
Co-Occurrence Patterns of Ustilago nuda and Pyrenophora graminea and Fungicide Contribution to Yield Gain in Barley under Fluctuating Climatic Conditions in Serbia



Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Origin
2.2. Field Trial
2.3. Disease and Fungicide Efficacy Assessments
- X—Number of plants infected with U. nuda or P. graminea in the seed-untreated plot;
- Y—Number of plants infected with U. nuda or P. graminea in the seed-treated plot.
2.4. Yield
- Y1—Maximum average grain yield in seed-treated plots obtained from four replications;
- Y2—Average grain yield in seed-untreated plots obtained from four replications.
2.5. Statistical Methods
3. Results
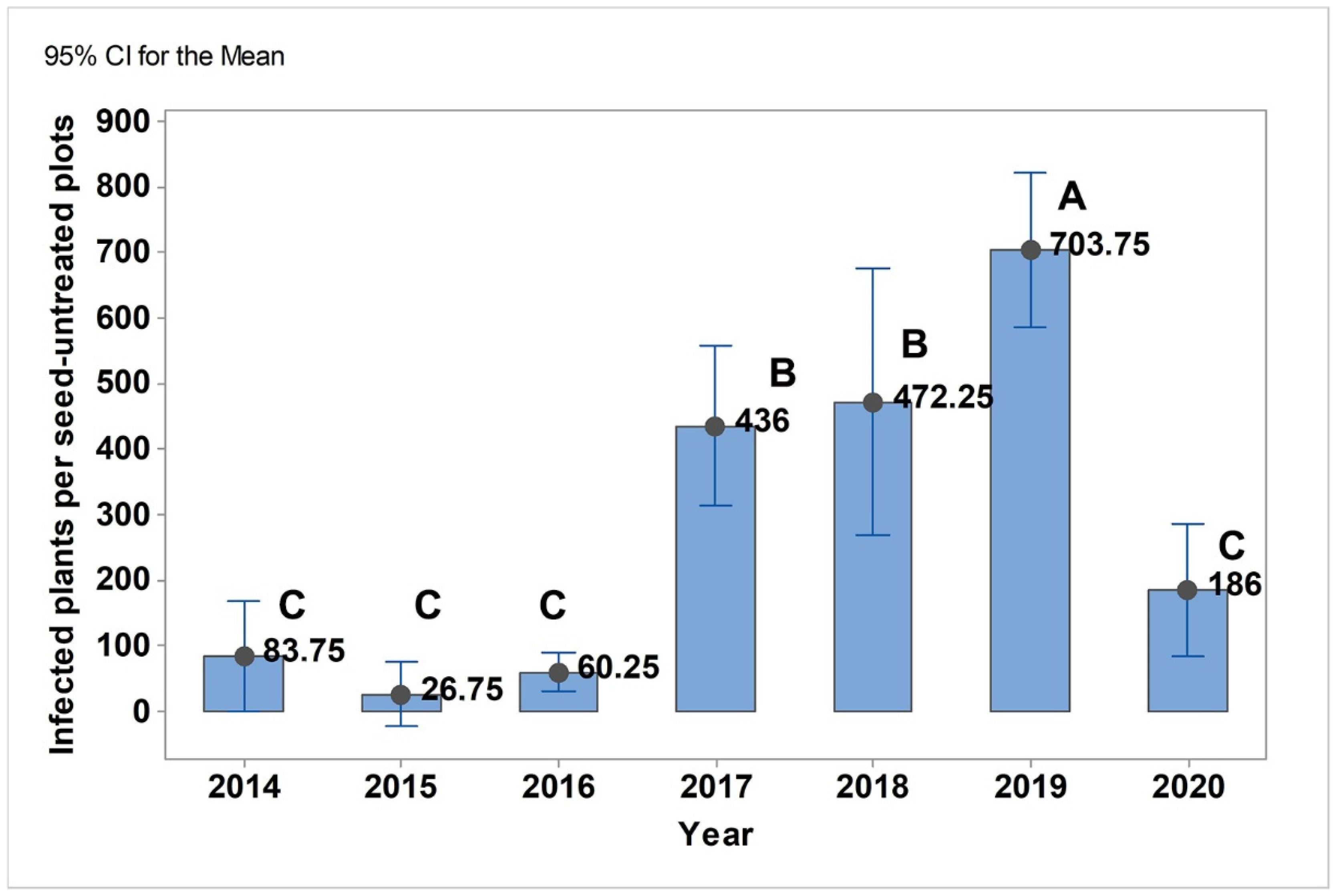
3.1. Co-Occurrence Patterns of U. nuda and P. graminea in Agro-Ecological Conditions in Serbia
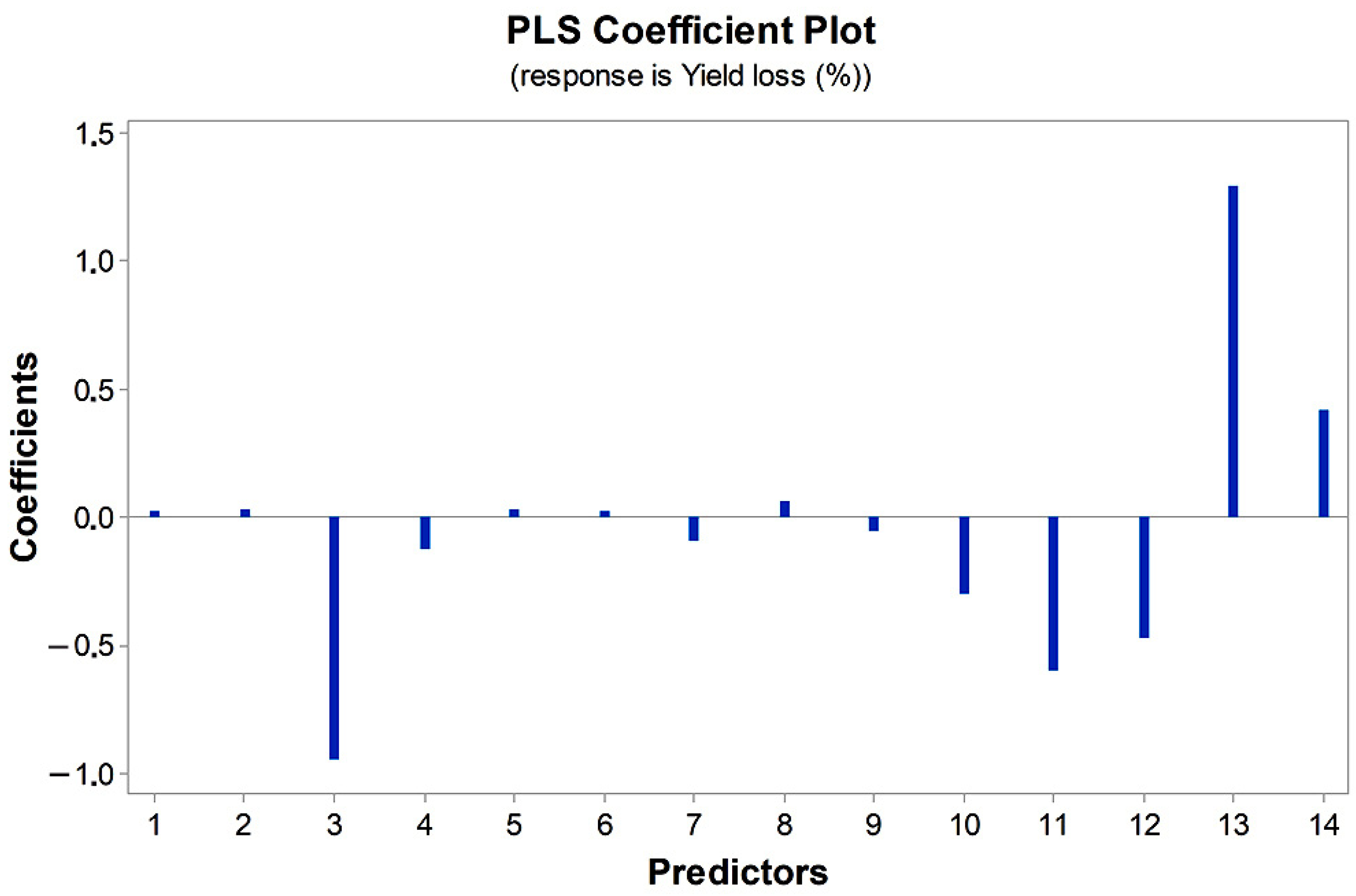
3.2. Combined Effect of Multiple Seed-Borne Diseases and Climatic Factors on Yield Loss in Seed-Untreated Plots
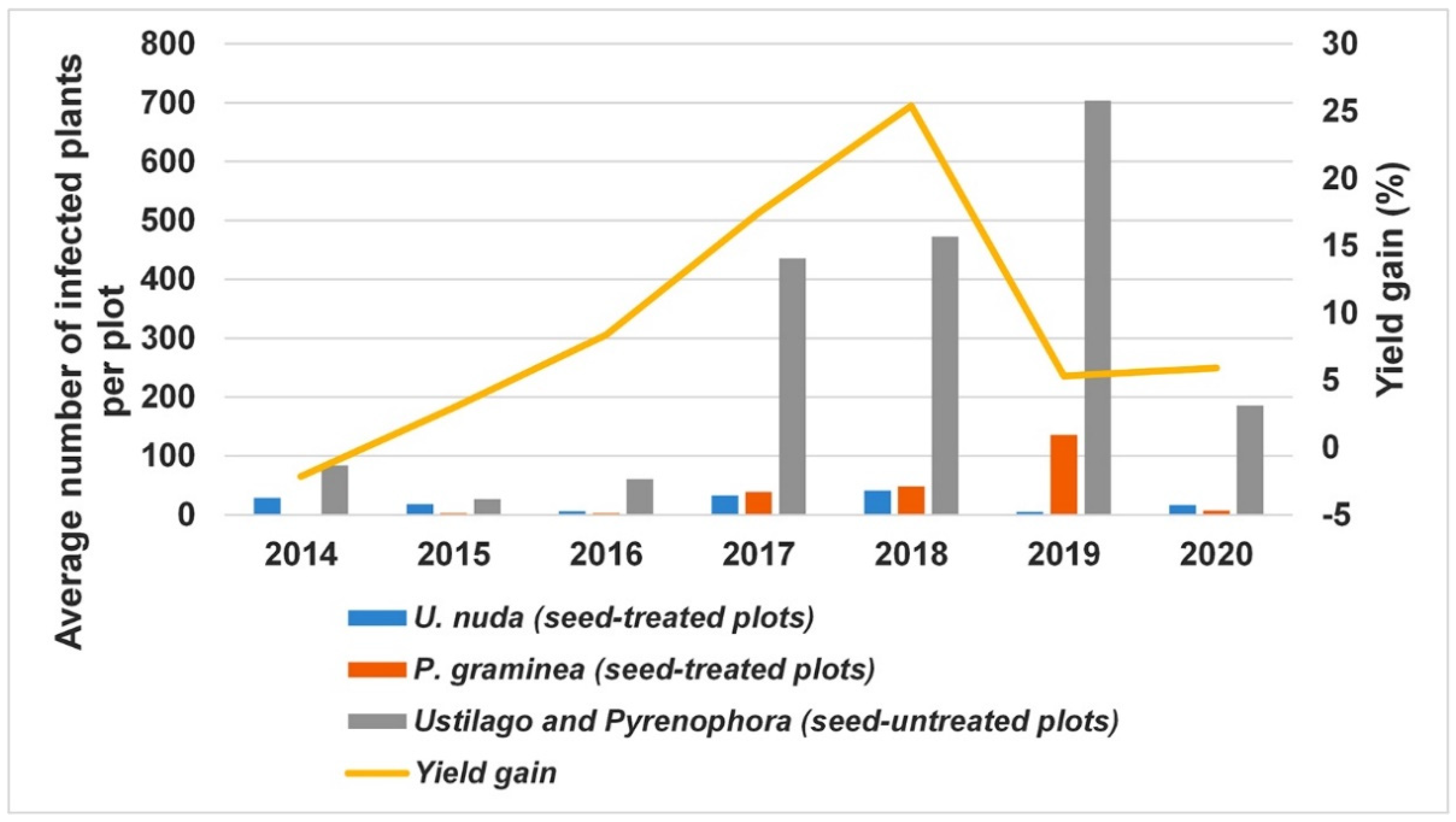
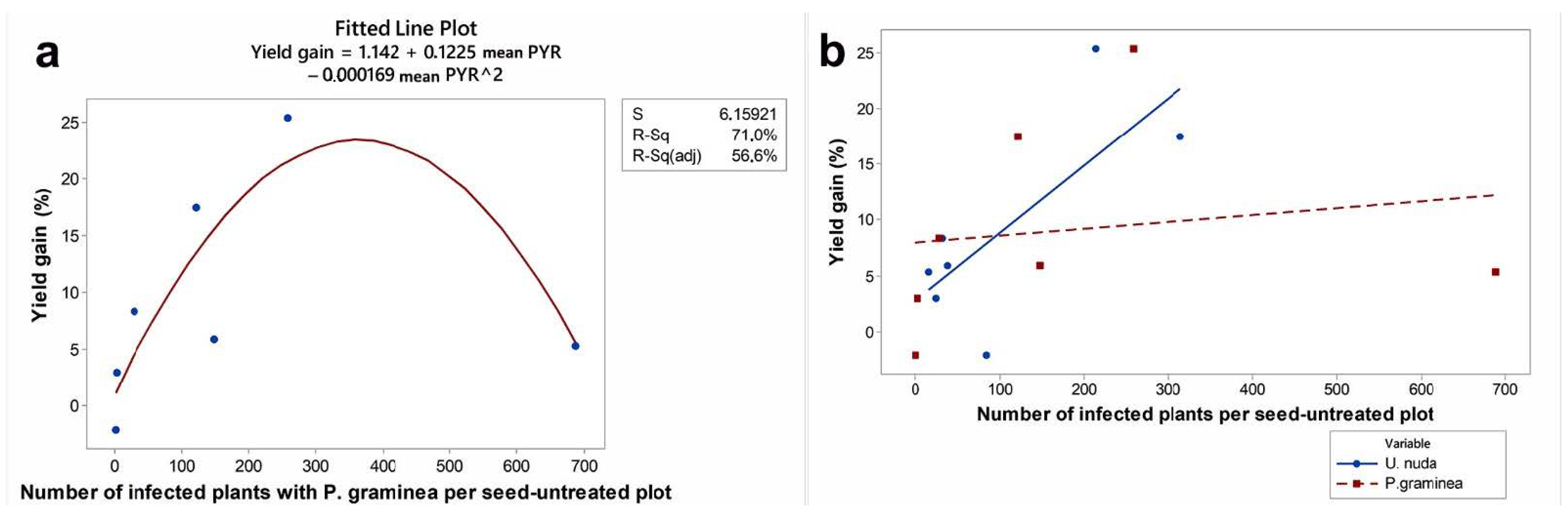
3.3. The Contribution of Fungicide Treatment to Yield Gain
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Oosterom, E.J.; Acevedo, E. Adaptation of Barley (Hordeum vulgare L.) to Harsh Mediterranean Environments. Euphytica 1992, 62, 1–14. [Google Scholar] [CrossRef]
- Baik, B.-K.; Ullrich, S.E. Barley for Food: Characteristics, Improvement, and Renewed Interest. J. Cereal Sci. 2008, 48, 233–242. [Google Scholar] [CrossRef]
- USDA Foreign Agricultural Services. Report Name: Grain and Feed Annual. Report Number: RB2020-0002. Available online: https://apps.fas.usda.gov/newgainapi/api/Report/DownloadReportByFileName?fileName=Grain%20and%20Feed%20Annual_Belgrade_Serbia_04-01-2020 (accessed on 1 October 2021).
- Wunderle, J.; Leclerque, A.; Schaffrath, U.; Slusarenko, A.; Koch, E. Assessment of the Loose Smut Fungi (Ustilago nuda and U. tritici) in Tissues of Barley and Wheat by Fluorescence Microscopy and Real-Time PCR. Eur. J. Plant Pathol. 2012, 133, 865–875. [Google Scholar] [CrossRef]
- Liu, J.; Huang, S.; Sun, M.; Liu, S.; Liu, Y.; Wang, W.; Zhang, X.; Wang, H.; Hua, W. An Improved Allele-Specific PCR Primer Design Method for SNP Marker Analysis and Its Application. Plant Methods 2012, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OEPP/EPPO. EPPO Standard PP 2/11(1) Guideline on Good Plant Protection Practice: Principles of Good Plant Protection Practice. Barley. Bullerin OEPP/EPPO Bull. 1997, 27, 339–361. [Google Scholar]
- Platenkamp, R. Investigation on the Infection Pathway of Drechslera graminea in Germinating Barley. K. Vet.-Og Landbohoeiskoles Aarsskrift 1976, 49–64. [Google Scholar]
- Bembelkacem, A.; Ceccarelli, S.; Amri, A.; Boulif, M. Variation in the Pathogenicity of 20 Algerian Isolates of “Pyrenophora graminea” Ito e Kur. on Nine Barley (“Hordeum vulgare” L.) Varieties. Var. Pathog. 2000, 1000–1007. [Google Scholar]
- Lamichhane, J.R.; You, M.P.; Laudinot, V.; Barbetti, M.J.; Aubertot, J.-N. Revisiting Sustainability of Fungicide Seed Treatments for Field Crops. Plant Dis. 2020, 104, 610–623. [Google Scholar] [CrossRef] [Green Version]
- Arabi, M.I.E.; Jawhar, M.; Al-Safadi, B.; Mirali, N. Yield Responses of Barley to Leaf Stripe (Pyrenophora graminea) under Experimental Conditions in Southern Syria. J. Phytopathol. 2004, 152, 519–523. [Google Scholar] [CrossRef]
- Tekauz, A.; Chiko, A.W. Leaf Stripe of Barley Caused by Pyrenophora graminea: Occurrence in Canada and Comparisons with Barley Stripe Mosaic. Can. J. Plant Pathol. 1980, 2, 152–158. [Google Scholar] [CrossRef]
- Heeb, L.; Jenner, E.; Cock, M.J.W. Climate-Smart Pest Management: Building Resilience of Farms and Landscapes to Changing Pest Threats. J. Pest Sci. 2019, 92, 951–969. [Google Scholar] [CrossRef]
- Alexandratos, N.; Bruinsma, J. World Agriculture towards 2030/2050: The 2012 Revision; AgEcon Search: Minneapolis, MN, USA, 2012. [Google Scholar]
- Chakraborty, S.; Newton, A.C. Climate Change, Plant Diseases and Food Security: An Overview. Plant Pathol. 2011, 60, 2–14. [Google Scholar] [CrossRef]
- White, J.W.; Hoogenboom, G.; Kimball, B.A.; Wall, G.W. Methodologies for Simulating Impacts of Climate Change on Crop Production. Field Crops Res. 2011, 124, 357–368. [Google Scholar] [CrossRef] [Green Version]
- Juroszek, P.; von Tiedemann, A. Climate Change and Potential Future Risks through Wheat Diseases: A Review. Eur. J. Plant Pathol. 2013, 136, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Lamichhane, J.R. Parsimonious Use of Pesticide-Treated Seeds: An Integrated Pest Management Framework. Trends Plant Sci. 2020, 25, 1070–1073. [Google Scholar] [CrossRef]
- You, M.P.; Rui, T.; Barbetti, M.J. Plant Genotype and Temperature Impact Simultaneous Biotic and Abiotic Stress-Related Gene Expression in Pythium-Infected Plants. Plant Pathol. 2020, 69, 655–668. [Google Scholar] [CrossRef]
- Vukovic, A.; Vujadinovic, M.; Rendulic, S.; Djurdjevic, V.; Ruml, M.; Babic, V.; Popovic, D. Global Warming Impact on Climate Change in Serbia for the Period 1961–2100. Therm. Sci. 2018, 2018, 168. [Google Scholar] [CrossRef]
- OEPP/EPPO. PP 1/019(4). Seed-Borne Cereal Fungi 2020. Bull. OEPP/EPPO Bull. 2020, 1–5. [Google Scholar]
- Bursac, Z.; Gauss, C.H.; Williams, D.K.; Hosmer, D.W. Purposeful Selection of Variables in Logistic Regression. Source Code Biol. Med. 2008, 3, 17. [Google Scholar] [CrossRef] [Green Version]
- Bryant, R.R.M.; McGrann, G.R.D.; Mitchell, A.R.; Schoonbeek, H.; Boyd, L.A.; Uauy, C.; Dorling, S.; Ridout, C.J. A Change in Temperature Modulates Defence to Yellow (Stripe) Rust in Wheat Line UC1041 Independently of Resistance Gene Yr36. BMC Plant Biol. 2014, 14, 10. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, L.K.; Jensen, J.D.; Nielsen, G.C.; Jensen, J.E.; Spliid, N.H.; Thomsen, I.K.; Justesen, A.F.; Collinge, D.B.; Jørgensen, L.N. Fusarium Head Blight of Cereals in Denmark: Species Complex and Related Mycotoxins. Phytopathology 2011, 101, 960–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woldemichael, M. D Importance, Biology, Epidemiology, and Management of Loose Smut (Ustilago nuda) of Barley (Hordeum vulgare): A Review. East Afr. J. Sci. 2019, 13, 89–108. [Google Scholar]
- Danko, J.; Michalikova, A. Influence of Some External Factors on the Germination of Chlamydospores of Ustilago tritici (Pers.). Jens Pol’nohospod. 1969, 15, 124–130. [Google Scholar]
- Tekauz, A.; Harper, F.R.; Davidson, J.G.N. Effect of Date of Seeding and Seed Treatment Fungicides on Infection of Barley by Pyrenophora graminea. Can. J. Plant Pathol. 1985, 7, 408–416. [Google Scholar] [CrossRef]
- Faccini, N.; Delbono, S.; Çelik Oğuz, A.; Cattivelli, L.; Valè, G.; Tondelli, A. Resistance of European Spring 2-Row Barley Cultivars to Pyrenophora graminea and Detection of Associated Loci. Agronomy 2021, 11, 374. [Google Scholar] [CrossRef]
- Glazebrook, J. Contrasting Mechanisms of Defense Against Biotrophic and Necrotrophic Pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Kissoudis, C.; van de Wiel, C.; Visser, R.G.F.; van der Linden, G. Enhancing Crop Resilience to Combined Abiotic and Biotic Stress through the Dissection of Physiological and Molecular Crosstalk. Front. Plant Sci. 2014, 5, 207. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, M.; Ishikawa, A.; Jikumaru, Y.; Seki, M.; Umezawa, T.; Asami, T.; Maruyama-Nakashita, A.; Kudo, T.; Shinozaki, K.; Yoshida, S.; et al. Antagonistic Interaction between Systemic Acquired Resistance and the Abscisic Acid–Mediated Abiotic Stress Response in Arabidopsis. Plant Cell 2008, 20, 1678–1692. [Google Scholar] [CrossRef] [Green Version]
- Jevtić, R.; Župunski, V.; Lalošević, M.; Župunski, L. Predicting Potential Winter Wheat Yield Losses Caused by Multiple Disease Systems and Climatic Conditions. Crop Prot. 2017, 99, 17–25. [Google Scholar] [CrossRef]
- Dawood, M.F.A.; Moursi, Y.S.; Amro, A.; Baenziger, P.S.; Sallam, A. Investigation of Heat-Induced Changes in the Grain Yield and Grains Metabolites, with Molecular Insights on the Candidate Genes in Barley. Agronomy 2020, 10, 1730. [Google Scholar] [CrossRef]
- Wiegmann, M.; Maurer, A.; Pham, A.; March, T.J.; Al-Abdallat, A.; Thomas, W.T.B.; Bull, H.J.; Shahid, M.; Eglinton, J.; Baum, M.; et al. Barley Yield Formation under Abiotic Stress Depends on the Interplay between Flowering Time Genes and Environmental Cues. Sci. Rep. 2019, 9, 6397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradhan, G.P.; Prasad, P.V.V.; Fritz, A.K.; Kirkham, M.B.; Gill, B.S. Effects of Drought and High Temperature Stress on Synthetic Hexaploid Wheat. Funct. Plant Biol. 2012, 39, 190–198. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Z.; Tao, F.; Palosuo, T.; Rötter, R.P. Impacts of Heat Stress on Leaf Area Index and Growth Duration of Winter Wheat in the North China Plain. Field Crops Res. 2018, 222, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Von Schmeling, B. Kulka Marshall Systemic Fungicidal Activity of 1,4-Oxathiin Derivatives. Science 1966, 152, 659–660. [Google Scholar] [CrossRef] [PubMed]
- Menzies, J.G. Carboxin Tolerant Strains of Ustilago Nuda and Ustilago Tritici in Canada. Can. J. Plant Pathol. 2008, 30, 498–502. [Google Scholar] [CrossRef]
- Menzies, J.; McLeod, R.; Tosi, L.; Cappelli, C. Occurrence of a Carboxin-Resistant Strain of Ustilago nuda in Italy. Phytopathol. Mediterr. 2005, 44, 216–219. [Google Scholar] [CrossRef]
- Magan, N.; Medina, A.; Aldred, D. Possible Climate-Change Effects on Mycotoxin Contamination of Food Crops Pre- and Postharvest. Plant Pathol. 2011, 60, 150–163. [Google Scholar] [CrossRef]
- Rodrigo, S.; Cuello-Hormigo, B.; Gomes, C.; Santamaria, O.; Costa, R.; Poblaciones, M.J. Influence of Fungicide Treatments on Disease Severity Caused by Zymoseptoria tritici, and on Grain Yield and Quality Parameters of Bread-Making Wheat under Mediterranean Conditions. Eur. J. Plant Pathol. 2015, 141, 99–109. [Google Scholar] [CrossRef]
- Benin, G.; Storck, L.; Marchioro, V.; Bornhofen, E.; Woyann, L.; Trevizan, D. Environment-Specific Selection to Identify High Yielding Wheat Genotypes and Response to Fungicide Application. Rev. Ceres 2017, 64, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Teng, P.S. Construction of Predictive Models: II. Forecasting Crop Losses. Adv. Plant Pathol. 1985, 3, 179–206. [Google Scholar]
- Hitaj, C.; Smith, D.J.; Code, A.; Wechsler, S.; Esker, P.D.; Douglas, M.R. Sowing Uncertainty: What We Do and Don’t Know about the Planting of Pesticide-Treated Seed. BioScience 2020, 70, 390–403. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Heading Date | Flowering Date (Period When Infection Occurred) * | Sowing Date | Year | Harvest Date |
|---|---|---|---|---|---|
| 2013 | 30 April | 3 May | 23 October | 2014 | 19 June |
| 2014 | 23 April | 26 April | 4 November | 2015 | 16 June |
| 2015 | 29 April | 2 May | 30 October | 2016 | 23 June |
| 2016 | 28 April | 1 May | 25 October | 2017 | 17 June |
| 2017 | 28 April | 1 May | 19 October | 2018 | 7 June |
| 2018 | 25 April | 28 April | 23 October | 2019 | 28 June |
| Treatments | Dosage Rate (Amount/1 kg of Seed) | |
|---|---|---|
| T1 | Seed-untreated plot (Control) | |
| T2 | RAXIL S 040 FS (20 g/L tebuconazole + 20 g/L triazoxide) | 1 mL |
| T3 | VITAVAX 200 FF (200 g/L carboxin + 200 g/L thiram) | 3 mL |
| T4 | MANKOGAL S (600 g/kg mancozeb) | 2 g |
| T5 | VIBRANCE DUO (25 g/L fludioxonil +25 g/L sedaxane) | 2 mL |
| T6 | RANCONA TRIO (5 g/L ipconazole + 13.3 g/L metalaxyl + 133 g/L carboxin) | 1 mL |
| T7 | CERTICOR 050 FS (30 g/L tebuconazole + 20 g/L metalaxyl -M) | 1 mL |
| T8 | CELEST EXTRA 050 FS (25 g/L fludioxonil + 25 g/L difenoconazole) | 2 mL |
| T9 | DIVIDEND EXTREME 115 FS (A12532C) (7.73% difenoconazole + 1.93% metalaxyl -M) | 1.5 mL |
| T10 | CELEST TOP 312.5 FS (262.5 g/L thiamethoxam + 25 g/L difenoconazole + 25 g/L fludioxonil) | 1.5 mL |
| T11 | YUNTA QUATTRO (6.7 g/L tebuconazole +33.3 g/L prothioconazole + 166.7 g/L imidacloprid +166.7 g/L clothianidin | 1.8 mL |
| T12 | VIAL TRUST FS (60 g/L tebuconazole+ 80 g/L thiabendazole) | 4 mL |
| T13 | LAMARDOR FS 400 (150 g/L tebuconazole + 250 g/L prothioconazole) | 0.2 mL |
| T14 | LAMARDOR FS 400 (150 g/L tebuconazole + 250 g/L prothioconazole) + GAUCHO 600 FS (600 g/L imidacloprid) | 0.2 + 1.7 mL |
| Source | DF | Adj SS | Adj MS | F Value | p Value |
|---|---|---|---|---|---|
| U. nuda | |||||
| Regression | 4 | 321,150 | 80,288 | 72.20 | 0.000 |
| Rainfall (flowering time in previous season) | 1 | 64,158 | 64,158 | 57.70 | 0.000 |
| Temperature (flowering time in previous season) | 1 | 22,899 | 22,899 | 20.59 | 0.000 |
| Humidity (flowering time in previous season) | 1 | 10,438 | 10,438 | 9.39 | 0.006 |
| Temperature in November | 1 | 187,980 | 187,980 | 169.04 | 0.000 |
| Error | 23 | 25,577 | 1112 | ||
| Total | 27 | 346,727 | |||
| P. graminea | |||||
| Regression | 5 | 1,418,983 | 283,797 | 103.80 | 0.000 |
| Rainfall (flowering time in previous growing season) | 1 | 193,806 | 193,806 | 7089 | 0.000 |
| Temperature (flowering time in previous season) | 1 | 713,053 | 713,053 | 260.81 | 0.000 |
| Humidity (flowering time in previous season) | 1 | 350,595 | 350,595 | 128.23 | 0.000 |
| Temperature (sowing time) | 1 | 402,556 | 402,556 | 147.24 | 0.000 |
| Rainfall (sowing time) | 1 | 167,962 | 167,962 | 61.43 | 0.000 |
| Error | 22 | 60,149 | 2734 | ||
| Total | 27 | 1,479,131 |
| Number of Predictors | R2 | R2pred | Mallows’ Cp | S | Joined Occurrence of U. nuda and P. graminea | T in November (°C) | T in February (°C) | T in April (°C) |
|---|---|---|---|---|---|---|---|---|
| 1 | 47.9 | 39.3 | 30.7 | 5.3481 | X | |||
| 1 | 47.1 | 38.9 | 31.5 | 5.3861 | X | |||
| 2 | 70.8 | 63.0 | 9.1 | 4.0848 | X | X | ||
| 2 | 64.5 | 57.1 | 15.6 | 4.5034 | X | X | ||
| 3 | 77.7 | 71.2 | 4.0 | 3.6455 | X | X | X | |
| 3 | 74.0 | 65.2 | 7.8 | 3.9355 | X | X | X | |
| 4 | 78.6 | 70.7 | 5.0 | 3.6485 | X | X | X | X |
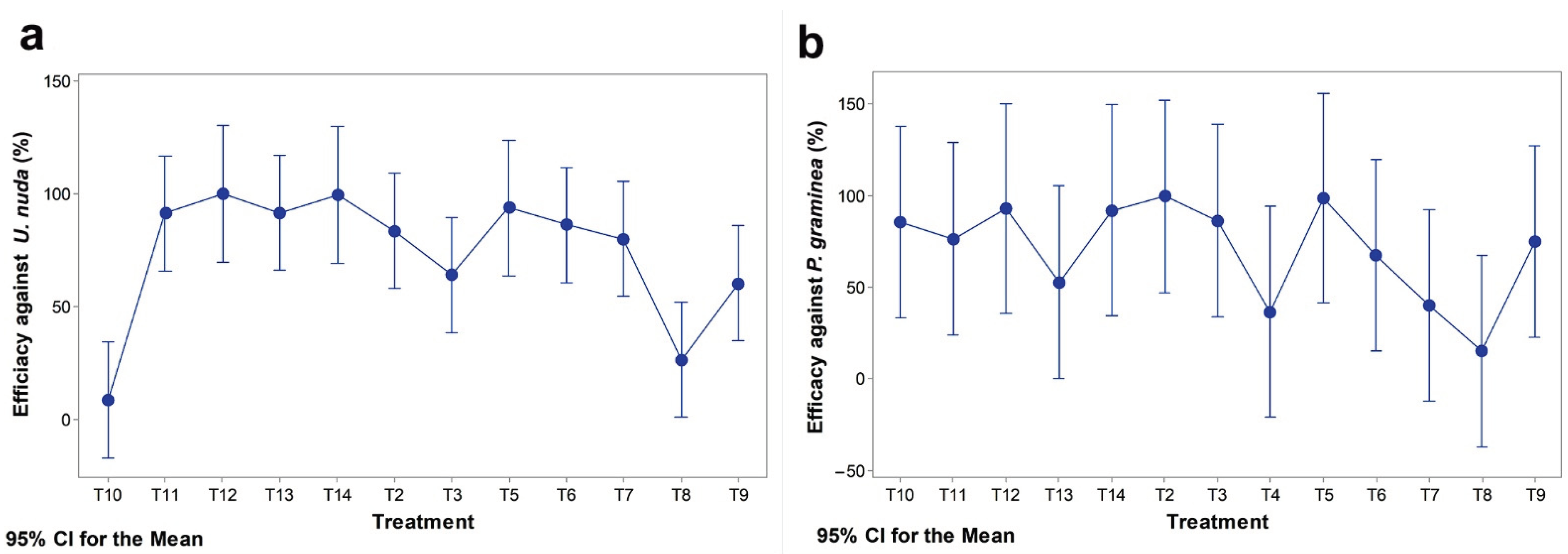
| Fungicide Treatment | Mean | Minimum | Maximum |
|---|---|---|---|
| U. nuda | |||
| T2 RAXIL S 040 FS (20 g/L tebuconazole +20 g/L triazoxide) | 83.5 A B | −4.1 | 100.0 |
| T3 VITAVAX 200 FF (200 g/L carboxin + 200 g/L thiram) | 64.1 A B C | −12.4 | 96.1 |
| T4 MANKOGAL S (600 g/kg mancozeb) | N/A * | ||
| T5 VIBRANCE DUO (25 g/L fludioxonil + 25 g/L sedaxane) | 93.80 A | 84.21 | 100.00 |
| T6 RANCONA TRIO (5 g/L ipconazole + 13.3 g/L metalaxyl + 133 g/L carboxin) | 86.29 A B | 60.82 | 100,00 |
| T7 CERTICOR 050 FS (30 g/L tebuconazole + 20 g/L metalaxyl -M) | 80.1 A B | −27.8 | 100.0 |
| T8 CELEST EXTRA 050 FS (25 g/L fludioxonil + 25 g/L difenoconazole) | 26.7 B C | −65.8 | 83.9 |
| T9 DIVIDEND EXTREME 115 FS (A12532C) (7.73% difenoconazole + 1.93% metalaxyl -M) | 60.4 A B C | −8.6 | 94.1 |
| T10 CELEST TOP 312.5 FS (262.5 g/L thiamethoxam + 25 g/L difenoconazole + 25 g/L fludioxonil) | 8.8 C | −59.7 | 70.6 |
| T11 YUNTA QUATTRO (6.7 g/L tebuconazole + 33.3 g/L prothioconazole + 166.7 g/L imidacloprid + 166.7 g/L clothianidin | 91.38 A | 69.74 | 100.00 |
| T12 VIAL TRUST FS (60 g/L tebuconazole + 80 g/L thiabendazole) | 100.00 A | 100.00 | 100.00 |
| T13 LAMARDOR FS 400 (150 g/L tebuconazole + 250 g/L prothioconazole) | 91.69 A | 43.30 | 100.00 |
| T14 LAMARDOR FS 400 (150 g/L tebuconazole + 250 g/L prothioconazole) + GAUCHO 600 FS (600 g/L imidacloprid) | 99.605 A | 98.026 | 100.000 |
| P. graminea | |||
| T2 RAXIL S 040 FS (20 g/L tebuconazole + 20 g/L triazoxide) | 99.488 A | 98.548 | 100.000 |
| T3 VITAVAX 200 FF (200 g/L carboxin + 200 g/L thiram) | 86.41 A | 53.18 | 100.00 |
| T4 MANKOGAL S (600 g/kg mancozeb) | 36.6 A | −39.0 | 85.0 |
| T5 VIBRANCE DUO (25 g/L fludioxonil + 25 g/L sedaxane) | 98.645 A | 94.869 | 100.000 |
| T6 RANCONA TRIO (5 g/L ipconazole + 13.3 g/L metalaxyl + 133 g/L carboxin) | 67.38 A | 50.00 | 86.99 |
| T7 CERTICOR 050 FS (30 g/L tebuconazole + 20 g/L metalaxyl -M) | 39.9 A | −80.0 | 87.2 |
| T8 CELEST EXTRA 050 FS (25 g/L fludioxonil + 25 g/L difenoconazole) | 15.1 A | −370.0 | 100.0 |
| T9 DIVIDEND EXTREME 115 FS (A12532C) (7.73% difenoconazole + 1.93% metalaxyl -M) | 74.7 A | −10.0 | 94.9 |
| T10 CELEST TOP 312.5 FS (262.5 g/L thiamethoxam + 25 g/L difenoconazole + 25 g/L fludioxonil) | 85.57 A | 60.00 | 98.62 |
| T11 YUNTA QUATTRO (6.7 g/L tebuconazole + 33.3 g/L prothioconazole + 166.7 g/L imidacloprid +166.7 g/L clothianidin | 76.45 A | 60.00 | 94.43 |
| T12 VIAL TRUST FS (60 g/L tebuconazole + 80 g/L thiabendazole) | 93.13 A | 80.90 | 98.23 |
| T13 LAMARDOR FS 400 (150 g/L tebuconazole + 250 g/L prothioconazole) | 52.8 A | −100.0 | 96.5 |
| T14 LAMARDOR FS 400 (150 g/L tebuconazole + 250 g/L prothioconazole) + GAUCHO 600 FS (600 g/L imidacloprid) | 91.96 A | 83.06 | 99.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jevtić, R.; Župunski, V.; Lalošević, M.; Brbaklić, L.; Orbović, B. Co-Occurrence Patterns of Ustilago nuda and Pyrenophora graminea and Fungicide Contribution to Yield Gain in Barley under Fluctuating Climatic Conditions in Serbia. J. Fungi 2022, 8, 542. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050542
Jevtić R, Župunski V, Lalošević M, Brbaklić L, Orbović B. Co-Occurrence Patterns of Ustilago nuda and Pyrenophora graminea and Fungicide Contribution to Yield Gain in Barley under Fluctuating Climatic Conditions in Serbia. Journal of Fungi. 2022; 8(5):542. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050542
Chicago/Turabian StyleJevtić, Radivoje, Vesna Župunski, Mirjana Lalošević, Ljiljana Brbaklić, and Branka Orbović. 2022. "Co-Occurrence Patterns of Ustilago nuda and Pyrenophora graminea and Fungicide Contribution to Yield Gain in Barley under Fluctuating Climatic Conditions in Serbia" Journal of Fungi 8, no. 5: 542. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050542