LncRNAs in Cardiomyocyte Maturation: New Window for Cardiac Regenerative Medicine

Department of Genetics, Faculty of Biological Sciences, Tarbiat Modares University, Tehran 111-14115, Iran
*
Author to whom correspondence should be addressed.
Non-Coding RNA 2021, 7(1), 20; https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7010020
Submission received: 6 February 2021
/
Revised: 5 March 2021
/
Accepted: 8 March 2021
/
Published: 10 March 2021
Abstract
:Cardiomyocyte (CM) maturation, which is characterized by structural, functional, and metabolic specializations, is the last phase of CM development that prepares the cells for efficient and forceful contraction throughout life. Over the past decades, CM maturation has gained increased attention due to the fact that pluripotent stem cell-derived CMs are structurally, transcriptionally, and functionally immature and embryonic-like, which causes a defect in cell replacement therapy. The current challenge is to discover and understand the molecular mechanisms, which control the CM maturation process. Currently, emerging shreds of evidence emphasize the role of long noncoding RNAs (lncRNAs) in regulating different aspects of CM maturation, including myofibril maturation, electrophysiology, and Ca2+ handling maturation, metabolic maturation and proliferation to hypertrophy transition. Here, we describe the structural and functional characteristics of mature CMs. Furthermore, this review highlights the lncRNAs as crucial regulators of different aspects in CM maturation, which have the potential to be used for mature CM production. With the current advances in oligonucleotide delivery; lncRNAs may serve as putative therapeutic targets to produce highly mature CMs for research and regenerative medicine.
1. Introduction
Directed differentiation of human pluripotent stem cells (hPSCs) to cardiomyocytes (CMs) has achieved remarkable progress over the past decade. These have provided powerful resources to study human development, translational research to discover disease mechanisms, new drug discovery as well as cell replacement therapy [1]. Despite tremendous achievements in producing the mature PSC-derived cardiomyocytes (PSC-CM) through tissue engineering-based methods, the highly mature CMs have yet to be achieved [2,3,4]. Most protocols generate immature CMs, which lack many attributes of adult CMs. Consequently, the generated cells cannot be used for efficient drug screening, modeling of adult-onset disease, and also as a source for the cell replacement therapy [5]. The current challenge is to discover and understand the molecular mechanisms, which control the CM maturation process. Cardiomyocyte maturation is the last phase of heart development characterized by structural, gene expression, metabolic, and functional specializations. Maturation is a complex trait controlled in different aspects by multiple signaling networks and regulators within the cytoplasm and nucleus [6]. In recent years, the advancement of novel technologies like microarray and next-generation sequencing (NGS) has revealed that 98% of noncoding sequences of the genome are actively transcribed and produce noncoding transcripts, including small and long noncoding RNAs (lncRNAs) [7]. The discovery of lncRNAs has expanded a new layer of regulation to the complexity of transcriptome. The emerging pieces of evidence demonstrate the important role of lncRNAs in CM differentiation and maturation [8]. Interestingly, a large number of lncRNAs are cardiac-specific or dynamically expressed during the differentiation and maturation processes. LncRNAs are the most heterogeneous group of regulators that can regulate gene expression through various mechanisms, mainly epigenetic and chromatin regulation mechanisms [9]. In this review, we first discuss the hallmarks of CM maturation following by lncRNAs description. Next, we introduce the most novel and important maturation-related lncRNAs and describe their corresponding molecular mechanism in regulating the CM maturation process.
2. Cardiomyocyte Maturation
The dynamic process of cardiac development is a highly orchestrated event that can be divided into three main stages, including specification, morphogenesis, and maturation [6]. A tight gene regulatory network is required to differentiate and specify cardiac lineages from mesodermal progenitor cells [1]. The morphogenesis step is followed by the proper organization of cardiac cells and other structural components of the heart, which is regulated at the molecular, cellular and tissue-level events [10]. At the last step, CM maturation, which is characterized by structural, functional and metabolic specializations, prepares the organ for efficient and forceful contraction and relaxation cycle throughout life [5]. The CM maturation process is an inclusive developmental program that drives a wide range of phenotypic changes, which lead to convert fetal CM to adult CM. The major biological processes involved in CM maturation are myofibril maturation, electrophysiology and Ca2+ handling maturation, metabolic maturation and proliferation to hypertrophy transition, which will be described in more detail [5,6]. Myofibrils, which are composed of repeating sections of sarcomeres, are the contractile apparatuses of CMs [11]. Myofibril maturation is characterized by the increase of sarcomere length, assembly and expansion; improvement of sarcomere alignment [12,13] and M-line formation [14], which results in a massive expansion of myofibrils (Figure 1A). The myofibrils organization enhancement is related to sarcomeric isoform switching through either transcriptional changes or alternative splicing [15]. In this process, several sarcomere components switch from a fetal to an adult isoform, which mainly includes TnnI1 to TnnI3 [16], Myh7 to Myh6 [17], Ttn-N2BA to Ttn-N2B [18] and Myl7 to Myl2 transition [19]. Cardiomyocyte contraction is activated by Ca2+ binding to the myofilaments. The electrical impulses and fluctuations of cytoplasmic Ca2+ concentration strongly regulate the activity of kinases, phosphatases, exchangers, ion channels, and transporters, which are critical for proper cardiomyocyte contraction [20]. The electrophysiology features of mature cardiomyocytes differ in important ways from immature cardiomyocytes [21]. The increased expression of ventricular ion channels like Scn5a [22], Kcnj12, Kcnj2 [23] and Ca2+ handling molecules, such as Cav1.2, Ryr2, and Serca [24], resulted in more negative resting membrane potential and faster upstroke velocity of mature cardiomyocytes [25,26]. Furthermore, the mature cardiomyocyte evolves transverse-tubules (T-tubules), which are an extension of cell members penetrating into the center of mature cardiomyocytes and containing large concentrations of ion channels, transporters, and pumps. The presence of T-tubules allows the rapid transmission of the action potential, synchronizing calcium release and regulating cellular calcium concentration (Figure 1B) [27,28,29]. The dramatic alterations in cardiomyocyte metabolism also occur during the maturation process. The well-defined metabolic shift occurred in fetal to neonatal transition in which the primary source of ATP generation changes from glycolysis to fatty acid β-oxidation [30,31]. The mature cardiomyocytes undergo various adaptations to compensate for this demand. This transition is supported by upregulation of metabolic transcriptional regulator likes Ppagrc1a, Para, Esrra and Nrf1/2 as well as upregulation of fatty acid metabolism and oxidative phosphorylation-related genes [32,33]. The mature cardiomyocytes are highly enriched in mitochondria and occupy up to 40% of cell volume. The cristae structures in mitochondria are densely organized, and the mitochondria are localized close to the SR and myofibrillar structures to support efficient ATP transition (Figure 1C) [34,35,36]. At the time of maturation, the cardiomyocyte proliferation rate declines rapidly through the proliferation-to-hypertrophy transition. This transition is supported through the downregulation of mitogenic signaling pathways. In the mammalian heart NRG1 and its co-receptor ERBB2 as a main players of Neuregulin-ErbB signaling pathway are essential for cardiomyocyte proliferation and can promote the regeneration of cardiac muscle cells [37]. Activation of Notch signaling pathway trigger the BMP10 activity, which in turn promote proliferation of cardiomyocytes through inhibition of the cell cycle inhibitor p57 [38]. It also regulates the transcription of CCND1 and induces cell cycle reentry and progression in neonatal cardiomyocytes [39]. Hippo signaling pathway is also an important regulator of cardiomyocyte proliferation and heart size [37]. The downregulation of cell-cycle-associated genes like Cdk1, Ccnb1 and Aurkb, as well as mentioned proliferation-associated signaling pathways, are the hallmark of this transition [40,41,42,43]. Despite the cell cycle exit, the postnatal heart size also increased more than 30-fold through the maturational hypertrophy [44]. Other aspect of proliferation-to-hypertrophy transition is polyploidization, in which the cardiomyocyte nuclei are polyploid due to DNA endoreplication without karyokinesis (Figure 1D, top) [45,46]. Cardiomyocytes polyploidization also induces the maturational hypertrophy and cause cell cycle exit in mature cardiomyocytes (Figure 1D, bottom) [47]. Finally, efficient integration of cardiomyocytes into cardiac tissues require adhesion like structures and costameres to facilitate extracellular matrix (ECM) attachment as well as intercalated discs (ICDs) structures. ICDs contain fascia adherens, desmosomes, and gap junctions, which support well organized cardiomyocytes junctions and synchronized contraction of cardiac cells [48,49,50,51].
3. Noncoding RNAs
The most recent studies indicate that the developmental complexity of higher eukaryotes is raised from the gene regulation process by a large number of regulators rather than the number of protein-coding genes [52]. The protein-coding genes represent only a small portion of the mammalian genome, while the other majority part produces different classes of noncoding RNAs (ncRNAs). Noncoding RNAs as an important hidden layer of regulation have pivotal roles in almost all biological processes. The ncRNAs can be classified based on their length into small (<200 nucleotides) and long noncoding RNAs (>200 nucleotides) [53,54,55].
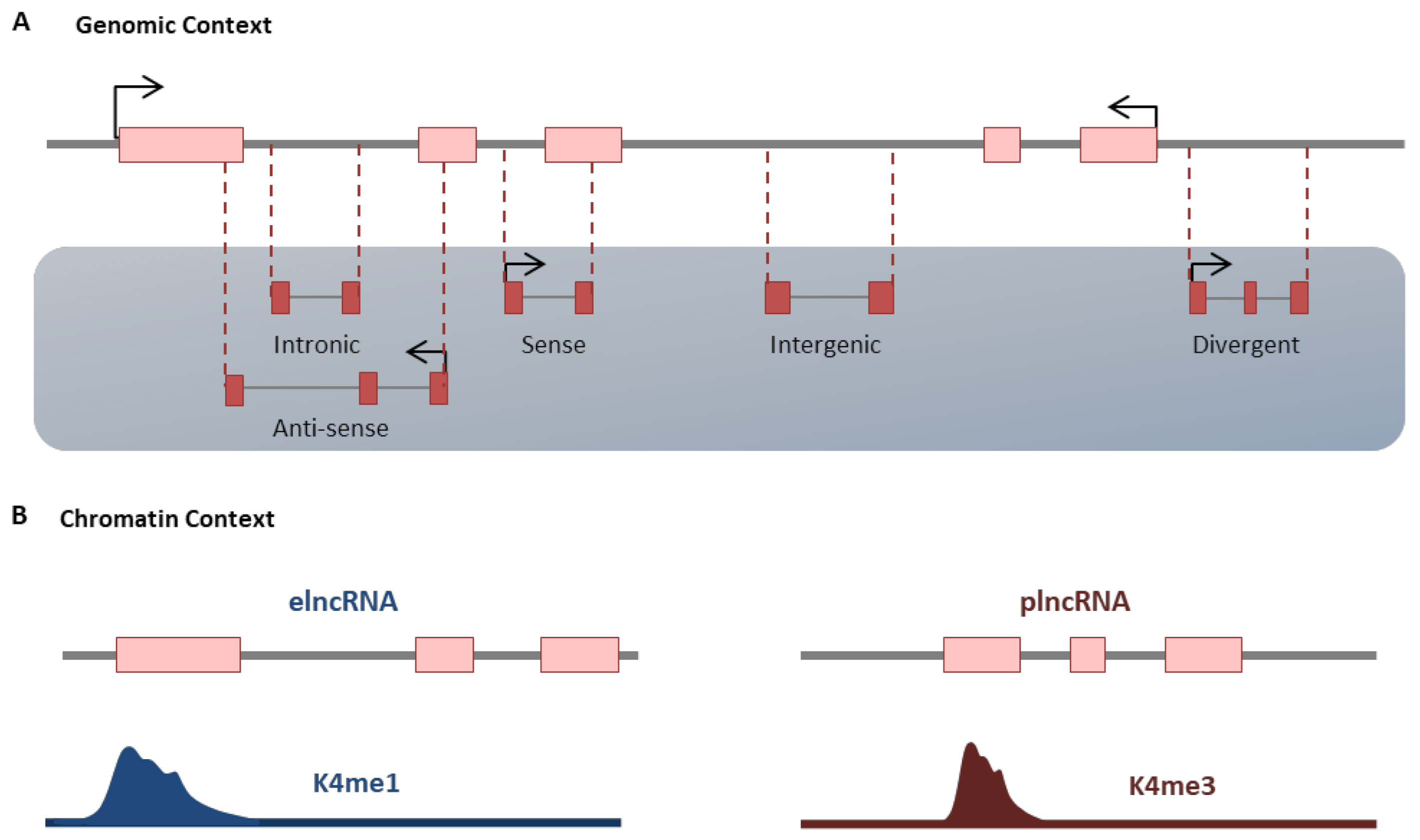
LncRNAs are the most diverse and heterogeneous class of noncoding RNAs [56]. Several transcriptional and post-transcriptional processes, including 5ʹ-capping, splicing and polyadenylation, are done to reach a mature form of lncRNAs [57,58]. Generally, lncRNAs have no open reading frame (ORF), while few studies shed light on the existence of lncRNAs with coding peptides [59]. The lncRNAs have a high degree of tissue specificity [60] and are conserved in either expression pattern or transcript structure despite their low sequence conservation [61,62,63]. The localization of lncRNAs is mainly related to their function. Some of them could be located in both the nucleus and the cytoplasm, while some reside only in the cytoplasm or permanently in the nucleus [64,65]. They could be classified according to their genomic position and orientation relative to their adjacent protein-coding gene [66]. The intergenic lncRNAs (lincRNAs), which do not have any overlap with ding genes, are the most defined group across the organisms [67]. Other groups reside inside the introns of protein-coding genes (intronic lncRNAs), or they are transcribed in the opposite direction of adjacent protein-coding genes (divergent lncRNAs). The lncRNAs can be transcribed from either sense or antisense strand of DNA (Figure 2A) [68,69,70]. They may also be classified based on the chromatin context, in which enhancer-associated (elncRNA) or promoter-associated (plncRNA) lncRNAs are characterized by K4me1 and K4me3 chromatin marks around their transcription start site (Figure 2B) [63]. The critical functions of lncRNAs belong to their ability to interact with regulatory proteins, DNA elements or even functional RNAs to regulate their interaction. LncRNAs can regulate transcription by recruiting transcription factors and chromatin modifier complexes through different mechanisms, including guide, signal, scaffold, or preventing their interaction through decoy mechanism [71,72,73]. LncRNAs also can facilitate the formation of the enhancer-promoter loop at the site of the target locus through regulating inter and intrachromosomal interaction [74,75]. Furthermore the local formation of R-loops (triple-stranded nucleic acid structures with RNA hybridized to duplex DNA) by some anti-sense lncRNAs recruit transcription cofactors to the promoter of corresponded sense mRNA and regulate its transcription [65,76]. The important function of lncRNAs during heart development, differentiation and disease is well recognized. Some important studies revealed the presence of a large number of lncRNAs, which are indispensable for cardiomyocyte maturation (Table 1). The sharp alteration of transcriptome, including lncRNAs at the time of maturation reflects the rapid adaptation of cardiomyocytes for metabolism, regeneration capacity as well as functional properties [8,33]. Below, we review the main lncRNAs involved in the cardiomyocyte maturation procedure.
3.1. Mhrt-LncRNA
The Myh-associated RNA transcript (Mhrt) is a cardiac-specific lncRNA, which is transcribed within the Myh genomic loci. The Myh genomic locus not only encodes the main cardiac contractile genes, including Myh6 and Myh7 but also miRNAs (miR-208a and miR-208b) and Mhrt lncRNA. Mhrt is transcribed in an antisense direction of Myh6 promoter into the genomic site of Myh7 [77]. The Mhrt expression level is increased gradually from embryonic hearts into mature adult hearts. As a cardiac-specific lncRNA, its expression is limited to the heart with no or minimal expression in other tissues [91]. This lncRNA has a different regulatory function in myofibril formation and hypertrophy at the stage of cardiomyocyte maturation. The lncRNA Mhrt has a tight correlation with the Myh6/Myh7 ratio and regulates their expression during the development through Brg1 inhibition. The BRG1 complex can oppositely regulate the expression of the Myh6 and Myh7 contractile genes through the recruitment of different repressors or activators. It can repress the expression of Myh6 by recruitment of chromatin repressor G9A and DNMT3 on Myh6 promoter and activate Myh7 expression through NFAT1 and AP-1 transcription factor recruitment on the site of Myh7 promoter. Mhrt lncRNA directly binds to the helicase domain of BRG1, which causes BRG1 to be sequestered from its target genomic DNA. This negative regulation of BRG1 by Mhrt is essential for the normal ratio of Myh6/Myh7 during the development and maturation (Figure 3) [77,116]. On the other side, Mhrt regulates cardiomyocyte hypertrophy through myocardin expression and function regulation. In the heart, myocardin, as a strong trans-activator, binds directly to SRF and activates cardiomyocytes restricted genes containing CarG boxes in their promoter region [117]. The Mhrt upregulation in the mature heart avoids myocardin expression out of control and protects the heart in the development process. Mhrt can raise the Hdac5 level and enrich the HDAC5/myocardin complex, which leads to myocardin deacetylation. Deacetylation loses the activity of myocardin to regulate the cardiac hypertrophy biomarkers expression (Figure 3) [91]. Mhrt lncRNA also inhibits the myocardin expression level through ce-miRNA mechanisms. It sponges miR-145-5p in a base-complementary pairing manner, which leads to regulating Klf4 expression as its target gene. Upregulation of Klf4 resulted in myocardin downregulation and development of myocardial hypertrophy (Figure 3) [92]. Briefly, Mhrt lncRNA have important roles at the time of maturation through regulating the expression of hypertrophy related markers as well as regulating Myh6/Myh7 ratio in the mature heart.
3.2. H19-LncRNA
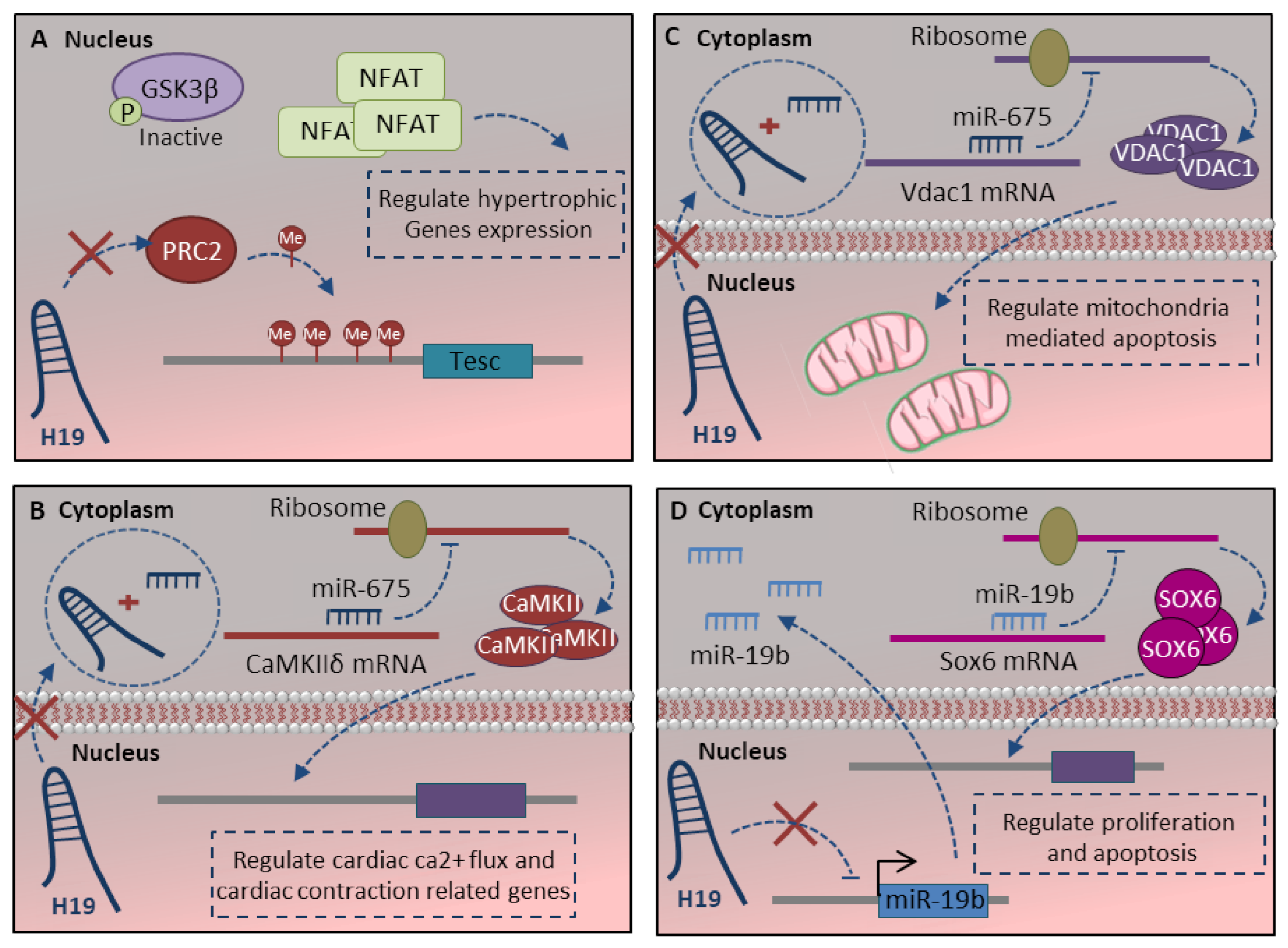
The H19-lncRNA is a highly conserved, imprinted, non-protein-coding gene. The base conservation, along with the highly conserved secondary structure, suggests its pivotal roles in development [96]. H19 has a high-level of expression through cardiomyocyte differentiation, while its expression strongly reduces after birth and through maturation [93]. H19 could regulate the cardiomyocyte maturation process at different steps, including apoptosis, proliferation, hypertrophy and contraction. H19-lncRNA has been demonstrated to regulate hypertrophy in cardiomyocyte cells [80]. H19 physically interacts with polycomb repressive complex 2 (PRC2) to suppress pro-hypertrophic NFAT signaling. Regarding the fact that H19 is downregulated during cardiac development, in the absence of H19, PRC2 hyper-methylates the TESC (tescalin) locus as a main inhibitor of NFAT. This leads to activate NFAT signaling and their corresponding hypertrophy-related target genes (Figure 4A) [93]. H19 also delicately regulates the function of proteins associate with cardiac contraction through the H19-miR-675 axis. It is shown that calcium/calmodulin-dependent protein kinase II Delta (CamkIIδ) is a direct target of H19-derived miR-675 (Figure 4B) [80]. CamkIIδ is the predominant cardiac isoform, which can phosphorylate ion channels and critical membrane proteins for cardiac electrical activity and structure like L-type Ca2+ and voltage-gated Na+ and K+ channels [118]. The absence of the H19-miR-675 axis in the mature heart increases CamkIIδ expression, which in turn regulates the protein associated with cardiac ca2+ flux, cardiac contraction and relaxation.
Other studies also indicate the pivotal role of the H19-miR-675 axis in the mitochondrial apoptotic pathway. The voltage-dependent anion channel 1 (Vdac1) is a direct target of miR-675, which is a key protein in the process of mitochondria-mediated apoptosis (Figure 4C) [94]. Moreover, H19-lncRNA regulates proliferation and apoptosis through the miR-19b-Sox6 axis. H19 inhibits miR19b expression, which leads to Sox6 target gene expression upregulation (Figure 4D). SOX6 is a multifunction transcription factor with critical roles in cell proliferation and apoptosis during the late-stage of cardiomyocyte differentiation [119]. Altogether absence of H19-lncRNA regulates the cardiomyocyte apoptosis, proliferation and protects cardiomyocytes by antiapoptotic effect through either H19-miR-675 or H19-miR-19b axis.
3.3. Ahit LncRNA
The antihypertrophic interrelated transcript (Ahit) lncRNA was recently reported that regulate hypertrophy in the postnatal heart. Ahit is highly enriched in the adult heart and is adjacent to Mef2a, which is a pivotal regulator of cardiac development [120]. While the critical function of MEF2 in cardiac development is clear, a novel function of MEF2 transcription factors in the postnatal heart was proposed. The MEF2 expression in the adult heart is at the basal level, and its upregulation to the level seen in the failing heart can promote chamber dilation, mechanical dysfunction, and dilated cardiomyopathy [121,122]. Ahit lncRNA regulates chromatin structure and expression of Mef2a through direct interaction with SUZ12 (suppressor of Zeste 12) and recruiting PRC2 chromatin remodeler at the promoter region of Mef2a locus. PRC2 modulates histone modification and increases H3K27me3 histone marks, which leads to Mef2a transcription repression [120]. Altogether, Ahit regulates the expression of the Mef2 family and prevents their abnormal function from inducing precise hypertrophy in mature adult cardiomyocytes.
3.4. Zfas1 and Dach1 LncRNAs
Intracellular Ca2+ homeostasis is a key factor to regulate cardiomyocyte contraction as well as cell viability through regulating mitochondrial-mediated apoptosis. This process is tightly regulated by an array of protein and nonprotein-coding genes. SERCA2a (SR Ca2+-ATPase 2a) is the main protein, which is indispensable for the normal intracellular Ca2+ handling process through regulating the Ca2+ reuptake into SR in cardiac muscles [123]. The more recent studies indicate that Serca2 expression and function are delicately regulated by lncRNAs. Zfas1 (zinc finger antisense 1), an antisense lncRNA to the 5′ end of the protein-coding gene Znfx1, is a cardiac-related lncRNA, which regulates cardiac apoptosis as well as contraction activity [4]. Zfas1 negatively regulates the Serca2a through either its expression repression or functional restriction (Figure 5A). This aims to decrease Ca2+ reuptake to the SR, which leads to cytosolic Ca2+ overload [81]. The studies indicate that Ca2+ overloads cause cardiac dysfunction [81] and induce mitochondria-mediated apoptosis [97].
More recently, a highly conserved lncRNA dachshund homolog 1 (Dach1) was also reported by the same group to have a main role in regulating the mature cardiomyocyte proliferation and function. Dach1 expression was gradually upregulated after birth in a mature heart. It has a hallmark role in the blockage of mitosis and proliferation in the mature heart through the Hippo signaling pathway (Figure 5A) [98]. It is well-known that the Hippo signaling pathway regulates the mitosis and proliferation of cardiomyocytes [124]. Dach1 directly interacts with protein phosphatase 1 alpha (PP1A) and reduces its dephosphorylation activity. This leads to improving yes-associated protein 1 (YAP1) phosphorylation, reduces its nuclear localization, which in turn inhibits cardiomyocyte proliferation in the mature heart [98]. Dach1 lncRNA also regulates the function of mature cardiomyocytes through regulating Ca2+ homeostasis in the cells. Dach1 directly interacts with SERCA2a protein, promote its ubiquitination and proteasome degradation (Figure 5A) [82]. All together, Zfas1 and Dach1 manipulate the cardiac function by regulating Ca2+ homeostasis, proliferation and apoptosis in mature cardiomyocytes.
3.5. Carl and Mdrl LncRNAs
Metabolic maturation, which is occurred at the level of prenatal to postnatal transition, is an important event in cardiomyocyte maturation. The increase in myofibril density and contractile components require more ATP production during postnatal hypertrophic growth. Mitochondria are highly dynamic, and their fusion and fission are crucial for maintaining mitochondrial function and fidelity [125]. The mitochondrial biogenesis is tightly regulated by a different network of proteins, miRNAs and lncRNAs. The cardiac apoptosis-related lncRNA (Carl), which is highly expressed at the adult heart, suppresses the cardiomyocytes fission and apoptosis by targeting miR-539 and Prohibitin 2 (PHB2) [88]. PHB2 has main roles in the functional and structural integrity of mitochondria, the cristae morphology and the formation of a super respiratory complex in mitochondria [126]. Carl lncRNA sponge miR-539 in the cardiomyocytes, which in turn upregulates the expression of Phb2 as its target gene (Figure 5B). PHB2 inhibits the fission in the mitochondria and suppresses apoptosis [88].
The mitochondrial fission and apoptosis are also regulated by other cardiomyocytes enriched lncRNA called mitochondrial dynamic related lncRNA (Mdrl). The studies indicate that nucleus enriched miR-361 inhibits pri- to pre-miR-484 processing through direct interaction with a primary transcript of miR-484. Mdrl lncRNA has direct interaction with miR-361 and downregulates its expression levels, which in turn promotes the processing of pri-miR-484 [89]. Upregulation of miR-484 represses Fis1 expression that is involved in mitochondrial fission and apoptosis (Figure 5B) [127]. Carl and Mdrl lncRNAs regulate mitochondrial fission and apoptosis through regulating the combination of miRNAs and transcription factors in adult cardiomyocytes.
3.6. Cpr and Sarrah LncRNAs
Most recent studies indicate the important function of lncRNA Cpr (cardiomyocyte proliferation regulator) in mature cardiomyocyte proliferation. The expression level of Cpr lncRNA is increased in postnatal and adult hearts and is enriched in the nucleus of cardiomyocytes cell type. This lncRNA attenuates the cardiomyocyte proliferation in postnatal and adult hearts. Cpr lncRNA directly interacts and recruits DNMT3A protein to enhance CpG methylation of minichromosomal maintenance 3 (Mcm3) promoter (Figure 5C) [78]. MCM3 belongs to the family of proteins involved in the initiation of eukaryotic genome replication and cell cycle progression [128]. The Mcm3 downregulation in mature cardiomyocytes inhibits cardiomyocyte proliferation. In addition, Liu et al. indicate that Cpr remarkably causes a hypertrophic response in mature cardiomyocytes, which has been shown by sarcomere organization, increased cell surface area, and elevated hypertrophy markers like Anf and Myh6 [78].
The association between lncRNAs and cardiomyocyte survival and function is coming to light more by other reports. The lncRNA Sarrah (short for SCOT1-antisense RNA regulated during aging in the Heart) was reported to have antiapoptotic and pro-survival effects in adult cardiomyocytes. Sarrah directly binds to the promoter region of the Nrf2 gene through RNA-DNA triple helix formation and recruits CRIP2 and p300 transcription factors (Figure 5C) [99]. The Nrf2 pathway promotes cell survival and is well-known for its antiapoptotic and cardio-protective effects [129]. The evolutionary conserved lncRNA Sarrah, as a regulator of cardiomyocyte survival, also enhance the contractile capacity of adult cardiomyocytes [99].
3.7. Other Novel LncRNAs
Most recently, more studies support the critical role of lncRNAs in cardiomyocyte maturation, especially through regulating the hypertrophy, proliferation and conduction system. Terminal differentiation-induced ncRNA (Tincr) regulates hypertrophy through direct interaction with EZH2 and regulates the CaMKII gene expression [83]. LncRNA uc.323 also regulates the expression of the cardiac hypertrophy-related gene, carnitine palmitoyltransferase 1b (Cpt1b), through direct interaction with EZH2 [109]. The regulation of hypertrophy was also supported by Magi1-IT1 lncRNA through inactivating Wnt/beta-catenin pathway via targeting the miR-302e/DKK1 axis [110]. LncRNA Snhg1 is a multifunction lncRNA in cardiomyocyte maturation. It can regulate both cardiomyocyte apoptosis by Snhg1/miR-195/Bcl2l2 [107] or Snhg1/miR-188-5p/PTEN [108] axis and cardiomyocytes hypertrophy through Snhg1/miR-15a-5p/Hmg1 axis [106]. The cardiac regeneration-associated regulator (Ecrar) and NR_045363 lncRNAs regulate cardiomyocyte proliferation and cell cycle progression through E2f1-Ecrar-ERK1/2 [111] and miR-216a/JAK2-STAT3 [114] signaling pathways, respectively. Cardiac conduction regulatory RNA (Ccrr) [84] and Kcna2 antisense RNA (Kcna2 AS) [85] lncRNAs have important roles in regulating the conduction system through regulating the connexin43 (Cx43) and Kcna2 genes expression. These recent studies support the pivotal role of lncRNAs in the maturation process. However, the exact mechanism of their action needs to be elaborated.
4. Therapeutic Target of LncRNAs in Cardiac Remodeling
The biological relevance of lncRNAs to different stages of cardiomyocyte maturation has triggered tremendous interest in their application as a therapeutic option. Nevertheless, there are several important issues that need to be considered before lncRNAs can be used as potential therapeutic targets in regeneration medicine. First, many lncRNAs are not conserved at the sequence level, and this makes the identification of human lncRNAs and their clinical testing challenging [130]. Second, as we described in this review, lncRNAs regulate gene expression through different mechanisms, including chromatin remodeling, miRNA regulation or binding to different proteins, RNA and even DNA elements. This phenomenon brings about potential off-target effects for lncRNAs expression, which should be considered. Consequently, the most promising lncRNAs as therapeutic targets are the ones, which their mechanism of action is well described. Finally, the delivery method of oligonucleotides and their duration of action in vivo should be considered before manipulating the candidate lncRNAs [131]. On the other hand, lncRNAs have high tissue specificity, which makes conserved lncRNAs a promising target for drug development, and this also helps to have less remote off-target effects [132]. In spite of the obstacles mentioned above, the role of CHROME [133] and H19 [134] lncRNAs in cardiovascular disease have been investigated in animal models. Of interest, H19 lncRNA with a pivotal role in the cardiomyocyte maturation process was studied in both mouse and mini-pig models. Li et al. reported that H19 lncRNA is a novel regulator of smooth muscle cell survival in abdominal aortic aneurysm development and progression [134]. All together, further works should be done to focus and clarify the lncRNAs regulatory network, investigate their role in known beneficial/harmful signaling pathways and optimize the delivery technology in animal models to promote the rapid clinical transformation of lncRNAs. In summary, based on the available in vitro and in vivo animal model, current studies with human cells and tissue and optimized delivery system, lncRNAs has potential to be considered as a novel therapeutic option in different filed of cardiovascular development specially maturation process.
5. Conclusions
Over the past decades, several differentiation protocols have been optimized to produce functional cardiomyocytes, which still are structurally, transcriptionally and functionally immature and somehow embryonic-like. In spite of the fact that differences between the immature and mature cardiomyocytes have been explained with details, underlying molecular mechanisms and corresponded regulators that mediate this transition still remain less elaborated. Critical regulators with combinational effects are needed to regulate and optimize cardiomyocyte maturation as a complex process. As previously discussed, several lncRNAs are expressed during the heart development and of interest cardiomyocyte maturation. LncRNAs are defined to regulate the maturation in different aspects, including myofibril maturation, electrophysiology and Ca2+ handling maturation, metabolic maturation and proliferation to hypertrophy transition. LncRNAs like Mhrt, H19, Zfas1 and Carl could act as important participants in promoting the maturation. As a result, they enhance a wide range of phenotypic changes, which promote the fetal to adult cardiomyocytes transition. This review highlights lncRNAs as the multi-effect regulators, which have the potential to promote mature cardiomyocyte production. Targeting the lncRNAs may be a novel approach to producing effective adult-like cardiomyocytes. However, we must consider several challenges, including the lack of sequence conservation, delivery strategy and off-target effect. With the current advances in oligonucleotide delivery and gene therapy, the studies in the next few years may demonstrate the possible role of lncRNAs as a therapeutic target to produce highly mature cardiomyocytes in cell replacement or other clinical therapies.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Liu, Y.-W.; Chen, B.; Yang, X.; Fugate, J.A.; Kalucki, F.A.; Futakuchi-Tsuchida, A.; Couture, L.; Vogel, K.W.; Astley, C.A.; Baldessari, A.; et al. Human embryonic stem cell–derived cardiomyocytes restore function in infarcted hearts of non-human primates. Nat. Biotechnol. 2018, 36, 597–605. [Google Scholar] [CrossRef]
- Huang, C.Y.; Maia-Joca, R.P.M.; Ong, C.S.; Wilson, I.; DiSilvestre, D.; Tomaselli, G.F.; Reich, D.H. Enhancement of human iPSC-derived cardiomyocyte maturation by chemical conditioning in a 3D environment. J. Mol. Cell. Cardiol. 2020, 138, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ronaldson-Bouchard, K.; Ma, S.P.; Yeager, K.; Chen, T.; Song, L.; Sirabella, D.; Morikawa, K.; Teles, D.; Yazawa, M.; Vunjak-Novakovic, G. Advanced maturation of human cardiac tissue grown from pluripotent stem cells. Nature 2018, 556, 239–243. [Google Scholar] [CrossRef]
- Bektik, E.; Cowan, D.B.; Wang, D.-Z. Long non-coding RNAs in atrial fibrillation: Pluripotent stem cell-derived cardiomyocytes as a model system. Int. J. Mol. Sci. 2020, 21, 5424. [Google Scholar] [CrossRef]
- Karbassi, E.; Fenix, A.; Marchiano, S.; Muraoka, N.; Nakamura, K.; Yang, X.; Murry, C.E. Cardiomyocyte maturation: Advances in knowledge and implications for regenerative medicine. Nat. Rev. Cardiol. 2020, 17, 341–359. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Pu, W.T. Cardiomyocyte maturation: New phase in development. Circ. Res. 2020, 126, 1086–1106. [Google Scholar] [CrossRef] [PubMed]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermüller, J.; Hofacker, I.L.; et al. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Touma, M.; Kang, X.; Zhao, Y.; Cass, A.A.; Gao, F.; Biniwale, R.; Coppola, G.; Xiao, X.; Reemtsen, B.; Wang, Y. Decoding the long noncoding RNA during cardiac maturation: A roadmap for functional discovery. Circ. Cardiovasc. Genet. 2016, 9, 395–407. [Google Scholar] [CrossRef]
- Devaux, Y.; Zangrando, J.; Schroen, B.; Creemers, E.E.; Pedrazzini, T.; Chang, C.-P.; Dorn, G.W., II; Thum, T.; Heymans, S. Long noncoding RNAs in cardiac development and ageing. Nat. Rev. Cardiol. 2015, 12, 415. [Google Scholar]
- Sizarov, A.; Ya, J.; de Boer, B.A.; Lamers, W.H.; Christoffels, V.M.; Moorman, A.F. Formation of the building plan of the human heart: Morphogenesis, growth, and differentiation. Circulation 2011, 123, 1125–1135. [Google Scholar] [CrossRef] [Green Version]
- Bird, S.; Doevendans, P.; Van Rooijen, M.; De La Riviere, A.B.; Hassink, R.; Passier, R.; Mummery, C. The human adult cardiomyocyte phenotype. Cardiovasc. Res. 2003, 58, 423–434. [Google Scholar] [CrossRef]
- Lundy, S.D.; Zhu, W.-Z.; Regnier, M.; Laflamme, M.A. Structural and functional maturation of cardiomyocytes derived from human pluripotent stem cells. Stem Cells Dev. 2013, 22, 1991–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iorga, B.; Schwanke, K.; Weber, N.; Wendland, M.; Greten, S.; Piep, B.; Dos Remedios, C.G.; Martin, U.; Zweigerdt, R.; Kraft, T.; et al. Differences in contractile function of myofibrils within human embryonic stem cell-derived cardiomyocytes vs. adult ventricular myofibrils are related to distinct sarcomeric protein isoforms. Front. Physiol. 2018, 8, 1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarkova, I.; Perriard, J.-C. The M-band: An elastic web that crosslinks thick filaments in the center of the sarcomere. Trends Cell Biol. 2005, 15, 477–485. [Google Scholar] [CrossRef]
- Reiser, P.J.; Portman, M.A.; Ning, X.-H.; Moravec, C.S. Human cardiac myosin heavy chain isoforms in fetal and failing adult atria and ventricles. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H1814–H1820. [Google Scholar] [CrossRef] [Green Version]
- Siedner, S.; Krüger, M.; Schroeter, M.; Metzler, D.; Roell, W.; Fleischmann, B.K.; Hescheler, J.; Pfitzer, G.; Stehle, R. Developmental changes in contractility and sarcomeric proteins from the early embryonic to the adult stage in the mouse heart. J. Physiol. 2003, 548, 493–505. [Google Scholar] [CrossRef]
- Mahdavi, V.; Lompre, A.; Chambers, A.; Nadal-Ginard, B. Cardiac myosin heavy chain isozymic transitions during development and under pathological conditions are regulated at the level of mRNA availability. Eur. Heart J. 1984, 5, 181–191. [Google Scholar] [CrossRef]
- Lahmers, S.; Wu, Y.; Call, D.R.; Labeit, S.; Granzier, H. Developmental control of titin isoform expression and passive stiffness in fetal and neonatal myocardium. Circ. Res. 2004, 94, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Kubalak, S.W.; Miller-Hance, W.C.; O’Brien, T.X.; Dyson, E.; Chien, K.R. Chamber specification of atrial myosin light chain-2 expression precedes septation during murine cardiogenesis. J. Biol. Chem. 1994, 269, 16961–16970. [Google Scholar] [CrossRef]
- Louch, W.E.; Koivumäki, J.T.; Tavi, P. Calcium signalling in developing cardiomyocytes: Implications for model systems and disease. J. Physiol. 2015, 593, 1047–1063. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Gao, S.; Nie, L.; Tang, M.; Huang, W.; Luo, H.; Hu, X.; Xi, J.; Zhu, M.; Zheng, Y.; et al. Molecular and functional changes in voltage-gated Na+ channels in cardiomyocytes during mouse embryogenesis. Circ. J. 2011, 75, 2071–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veerman, C.C.; Mengarelli, I.; Lodder, E.M.; Kosmidis, G.; Bellin, M.; Zhang, M.; Dittmann, S.; Guan, K.; Wilde, A.A.; Schulze-Bahr, E.; et al. Switch From Fetal to Adult SCN 5A Isoform in Human Induced Pluripotent Stem Cell–Derived Cardiomyocytes Unmasks the Cellular Phenotype of a Conduction Disease–Causing Mutation. J. Am. Heart Assoc. 2017, 6, e005135. [Google Scholar] [CrossRef]
- Goversen, B.; van der Heyden, M.A.; van Veen, T.A.; de Boer, T.P. The immature electrophysiological phenotype of iPSC-CMs still hampers in vitro drug screening: Special focus on IK1. Pharmacol. Ther. 2018, 183, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Boutjdir, M. Gene expression of SERCA2a and L-and T-type Ca channels during human heart development. Pediatr. Res. 2001, 50, 569–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bers, D.M. Cardiac excitation–contraction coupling. Nature 2002, 415, 198–205. [Google Scholar] [CrossRef]
- Peinkofer, G.; Burkert, K.; Urban, K.; Krausgrill, B.; Hescheler, J.; Saric, T.; Halbach, M. From early embryonic to adult stage: Comparative study of action potentials of native and pluripotent stem cell-derived cardiomyocytes. Stem Cells Dev. 2016, 25, 1397–1406. [Google Scholar] [CrossRef]
- Lieu, D.K.; Liu, J.; Siu, C.-W.; McNerney, G.P.; Tse, H.-F.; Abu-Khalil, A.; Huser, T.; Li, R.A. Absence of transverse tubules contributes to non-uniform ca2+ wavefronts in mouse and human embryonic stem cell–derived cardiomyocytes. Stem Cells Dev. 2009, 18, 1493–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, T.; Shaw, R.M. Cardiac T-tubule microanatomy and function. Physiol. Rev. 2017, 97, 227–252. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Pu, W.T. The architecture and function of cardiac dyads. Biophys. Rev. 2020, 12, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Lopaschuk, G.D.; Jaswal, J.S. Energy metabolic phenotype of the cardiomyocyte during development, differentiation, and postnatal maturation. J. Cardiovasc. Pharmacol. 2010, 56, 130–140. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Y.; Dorn, G.W. Mitochondrial fusion is essential for organelle function and cardiac homeostasis. Circ. Res. 2011, 109, 1327–1331. [Google Scholar] [CrossRef]
- Sim, C.B.; Ziemann, M.; Kaspi, A.; Harikrishnan, K.; Ooi, J.; Khurana, I.; Chang, L.; Hudson, J.E.; El-Osta, A.; Porrello, E.R. Dynamic changes in the cardiac methylome during postnatal development. FASEB J. 2015, 29, 1329–1343. [Google Scholar] [CrossRef] [Green Version]
- Uosaki, H.; Cahan, P.; Lee, D.I.; Wang, S.; Miyamoto, M.; Fernandez, L.; Kass, D.A.; Kwon, C. Transcriptional landscape of cardiomyocyte maturation. Cell Rep. 2015, 13, 1705–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaper, J.; Meiser, E.; Stämmler, G. Ultrastructural morphometric analysis of myocardium from dogs, rats, hamsters, mice, and from human hearts. Circ. Res. 1985, 56, 377–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seppet, E.K.; Kaambre, T.; Sikk, P.; Tiivel, T.; Vija, H.; Tonkonogi, M.; Sahlin, K.; Kay, L.; Appaix, F.; Braun, U.; et al. Functional complexes of mitochondria with Ca, MgATPases of myofibrils and sarcoplasmic reticulum in muscle cells. Biochim. Biophys. Acta BBA Bioenergy 2001, 1504, 379–395. [Google Scholar] [CrossRef] [Green Version]
- Dai, D.-F.; Danoviz, M.E.; Wiczer, B.; Laflamme, M.A.; Tian, R. Mitochondrial maturation in human pluripotent stem cell derived cardiomyocytes. Stem Cells Int. 2017, 2017, 5153625. [Google Scholar] [CrossRef]
- Yutzey, K.E. Neuregulin 1 makes heart muscle. Nature 2015, 520, 445–446. [Google Scholar] [CrossRef] [PubMed]
- Grego-Bessa, J.; Luna-Zurita, L.; del Monte, G.; Bolós, V.; Melgar, P.; Arandilla, A.; Garratt, A.N.; Zang, H.; Mukouyama, Y.-S.; Chen, H.; et al. Notch signaling is essential for ventricular chamber development. Dev. Cell 2007, 12, 415–429. [Google Scholar] [CrossRef] [Green Version]
- Campa, V.M.; Gutiérrez-Lanza, R.; Cerignoli, F.; Díaz-Trelles, R.; Nelson, B.; Tsuji, T.; Barcova, M.; Jiang, W.; Mercola, M. Notch activates cell cycle reentry and progression in quiescent cardiomyocytes. J. Cell Biol. 2008, 183, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Heallen, T.; Morikawa, Y.; Leach, J.; Tao, G.; Willerson, J.T.; Johnson, R.L.; Martin, J.F. Hippo signaling impedes adult heart regeneration. Development 2013, 140, 4683–4690. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Guo, H.; Cao, Y.; Zohrabian, S.; Zhou, P.; Ma, Q.; VanDusen, N.; Guo, Y.; Zhang, J.; Stevens, S.M.; et al. Acetylation of VGLL4 regulates Hippo-YAP signaling and postnatal cardiac growth. Dev. Cell 2016, 39, 466–479. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, T.M.; Ang, Y.-S.; Radzinsky, E.; Zhou, P.; Huang, Y.; Elfenbein, A.; Foley, A.; Magnitsky, S.; Srivastava, D. Regulation of cell cycle to stimulate adult cardiomyocyte proliferation and cardiac regeneration. Cell 2018, 173, 104–116.e112. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F.; Meng, Q.; Yu, Y.; Shao, L.; Shen, Z. Adult Cardiomyocyte Proliferation: A New Insight for Myocardial Infarction Therapy. J. Cardiovasc. Transl. Res. 2020, 1–10. [Google Scholar] [CrossRef]
- Yatscoff, M.A.; Jaswal, J.S.; Grant, M.R.; Greenwood, R.; Lukat, T.; Beker, D.L.; Rebeyka, I.M.; Lopaschuk, G.D. Myocardial hypertrophy and the maturation of fatty acid oxidation in the newborn human heart. Pediatr. Res. 2008, 64, 643–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, M.; Barske, L.; Van Handel, B.; Rau, C.D.; Gan, P.; Sharma, A.; Parikh, S.; Denholtz, M.; Huang, Y.; Yamaguchi, Y.; et al. Frequency of mononuclear diploid cardiomyocytes underlies natural variation in heart regeneration. Nat. Genet. 2017, 49, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- González-Rosa, J.M.; Sharpe, M.; Field, D.; Soonpaa, M.H.; Field, L.J.; Burns, C.E.; Burns, C.G. Myocardial polyploidization creates a barrier to heart regeneration in zebrafish. Dev. Cell 2018, 44, 433–446.e437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhang, C.-H.; Ammanamanchi, N.; Suresh, S.; Lewarchik, C.; Rao, K.; Uys, G.M.; Han, L.; Abrial, M.; Yimlamai, D.; et al. Control of cytokinesis by β-adrenergic receptors indicates an approach for regulating cardiomyocyte endowment. Sci. Transl. Med. 2019, 11, eaaw6419. [Google Scholar] [CrossRef]
- Peter, A.K.; Cheng, H.; Ross, R.S.; Knowlton, K.U.; Chen, J. The costamere bridges sarcomeres to the sarcolemma in striated muscle. Prog. Pediatric Cardiol. 2011, 31, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Paige, S.L.; Plonowska, K.; Xu, A.; Wu, S.M. Molecular regulation of cardiomyocyte differentiation. Circ. Res. 2015, 116, 341–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermij, S.H.; Abriel, H.; van Veen, T.A. Refining the molecular organization of the cardiac intercalated disc. Cardiovasc. Res. 2017, 113, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Epifantseva, I.; Shaw, R.M. Intracellular trafficking pathways of Cx43 gap junction channels. Biochim. Biophys. Acta BBA Biomembr. 2018, 1860, 40–47. [Google Scholar] [CrossRef]
- Morris, K.V.; Mattick, J.S. The rise of regulatory RNA. Nat. Rev. Genet. 2014, 15, 423–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotini, A.; Martínez-Sarrà, E.; Pozzo, E.; Sampaolesi, M. Interactions between microRNAs and long non-coding RNAs in cardiac development and repair. Pharmacol. Res. 2018, 127, 58–66. [Google Scholar] [CrossRef]
- Kay, M.; Soltani, B.M.; Aghdaei, F.H.; Ansari, H.; Baharvand, H. Hsa-miR-335 regulates cardiac mesoderm and progenitor cell differentiation. Stem Cell Res. Ther. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Ekhteraei-Tousi, S.; Mohammad-Soltani, B.; Sadeghizadeh, M.; Mowla, S.J.; Parsi, S.; Soleimani, M. Inhibitory effect of hsa-miR-590-5p on cardiosphere-derived stem cells differentiation through downregulation of TGFB signaling. J. Cell. Biochem. 2015, 116, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Jafarzadeh, M.; Soltani, B.M. Long Noncoding RNA LOC400043 (LINC02381) Inhibits Gastric Cancer Progression Through Regulating Wnt Signaling Pathway. Front. Oncol. 2020, 10, 2189. [Google Scholar] [CrossRef]
- Melé, M.; Mattioli, K.; Mallard, W.; Shechner, D.M.; Gerhardinger, C.; Rinn, J.L. Chromatin environment, transcriptional regulation, and splicing distinguish lincRNAs and mRNAs. Genome Res. 2017, 27, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlackow, M.; Nojima, T.; Gomes, T.; Dhir, A.; Carmo-Fonseca, M.; Proudfoot, N.J. Distinctive patterns of transcription and RNA processing for human lincRNAs. Mol. Cell 2017, 65, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Osak, M.; Bogu, G.K.; Stanton, L.W.; Johnson, R.; Lipovich, L. Genome-wide computational identification and manual annotation of human long noncoding RNA genes. RNA 2010, 16, 1478–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Washietl, S.; Kellis, M.; Garber, M. Evolutionary dynamics and tissue specificity of human long noncoding RNAs in six mammals. Genome Res. 2014, 24, 616–628. [Google Scholar] [CrossRef] [Green Version]
- Marques, A.C.; Ponting, C.P. Catalogues of mammalian long noncoding RNAs: Modest conservation and incompleteness. Genome Biol. 2009, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulitsky, I.; Shkumatava, A.; Jan, C.H.; Sive, H.; Bartel, D.P. Conserved function of lincRNAs in vertebrate embryonic development despite rapid sequence evolution. Cell 2011, 147, 1537–1550. [Google Scholar] [CrossRef] [Green Version]
- Kapusta, A.; Feschotte, C. Volatile evolution of long noncoding RNA repertoires: Mechanisms and biological implications. Trends Genet. 2014, 30, 439–452. [Google Scholar] [CrossRef] [Green Version]
- Mercer, T.R.; Dinger, M.E.; Sunkin, S.M.; Mehler, M.F.; Mattick, J.S. Specific expression of long noncoding RNAs in the mouse brain. Proc. Natl. Acad. Sci. USA 2008, 105, 716–721. [Google Scholar] [CrossRef] [Green Version]
- Rinn, J.L.; Chang, H.Y. Long noncoding RNAs: Molecular modalities to organismal functions. Annu. Rev. Biochem. 2020, 89, 283–308. [Google Scholar] [CrossRef]
- Laurent, G.S.; Wahlestedt, C.; Kapranov, P. The Landscape of long noncoding RNA classification. Trends Genet. 2015, 31, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Cabili, M.N.; Trapnell, C.; Goff, L.; Koziol, M.; Tazon-Vega, B.; Regev, A.; Rinn, J.L. Integrative annotation of human large intergenic noncoding RNAs reveals global properties and specific subclasses. Genes Dev. 2011, 25, 1915–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, E.J.; Chin-Inmanu, K.; Jia, H.; Lipovich, L. Sense-antisense gene pairs: Sequence, transcription, and structure are not conserved between human and mouse. Front. Genet. 2013, 4, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Lu, J.Y.; Liu, L.; Yin, Y.; Chen, C.; Han, X.; Wu, B.; Xu, R.; Liu, W.; Yan, P.; et al. Divergent lncRNAs regulate gene expression and lineage differentiation in pluripotent cells. Cell Stem Cell 2016, 18, 637–652. [Google Scholar] [CrossRef]
- Zampetaki, A.; Albrecht, A.; Steinhofel, K. Long non-coding RNA structure and function: Is there a link? Front. Physiol. 2018, 9, 1201. [Google Scholar] [CrossRef] [PubMed]
- Vance, K.W.; Ponting, C.P. Transcriptional regulatory functions of nuclear long noncoding RNAs. Trends Genet. 2014, 30, 348–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, R.B.-T.; Ulitsky, I. The functions of long noncoding RNAs in development and stem cells. Development 2016, 143, 3882–3894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjeije, H.; Soltani, B.M.; Behmanesh, M.; Zali, M.R. YWHAE long non-coding RNA competes with miR-323a-3p and miR-532-5p through activating K-Ras/Erk1/2 and PI3K/Akt signaling pathways in HCT116 cells. Hum. Mol. Genet. 2019, 28, 3219–3231. [Google Scholar] [CrossRef]
- Ounzain, S.; Pedrazzini, T. Super-enhancer lncs to cardiovascular development and disease. Biochim. Biophys. Acta BBA Mol. Cell Res. 2016, 1863, 1953–1960. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.-W.; Wang, Y.; Chen, L.-L. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef]
- Han, P.; Li, W.; Lin, C.-H.; Yang, J.; Shang, C.; Nurnberg, S.T.; Jin, K.K.; Xu, W.; Lin, C.-Y.; Lin, C.-J.; et al. A long noncoding RNA protects the heart from pathological hypertrophy. Nature 2014, 514, 102–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponnusamy, M.; Liu, F.; Zhang, Y.-H.; Li, R.-B.; Zhai, M.; Liu, F.; Zhou, L.-Y.; Liu, C.-Y.; Yan, K.-W.; Dong, Y.-H.; et al. Long noncoding RNA CPR (cardiomyocyte proliferation regulator) regulates cardiomyocyte proliferation and cardiac repair. Circulation 2019, 139, 2668–2684. [Google Scholar] [CrossRef]
- Kang, X.; Zhao, Y.; Van Arsdell, G.; Nelson, S.F.; Touma, M. Ppp1r1b-lncRNA inhibits PRC2 at myogenic regulatory genes to promote cardiac and skeletal muscle development in mouse and human. RNA 2020, 26, 481–491. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; An, X.; Li, Z.; Song, Y.; Li, L.; Zuo, S.; Liu, N.; Yang, G.; Wang, H.; Cheng, X. The H19 long noncoding RNA is a novel negative regulator of cardiomyocyte hypertrophy. Cardiovasc. Res. 2016, 111, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Jiao, L.; Sun, L.; Li, Y.; Gao, Y.; Xu, C.; Shao, Y.; Li, M.; Li, C.; Lu, Y.; et al. LncRNA ZFAS1 as a SERCA2a inhibitor to cause intracellular Ca2+ overload and contractile dysfunction in a mouse model of myocardial infarction. Circ. Res. 2018, 122, 1354–1368. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Zhang, Y.; Zhao, Y.; Wang, J.; Li, T.; Zhang, Y.; Jiang, Y.; Jin, X.; Xue, G.; Li, P.; et al. Long Noncoding RNA–DACH1 (Dachshund Homolog 1) Regulates Cardiac Function by Inhibiting SERCA2a (Sarcoplasmic Reticulum Calcium ATPase 2a). Hypertension 2019, 74, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Chen, G.; Lv, F.; Liu, Y.; Tian, H.; Tao, R.; Jiang, R.; Zhang, W.; Zhuo, C. LncRNA TINCR attenuates cardiac hypertrophy by epigenetically silencing CaMKII. Oncotarget 2017, 8, 47565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Sun, L.; Xuan, L.; Pan, Z.; Hu, X.; Liu, H.; Bai, Y.; Jiao, L.; Li, Z.; Cui, L.; et al. Long non-coding RNA CCRR controls cardiac conduction via regulating intercellular coupling. Nat. Commun. 2018, 9, 4176. [Google Scholar] [CrossRef] [Green Version]
- Long, Q.Q.; Wang, H.; Gao, W.; Fan, Y.; Li, Y.F.; Ma, Y.; Yang, Y.; Shi, H.J.; Chen, B.R.; Meng, H.Y.; et al. Long noncoding RNA Kcna2 antisense RNA contributes to ventricular arrhythmias via silencing Kcna2 in rats with congestive heart failure. J. Am. Heart Assoc. 2017, 6, e005965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, P.; Yang, M.; Ren, H.; Shen, G.; Chen, J.; Zhang, J.; Liu, J.; Sun, C. Long noncoding RNA MALAT1 downregulates cardiac transient outward potassium current by regulating miR-200c/HMGB1 pathway. J. Cell. Biochem. 2018, 119, 10239–10249. [Google Scholar] [CrossRef]
- Dai, W.; Chao, X.; Li, S.; Zhou, S.; Zhong, G.; Jiang, Z. Long Noncoding RNA HOTAIR Functions as a Competitive Endogenous RNA to Regulate Connexin43 Remodeling in Atrial Fibrillation by Sponging MicroRNA-613. Cardiovasc. Ther. 2020, 2020, 5925342. [Google Scholar] [CrossRef]
- Wang, K.; Long, B.; Zhou, L.-Y.; Liu, F.; Zhou, Q.-Y.; Liu, C.-Y.; Fan, Y.-Y.; Li, P.-F. CARL lncRNA inhibits anoxia-induced mitochondrial fission and apoptosis in cardiomyocytes by impairing miR-539-dependent PHB2 downregulation. Nat. Commun. 2014, 5, 3596. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Sun, T.; Li, N.; Wang, Y.; Wang, J.-X.; Zhou, L.-Y.; Long, B.; Liu, C.-Y.; Liu, F.; Li, P.-F. MDRL lncRNA regulates the processing of miR-484 primary transcript by targeting miR-361. PloS Genet. 2014, 10, e1004467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zou, X.; Liu, F. Silencing TTTY15 mitigates hypoxia-induced mitochondrial energy metabolism dysfunction and cardiomyocytes apoptosis via TTTY15/let-7i-5p and TLR3/NF-κB pathways. Cell Signal. 2020, 76, 109779. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Xu, Y.; Liang, C.; Xing, W.; Zhang, T. The mechanism of myocardial hypertrophy regulated by the interaction between mhrt and myocardin. Cell Signal. 2018, 43, 11–20. [Google Scholar] [CrossRef]
- Xu, Y.; Luo, Y.; Liang, C.; Zhang, T. LncRNA-Mhrt regulates cardiac hypertrophy by modulating the miR-145a-5p/KLF4/myocardin axis. J. Mol. Cell. Cardiol. 2020, 139, 47–61. [Google Scholar] [CrossRef]
- Viereck, J.; Bührke, A.; Foinquinos, A.; Chatterjee, S.; Kleeberger, J.A.; Xiao, K.; Janssen-Peters, H.; Batkai, S.; Ramanujam, D.; Kraft, T.; et al. Targeting muscle-enriched long non-coding RNA H19 reverses pathological cardiac hypertrophy. Eur. Heart J. 2020, 41, 3462–3474. [Google Scholar]
- Li, X.; Wang, H.; Yao, B.; Xu, W.; Chen, J.; Zhou, X. lncRNA H19/miR-675 axis regulates cardiomyocyte apoptosis by targeting VDAC1 in diabetic cardiomyopathy. Sci. Rep. 2016, 6, 36340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.; Dong, B.; Chen, M.; Yao, C. LncRNA H19 suppresses pyroptosis of cardiomyocytes to attenuate myocardial infarction in a PBX3/CYP1B1-dependent manner. Mol. Cell. Biochem. 2021, 476, 1387–1400. [Google Scholar] [CrossRef] [PubMed]
- Juan, V.; Crain, C.; Wilson, C. Evidence for evolutionarily conserved secondary structure in the H19 tumor suppressor RNA. Nucleic Acids Res. 2000, 28, 1221–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, L.; Li, M.; Shao, Y.; Zhang, Y.; Gong, M.; Yang, X.; Wang, Y.; Tan, Z.; Sun, L.; Xuan, L.; et al. lncRNA-ZFAS1 induces mitochondria-mediated apoptosis by causing cytosolic Ca 2+ overload in myocardial infarction mice model. Cell Death Dis. 2019, 10, 942. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Ma, W.; Wang, X.; Sukhareva, N.; Hua, B.; Zhang, L.; Xu, J.; Li, X.; Li, S.; Liu, S.; et al. Targeting LncDACH1 promotes cardiac repair and regeneration after myocardium infarction. Cell Death Differ. 2020, 27, 2158–2175. [Google Scholar] [CrossRef]
- Trembinski, D.J.; Bink, D.I.; Theodorou, K.; Sommer, J.; Fischer, A.; van Bergen, A.; Kuo, C.-C.; Costa, I.G.; Schürmann, C.; Leisegang, M.S.; et al. Aging-regulated anti-apoptotic long non-coding RNA Sarrah augments recovery from acute myocardial infarction. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, X.-J.; Ji, Y.-X.; Zhang, P.; Deng, K.-Q.; Gong, J.; Ren, S.; Wang, X.; Chen, I.; Wang, H.; et al. The long noncoding RNA Chaer defines an epigenetic checkpoint in cardiac hypertrophy. Nat. Med. 2016, 22, 1131. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liang, Y.; Huang, X.; Guo, X.; Liu, Y.; Zhong, J.; Yuan, J. STAT3-induced upregulation of lncRNA MEG3 regulates the growth of cardiac hypertrophy through miR-361-5p/HDAC9 axis. Sci. Rep. 2019, 9, 460. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, J.; Geng, J.; Chen, F.; Wei, Z.; Liu, C.; Zhang, X.; Li, Q.; Zhang, J.; Gao, L.; et al. Long non-coding RNA MEG3 knockdown attenuates endoplasmic reticulum stress-mediated apoptosis by targeting p53 following myocardial infarction. J. Cell. Mol. Med. 2019, 23, 8369–8380. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.; Li, T.; Li, X.; Xu, C.; Liu, Q.; Jiang, H.; Li, Y.; Liu, Y.; Yan, H.; Huang, Q.; et al. The lncRNA Plscr4 controls cardiac hypertrophy by regulating miR-214. Mol. Ther. Nucleic Acids 2018, 10, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Viereck, J.; Kumarswamy, R.; Foinquinos, A.; Xiao, K.; Avramopoulos, P.; Kunz, M.; Dittrich, M.; Maetzig, T.; Zimmer, K.; Remke, J.; et al. Long noncoding RNA Chast promotes cardiac remodeling. Sci. Transl. Med. 2016, 8, 326ra22. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, F.; Zhou, L.-Y.; Long, B.; Yuan, S.-M.; Wang, Y.; Liu, C.-Y.; Sun, T.; Zhang, X.-J.; Li, P.-F. The long noncoding RNA CHRF regulates cardiac hypertrophy by targeting miR-489. Circ. Res. 2014, 114, 1377–1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, S.M.; Li, H.; Shu, Q.; Wu, W.J.; Luo, X.M.; Lu, L. LncRNA SNHG1 exerts a protective role in cardiomyocytes hypertrophy via targeting miR-15a-5p/HMGA1 axis. Cell Biol. Int. 2020, 44, 1009–1019. [Google Scholar] [CrossRef]
- Zhang, N.; Meng, X.; Mei, L.; Hu, J.; Zhao, C.; Chen, W. The long non-coding RNA SNHG1 attenuates cell apoptosis by regulating miR-195 and BCL2-like protein 2 in human cardiomyocytes. Cell. Physiol. Biochem. 2018, 50, 1029–1040. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Zhang, Z.; Lu, X.; Yang, X. Long non-coding RNA SNHG15 regulates cardiomyocyte apoptosis after hypoxia/reperfusion injury via modulating miR-188-5p/PTEN axis. Arch. Physiol. Biochem. 2020, 1–8. [Google Scholar] [CrossRef]
- Sun, Y.; Fan, W.; Xue, R.; Dong, B.; Liang, Z.; Chen, C.; Li, J.; Wang, Y.; Zhao, J.; Huang, H.; et al. Transcribed Ultraconserved Regions, Uc. 323, Ameliorates Cardiac Hypertrophy by Regulating the Transcription of CPT1b (Carnitine Palmitoyl transferase 1b). Hypertension 2020, 75, 79–90. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, F.; Wang, F.; Wu, N. Long noncoding RNA MAGI1-IT1 regulates cardiac hypertrophy by modulating miR-302e/DKK1/Wnt/beta-catenin signaling pathway. J. Cell. Physiol. 2020, 235, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, X.; Li, B.; Wang, H.; Li, M.; Huang, S.; Sun, Y.; Chen, G.; Si, X.; Huang, C. Long non-coding RNA ECRAR triggers post-natal myocardial regeneration by activating ERK1/2 signaling. Mol. Ther. 2019, 27, 29–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Li, H.; Li, X.; Li, B.; Zhong, L.; Huang, S.; Zheng, H.; Li, M.; Jin, G.; Liao, W.; et al. Loss of long non-coding RNA CRRL promotes cardiomyocyte regeneration and improves cardiac repair by functioning as a competing endogenous RNA. J. Mol. Cell. Cardiol. 2018, 122, 152–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.; Li, C.; Feng, J.; Zhang, J.; Fang, Y. lncRNA UCA1 is a novel regulator in cardiomyocyte hypertrophy through targeting the miR-184/HOXA9 axis. Cardiorenal Med. 2018, 8, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, X.; Shen, D.; Ge, D.; Chen, J.; Pei, J.; Li, Y.; Yue, Z.; Feng, J.; Chu, M.; et al. A long noncoding RNA NR_045363 controls cardiomyocyte proliferation and cardiac repair. J. Mol. Cell. Cardiol. 2019, 127, 105–114. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Cai, B.; Li, Y.; Jiang, Y.; Fu, X.; Zhao, Y.; Gao, H.; Yang, Y.; Yang, J.; et al. The long noncoding RNA lncCIRBIL disrupts the nuclear translocation of Bclaf1 alleviating cardiac ischemia–reperfusion injury. Nat. Commun. 2021, 12, 522. [Google Scholar]
- Hang, C.T.; Yang, J.; Han, P.; Cheng, H.-L.; Shang, C.; Ashley, E.; Zhou, B.; Chang, C.-P. Chromatin regulation by Brg1 underlies heart muscle development and disease. Nature 2010, 466, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Lu, M.M.; Cheng, L.; Yuan, L.-J.; Zhu, X.; Stout, A.L.; Chen, M.; Li, J.; Parmacek, M.S. Myocardin is required for cardiomyocyte survival and maintenance of heart function. Proc. Natl. Acad. Sci. USA 2009, 106, 18734–18739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaminathan, P.D.; Purohit, A.; Hund, T.J.; Anderson, M.E. Calmodulin-dependent protein kinase II: Linking heart failure and arrhythmias. Circ. Res. 2012, 110, 1661–1677. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Xu, H.; Cheng, J.; Zhang, Y.; Gao, C.; Fan, T.; Peng, B.; Li, B.; Liu, L.; Cheng, Z. Downregulation of long non-coding RNA H19 promotes P19CL6 cells proliferation and inhibits apoptosis during late-stage cardiac differentiation via miR-19b-modulated Sox6. Cell Biosci. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Yang, Y.; Xu, Z.; Lan, C.; Chen, C.; Li, C.; Chen, Z.; Yu, C.; Xia, X.; Liao, Q.; et al. Long Noncoding RNA Ahit Protects Against Cardiac Hypertrophy Through SUZ12 (Suppressor of Zeste 12 Protein Homolog)-Mediated Downregulation of MEF2A (Myocyte Enhancer Factor 2A). Circ. Heart Fail. 2020, 13, e006525. [Google Scholar] [CrossRef]
- Pereira, A.H.M.; Cardoso, A.C.; Consonni, S.R.; Oliveira, R.R.; Saito, A.; Vaggione, M.L.B.; Matos-Souza, J.R.; Carazzolle, M.F.; Goncalves, A.; Fernandes, J.L. MEF2C repressor variant deregulation leads to cell cycle re-entry and development of heart failure. EBioMedicine 2020, 51, 102571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Oort, R.; van Rooij, E.; Bourajjaj, M.; Schimmel, J.; Jansen, M.; van der Nagel, R.; Doevendans, P.; Schneider, M.; van Echteld, C.; De Windt, L.J. MEF2 activates a genetic program promoting chamber dilation and contractile dysfunction in calcineurin-induced heart failure. Circulation 2006, 114, 298–308. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ni, J.; Lan, L.; Michael, S.; Rui, G.; Xiyun, B.; Xiaozhi, L.; Guanwei, F. SERCA2a: A key protein in the Ca2+ cycle of the heart failure. Heart Fail. Rev. 2019, 25, 523–535. [Google Scholar]
- Tian, Y.; Liu, Y.; Wang, T.; Zhou, N.; Kong, J.; Chen, L.; Snitow, M.; Morley, M.; Li, D.; Petrenko, N.; et al. A microRNA-Hippo pathway that promotes cardiomyocyte proliferation and cardiac regeneration in mice. Sci. Transl. Med. 2015, 7, 279ra38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pohjoismäki, J.L.; Goffart, S. The role of mitochondria in cardiac development and protection. Free Radic. Biol. Med. 2017, 106, 345–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, D.; Jian, C.; Peng, Q.; Hou, T.; Wu, K.; Shang, B.; Zhao, M.; Wang, Y.; Zheng, W.; Ma, Q.; et al. Prohibitin 2 deficiency impairs cardiac fatty acid oxidation and causes heart failure. Cell Death Dis. 2020, 11, 181. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Long, B.; Jiao, J.-Q.; Wang, J.-X.; Liu, J.-P.; Li, Q.; Li, P.-F. miR-484 regulates mitochondrial network through targeting Fis1. Nat. Commun. 2012, 3, 781. [Google Scholar] [CrossRef]
- Forsburg, S.L. Eukaryotic MCM proteins: Beyond replication initiation. Microbiol. Mol. Biol. Rev. 2004, 68, 109–131. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Tan, B.; Yang, Z.; Yu, X.; Chen, L.; Ran, D.; Xu, Q.; Zhou, X. Nrf2/ARE pathway activation is involved in negatively regulating heat-induced apoptosis in non-small cell lung cancer cells. Acta Biochim. Biophys. Sin. 2020, 52, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Boon, R.A.; Jaé, N.; Holdt, L.; Dimmeler, S. Long noncoding RNAs: From clinical genetics to therapeutic targets? J. Am. Coll. Cardiol. 2016, 67, 1214–1226. [Google Scholar] [CrossRef]
- Choong, O.K.; Lee, D.S.; Chen, C.-Y.; Hsieh, P.C. The roles of non-coding RNAs in cardiac regenerative medicine. Non-Coding RNA Res. 2017, 2, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Sarropoulos, I.; Marin, R.; Cardoso-Moreira, M.; Kaessmann, H. Developmental dynamics of lncRNAs across mammalian organs and species. Nature 2019, 571, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, E.J.; van Solingen, C.; Scacalossi, K.R.; Ouimet, M.; Afonso, M.S.; Prins, J.; Koelwyn, G.J.; Sharma, M.; Ramkhelawon, B.; Carpenter, S.; et al. The long noncoding RNA CHROME regulates cholesterol homeostasis in primates. Nat. Metab. 2019, 1, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Y.; Busch, A.; Jin, H.; Chernogubova, E.; Pelisek, J.; Karlsson, J.; Sennblad, B.; Liu, S.; Lao, S.; Hofmann, P.; et al. H19 induces abdominal aortic aneurysm development and progression. Circulation 2018, 138, 1551–1568. [Google Scholar] [CrossRef] [PubMed]
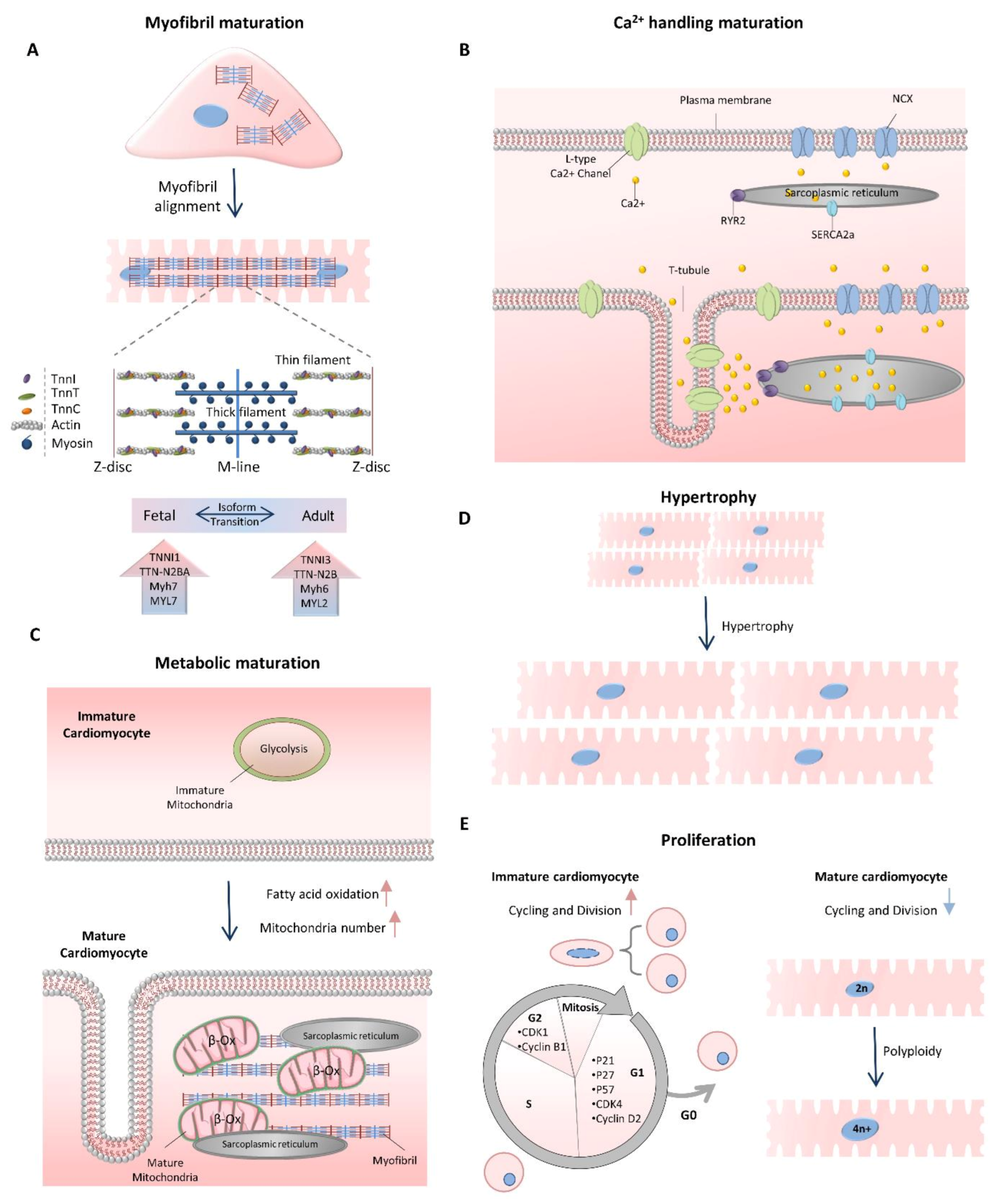
Figure 1.
Cardiomyocyte maturation characteristics. (A) Cardiomyocytes undergo myofibril maturation, which is followed by increasing the sarcomere length, assembly and expansion; improvement of sarcomere alignment and M-line formation. (B) Calcium handling maturation is mediated by the development of T-tubules, expression of calcium handling proteins and increased volume of calcium stores of the sarcoplasmic reticulum. (C) Metabolic maturation causes the switch of glycolysis to fatty acid β-oxidation and increases in mitochondria number. The cristae structures in mitochondria are densely organized, and the mitochondria are localized close to the SR and myofibrillar structures to support efficient ATP transition. (D,E) Proliferation-to-hypertrophy transition occurs at the time of maturation. The postnatal heart size is increased through maturational hypertrophy followed by polyploidization (top) cardiomyocyte proliferation rate declines through downregulation of cell-cycle-associated genes (bottom).
Figure 1.
Cardiomyocyte maturation characteristics. (A) Cardiomyocytes undergo myofibril maturation, which is followed by increasing the sarcomere length, assembly and expansion; improvement of sarcomere alignment and M-line formation. (B) Calcium handling maturation is mediated by the development of T-tubules, expression of calcium handling proteins and increased volume of calcium stores of the sarcoplasmic reticulum. (C) Metabolic maturation causes the switch of glycolysis to fatty acid β-oxidation and increases in mitochondria number. The cristae structures in mitochondria are densely organized, and the mitochondria are localized close to the SR and myofibrillar structures to support efficient ATP transition. (D,E) Proliferation-to-hypertrophy transition occurs at the time of maturation. The postnatal heart size is increased through maturational hypertrophy followed by polyploidization (top) cardiomyocyte proliferation rate declines through downregulation of cell-cycle-associated genes (bottom).
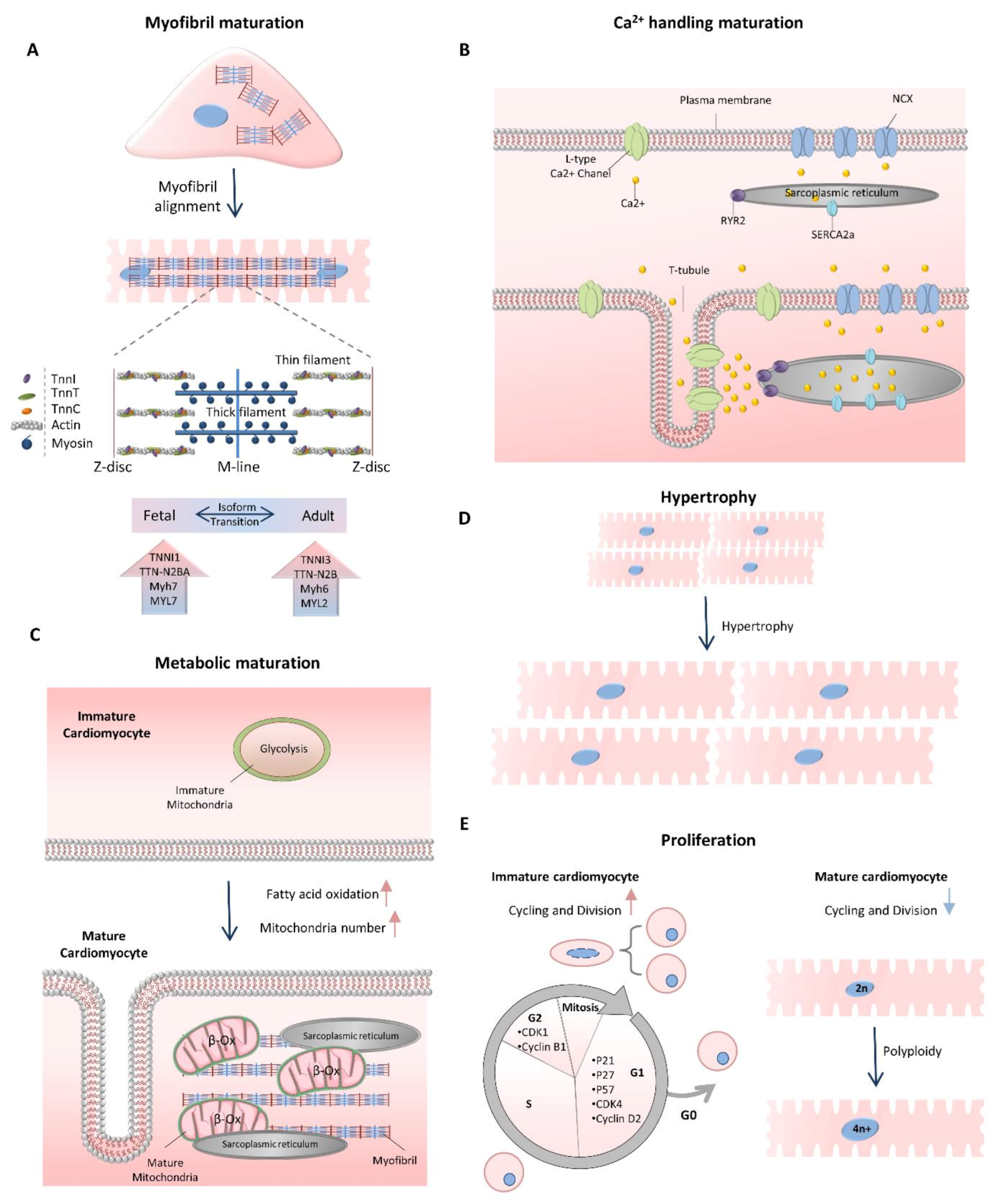
Figure 2.
Classification of LncRNA. (A) LncRNAs may be divided based on their position and orientation relative to protein-coding genes (genomic context) or by distinct chromatin marks around their transcription start-sites (chromatin context). (B) More details are described within the text.
Figure 2.
Classification of LncRNA. (A) LncRNAs may be divided based on their position and orientation relative to protein-coding genes (genomic context) or by distinct chromatin marks around their transcription start-sites (chromatin context). (B) More details are described within the text.
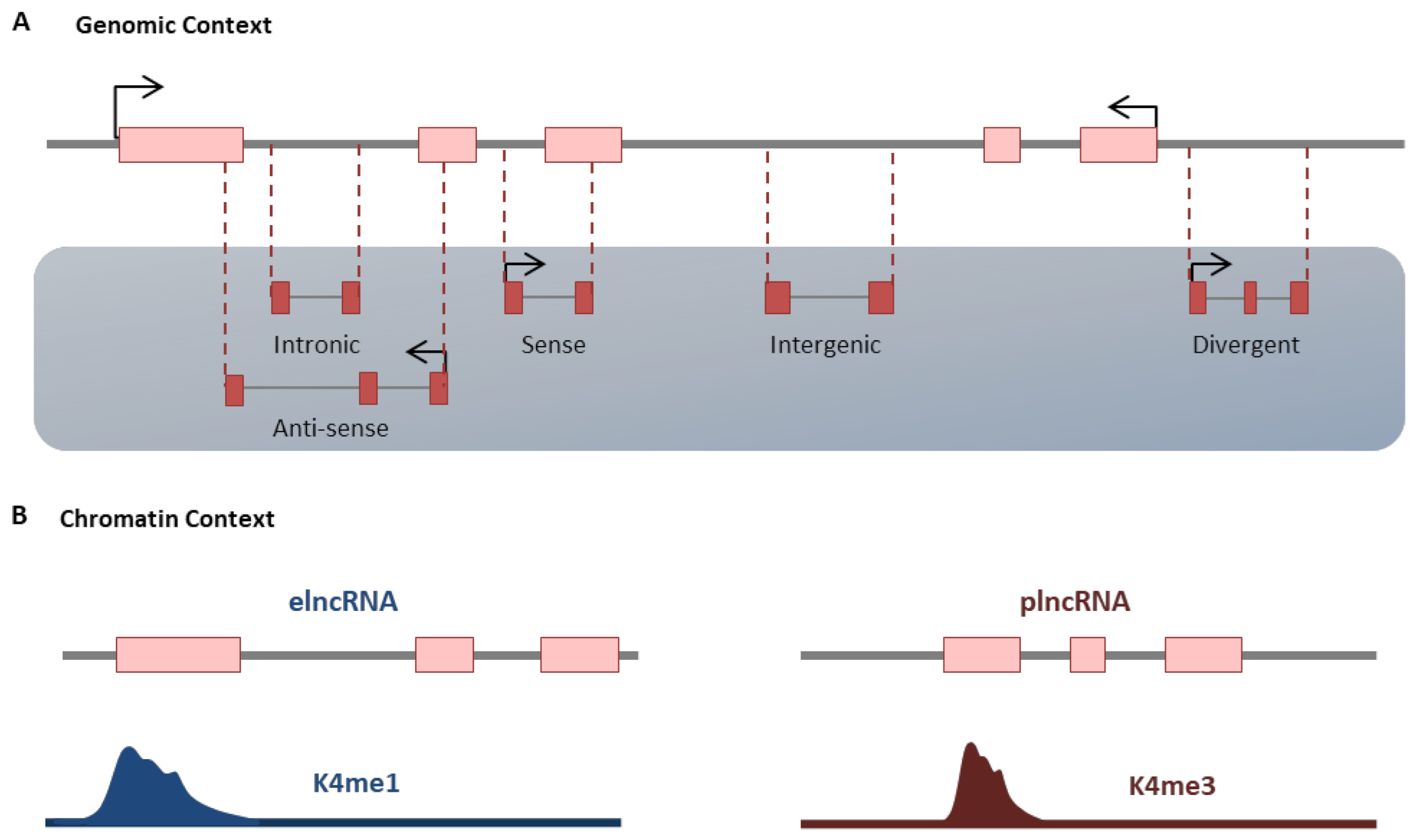
Figure 3.
Roles of Mhrt-LncRNA in cardiomyocyte maturation. Mhrt-lncRNA has a different regulatory function in myofibril formation and hypertrophy at the stage of cardiomyocyte maturation. Mhrt regulates the Myh6/Myh7 ratio and their expression during the development through Brg1 inhibition. Cardiomyocyte hypertrophy regulation is also occurred by Mhrt through myocardin expression and function regulation. More details are described within the text.
Figure 3.
Roles of Mhrt-LncRNA in cardiomyocyte maturation. Mhrt-lncRNA has a different regulatory function in myofibril formation and hypertrophy at the stage of cardiomyocyte maturation. Mhrt regulates the Myh6/Myh7 ratio and their expression during the development through Brg1 inhibition. Cardiomyocyte hypertrophy regulation is also occurred by Mhrt through myocardin expression and function regulation. More details are described within the text.
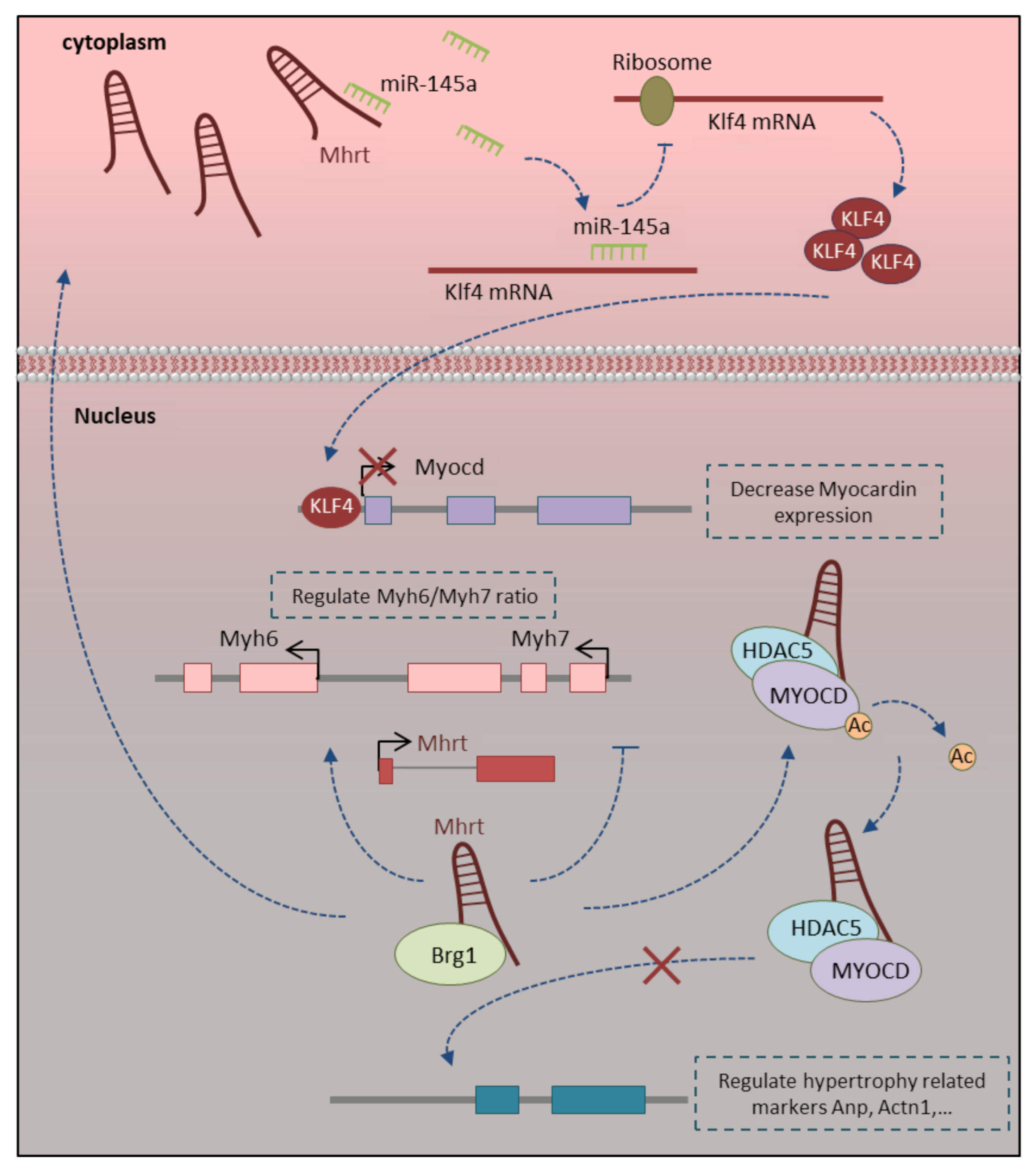
Figure 4.
Regulatory effect of H19-LncRNA in cardiomyocyte maturation. (A) H19 regulates the cardiomyocyte maturation process at different steps, including apoptosis, proliferation, hypertrophy and contraction. A. H19 physically interacts with PRC2 to suppress pro-hypertrophic NFAT signaling. (B) H19 regulates the cardiac ca2+ flux and cardiac contraction function through the H19-miR-675-CamkIIδ axis. (C,D) The regulatory function of H19 lncRNA in mitochondrial apoptotic pathway and cardiomyocyte proliferation is mediated through either H19-miR-675-Vdac1 (C) or H19-miR-19b-Sox6 (D) axis. More details are described within the text.
Figure 4.
Regulatory effect of H19-LncRNA in cardiomyocyte maturation. (A) H19 regulates the cardiomyocyte maturation process at different steps, including apoptosis, proliferation, hypertrophy and contraction. A. H19 physically interacts with PRC2 to suppress pro-hypertrophic NFAT signaling. (B) H19 regulates the cardiac ca2+ flux and cardiac contraction function through the H19-miR-675-CamkIIδ axis. (C,D) The regulatory function of H19 lncRNA in mitochondrial apoptotic pathway and cardiomyocyte proliferation is mediated through either H19-miR-675-Vdac1 (C) or H19-miR-19b-Sox6 (D) axis. More details are described within the text.

Figure 5.
The regulatory function of main lncRNAs related to cardiomyocyte maturation. (A) Zfas1 and Dach1 manipulate the cardiac function by regulating Ca2+ homeostasis, proliferation and apoptosis in mature cardiomyocytes. This happened through negative regulation of Serca2a through either its expression repression or functional restriction and regulating the Hippo signaling pathway. (B) Carl and Mdrl-lncRNAs regulate mitochondrial fission and apoptosis through regulating the combination of miRNAs and transcription factors in adult cardiomyocytes. Carl lncRNA sponges miR-539 in the cardiomyocytes, which in turn upregulates the expression of Phb2 as its target gene. Mdrl-lncRNA has direct interaction with miR-361 and downregulates its expression levels, which in turn promotes the processing of pri-miR-484 and represses the Fis1 expression. (C) Cpr lncRNA attenuates the cardiomyocyte proliferation in postnatal and adult hearts through direct interaction and recruitment of DNMT3A protein Mcm3 promoter to repress its expression. Sarrah lncRNA regulates cardiomyocyte survival and enhances the contractile capacity of adult cardiomyocytes through activating the NRF2 signaling pathway. Sarrah directly binds to the promoter region of the Nrf2 gene through RNA-DNA triple helix formation and recruits CRIP2 and p300 transcription factors to induce its expression. More details are described within the text.
Figure 5.
The regulatory function of main lncRNAs related to cardiomyocyte maturation. (A) Zfas1 and Dach1 manipulate the cardiac function by regulating Ca2+ homeostasis, proliferation and apoptosis in mature cardiomyocytes. This happened through negative regulation of Serca2a through either its expression repression or functional restriction and regulating the Hippo signaling pathway. (B) Carl and Mdrl-lncRNAs regulate mitochondrial fission and apoptosis through regulating the combination of miRNAs and transcription factors in adult cardiomyocytes. Carl lncRNA sponges miR-539 in the cardiomyocytes, which in turn upregulates the expression of Phb2 as its target gene. Mdrl-lncRNA has direct interaction with miR-361 and downregulates its expression levels, which in turn promotes the processing of pri-miR-484 and represses the Fis1 expression. (C) Cpr lncRNA attenuates the cardiomyocyte proliferation in postnatal and adult hearts through direct interaction and recruitment of DNMT3A protein Mcm3 promoter to repress its expression. Sarrah lncRNA regulates cardiomyocyte survival and enhances the contractile capacity of adult cardiomyocytes through activating the NRF2 signaling pathway. Sarrah directly binds to the promoter region of the Nrf2 gene through RNA-DNA triple helix formation and recruits CRIP2 and p300 transcription factors to induce its expression. More details are described within the text.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
LncRNAs with potential roles in the cardiomyocyte maturation process.
| Maturation Characteristics | LncRNA | Function | Regulated Target | Ref |
|---|---|---|---|---|
| Myofibril formation | Mhrt | Regulation of Myh6/Myh7 ratio | Brg1 | [77] |
| Cpr | Sarcomere organization | Mcm3 | [78] | |
| Ppp1r1b | Cardiac myogenesis regulation | Tcap | [79] | |
| Electrophysiology | H19 | CaMKIIδ expression regulation | miR-675 | [80] |
| Zfas1 | Ca2+ homeostasis regulation | Serca2a | [81] | |
| Dach1 | Ca2+ homeostasis regulation | Serca2a | [82] | |
| Tincr | CaMKII expression regulation | EZH2 | [83] | |
| Ccrr | Cardiac conduction regulation | CIP85 | [84] | |
| Kcna2-AS | Cardiac conduction regulation | Kv1.2 | [85] | |
| Malat1 | Cardiac conduction regulation | miR-200c | [86] | |
| ZNF593-AS | Cardiac conduction regulation | HNRNPC | ||
| Hotair | Regulate Cx43 expression | miR-613 | [87] | |
| Metabolism | Carl | Regulate metabolic maturation | miR-539 | [88] |
| Mdrl | Regulate metabolic maturation | miR-361miR-484 | [89] | |
| TTTY15 | Regulate mitochondrial energy metabolism | let-7i-5p | [90] | |
| Proliferation and hypertrophy | Mhrt | Hypertrophy regulation | Hdac5 | [91] |
| Hypertrophy regulation/Myocd regulation | miR-145-5p | [92] | ||
| H19 | Hypertrophy regulation/NFAT signaling | Prc2 | [93] | |
| Mitochondria-mediated apoptosis regulation by PBX3-dependent way | miR-675 CYP1B1 | [94] | ||
| [95] | ||||
| Ahit | Hypertrophy regulation/Mef2A regulation | Suz12-Prc2 | [96] | |
| Zfas1 | Mitochondria-mediated apoptosis | Serca2a | [97] | |
| Dach1 | Proliferation regulation/Hippo signaling | PP1A | [98] | |
| Cpr | Proliferation regulation | Mcm3 | [78] | |
| Sarrah | Apoptosis and survival regulation/Nrf2 pathway | CRIP2 and p300 | [99] | |
| Chaer | Hypertrophy regulation | Prc2 | [100] | |
| Meg3 | Hypertrophy regulation | miR-361-5p | [101] | |
| Apoptosis regulation/NF-κB signaling | P53 | [102] | ||
| Plscr4 | Hypertrophy regulation | miR-214 | [103] | |
| Chast | Hypertrophy regulation | Plekhm1 | [104] | |
| Chrf | Hypertrophy regulation | miR-489 | [105] | |
| Snhg1 | Hypertrophy regulation | miR-15a-5p | [106] | |
| Apoptosis regulation | miR-195 | [107] | ||
| Apoptosis regulation | miR-188-5p | [108] | ||
| Uc.323 | Hypertrophy regulation | Ezh2 | [109] | |
| Magi1-IT1 | Hypertrophy regulation/Wnt signaling | miR-302e | [110] | |
| Ecrar | Proliferation regulation/ERK1/2 signaling | ERK1/2 | [111] | |
| Crrl | Proliferation regulation/HOPX regulation | miR-199a-3p | [112] | |
| Uca1 | Hypertrophy regulation | miR-184 | [113] | |
| NR_045363 | Proliferation regulation/JAK2-STAT3 signaling | miR-216a | [114] | |
| lncCIRBIL | Apoptosis regulation | Bclaf1 | [115] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kay, M.; Soltani, B.M. LncRNAs in Cardiomyocyte Maturation: New Window for Cardiac Regenerative Medicine. Non-Coding RNA 2021, 7, 20. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7010020
AMA Style
Kay M, Soltani BM. LncRNAs in Cardiomyocyte Maturation: New Window for Cardiac Regenerative Medicine. Non-Coding RNA. 2021; 7(1):20. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7010020
Chicago/Turabian StyleKay, Maryam, and Bahram M. Soltani. 2021. "LncRNAs in Cardiomyocyte Maturation: New Window for Cardiac Regenerative Medicine" Non-Coding RNA 7, no. 1: 20. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7010020
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.