimicrob. Agents Chemother. 1998, 42, 2645–2649. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Effron, G.; Lee, S.; Park, S.; Cleary, J.D.; Perlin, D.S. Effect of Candida glabrata FKS1 and FKS2 mutations on echinocandin sensitivity and kinetics of 1,3-beta-D-glucan synthase: Implication for the existing susceptibility breakpoint. Antimicrob. Agents Chemother. 2009, 53, 3690–3699. [Google Scholar] [CrossRef] [Green Version]
- Dannaoui, E.; Desnos-Ollivier, M.; Garcia-Hermoso, D.; Grenouillet, F.; Cassaing, S.; Baixench, M.T.; Bretagne, S.; Dromer, F.; Lortholary, O.; French Mycoses Study Group. Candida spp. with acquired echinocandin resistance, France, 2004–2010. Emerg. Infect. Dis. 2012, 18, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Alexander, B.D.; Johnson, M.D.; Pfeiffer, C.D.; Jimenez-Ortigosa, C.; Catania, J.; Booker, R.; Castanheira, M.; Messer, S.A.; Perlin, D.S.; Pfaller, M.A. Increasing echinocandin resistance in Candida glabrata: Clinical failure correlates with presence of FKS mutations and elevated minimum inhibitory concentrations. Clin. Infect. Dis. 2013, 56, 1724–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messer, S.A.; Carvalhaes, C.G.; Castanheira, M.; Pfaller, M.A. In vitro activity of isavuconazole versus opportunistic filamentous fungal pathogens from the SENTRY antifungal surveillance program, 2017-2018. Diagn. Microbiol. Infect. Dis. 2020, 97, 115007. [Google Scholar] [CrossRef]
- Fuller, J.; Dingle, T.C.; Bull, A.; Shokoples, S.; Laverdiere, M.; Baxter, M.R.; Adam, H.J.; Karlowsky, J.A.; Zhanel, G.G.; Canadian Antimicrobial Resistance Alliance; et al. Species distribution and antifungal susceptibility of invasive Candida isolates from Canadian hospitals: Results of the CANWARD 2011-16 study. J. Antimicrob. Chemother. 2019, 74, iv48–iv54. [Google Scholar] [CrossRef] [PubMed]
- Astvad, K.M.T.; Johansen, H.K.; Roder, B.L.; Rosenvinge, F.S.; Knudsen, J.D.; Lemming, L.; Schonheyder, H.C.; Hare, R.K.; Kristensen, L.; Nielsen, L.; et al. Update from a 12-year nationwide fungemia surveillance: Increasing intrinsic and acquired resistance causes concern. J. Clin. Microbiol. 2018, 56, e01564-17. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Herrera, E.; Frias-De-Leon, M.G.; Hernandez-Castro, R.; Garcia-Salazar, E.; Arenas, R.; Ocharan-Hernandez, E.; Rodriguez-Cerdeira, C. Antifungal resistance in clinical isolates of Candida glabrata in Ibero-America. J. Fungi 2021, 8, 14. [Google Scholar] [CrossRef]
- Aldejohann, A.M.; Herz, M.; Martin, R.; Walther, G.; Kurzai, O. Emergence of resistant Candida glabrata in Germany. JAC Antimicrob. Resist. 2021, 3, dlab122. [Google Scholar] [CrossRef]
- Vitale, R.G. Role of antifungal combinations in difficult to treat Candida infections. J. Fungi 2021, 7, 731. [Google Scholar] [CrossRef]
- Vitale, R.G.; Afeltra, J.; Dannaoui, E. Antifungal combinations. Methods Mol. Med. 2005, 118, 143–152. [Google Scholar] [CrossRef]
- Perfect, J.R.; Dismukes, W.E.; Dromer, F.; Goldman, D.L.; Graybill, J.R.; Hamill, R.J.; Harrison, T.S.; Larsen, R.A.; Lortholary, O.; Nguyen, M.H.; et al. Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2010, 50, 291–322. [Google Scholar] [CrossRef] [Green Version]
- Kontoyiannis, D.P.; Lewis, R.E. Toward more effective antifungal therapy: The prospects of combination therapy. Br. J. Haematol. 2004, 126, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.D.; MacDougall, C.; Ostrosky-Zeichner, L.; Perfect, J.R.; Rex, J.H. Combination antifungal therapy. Antimicrob. Agents Chemother. 2004, 48, 693–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, P.; Nikolskiy, I.; Bidaud, A.L.; Sommer, F.; Bange, G.; Dannaoui, E. In vitro synergy of isavuconazole combined with colistin against common Candida species. Front. Cell Infect. Microbiol. 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, P.; Nikolskiy, I.; Bidaud, A.L.; Sommer, F.; Bange, G.; Dannaoui, E. In vitro activity of amphotericin B in combination with colistin against fungi responsible for invasive infections. J. Fungi 2022, 8, 115. [Google Scholar] [CrossRef]
- Schwarz, P.; Bidaud, A.L.; Dannaoui, E. In vitro synergy of isavuconazole in combination with colistin against Candida auris. Sci. Rep. 2020, 10, 21448. [Google Scholar] [CrossRef]
- Schwarz, P.; Djenontin, E.; Dannaoui, E. Colistin and isavuconazole interact synergistically in vitro against Aspergillus nidulans and Aspergillus niger. Microorganisms 2020, 8, 1447. [Google Scholar] [CrossRef]
- Schwarz, P.; Dannaoui, E. In vitro interaction between isavuconazole and tacrolimus, cyclosporin A, or sirolimus against Aspergillus Species. J. Fungi 2020, 6, 103. [Google Scholar] [CrossRef]
- Schwarz, P.; Schwarz, P.V.; Felske-Zech, H.; Dannaoui, E. In vitro interactions between isavuconazole and tacrolimus, cyclosporin A or sirolimus against Mucorales. J. Antimicrob. Chemother. 2019, 74, 1921–1927. [Google Scholar] [CrossRef]
- Seifert, H.; Aurbach, U.; Stefanik, D.; Cornely, O. In vitro activities of isavuconazole and other antifungal agents against Candida bloodstream isolates. Antimicrob. Agents Chemother. 2007, 51, 1818–1821. [Google Scholar] [CrossRef] [Green Version]
- Maertens, J.A.; Raad, I.I.; Marr, K.A.; Patterson, T.F.; Kontoyiannis, D.P.; Cornely, O.A.; Bow, E.J.; Rahav, G.; Neofytos, D.; Aoun, M.; et al. Isavuconazole versus voriconazole for primary treatment of invasive mould disease caused by Aspergillus and other filamentous fungi (SECURE): A phase 3, randomised-controlled, non-inferiority trial. Lancet 2016, 387, 760–769. [Google Scholar] [CrossRef]
- Kullberg, B.J.; Viscoli, C.; Pappas, P.G.; Vazquez, J.; Ostrosky-Zeichner, L.; Rotstein, C.; Sobel, J.D.; Herbrecht, R.; Rahav, G.; Jaruratanasirikul, S.; et al. Isavuconazole versus caspofungin in the treatment of candidemia and other invasive Candida infections: The ACTIVE trial. Clin. Infect. Dis. 2019, 68, 1981–1989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellsworth, M.; Ostrosky-Zeichner, L. Isavuconazole: Mechanism of action, clinical efficacy, and resistance. J. Fungi 2020, 6, 324. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Lee, H.; Ko, H.J.; Woo, E.R.; Lee, D.G. Fungicidal effect of isoquercitrin via inducing membrane disturbance. Biochim. Biophys. Acta 2015, 1848, 695–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, P.; Bretagne, S.; Gantier, J.C.; Garcia-Hermoso, D.; Lortholary, O.; Dromer, F.; Dannaoui, E. Molecular identification of zygomycetes from culture and experimentally infected tissues. J. Clin. Microbiol. 2006, 44, 340–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidaud, A.L.; Schwarz, P.; Herbreteau, G.; Dannaoui, E. Techniques for the assessment of in vitro and in vivo antifungal combinations. J. Fungi 2021, 7, 113. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.C.; Meletiadis, J.; Mouton, J.W.; Lagrou, K.; Hamal, P.; Guinea, J. Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Yeasts (EUCAST Definitive Document E.Def 7.3.2). 2020. Available online: https://www.eucast.org/astoffungi/methodsinantifungalsusceptibilitytesting/susceptibility_testing_of_yeasts/ (accessed on 1 May 2022).
- Odds, F.C. Synergy, antagonism, and what the chequerboard puts between them. J. Antimicrob. Chemother. 2003, 52, 1. [Google Scholar] [CrossRef]
- Di Veroli, G.Y.; Fornari, C.; Wang, D.; Mollard, S.; Bramhall, J.L.; Richards, F.M.; Jodrell, D.I. Combenefit: An interactive platform for the analysis and visualization of drug combinations. Bioinformatics 2016, 32, 2866–2868. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [Green Version]
- Beckman, C.H. Phenolic-storing cells: Keys to programmed cell death and periderm formation in wilt disease resistance and in general defence responses in plants? Physiol. Mol. Plant Pathol. 2000, 57, 101–110. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Cushnie, T.P.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Faustino, M.V.; Pinto, D.C.G.A.; Gonçalves, M.J.; Salgueiro, L.; Silveira, P.; Silva, A.M.S. Calendula L. species polyphenolic profile and in vitro antifungal activity. J. Funct. Foods 2018, 45, 254–267. [Google Scholar] [CrossRef]
- Foss, S.R.; Nakamura, C.V.; Ueda-Nakamura, T.; Cortez, D.A.; Endo, E.H.; Dias Filho, B.P. Antifungal activity of pomegranate peel extract and isolated compound punicalagin against dermatophytes. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 32. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Joya, J.A.; Pastrana-Castro, L.; Nieto-Oropeza, D.; Ventura-Sobrevilla, J.; Rojas-Molina, R.; Aguilar, C.N. The physicochemical, antifungal and antioxidant properties of a mixed polyphenol based bioactive film. Heliyon 2018, 4, e00942. [Google Scholar] [CrossRef] [Green Version]
- Ascacio-Valdes, J.; Burboa, E.; Aguilera-Carbo, A.F.; Aparicio, M.; Perez-Schmidt, R.; Rodriguez, R.; Aguilar, C.N. Antifungal ellagitannin isolated from Euphorbia antisyphilitica Zucc. Asian Pac. J. Trop. Biomed. 2013, 3, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Kanwal, Q.; Hussain, I.; Latif Siddiqui, H.; Javaid, A. Antifungal activity of flavonoids isolated from mango (Mangifera indica L.) leaves. Nat. Prod. Res. 2010, 24, 1907–1914. [Google Scholar] [CrossRef]
- Rongai, D.; Pulcini, P.; Pesce, B.; Milano, F. Antifungal activity of pomegranate peel extract against fusarium wilt of tomato. Eur. J. Plant Pathol. 2017, 147, 229–238. [Google Scholar] [CrossRef]
- Filho, A.A.O.; Oliveira, H.B.D.; de Sousa, J.P.; Meireles, D.R.P.; de Alzevedo Maia, G.L.; Filho, J.M.B.; de Siqueira Júnior, J.P.; de Oliveira Lima, E. In vitro anti-Candida activity and mechanism of action of the flavonoid isolated from Praxelis clematidea against Candida albicans species. J. Appl. Pharm. Sci. 2016, 6, 066–069. [Google Scholar] [CrossRef] [Green Version]
- Meragelman, T.L.; Tucker, K.D.; McCloud, T.G.; Cardellina, J.H., 2nd; Shoemaker, R.H. Antifungal flavonoids from Hildegardia barteri. J. Nat. Prod. 2005, 68, 1790–1792. [Google Scholar] [CrossRef]
- Nair, M.S.; Saxena, A.; Kaur, C. Characterization and antifungal activity of pomegranate peel extract and its use in polysaccharide-based edible coatings to extend the shelf-life of Capsicum (Capsicum annuum L.). Food Bioprocess Technol. 2018, 11, 1317–1327. [Google Scholar] [CrossRef]
- Yamaguchi, M.U.; Garcia, F.P.; Cortez, D.A.; Ueda-Nakamura, T.; Filho, B.P.; Nakamura, C.V. Antifungal effects of Ellagitannin isolated from leaves of Ocotea odorifera (Lauraceae). Antonie. Van. Leeuwenhoek 2011, 99, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Jin, Q.; Jin, H.G.; Ko, H.J.; Woo, E.R. Phenolic compounds with IL-6 inhibitory activity from Aster yomena. Arch Pharm. Res. 2014, 37, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Makino, T.; Kanemaru, M.; Okuyama, S.; Shimizu, R.; Tanaka, H.; Mizukami, H. Anti-allergic effects of enzymatically modified isoquercitrin (alpha-oligoglucosyl quercetin 3-O-glucoside), quercetin 3-O-glucoside, alpha-oligoglucosyl rutin, and quercetin, when administered orally to mice. J. Nat. Med. 2013, 67, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.; Woo, E.R.; Lee, D.G. Effect of isoquercitrin on membrane dynamics and apoptosis-like death in Escherichia coli. Biochim. Biophys. Acta Biomembr. 2018, 1860, 357–363. [Google Scholar] [CrossRef]
- Jayachandran, M.; Zhang, T.; Wu, Z.; Liu, Y.; Xu, B. Isoquercetin regulates SREBP-1C via AMPK pathway in skeletal muscle to exert antihyperlipidemic and anti-inflammatory effects in STZ induced diabetic rats. Mol. Biol. Rep. 2020, 47, 593–602. [Google Scholar] [CrossRef]
- Rogerio, A.P.; Kanashiro, A.; Fontanari, C.; da Silva, E.V.; Lucisano-Valim, Y.M.; Soares, E.G.; Faccioli, L.H. Anti-inflammatory activity of quercetin and isoquercitrin in experimental murine allergic asthma. Inflamm. Res. 2007, 56, 402–408. [Google Scholar] [CrossRef]
- Jung, S.H.; Kim, B.J.; Lee, E.H.; Osborne, N.N. Isoquercitrin is the most effective antioxidant in the plant Thuja orientalis and able to counteract oxidative-induced damage to a transformed cell line (RGC-5 cells). Neurochem. Int. 2010, 57, 713–721. [Google Scholar] [CrossRef]
- Yun, J.; Woo, E.R.; Lee, D.L. Isoquercitrin, isolated from Aster yomena triggers ROS-mediated apoptosis in Candida albicans. J. Funct. Foods 2016, 22, 347–357. [Google Scholar] [CrossRef]
- Desnos-Ollivier, M.; Bretagne, S.; Boullie, A.; Gautier, C.; Dromer, F.; Lortholary, O.; French Mycoses Study Group. Isavuconazole MIC distribution of 29 yeast species responsible for invasive infections (2015-2017). Clin. Microbiol. Infect. 2019, 25, 634.e1–634.e4. [Google Scholar] [CrossRef]
- Jorgensen, K.M.; Astvad, K.M.T.; Hare, R.K.; Arendrup, M.C. EUCAST susceptibility testing of isavuconazole: MIC data for contemporary clinical mold and yeast isolates. Antimicrob. Agents Chemother. 2019, 63, e00073-19. [Google Scholar] [CrossRef] [Green Version]
- Sickles, E.A.; Greene, W.H.; Wiernik, P.H. Clinical presentation of infection in granulocytopenic patients. Arch. Intern. Med. 1975, 135, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, M.; Kannan, A.; Stojkovic, D.S.; Glamoclija, J.; Calhelha, R.C.; Ferreira, I.; Sanglard, D.; Sokovic, M. Flavones, flavonols, and glycosylated derivatives-impact on Candida albicans growth and virulence, expression of CDR1 and ERG11, cytotoxicity. Pharmaceuticals 2020, 14, 27. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Li, H.; Zhang, W.; Lu, D.; Chen, Y.; Yin, J.; Meng, Y.; Ying, X.; Kang, T. Pharmacokinetic study of isoquercitrin in rat plasma after intravenous administration at three different doses. Braz. J. Pharm. Sci. 2013, 49, 435–441. [Google Scholar] [CrossRef] [Green Version]
- Kontoyiannis, D.P.; Lewis, R.E.; Alexander, B.D.; Lortholary, O.; Dromer, F.; Gupta, K.L.; John, G.T.; Del Busto, R.; Klintmalm, G.B.; Somani, J.; et al. Calcineurin inhibitor agents interact synergistically with antifungal agents in vitro against Cryptococcus neoformans isolates: Correlation with outcome in solid organ transplant recipients with cryptococcosis. Antimicrob. Agents Chemother. 2008, 52, 735–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Woo, E.R.; Lee, D.G. Synergistic antifungal activity of isoquercitrin: Apoptosis and membrane permeabilization related to reactive oxygen species in Candida albicans. IUBMB Life 2019, 71, 283–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castanheira, M.; Messer, S.A.; Rhomberg, P.R.; Dietrich, R.R.; Jones, R.N.; Pfaller, M.A. Isavuconazole and nine comparator antifungal susceptibility profiles for common and uncommon Candida species collected in 2012: Application of new CLSI clinical breakpoints and epidemiological cutoff values. Mycopathologia 2014, 178, 1–9. [Google Scholar] [CrossRef]
- Sanglard, D.; Coste, A.T. Activity of isavuconazole and other azoles against Candida clinical isolates and yeast model systems with known azole resistance mechanisms. Antimicrob. Agents Chemother. 2016, 60, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Pristov, K.E.; Ghannoum, M.A. Resistance of Candida to azoles and echinocandins worldwide. Clin. Microbiol. Infect. 2019, 25, 792–798. [Google Scholar] [CrossRef]


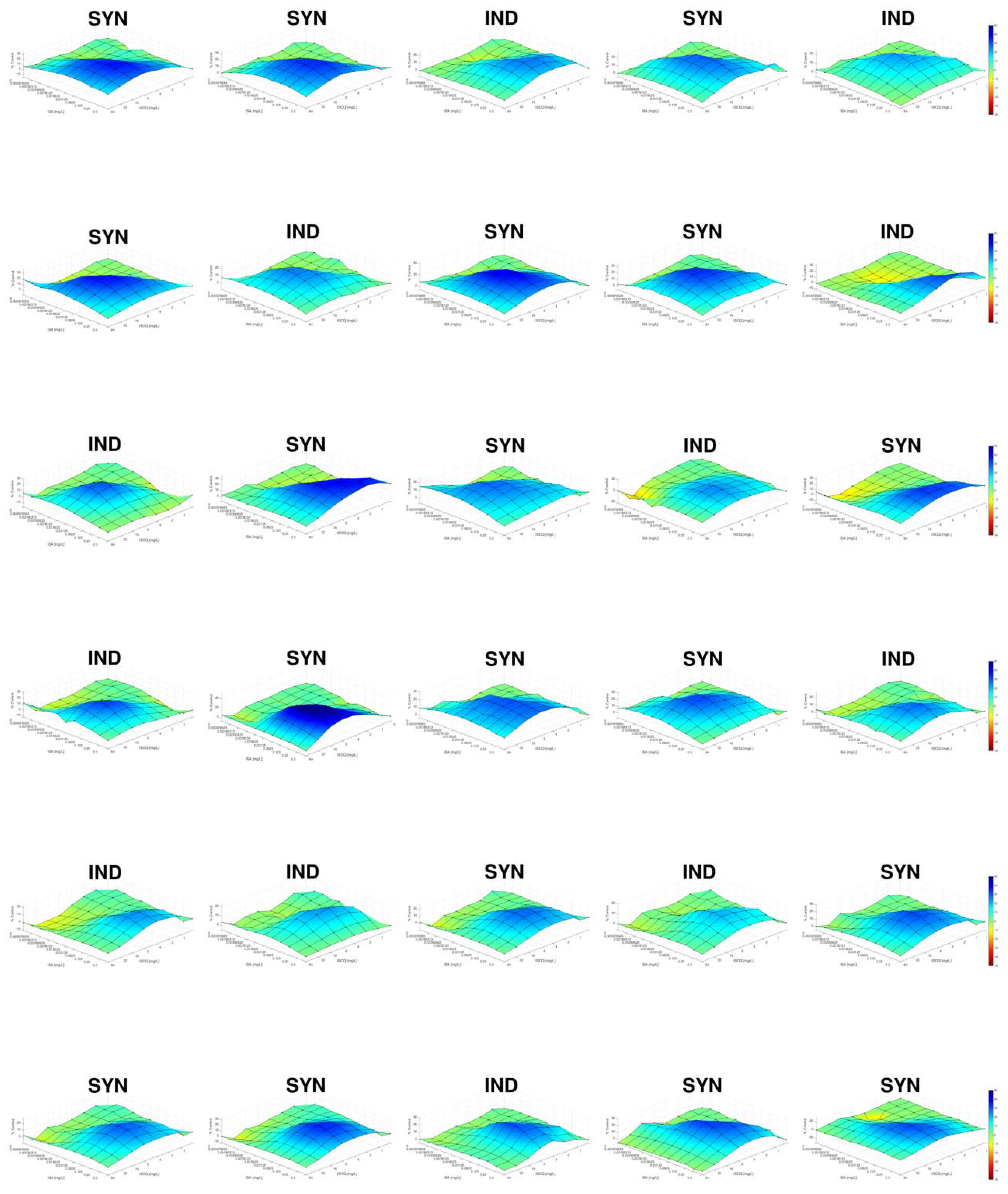{kind=link}
| Species | Collection Number | Checkerboard MICs (µg/mL) | Response Surface Analysis | |||||
|---|---|---|---|---|---|---|---|---|
| ISA | ISOQ | ISA/ISOQ | FICI | INTPN | ΣSYN-ANT (ΣSYN; ΣANT) | INTPN | ||
| C. albicans | V2105126 | 0.002 | 128 | 0.001/64 | 1 | IND | 17.47 (19.03; −1.56) | IND |
| C. albicans | N2101578 | 0.004 | 128 | 0.004/1 | 1.0078 | IND | −7.17 (8.40; −15.57) | IND |
| C. albicans | V2105568 | 0.001 | 64 | 0.00006/32 | 0.5625 | IND | 27.02 (27.98; −0.96) | IND |
| C. albicans | N2101577 | 0.002 | 128 | 0.002/1 | 1.0078 | IND | 34.95 (35.70; −0.75) | IND |
| C. albicans | V2105825iso3 | 0.001 | 32 | 0.0005/16 | 1 | IND | 22.97 (23.54; −0.57) | IND |
| C. albicans | ATCC 14053 | 0.002 | 32 | 0.001/1 | 0.5313 | IND | 16.83 (18.45; −1.62) | IND |
| C. albicans | V2105529 | 0.001 | 32 | 0.0005/8 | 0.75 | IND | 28.19 (28.66; −0.47) | IND |
| C. albicans | V2106139 | 0.002 | 64 | 0.00006/32 | 0.5313 | IND | 0.85 (10.26: −9.41) | IND |
| C. albicans | V2106041 | 0.001 | 128 | 0.001/1 | 1.0078 | IND | 11.79 (12.27; −0.48) | IND |
| C. albicans | V2106305 | 0.004 | 128 | 0.002/1 | 0.5078 | IND | 12.60 (13.34; −0.74) | IND |
| C. glabrata | V2105272 | 0.5 | 64 | 0.06/4 | 0.1875 | SYN | 93.42 (93.60; −0.18) | SYN |
| C. glabrata | V2105282 | 0.5 | 64 | 0.016/8 | 0.1563 | SYN | 84.19 (84.83; −0.64) | SYN |
| C. glabrata | N2101711 | 0.125 | 32 | 0.03/2 | 0.3125 | SYN | 55.66 (56.95; −1.29) | IND |
| C. glabrata | V2105636 | 0.125 | 32 | 0.016/4 | 0.25 | SYN | 70.11 (70.56; −0.45) | SYN |
| C. glabrata | DSM 70614 | 0.125 | 16 | 0.03/2 | 0.375 | SYN | 46.95 (48.26; −1.31) | IND |
| C. krusei | V2105825iso4 | 0.06 | 128 | 0.06/1 | 1.0078 | IND | 22.13 (24.96; −2.83) | IND |
| C. krusei | V2105866 | 0.06 | 128 | 0.06/1 | 1.0078 | IND | −38.29 (1.94; −40.23) | IND |
| C. krusei | V2106177 | 0.06 | 128 | 0.03/1 | 0.5078 | IND | 9.70 (19.83; −10.13) | IND |
| C. krusei | V2105920 | 0.06 | 128 | 0.03/1 | 0.5078 | IND | −4.50 (10.81; −15.31) | IND |
| C. krusei | ATCC 6258 | 0.06 | 128 | 0.03/2 | 0.5156 | IND | −30.31 (5.67; −35.98) | IND |
| C. parapsilosis | V2105056 | 0.008 | 128 | 0.008/1 | 1.0078 | IND | −1.10 (11.54; −12.64) | IND |
| C. parapsilosis | V2105223 | 0.008 | 128 | 0.004/16 | 0.625 | IND | −40.04 (9.22; −49.26) | IND |
| C. parapsilosis | B2107379 | 0.008 | 128 | 0.008/1 | 1.0078 | IND | 6.03 (13.39; −7.36) | IND |
| C. parapsilosis | ATCC 22019 | 0.016 | 128 | 0.016/1 | 1.0078 | IND | −8.18 (5.27; −13.45) | IND |
| C. tropicalis | V2105128 | 0.008 | 128 | 0.008/1 | 1.0078 | IND | −5.10 (4.14, −9.24) | IND |
| C. tropicalis | V2105245 | 0.008 | 128 | 0.008/1 | 1.0078 | IND | −9.39 (2.91; −12.30) | IND |
| C. tropicalis | V2105598 | 0.008 | 128 | 0.008/1 | 1.0078 | IND | 15.03 (16.16; −1.13) | IND |
| C. tropicalis | B1907975 | 0.008 | 128 | 0.008/1 | 1.0078 | IND | −2.05 (6.51; −8.56) | IND |
| C. tropicalis | V2106298 | 0.008 | 128 | 0.008/1 | 1.0078 | IND | −41.98 (1.63; −43.61) | IND |
| C. kefyr | V2106126 | 0.002 | 128 | 0.001/32 | 0.75 | IND | 7.71 (12.12; −4.41) | IND |
| C. kefyr | N2101899 | 0.0005 | 32 | 0.0002/8 | 0.75 | IND | 9.04 (17.00; −7.96) | IND |
| C. kefyr | N2102541 | 0.002 | 128 | 0.001/8 | 0.5625 | IND | 8.52 (18.35; −9.83) | IND |
| C. kefyr | V2107293 | 0.004 | 128 | 0.004/16 | 1.125 | IND | −4.30 (7.64; −11.94) | IND |
| C. kefyr | V2107534 | 0.0005 | 128 | 0.0005/2 | 1.0156 | IND | 10.99 (16.08; −5.09) | IND |
| C. kefyr | V2108462 | 0.002 | 128 | 0.002/2 | 1.0156 | IND | 10.42 (24.18; −13.76) | IND |
| Species | Collection Number | Checkerboard MICs (µg/mL) | Response Surface Analysis | |||||
|---|---|---|---|---|---|---|---|---|
| ISA | ISOQ | ISA/ISOQ | FICI | INTPN | ΣSYN-ANT (ΣSYN; ΣANT) | INTPN | ||
| C. glabrata | U2105834 | 0.25 | 32 | 0.03/4 | 0.25 | SYN | 81.95 (83.97; −2.02) | SYN |
| C. glabrata | V2105576 | 0.25 | 32 | 0.03/4 | 0.25 | SYN | 53.99 (56.11; −2.12) | IND |
| C. glabrata | N2102530 | 0.5 | 32 | 0.06/2 | 0.1875 | SYN | 95.80 (96.38; −0.58) | SYN |
| C. glabrata | U2106503 | 0.5 | 32 | 0.06/2 | 0.1875 | SYN | 82.48 (82.62; −0.14) | SYN |
| C. glabrata | U2106602 | 0.5 | 16 | 0.125/2 | 0.375 | SYN | 25.99 (37.29; −11.30) | IND |
| C. glabrata | U2106664 | 0.125 | 32 | 0.016/4 | 0.25 | SYN | 47.50 (48.14; −0.64) | IND |
| C. glabrata | U2106745 | 0.25 | 32 | 0.03/4 | 0.25 | SYN | 78.88 (81.92; −3.04) | SYN |
| C. glabrata | U2107113 | 0.25 | 64 | 0.03/2 | 0.1563 | SYN | 76.94 (77.70; −0.76) | SYN |
| C. glabrata | U2107210 | 0.125 | 64 | 0.03/4 | 0.3125 | SYN | 51.26 (54.21; −2.95) | IND |
| C. glabrata | U2107214 | 0.25 | 64 | 0.06/8 | 0.375 | SYN | 56.24 (61.55; −5.31) | SYN |
| C. glabrata | V2107409 | 0.25 | 64 | 0.03/8 | 0.25 | SYN | 55.26 (56.33; −1.07) | IND |
| C. glabrata | N2102703 | 0.5 | 64 | 0.06/4 | 0.1875 | SYN | 93.46 (94.17; −0.71) | SYN |
| C. glabrata | N2102712 | 1 | 64 | 0.06/4 | 0.125 | SYN | 87.70 (87.85; −0.15) | SYN |
| C. glabrata | N2102714 | 0.25 | 64 | 0.03/4 | 0.1875 | SYN | 77.59 (77.66; −0.07) | SYN |
| C. glabrata | U2107517 | 0.5 | 64 | 0.06/8 | 0.25 | SYN | 51.55 (53.74; −2.19) | IND |
| C. glabrata | U2107630 | 0.25 | 32 | 0.06/4 | 0.375 | SYN | 35.48 (39.20; −3.72) | IND |
| C. glabrata | U2107836 | 0.25 | 64 | 0.03/8 | 0.25 | SYN | 39.54 (40.73; −1.19) | IND |
| C. glabrata | V2108007 | 0.25 | 64 | 0.03/8 | 0.25 | SYN | 57.34 (58.43; −1.09) | SYN |
| C. glabrata | V2108459 | 0.25 | 64 | 0.06/2 | 0.2813 | SYN | 37.39 (39.26; −1.87) | IND |
| C. glabrata | B2109750 | 0.25 | 64 | 0.03/8 | 0.25 | SYN | 70.15 (70.26; −0.11) | SYN |
| C. glabrata | A2100553 | 0.25 | 64 | 0.03/8 | 0.25 | SYN | 60.71 (61.60; −0.89) | SYN |
| C. glabrata | U2107634 | 0.5 | 64 | 0.06/4 | 0.1875 | SYN | 72.52 (73.38; −0.86) | SYN |
| C. glabrata | U2107796 | 0.25 | 16 | 0.06/4 | 0.5 | SYN | 55.06 (55.47; −0.41) | IND |
| C. glabrata | U2108032 | 0.125 | 16 | 0.016/4 | 0.375 | SYN | 65.49 (66.03; −0.54) | SYN |
| C. glabrata | U2107634 | 0.5 | 32 | 0.06/2 | 0.1875 | SYN | 64.03 (65.77; −1.74) | SYN |
| Species (Strains), Interpretation Model | % of Strains with the Following Interaction | ||
|---|---|---|---|
| Synergy | Indifference | Antagonism | |
| C. albicans (10), FICI | 0 | 100 | 0 |
| C. albicans (10), RSA | 0 | 100 | 0 |
| C. glabrata (30), FICI | 100 | 00 | 0 |
| C. glabrata (30), RSA | 60 | 40 | 0 |
| C. krusei (5) FICI | 0 | 100 | 0 |
| C. krusei (5), RSA | 0 | 100 | 0 |
| C. parapsilosis (4), FICI | 0 | 100 | 0 |
| C. parapsilosis (4), RSA | 0 | 100 | 0 |
| C. tropicalis (5), FICI | 0 | 100 | 0 |
| C. tropicalis (5), RSA | 0 | 100 | 0 |
| C. kefyr (6), FICI | 0 | 100 | 0 |
| C. kefyr (6), RSA | 0 | 100 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwarz, P.V.; Nikolskiy, I.; Dannaoui, E.; Sommer, F.; Bange, G.; Schwarz, P. Synergistic In Vitro Interaction of Isavuconazole and Isoquercitrin against Candida glabrata. J. Fungi 2022, 8, 525. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050525
Schwarz PV, Nikolskiy I, Dannaoui E, Sommer F, Bange G, Schwarz P. Synergistic In Vitro Interaction of Isavuconazole and Isoquercitrin against Candida glabrata. Journal of Fungi. 2022; 8(5):525. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050525
Chicago/Turabian StyleSchwarz, Petra V., Ilya Nikolskiy, Eric Dannaoui, Frank Sommer, Gert Bange, and Patrick Schwarz. 2022. "Synergistic In Vitro Interaction of Isavuconazole and Isoquercitrin against Candida glabrata" Journal of Fungi 8, no. 5: 525. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050525