
The Search for Cryptic L-Rhamnosyltransferases on the Sporothrix schenckii Genome

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hardware and Software Environment Used
2.2. Sporothrix Schenckii Proteome and PFAM/TIGRFAM Hidden Markov Models Profiles
2.3. Construction of Ad Hoc Eukaryotic Rhamnosyltransferases HMM Profile and Hmmer Searches
2.4. In Silico Structural Analysis of Putative Rhamnosyltransferases Proteins
2.5. Strains and Culture Media
2.6. Analysis of Gene Expression
2.7. Heterologous Gene Expression in Escherichia coli and Protein Purification
2.8. Rhamnosyltransferase Assays
2.9. Statistical Analysis
3. Results
3.1. Identification of Putative Genes Encoding for Rahmosyltranferases
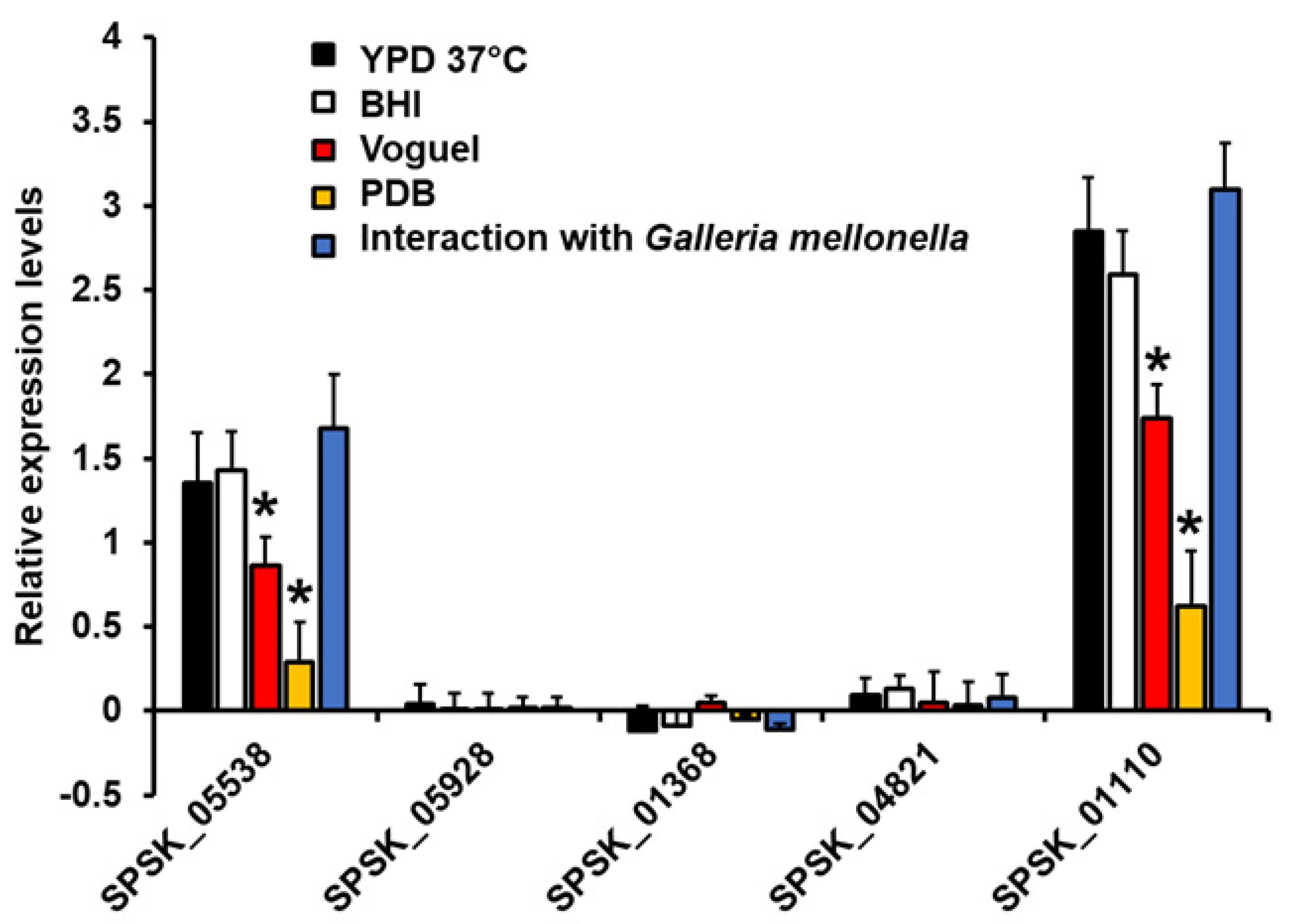
3.2. Expression Analysis of Five Putative Rhamnosyltransferases on Sporothrix Schenckii
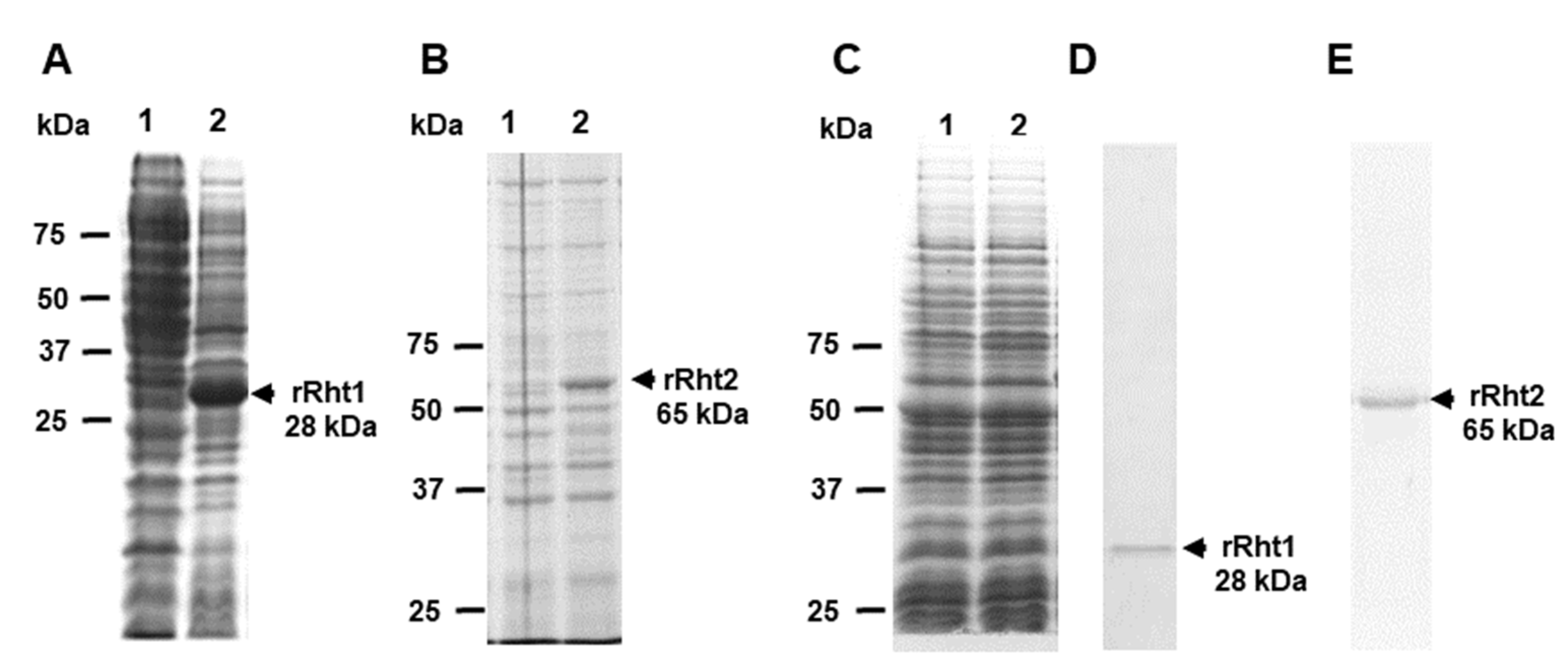
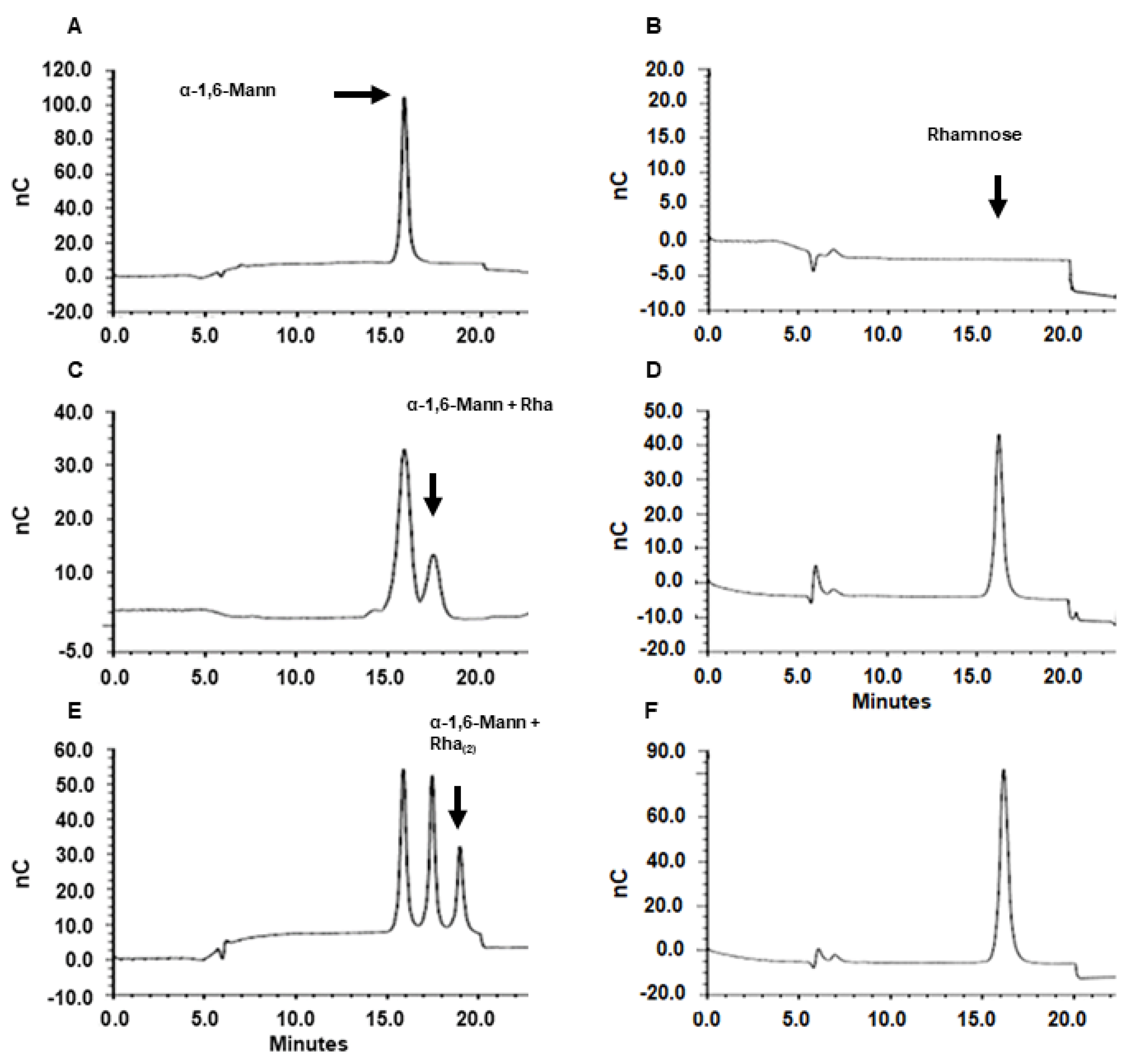
3.3. Sporothrix schenckii RHT1 and RHT2 Expression in Escherichia coli and Characterization of the Enzyme Activity
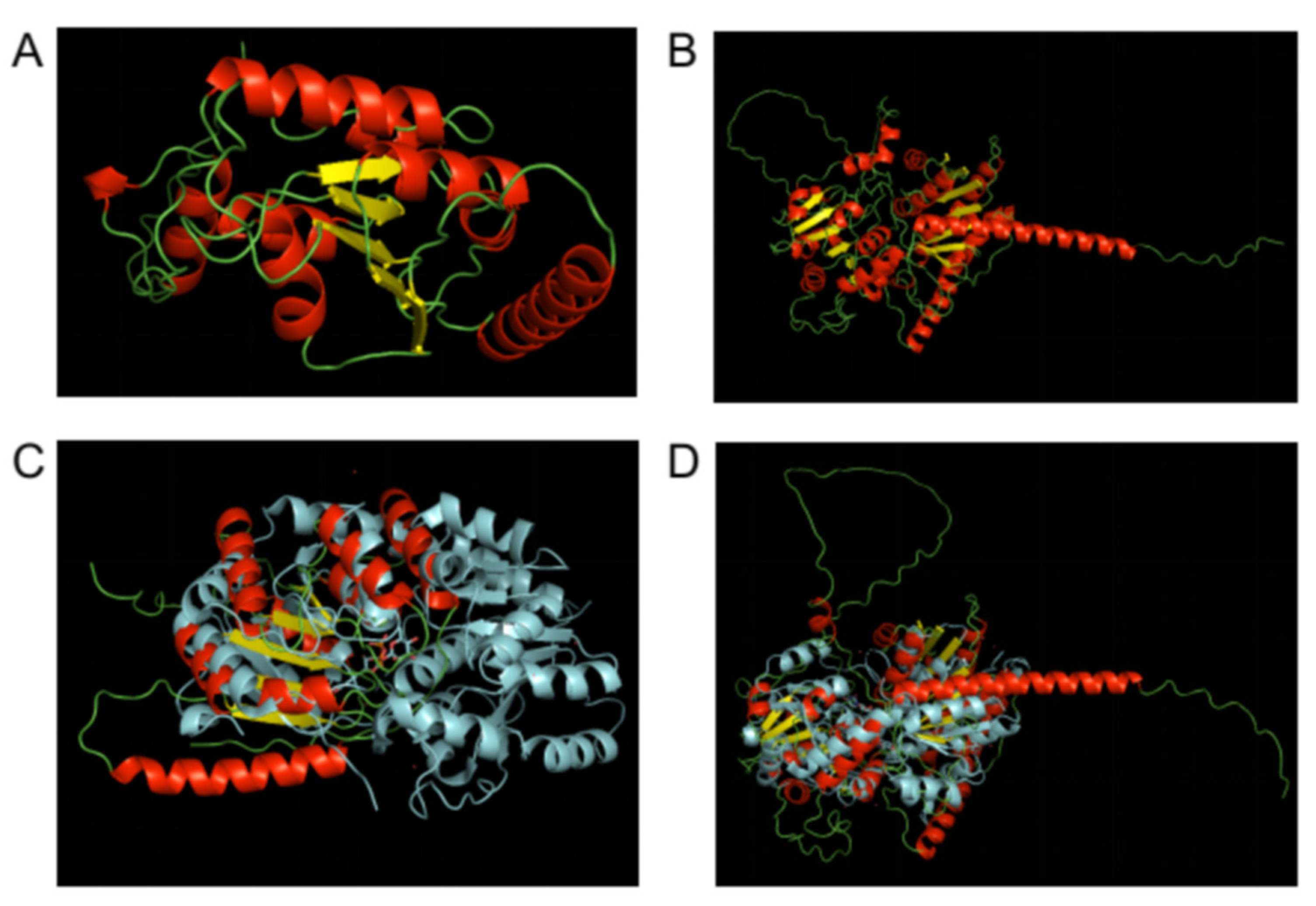
3.4. In Silico Structural Analysis of Sporothrix Schenckii Rht1 and Rht2 Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, M.C.; Gurr, S.J.; Cuomo, C.A.; Blehert, D.S.; Jin, H.; Stukenbrock, E.H.; Stajich, J.E.; Kahmann, R.; Boone, C.; Denning, D.W.; et al. Threats posed by the fungal kingdom to humans, wildlife, and agriculture. mBio 2020, 11, e00449-20. [Google Scholar] [CrossRef] [PubMed]
- Dalhoff, A. Does the use of antifungal agents in agriculture and food foster polyene resistance development? A reason for concern. J. Glob. Antimicrob. Resist. 2018, 13, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Brauer, V.S.; Rezende, C.P.; Pessoni, A.M.; De Paula, R.G.; Rangappa, K.S.; Nayaka, S.C.; Gupta, V.K.; Almeida, F. Antifungal agents in agriculture: Friends and foes of public health. Biomolecules 2019, 9, 521. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Bing, J.; Hu, T.; Ennis, C.L.; Nobile, C.J.; Huang, G. Candida auris: Epidemiology, biology, antifungal resistance, and virulence. PLoS Pathog. 2020, 16, e1008921. [Google Scholar] [CrossRef]
- Staniszewska, M. Virulence Factors in Candida species. Curr. Protein Pept. Sci. 2020, 21, 313–323. [Google Scholar] [CrossRef]
- Alspaugh, J.A. Virulence mechanisms and Cryptococcus neoformans pathogenesis. Fungal Genet. Biol. 2015, 78, 55–58. [Google Scholar] [CrossRef] [Green Version]
- Ardizzoni, A.; Wheeler, R.T.; Pericolini, E. It takes two to tango: How a dysregulation of the innate immunity, coupled with Candida virulence, triggers VVC onset. Front. Microbiol. 2021, 12, 692491. [Google Scholar] [CrossRef]
- Tamez-Castrellón, A.K.; Romeo, O.; García-Carnero, L.C.; Lozoya-Pérez, N.E.; Mora-Montes, H.M. Virulence factors in Sporothrix schenckii, one of the causative agents of sporotrichosis. Curr. Protein Pept. Sci. 2020, 21, 295–312. [Google Scholar] [CrossRef]
- Lopes-Bezerra, L.M.; Walker, L.A.; Niño-Vega, G.; Mora-Montes, H.M.; Neves, G.W.P.; Villalobos-Duno, H.; Barreto, L.; Garcia, K.; Franco, B.; Martínez-Álvarez, J.A.; et al. Cell walls of the dimorphic fungal pathogens Sporothrix schenckii and Sporothrix brasiliensis exhibit bilaminate structures and sloughing of extensive and intact layers. PLoS Negl. Trop. Dis. 2018, 12, e0006169. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Jiménez, D.F.; Pérez-García, L.A.; Martínez-Álvarez, J.A.; Mora-Montes, H.M. Role of the fungal cell wall in pathogenesis and antifungal resistance. Curr. Fungal Infect. Rep. 2012, 6, 275–282. [Google Scholar] [CrossRef]
- Klis, F.M.; de Groot, P.; Hellingwerf, K. Molecular organization of the cell wall of Candida albicans. Med. Mycol. 2001, 39 (Suppl. S1), 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latgé, J.-P.; Beauvais, A.; Chamilos, G. The cell wall of the human fungal pathogen Aspergillus fumigatus: Biosynthesis, organization, immune response, and virulence. Ann. Rev. Microbiol. 2017, 71, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Álvarez, J.A.; Pérez-García, L.A.; Mellado-Mojica, E.; López, M.G.; Martínez-Duncker, I.; Lópes-Bezerra, L.M.; Mora-Montes, H.M. Sporothrix schenckii sensu stricto and Sporothrix brasiliensis are differentially recognized by human peripheral blood mononuclear cells. Front. Microbiol. 2017, 8, 843. [Google Scholar] [CrossRef]
- Lozoya-Pérez, N.E.; Clavijo-Giraldo, D.M.; Martínez-Duncker, I.; García-Carnero, L.C.; López-Ramírez, L.A.; Niño-Vega, G.A.; Mora-Montes, H.M. Influences of the culturing media in the virulence and cell wall of Sporothrix schenckii, Sporothrix brasiliensis, and Sporothrix globosa. J. Fungi 2020, 6, 323. [Google Scholar] [CrossRef]
- Hoyer, L.L.; Cota, E. Candida albicans Agglutinin-Like Sequence (Als) family vignettes: A review of Als protein structure and function. Front. Microbiol. 2016, 7, 280. [Google Scholar] [CrossRef] [Green Version]
- Staniszewska, M.; Bondaryk, M.; Ochal, Z. Contribution of aspartic proteases in Candida virulence. protease inhibitors against candida infections. Curr. Protein Pept. Sci. 2017, 18, 1050–1062. [Google Scholar] [CrossRef]
- Lopes-Bezerra, L.M. Sporothrix schenckii cell wall peptidorhamnomannans. Front. Microbiol. 2011, 2, 243. [Google Scholar] [CrossRef] [Green Version]
- García-Carnero, L.C.; Salinas-Marín, R.; Lozoya-Pérez, N.E.; Wrobel, K.; Wrobel, K.; Martínez-Duncker, I.; Niño-Vega, G.A.; Mora-Montes, H.M. The heat shock protein 60 and Pap1 participate in the Sporothrix schenckii-host interaction. J. Fungi 2021, 7, 960. [Google Scholar] [CrossRef]
- Mora-Montes, H.M.; Ponce-Noyola, P.; Villagómez-Castro, J.C.; Gow, N.A.R.; Flores-Carreón, A.; López-Romero, E. Protein glycosylation in Candida. Future Microbiol. 2009, 4, 1167–1183. [Google Scholar] [CrossRef]
- Lozoya-Pérez, N.E.; Casas-Flores, S.; de Almeida, J.R.F.; Martínez-Álvarez, J.A.; López-Ramírez, L.A.; Jannuzzi, G.P.; Trujillo-Esquivel, E.; Estrada-Mata, E.; Almeida, S.R.; Franco, B.; et al. Silencing of OCH1 unveils the role of Sporothrix schenckii N-linked glycans during the host-fungus interaction. Infect. Drug Resist. 2019, 12, 67–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Nadales, E.; Nogueira, M.F.; Baldin, C.; Castanheira, S.; El Ghalid, M.; Grund, E.; Lengeler, K.; Marchegiani, E.; Mehrotra, P.V.; Moretti, M.; et al. Fungal model systems and the elucidation of pathogenicity determinants. Fungal Genet. Biol. 2014, 70, 42–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Gaviria, M.; Vargas-Macías, A.P.; García-Carnero, L.C.; Martínez-Duncker, I.; Mora-Montes, H.M. Role of protein glycosylation in interactions of medically relevant fungi with the host. J. Fungi 2021, 7, 875. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, P.; Boshoven, J.C.; Salas, O.; Bowler, K.; Islam, M.T.; Saber, M.K.; van den Berg, G.C.; Bar-Peled, M.; Thomma, B.P. Rhamnose synthase activity is required for pathogenicity of the vascular wilt fungus Verticillium dahliae. Mol. Plant Pathol. 2017, 18, 347–362. [Google Scholar] [CrossRef] [Green Version]
- Lopes, L.C.; da Silva, M.I.; Bittencourt, V.C.; Figueiredo, R.T.; Rollin-Pinheiro, R.; Sassaki, G.L.; Bozza, M.T.; Gorin, P.A.; Barreto-Bergter, E. Glycoconjugates and polysaccharides from the Scedosporium/Pseudallescheria boydii complex: Structural characterisation, involvement in cell differentiation, cell recognition and virulence. Mycoses 2011, 54 (Suppl. S3), 28–36. [Google Scholar] [CrossRef]
- Tamez-Castrellón, A.K.; van der Beek, S.L.; López-Ramírez, L.A.; Martínez-Duncker, I.; Lozoya-Pérez, N.E.; van Sorge, N.M.; Mora-Montes, H.M. Disruption of protein rhamnosylation affects the Sporothrix schenckii-host interaction. Cell Surf. 2021, 7, 100058. [Google Scholar] [CrossRef]
- Villalobos-Duno, H.L.; Barreto, L.A.; Alvarez-Aular, Á.; Mora-Montes, H.M.; Lozoya-Pérez, N.E.; Franco, B.; Lopes-Bezerra, L.M.; Niño-Vega, G.A. Comparison of cell wall polysaccharide composition and structure between strains of Sporothrix schenckii and Sporothrix brasiliensis. Front. Microbiol. 2021, 12, 726958. [Google Scholar] [CrossRef]
- Lopes-Bezerra, L.M.; Mora-Montes, H.M.; Zhang, Y.; Nino-Vega, G.; Rodrigues, A.M.; de Camargo, Z.P.; de Hoog, S. Sporotrichosis between 1898 and 2017: The evolution of knowledge on a changeable disease and on emerging etiological agents. Med. Mycol. 2018, 56, 126–143. [Google Scholar] [CrossRef]
- Mora-Montes, H.M.; Dantas Ada, S.; Trujillo-Esquivel, E.; de Souza Baptista, A.R.; Lopes-Bezerra, L.M. Current progress in the biology of members of the Sporothrix schenckii complex following the genomic era. FEMS Yeast Res. 2015, 15, fov065. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, M.M.; de Almeida, L.G.; Kubitschek-Barreira, P.; Alves, F.L.; Kioshima, E.S.; Abadio, A.K.; Fernandes, L.; Derengowski, L.S.; Ferreira, K.S.; Souza, R.C.; et al. Comparative genomics of the major fungal agents of human and animal Sporotrichosis: Sporothrix schenckii and Sporothrix brasiliensis. BMC Genom. 2014, 15, 943. [Google Scholar] [CrossRef] [Green Version]
- Bohl, T.; Bai, L.; Li, H. Recent progress in structural studies on the GT-C superfamily of protein glycosyltransferases. Sub-Cell. Biochem. 2021, 96, 259–271. [Google Scholar] [CrossRef]
- Breton, C.; Fournel-Gigleux, S.; Palcic, M.M. Recent structures, evolution and mechanisms of glycosyltransferases. Curr. Opin. Struct. Biol. 2012, 22, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Breton, C.; Snajdrová, L.; Jeanneau, C.; Koca, J.; Imberty, A. Structures and mechanisms of glycosyltransferases. Glycobiology 2006, 16, 29r–37r. [Google Scholar] [CrossRef] [PubMed]
- Giosa, D.; Felice, M.R.; Giuffrè, L.; Aiese Cigliano, R.; Paytuví-Gallart, A.; Lo Passo, C.; Barresi, C.; D’Alessandro, E.; Huang, H.; Criseo, G.; et al. Transcriptome-wide expression profiling of Sporothrix schenckii yeast and mycelial forms and the establishment of the Sporothrix Genome DataBase. Microb. Genom. 2020, 6, mgen000445. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2022, 50, D20–D26. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Burley, S.K.; Bhikadiya, C.; Bi, C.; Bittrich, S.; Chen, L.; Crichlow, G.V.; Christie, C.H.; Dalenberg, K.; Di Costanzo, L.; Duarte, J.M.; et al. RCSB Protein Data Bank: Powerful new tools for exploring 3D structures of biological macromolecules for basic and applied research and education in fundamental biology, biomedicine, biotechnology, bioengineering and energy sciences. Nucleic Acids Res. 2021, 49, D437–D451. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Dek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Almeida, A.; Beracochea, M.; Boland, M.; Burgin, J.; Cochrane, G.; Crusoe, M.R.; Kale, V.; Potter, S.C.; Richardson, L.J.; et al. MGnify: The microbiome analysis resource in 2020. Nucleic Acids Res. 2019, 48, D570–D578. [Google Scholar] [CrossRef]
- Mirdita, M.; von den Driesch, L.; Galiez, C.; Martin, M.J.; Sding, J.; Steinegger, M. Uniclust databases of clustered and deeply annotated protein sequences and alignments. Nucleic Acids Res. 2017, 45, D170–D176. [Google Scholar] [CrossRef] [PubMed]
- Mirdita, M.; Steinegger, M.; Sding, J. MMseqs2 desktop and local web server app for fast, interactive sequence searches. Bioinformatics 2019, 35, 2856–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold-Making protein folding accessible to all. bioRxiv 2022. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trujillo-Esquivel, E.; Martínez-Álvarez, J.A.; Clavijo-Giraldo, D.M.; Hernández, N.V.; Flores-Martínez, A.; Ponce-Noyola, P.; Mora-Montes, H.M. The Sporothrix schenckii gene encoding for the ribosomal protein L6 has constitutive and stable expression and works as an endogenous control in gene expression analysis. Front. Microbiol. 2017, 8, 1676. [Google Scholar] [CrossRef] [PubMed]
- Robledo-Ortiz, C.I.; Flores-Carreón, A.; Hernández-Cervantes, A.; Álvarez-Vargas, A.; Lee, K.K.; Díaz-Jiménez, D.F.; Munro, C.A.; Cano-Canchola, C.; Mora-Montes, H.M. Isolation and functional characterization of Sporothrix schenckii ROT2, the encoding gene for the endoplasmic reticulum glucosidase II. Fungal Biol. 2012, 116, 910–918. [Google Scholar] [CrossRef] [PubMed]
- Trujillo-Esquivel, E.; Franco, B.; Flores-Martínez, A.; Ponce-Noyola, P.; Mora-Montes, H.M. Purification of single-stranded cDNA based on RNA degradation treatment and adsorption chromatography. Nucleosides Nucleotides Nucleic Acids 2016, 35, 404–409. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Martínez-Álvarez, J.A.; García-Carnero, L.C.; Kubitschek-Barreira, P.H.; Lozoya-Pérez, N.E.; Belmonte-Vázquez, J.L.; de Almeida, J.R.; Gómez-Infante, A.d.J.; Curty, N.; Villagómez-Castro, J.C.; Peña-Cabrera, E.; et al. Analysis of some immunogenic properties of the recombinant Sporothrix schenckii Gp70 expressed in Escherichia coli. Future Microbiol. 2019, 14, 397–410. [Google Scholar] [CrossRef]
- Chevallet, M.; Luche, S.; Rabilloud, T. Silver staining of proteins in polyacrylamide gels. Nat. Protoc. 2006, 1, 1852–1858. [Google Scholar] [CrossRef] [Green Version]
- Yonekura-Sakakibara, K.; Tohge, T.; Niida, R.; Saito, K. Identification of a flavonol 7-O-rhamnosyltransferase gene determining flavonoid pattern in Arabidopsis by transcriptome coexpression analysis and reverse genetics. J. Biol. Chem. 2007, 282, 14932–14941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora-Montes, H.M.; López-Romero, E.; Zinker, S.; Ponce-Noyola, P.; Flores-Carreón, A. Hydrolysis of Man9GlcNAc2 and Man8GlcNAc2 oligosaccharides by a purified α-mannosidase from Candida albicans. Glycobiology 2004, 14, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Plaine, A.; Walker, L.; Da Costa, G.; Mora-Montes, H.M.; McKinnon, A.; Gow, N.A.; Gaillardin, C.; Munro, C.A.; Richard, M.L. Functional analysis of Candida albicans GPI-anchored proteins: Roles in cell wall integrity and caspofungin sensitivity. Fungal Genet. Biol. 2008, 45, 1404–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, N.V.; López-Ramírez, L.A.; Díaz-Jiménez, D.F.; Mellado-Mojica, E.; Martínez-Duncker, I.; López, M.G.; Mora-Montes, H.M. Saccharomyces cerevisiae KTR4, KTR5 and KTR7 encode mannosyltransferases differentially involved in the N- and O-linked glycosylation pathways. Res. Microbiol. 2017, 168, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, P.M.; Deleury, E.; Davies, G.J.; Henrissat, B. An evolving hierarchical family classification for glycosyltransferases. J. Mol. Biol. 2003, 328, 307–317. [Google Scholar] [CrossRef]
- Gloster, T.M. Advances in understanding glycosyltransferases from a structural perspective. Curr. Opin. Struct. Biol. 2014, 28, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Chang, A.; Singh, S.; Helmich, K.E.; Goff, R.D.; Bingman, C.A.; Thorson, J.S.; Phillips, G.N., Jr. Complete set of glycosyltransferase structures in the calicheamicin biosynthetic pathway reveals the origin of regiospecificity. Proc. Natl. Acad. Sci. USA 2011, 108, 17649–17654. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Weldeghiorghis, T.; Zhang, G.; Imperiali, B.; Prestegard, J.H. Solution structure of Alg13: The sugar donor subunit of a yeast N-acetylglucosamine transferase. Structure 2008, 16, 965–975. [Google Scholar] [CrossRef] [Green Version]
- Zong, G.; Fei, S.; Liu, X.; Li, J.; Gao, Y.; Yang, X.; Wang, X.; Shen, Y. Crystal structures of rhamnosyltransferase UGT89C1 from Arabidopsis thaliana reveal the molecular basis of sugar donor specificity for UDP-β-l-rhamnose and rhamnosylation mechanism. Plant J. Cell Mol. Biol. 2019, 99, 257–269. [Google Scholar] [CrossRef]
- Campbell, J.A.; Davies, G.J.; Bulone, V.; Henrissat, B. A classification of nucleotide-diphospho-sugar glycosyltransferases based on amino acid sequence similarities. Biochem. J. 1997, 326 Pt 3, 929–939. [Google Scholar] [CrossRef]
- Drula, E.; Garron, M.L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef] [PubMed]
- Wiggins, C.A.; Munro, S. Activity of the yeast MNN1 α-1,3-mannosyltransferase requires a motif conserved in many other families of glycosyltransferases. Proc. Natl. Acad. Sci. USA 1998, 95, 7945–7950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellokumpu, S.; Hassinen, A.; Glumoff, T. Glycosyltransferase complexes in eukaryotes: Long-known, prevalent but still unrecognized. Cell. Mol. Life Sci. CMLS 2016, 73, 305–325. [Google Scholar] [CrossRef] [PubMed]
- Grewal, R.K.; Shaikh, A.R.; Gorle, S.; Kaur, M.; Videira, P.A.; Cavallo, L.; Chawla, M. Structural insights in mammalian sialyltransferases and fucosyltransferases: We have come a long way, but it is still a long way down. Molecules 2021, 26, 5203. [Google Scholar] [CrossRef] [PubMed]
- Wagstaff, B.A.; Zorzoli, A.; Dorfmueller, H.C. NDP-rhamnose biosynthesis and rhamnosyltransferases: Building diverse glycoconjugates in nature. Biochem. J. 2021, 478, 685–701. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GENE Symbol | mRNA AccnNumber | GeneID | Protein AccnNumber | Gene Description | Length |
|---|---|---|---|---|---|
| SPSK_01368 | XM_016728283.1 | gi|27663560 | gb|XP_016590217.1 | Sterol 3-beta-glucosyltransferase mRNA | 1836 |
| SPSK_04821 | XM_016731584.1 | gi|27666861 | gb|XP_016583511.1 | UDP-transferase mRNA | 1415 |
| SPSK_05538 | XM_016732282.1 | gi|27667559 | gb|XP_016583713.1 | beta-1,4-N-acetylglucosaminyltransferase mRNA | 232 |
| SPSK_05928 | XM_016732636.1 | gi|27667913 | gb|XP_016591800.1 | glucosylltransferase family 28 protein mRNA | 920 |
| SPSK_01110 | XM_016728041.1 | gi|27663318 | gb|XP_016584143.1 | hypothetical protein mRNA | 576 |
| Seq ID | aln Start | aln End | hmm Acc | hmm Name | Bitscore | Indiv. E-Value | Cond. E-Value | Clan |
|---|---|---|---|---|---|---|---|---|
| SPSK_05538 | 16 | 192 | PF04101.19 | Glyco_tran_28_C | 57.35 | 2e-15 | 9.9e-20 | CL0113 |
| SPSK_05928 | 71 | 113 | PF03033.23 | Glyco_transf_28 | 32.35 | 9.2e-8 | 9.3e-12 | CL0113 |
| 479 | 549 | PF00201.21 | UDPGT | 21.39 | 1e-4 | 1e-8 | CL0113 | |
| SPSK_01368 | 1022 | 1128 | PF02893.23 | GRAM | 71.46 | 5.6e-20 | 8.5e-24 | CL0266 |
| 1237 | 1372 | PF03033.23 | Glyco_transf_28 | 99.87 | 1.3e-28 | 2e-32 | CL0113 | |
| 505 | 598 | PF00169.32 | PH | 48.85 | 8.5e-13 | 1.3e-16 | CL0266 | |
| SPSK_04821 | 415 | 576 | PF03033.23 | Glyco_transf_28 | 54.65 | 1.2e-14 | 1.2e-18 | CL0113 |
| 677 | 823 | PF00201.21 | UDPGT | 30.15 | 2.3e-7 | 2.3e-11 | CL0113 | |
| SPSK_01110 | 402 | 482 | PF00201.21 | UDPGT | 26.23 | 3.5e-6 | 1.8e-10 | CL0113 |
| Empty Vector | rRth1 | rRth2 | |
|---|---|---|---|
| No sugar donor | 0.03 ± 0.02 * | 0.01 ± 0.01 | 0.03 ± 0.02 |
| + UDP-Rhamnose | 0.08 ± 0.06 | 198.7 ± 34.4 ** | 215.4 ± 28.6 ** |
| + UDP-Glucose | 0.7 ± 0.4 | 0.9 ± 0.6 | 0.6 ± 0.4 |
| + GDP-Mannose | 1.5 ± 0.7 | 2.1 ± 1.4 | 1.5 ± 0.8 |
| + UDP-N-Acetylglucosamine | 0.3 ± 0.2 | 0.4 ± 0.2 | 0. 5 ± 0.3 |
| + UDP-Galactose | 0.5 ± 0.3 | 0.9 ± 0.7 | 0.7 ± 0.4 |
| + UDP-Rhamnose and heat-inactivated protein | 0.1 ± 0.09 | 0.1 ± 0.08 | 0.3 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mora-Montes, H.M.; García-Gutiérrez, K.; García-Carnero, L.C.; Lozoya-Pérez, N.E.; Ramirez-Prado, J.H. The Search for Cryptic L-Rhamnosyltransferases on the Sporothrix schenckii Genome. J. Fungi 2022, 8, 529. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050529
Mora-Montes HM, García-Gutiérrez K, García-Carnero LC, Lozoya-Pérez NE, Ramirez-Prado JH. The Search for Cryptic L-Rhamnosyltransferases on the Sporothrix schenckii Genome. Journal of Fungi. 2022; 8(5):529. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050529
Chicago/Turabian StyleMora-Montes, Héctor M., Karina García-Gutiérrez, Laura C. García-Carnero, Nancy E. Lozoya-Pérez, and Jorge H. Ramirez-Prado. 2022. "The Search for Cryptic L-Rhamnosyltransferases on the Sporothrix schenckii Genome" Journal of Fungi 8, no. 5: 529. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050529