
Emerging Data on the Diversity of Molecular Mechanisms Involving C/D snoRNAs


Campus Brabois Santé, Université de Lorraine, CNRS, IMoPA, F-54000 Nancy, France
*
Authors to whom correspondence should be addressed.
Non-Coding RNA 2021, 7(2), 30; https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7020030
Submission received: 12 April 2021
/
Revised: 28 April 2021
/
Accepted: 30 April 2021
/
Published: 6 May 2021
(This article belongs to the Special Issue Non-coding RNA: 5th Anniversary)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Box C/D small nucleolar RNAs (C/D snoRNAs) represent an ancient family of small non-coding RNAs that are classically viewed as housekeeping guides for the 2′-O-methylation of ribosomal RNA in Archaea and Eukaryotes. However, an extensive set of studies now argues that they are involved in mechanisms that go well beyond this function. Here, we present these pieces of evidence in light of the current comprehension of the molecular mechanisms that control C/D snoRNA expression and function. From this inventory emerges that an accurate description of these activities at a molecular level is required to let the snoRNA field enter in a second age of maturity.
1. Introduction
On several occasions, the fortuitous identification of RNAs with odd features turned out to be at the origin of a new family of non-coding RNAs (ncRNAs). A perfect example is the characterization of development timing mutants in C. elegans, leading to the identification of tiny ncRNAs originally named small temporary RNAs (stRNAs) lin-4 and let-7 [1,2]. These two RNAs were, in fact, part of the microRNAs class, which now has more than 1000 members. The discovery of the box C/D small nucleolar RNAs (C/D snoRNAs) is another striking case. Abundant low molecular weight RNAs were first identified in nuclear [3] and nucleolar fractions [4,5]. Nucleolar U3 RNA was later demonstrated contributing at an early stage of the ribosomal RNA (rRNA) processing pathway by acting as a chaperone folding the nascent primary transcript for the first endonucleolytic cleavages [6,7,8,9]. Next, U3 conserved sequences and common associated proteins were subsequently identified for other nucleolar ncRNAs leading to classify hundreds of snoRNAs with U3 in the family of the box C/D snoRNAs [10,11,12,13,14]. A large number of the box C/D snoRNAs were shown forming small nucleolar ribonucleoproteins (snoRNPs) to participate in ribosome biogenesis (for recent reviews, see [15,16,17,18,19]). However, U3 snoRNP appeared singular since most of the box C/D snoRNAs are components of RNP enzymes catalyzing the site-specific 2′-O-methylation of functionally important regions in rRNA. Concomitantly, a second large family of snoRNAs with conserved sequences boxes H and ACA were discovered [20,21,22] (for reviews, see [15,23]). The box H/ACA snoRNAs form snoRNPs that are RNA-guided modification enzymes targeting uridines for their isomerization into pseudouridines. As for box C/D snoRNPs, a few H/ACA snoRNAs are also involved in pre-rRNA processing machinery [24,25] (for reviews, see [15,16,18,19]). Subsequently, small Cajal Bodies RNAs (scaRNAs) with similar features as snoRNAs and forming equivalent catalytic RNPs were found enriched in Cajal bodies (CBs, for a review, see [26]). In these nuclear bodies, some are the catalysts for ribose 2′-O-methylation and for pseudouridylation inside functionally important sequences of spliceosomal small nuclear RNAs (snRNAs) (for a review, see [27]). Hence, RNA-guided modification is not the prerogative of snoRNPs, and scaRNPs illustrated a potential of diversity towards substrate specialization depending on their nuclear localization.
Data accumulated this last decade added diversity to the pattern of expression, the place, the partners, and the functions of snoRNAs that could not still be perceived as confined to ribosome-associated functions. On the other hand, several of these RNAs still have no identified function, while some of them have been identified as key actors in human pathologies. Here, we compare novel features recently discovered for box C/D snoRNPs with the conventional ones that are firmly established. We focus on the box C/D sno/scaRNAs and their associated RNPs as more structure, and function diversity is up to now identified among this subclass compared with box H/ACA sno/scaRNPs. Importantly, complementary and detailed information on the (patho)physiology of both C/D and H/ACA snoRNAs, including in vivo knockout models, are available in recent reviews [15,28,29,30,31,32,33].
2. Molecular Features for the Canonical Paradigm of RNA-Guided Activity: The Box C/D RNP Methyltransferases
It is probable that eukaryotic snoRNAs originate from an ancestor present before the separation between Eukarya and Archaea (for a review, see [34]). Indeed, small RNAs with box C/D or box H/ACA snoRNA features, named sRNAs, are also acting for RNA-guided modification in archaeons [35,36,37,38,39,40,41] (for a review, see [42,43]). This phylogenetic feature highlights the essentiality of the cellular activity of these systems that could have expanded in eukaryotic cells with other functions and mechanisms of action.
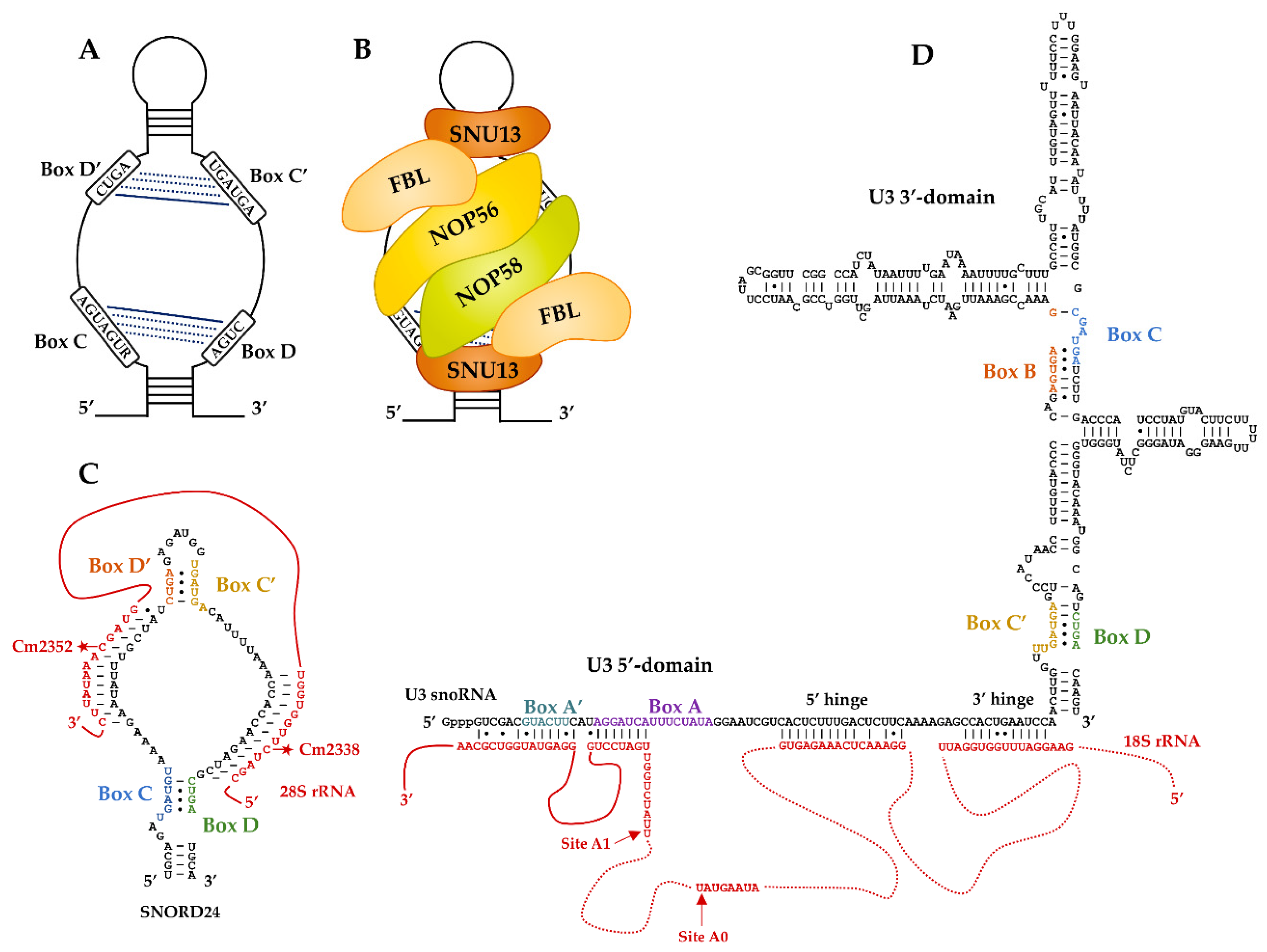
The box C/D snoRNAs have an average length of 60–90 nucleotides and are folded as a single irregular hairpin structure. They possess conserved sequence patterns forming a box C (consensus sequence 5′-RUGAUGA-3′ where R = purine) and a box D (consensus sequence 5′-CUGA-3′), which are located at the 5′- and 3′-ends of snoRNAs, respectively. In most cases, additional boxes designated C′ and D′ are present in the internal region of the transcript; however, these sequences are less well conserved and sometimes degenerated [11,44]. In the secondary structure of the snoRNA transcripts, the conserved boxes are brought together and form the C/D and C′/D′ motifs (Figure 1A). The C/D motif corresponds to a structural element organized as a kink-turn (K-turn), which is characterized by a 3-nucleotide asymmetric internal bulge surrounded by two stems I and II and comprising at the edge of stem II tandem G:A and A:G sheared base pairs. In the 3D structure, a sharp bend of the ribose-phosphate backbone is formed that is stabilized by stacking interactions and featuring a protruded uridine invariably present in the internal budge [14,45,46,47] (for a review, see [48]). This type of motif is also present in U4 snRNA [14,49,50] and in both box C/D and box H/ACA archaeal sRNAs [38,51,52]. The motif C′/D′ in archaeal sRNAs is organized as a K-loop lacking stem I and closed by a terminal loop [53]. The box C/D snoRNAs K-turn motif constitutes a specific binding site for Snu13p/SNU13 (in Yeast and Human, respectively) that nucleates RNP formation through the subsequent recruitment of Nop58p/NOP58 and the SAM-dependent 2′-O-methylase Nop1p/FBL [14,54,55]. The C′/D′ motif constitutes a secondary Snu13p/SNU13 binding site leading to the assembly of a second half-particle through recruitment of the Nop56p/NOP56–Nop1p/FBL module [50,56,57] (Figure 1B). Based on structural studies performed on the archaeal components [58,59] (for a review, see [15]), it has been proposed that the sub-RNPs assembled on each RNA motif are connected through the coiled-coil dimer of paralogous Nop56p/NOP56 and Nop58p/NOP58. Recent reconstitution of C/D snoRNP particle from the thermophilic yeast Chaetomium thermophilum indicates that an in vivo assembly machinery allows the specific assembly of Nop58 and Nop56 on the C/D and C′/D′ motifs, respectively [60].
Guide activity of box C/D snoRNAs relies on the sequence located immediately upstream of boxes D and D′ that forms base pairs with the RNA substrate and specifies the site of ribose 2′-O-methylation. The modified residue is the fifth nucleotide upstream of the D or D′ sequences [11,63] (Figure 1C). Information on the molecular bases for the positioning of the active site of the catalytic subunit of the RNP was gained by structural studies performed with reconstituted archaeal box C/D RNPs [58,64]. The sequence potentially available for substrate binding usually has a length of 10 to 21 nucleotides [11,65,66]. One study based on 3D structures of artificial yeast sRNPs hybridized to RNA substrates proposed that a maximum of 10 bp can be accommodated by such particles [67]. Conversely, a subset of Saccharomyces cerevisiae, human, and Arabidopsis thaliana box C/D snoRNAs were shown to have extra base-paired regions leading to enhanced methylation [44,68,69].
3. U3, the First Identified Atypical Box C/D RNP
As mentioned in the introduction, U3 has atypical features. First, although it contains the 2′-O-methyltransferase Nop1p/FBL, the U3 snoRNP has never been shown to be a catalyst for RNA 2′-O-methylation. Second, the U3 snoRNA contains four phylogenetically conserved sequences that form two non-conventional box C/D-like motifs named C′/D and B/C (Figure 1D). Each forms a K-turn structure recognized by Snu13p/SNU13 [14,70,71,72]. However, protein assembly on these two motifs is dissymmetric. A specific U3 core protein Rrp9p in yeast and U3-55K in human [73,74,75] is recruited on the B/C motif in a Snu13p/SNU13-dependent manner [72,76,77,78] together with Nop56p/NOP56 and Nop1p/FBL [79,80]. In contrast, the classical set of C/D box proteins Snu13p/SNU13, Nop58p/NOP58, and Nop1p/FBL is recruited on the C′/D motif [14,81]. Recent cryo-EM structures from S. cerevisiae and Chaetomium thermophilum confirmed this pseudo-symmetric organization of U3 snoRNP [61,82,83,84].
Compared with other C/D snoRNAs, U3 has a long 5′-terminal region needed for the function of the particle during pre-rRNA processing [7,85,86,87,88,89]. Pre-rRNA maturation begins in terminal knobs corresponding to the packaging of the nascent rRNA primary transcript into a large structure, named small subunit (SSU) processome. The U3 snoRNP contributes to the assembly of the SSU processome [9]. The U3 snoRNA plays a central role by forming base-pair interactions with several sequences of the 5′ external transcribed spacer (5′-ETS) and the 18S rRNA (Figure 1D). It is viewed as an organizing chaperone to RNA folding into 5 distinct helices that potentiate early cleavage at sites A0 and A1 in the 5′-ETS. The occurrence of helices was investigated by biochemical and genetic approaches [7,62,85,86,87,88,89]. The cryoEM structures confirmed that several of them are formed within the processome [61,82,83,84].
4. Biogenesis of Box C/D RNP: Assembly and Nuclear Journey
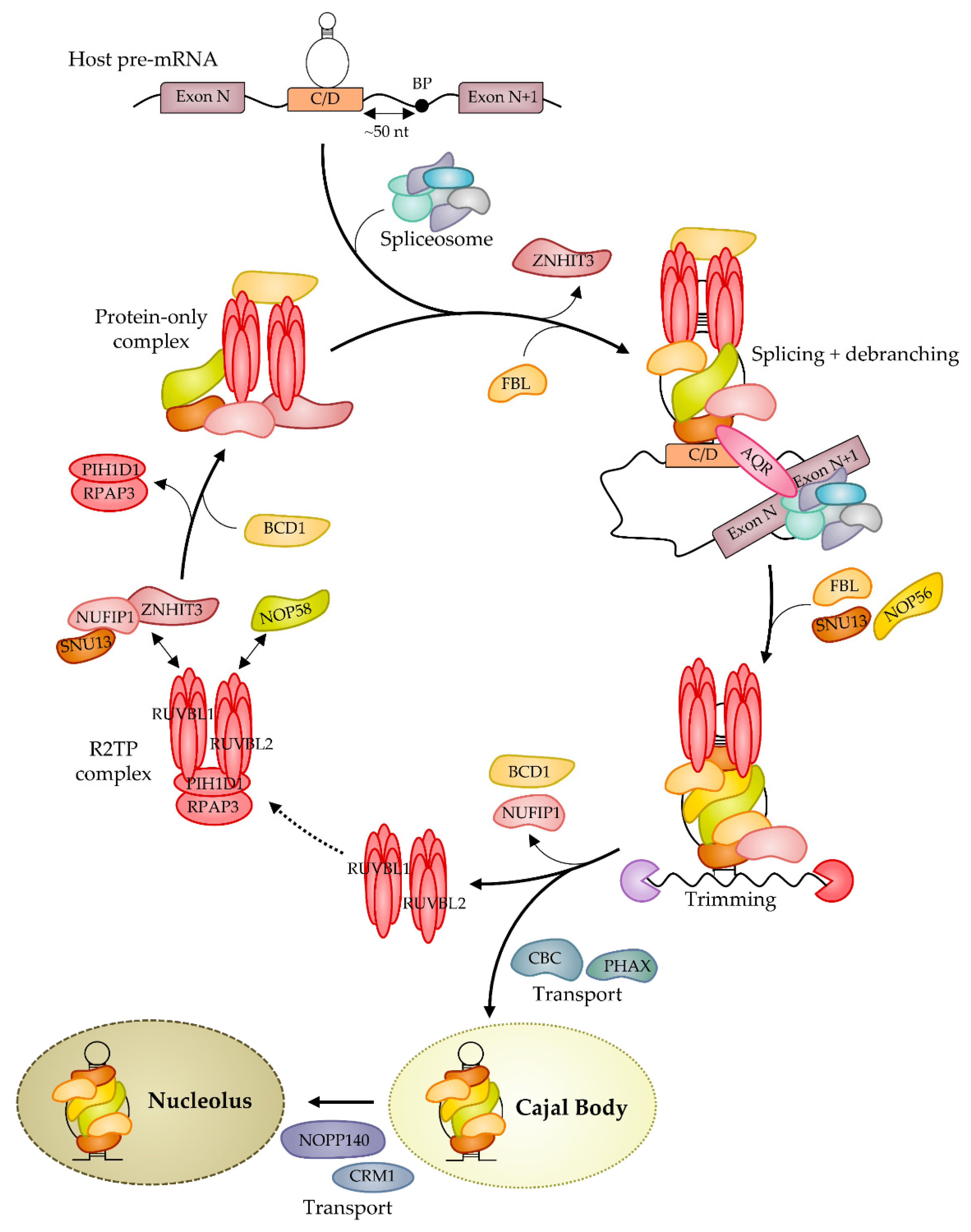
Biogenesis of snoRNPs is a complex process involving trans-acting factors and starting on nascent transcripts in a co-transcriptional fashion [15,33,90] (Figure 2). In most organisms, snoRNAs are transcribed as long precursors by RNA pol II that are trimmed by ribonucleases at 3′- or both 5′- and 3′-ends to generate mature snoRNAs (for a review, see [30]). Early recognition of snoRNA elements by core RNP proteins protects RNA from degradation during the processing. The specific mode of processing is determined by the genomic organization. In vertebrates, a subset of snoRNAs is encoded by independent transcriptional units, but the majority are embedded in introns of host protein-coding genes. In contrast, to box H/ACA snoRNAs, most box C/D snoRNAs are present at a conserved position upstream of the 3′ splicing acceptor site [91,92], and an optimal distance of ~50 nucleotides upstream of the branch point contributes to efficient box C/D snoRNP assembly [93] (for a review, see [94]). This situation favors splicing-dependent assembly of intronic box C/D snoRNPs and the general splicing factor Aquarius AQR (alias Intron Binding Protein IBP160 [95]) was shown to aid RNP assembly [96]. However, some intronic C/D snoRNA genes are found at divergent locations, such as the human repeated snord116 genes that are each sited ~250 bp after the 5′-splice site of the host gene. Then, considering independent snoRNA-encoded transcripts like the U3 snoRNA, the presence of the core proteins was shown to be linked to 3′-end processing and transcription termination [97,98].
Contrary to archaeal sRNPs [37], the assembly of eukaryotic snoRNPs is not a mechanism that occurs autonomously in vitro. Independently of the coupling with splicing machinery as described above, biogenesis requires trans-acting factors involved in RNP assembly and in nuclear trafficking before the mature RNP reaches the nucleolus. Three independent modules Rsa1p:Hit1p/NUFIP1:ZNHIT3 [98,99], the R2TP complex [100,101,102] and Bcd1p/BCD1(ZNHIT6) [90,101,103,104] were identified contributing to snoRNP assembly. Inactive pre-RNPs are first assembled on nascent transcripts before reaching the CBs where they become mature and functional (for a review, see [33,105]). A model has been proposed postulating that the formation of human box C/D snoRNPs requires the formation of an RNA-free pre-complex composed of the two core proteins SNU13 and NOP58 as well as five assembly factors NUFIP1, ZNHIT3, BCD1, RUVBL1, and RUVBL2 [99]. Rsa1p/NUFIP1 is a platform protein stabilized by the factor Hit1p/ZNHIT3 that forms a tertiary complex with Snu13p/SNU13 [71,99,101]. The two ATPases associated with diverse cellular activities (AAA+) RUVBL1 and RUVBL2 (RUVBL1/2, alias TIP49, and TIP48) are members of the complex R2TP, which also includes PIH1D1 (Pih1p or Nop17p in yeast) and RPAP3 (Tah1p in yeast) [100,106,107,108]. This complex cooperates with the chaperone HSP90 and participates in the stabilization and the recruitment of NOP58 on the RNP by direct interaction with PIH1D1 [109]. NUFIP1 interacts with SNU13 during the first stages of maturation and blocks the catalytic activity of the complex [99]. ZNHIT3 is then released, and the C/D box snoRNA, as well as FBL, are recruited before binding of the second molecule of FBL and NOP56. NUFIP1 dissociates from the complex, generating a rotation of the catalytic module of FBL, leading to an active C/D box snoRNP complex. Several proteins manage the nuclear trafficking of the pre-snoRNPs, such as CBC and PHAX for the transfer to CBs (or WDR79 [110,111] and TDP-43 [112] for the scaRNPs) and NOPP140 and CRM1 that control the nucleolar localization [33,113]. All categories of snoRNA traffic through CBs [114]. The cap m7Gppp is a key determinant for U3 to reach CBs, and after hypermethylation, CREM intervenes for reaching nucleoli [115,116] (for reviews, see [113,117]).
5. The Diversity of C/D snoRNA Maturation Forms
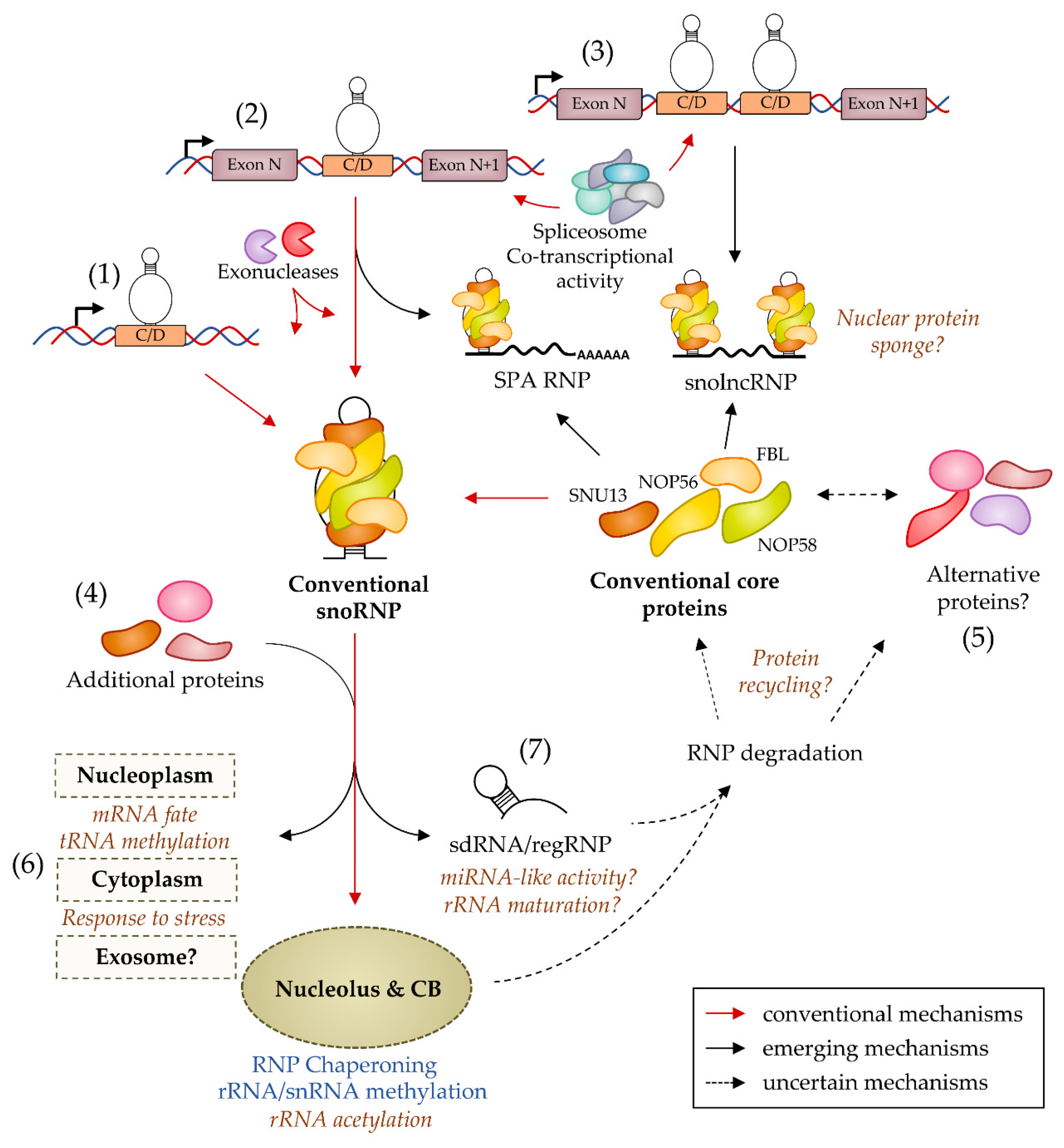
Precursor snoRNA processing accepts different outcomes, at least in Mammals. First, the pre-snoRNA could be only partially matured to generate long non-coding RNAs (lncRNAs) flanked by snoRNA sequences at one or both extremities (Figure 3). In humans, these RNA species were first described as products of the tandemly repeated snord116 genes at the imprinted PWS locus. This cluster is transcribed as one (or several) precursor RNA(s) whose introns host the snord116 sequences. Intron processing generates conventional snord116 snoRNPs [118] that reach the nucleolus, while the spliced host transcripts accumulate near the transcription site [119]. However, some molecules may undergo unusual processing when certain exons are not spliced, leaving two consecutive snoRNAs that are then trimmed only at the 5′-end for the first and 3′-end for the second. Thus, these so-called snolncRNAs are stabilized by two snoRNP structures at their extremities [120]. Subsequently, snord116 repeats were found to generate lncRNAs containing a 5′-end snoRNP and a polyadenylated 3′-end, called SPA (5′ snoRNA capped and polyadenylated lncRNA [121]). The works characterizing both lncRNAs also proposed that they act as protein factor sponges to control mRNA splicing and gene expression (Figure 3). Since then, several other lncRNAs harboring either C/D or H/ACA snoRNA sequences at their extremities have been described in human cell lines [120]. The nucleolar-specific lncRNA called LoNA is proposed to sequester the core protein FBL via its snoRNA-like 3′-end, which in turn modulates rRNA methylation levels [122]. In addition, the splicing of the human Nop56 pre-mRNA is controlled by an intron-hosted orphan snoRNA called snord86 whose structuration restricts the usage of a nearby splice site. Consequently, an excess of NOP56 protein is expected to favor the production of a cytoplasmic RNA capped at its 5′-end by the snoRNA snord86. This unusual lncRNA is generated at the expense of Nop56 mRNA production, which may constitute a negative feedback mechanism regulating NOP56 protein expression [123]. It is interesting to note that in every case the production of a conventional snoRNA is detected, which opens the possibility that the two maturation processes are in competition or that the snolncRNAs behave as stable intermediate for snoRNA synthesis.
Conventional C/D snoRNAs are also suspected of exhibiting diversity in size. Two distinct isoforms of human C/D snoRNAs differing in their terminal stem length were reported in both normal and cancer cell lines [124]. Such variations involving ± 1–2 bp at the extremities of the basal stem have been recurrently observed by us and others. More surprisingly, the short forms were proposed to form non-canonical snoRNPs based on their lower sensitivity towards NOP58 downregulation. Unfortunately, a precise characterization of these alternative RNPs and the molecular mechanisms that direct their biogenesis has not been reported yet. More surprisingly, the generalization of RNA seq analyses has unveiled another category of snoRNA gene products in the form of short fragments ranging from ~ 20 to 100 nucleotides in size that are collectively called snoRNA-derived RNAs (sdRNAs) or processed snoRNAs (psnoRNAs) (Figure 3). These sdRNAs have been reported in several organisms such as Yeast [125], Protozoans [126], Angiosperms [127], Human [128,129], and Viruses [130] and also include scaRNA derivatives [129,131]. In Human, sdRNAs have been particularly associated with cancers [132,133], and, more largely, they seem to characterize stress conditions such as evidenced in Saccharomyces cerevisiae [134] and Wheat [127]. Theoretically, the origin of sdRNAs could derive from regular snoRNAs or being produced directly from precursor RNAs. However, some observations favor the first hypothesis as sdRNAs largely correspond to sequences present in mature snoRNAs and not to surrounding regions. In addition, a study identifying a strong enrichment of sdRNAs corresponding to both extremities of the snord44 gene in HeLa cells reported that their expression was dependent on the core proteins FBL and NOP58 [135]. The relative expression of snoRNAs and associated sdRNAs seems to vary quite independently in Saccharomyces cerevisiae [134] and in Human [136], suggesting that they are not simply produced by snoRNA turnover. Nevertheless, the mechanisms of sdRNA biogenesis and regulation are poorly understood. Interestingly enough, FBL has been recently shown to exhibit an RNase activity regulated by the presence of phosphatidic acid [137], which opens the possibility that snoRNA processing is an intrinsic property of C/D snoRNPs upon specific stimuli. The interaction of sdRNAs with proteins is largely unknown, except for a small subset of sdRNAs that associate with proteins of the Argonaute family and harbor microRNA-like [129,138] or piRNA-like [139] properties. The biogenesis of the formers, also called sno-miRs, has been partially evaluated. Based on few studies, the generation of the microRNA-like fragments (also called sno-miRs) requires [129,138] or not [140] the RNase III activity of DICER and are dependent [141,142] or not [140] on the RNase III activity of DROSHA. It could be expected that the study of additional sno-miRs—if they exist [143]—could clarify the mode(s) of biogenesis. Conversely, it is not known whether most of the sdRNAs reported under various cellular conditions are functional or part of the degradome, which leads to a little-addressed question: How and when are snoRNAs degraded, and how is this regulated? To date, we have no answers, although they are crucial for understanding the fate of snoRNPs.
6. The Multiple Regulations of C/D snoRNP Biogenesis
In Mammals, the expression of intronic snoRNA genes could be uncoupled from the expression of host genes, e.g., due to alternative splicing and nonsense-mediated decay [144]. Originally, the fact that, in general, host genes exhibit a high transcription rate has suggested that an important regulation step of snoRNA expression occurs at the step of RNP biogenesis. The sophistication of eukaryotic C/D snoRNP assembly involving dedicated factors may represent a site of opportunity for regulation. In agreement, N6-methylation of adenine could interfere with K-turn formation and Snu13p/SNU13 binding, thus preventing the first step of protein assembly on the nascent snoRNA in both Yeast and Human [145,146]. Moreover, several post-translational modifications of core proteins have been identified: NOP58 stability has been shown to depends on sumoylation [147,148], and the stabilization of FBL, NOP58, and NOP56 by O-GlcNAcylation is essential for snoRNP assembly [149]. However, the elucidation of the pathways controlling these modifications is pending. The formation of the chaperone complex R2TP is dependent on nutriment availability that controls, via the mTOR pathway, the nuclear import of the R2TP components Pih1p and Tah1p by Crm1p and Kap121p in Saccharomyces cerevisiae [150]. As the R2TP is necessary for Nop58p stability, this mechanism coordinates cell growth with C/D snoRNP assembly. Other proteins could modulate biogenesis. This is the case of the human H3K27 methyltransferase EZH2 that reinforces the FBL-NOP56 interaction by directly interacting with both proteins [151,152], whereas the RNA-binding protein NPM1 is suspected to modulate biogenesis with pathological consequences in dyskeratosis congenita [153]. However, for the latter, the mechanistic evidence are not clear yet. EZH2 illustrates the emerging link between C/D snoRNP assembly and chromatin effectors nicely. In Saccharomyces cerevisiae, the assembly factor Bcd1p interacts directly with the histone chaperone Rtt106p to modulate its recruitment at active genes [154]. Rsa1p with its partner Hit1p contribute to rDNA compaction by modulating Condensin accessibility [155]. In addition, the R2TP subunits RUVBL1 and RUVBL2 are components of several chromatin modification complexes, including the NuA4 histone acetyltransferase complex [156] and the chromatin remodeling complexes SWR1 [157] and INO80 [158]. C/D snoRNP assembly factors are playing other cellular functions, e.g., the R2TP provides a platform for the assembly and maturation of multiple protein complexes such as U4 snRNP-specific proteins, PIKK or RNA polymerase complexes (for a review, see [159]), and the assembly factor Rsa1p is required for the loading of Rpl10p onto the 60S subunit [160]. While these interactions between snoRNP biogenesis and different regulatory systems theoretically permit coordination, this possibility remains largely to be tested.
7. The Strange Case of Alternative C/D snoRNPs
An intriguing question regarding the regulation of C/D snoRNP biogenesis concerns the possibility of generating RNPs with different protein compositions. Some studies using fractionation and pull-down assays have proposed the presence of snoRNAs in complexes with abundant nuclear proteins such as splicing factors but devoid of FBL [161,162]. However, the particles have not been clearly identified yet, and several caveats remain to be evacuated. Co-fractionation does not mean association, and the absence of FBL detection does not mean absence of the protein. In addition, whether the snoRNAs identified correspond to precursors or degradation fragments has not been systematically analyzed. The existence of alternative RNPs clearly deserves supplementary characterization, but if confirmed, it would be particularly interesting to identify whether these complexes are diverted from conventional snoRNP biogenesis or degradation paths. For what is known currently, the first RNA determinant of snoRNP biogenesis is the K-turn formed by the C and D boxes that are recognized by SNU13. Then, the possibility for a snoRNA to assemble with an alternative set of proteins suggests either that (i) after recognition by SNU13 and at any step of the biogenesis, the assembly process is diverted towards a different set of proteins that ultimately competes with conventional core proteins or (ii) an RNA-binding protein other than SNU13 interacts specifically with the K-turn or another snoRNA feature, allowing the biogenesis of the alternative snoRNP. Alternatively, a remodeling process leading to protein exchange may intervene on a mature C/D snoRNP, e.g., in substitution of its recycling. Thus, in addition to fine biochemical characterization, it would be important to test the dependence of these RNPs on conventional assembly factors and core proteins.
8. The Diversity of C/D snoRNA Expression
By virtue of their role in the highly coordinated process of ribosome production, snoRNAs have long been considered housekeeping actors of cell function. In agreement, their expression largely correlates with the expression of other players of ribosome synthesis and translation. In Ascomycetes, the transcription of box C/D snoRNA genes is controlled by a Homol-D box that also controls the transcription of ribosomal protein genes [163], thus providing coordination [164]. Several genes coding for snoRNP components or related regulators is under the control of the transcription factor MYC in Drosophila and Human [165], which also controls numerous genes involved in ribosome biogenesis and protein synthesis [166]. In Metazoans, most snoRNA genes are located in introns of coding genes that exhibit a bias towards nucleolar function, ribosome structure, and protein synthesis [167]. In complement, several lncRNA genes that host snoRNA genes belong to the 5′TOP (5′-terminal oligopyrimidine) gene family [168,169] characterized by high-level transcription and growth-dependent regulation [170]. In Human, transcription of the few independent snoRNA genes is linked to cell proliferation by the transcriptional cofactors and RNA helicases DDX5 and DDX17 that recruit the histone deacetylase HDAC1 at promoter regions [171,172]. Yet, a more complex expression pattern is emerging from the numerous transcriptomic analyses accumulated in recent years. This is especially prominent in cancer as biopsies typically show altered expression of subsets of snoRNA genes [173,174,175] (for a review, see [176]) and have suggested their potential as diagnostic or prognostic markers [176]. Hence, several snoRNAs have been proposed as proto-oncogenes or tumor suppressors based on in vitro analyses [177,178,179,180,181]. They are also actors of other conditions, e.g., the Prader Willi syndrome (PWS) for the snord116 snoRNAs (for a review, see [182]), or the leukoencephalopathy with brain calcifications and cysts (LCC) for the U8 snoRNA [183]. In addition, C/D snoRNA genes have been found in some viral genomes where they are expected to play specific functions as suggested by the v-snoRNA1 produced by the Epstein-Barr virus, whose expression is drastically enhanced during the lytic cycle [130]. If most viruses do not encode their own snoRNAs, C/D snoRNAs are recurrently deregulated upon viral infection, e.g., by the Chikungunya virus (CHIKV) [184], by the Murine Cytomegalovirus (MCMV) [185], or the porcine reproductive and respiratory syndrome virus (PRRSV) [186]. Moreover, gene-trap insertional mutagenesis testing 12 distinct viruses has also revealed that, among other host genes, several human snoRNA genes participate in viral replication [187] (for an extensive review of the functional interactions between viruses and C/D snoRNAs, see [188]). Apart from pathological conditions, it appears that the expression of snoRNAs could be uncoupled from the level of ribosome synthesis. In Nematode, some snoRNAs display developmentally variable expression [189], while several snoRNAs from Arabidopsis and Human have been reported to be circadian clock-regulated [190,191]. C/D snoRNAs could also exhibit tissue-specificity, e.g., a strong tropism towards cerebral expression for several mammalian-specific snoRNAs [192,193,194]. The emerging versatility in the expression of many C/D snoRNAs questions their role. Going back to the ribosome, a reflection of this variability may be seen in the variable methylation levels observed at certain rRNA positions in different tissues or cellular conditions, giving rise to the concept of the specialized ribosome [195]. Indeed, some recent reports point to variations in rRNA methylation levels during mouse development [196], during human tumorigenesis [197,198], or in cell line models [199]. It suggests that the expression and/or catalytic activity of C/D snoRNPs could be finely regulated for adaptations that are beginning to be described. In this line, modifications due to high levels of FBL in cancers have been linked to impaired translation fidelity [198], fueling the notion of ribosomopathy associated with elevated cancer risk (for a review, see [200]). Interestingly, a similar scheme likely applies to snRNA methylation levels that modulate mRNA splicing [201].
9. The Diversity of C/D snoRNA Molecular Partners and Targets
Accumulating data support the notion that C/D snoRNAs perform molecular functions beyond the conventional ones. First, snoRNAs can methylate a broader set of targets than expected. It includes human tRNAs, affecting their nucleolytic cleavage [202], while in Archaea, the methylation of tRNAs by sRNAs was already described two decades ago [203,204,205]. In addition, human C/D snoRNAs could direct mRNA methylation and hence modulate the translation level of their target [206]. Interestingly, these data suggest that eukaryotic C/D snoRNPs could perform functions outside their main sites of accumulation, i.e., the nucleolus and CBs, which was already suggested by the fact that scaRNAs exhibit snRNA modification activity in the nucleoplasm in the absence of CBs [207]. Conversely, C/D snoRNPs target rRNAs in the nucleolus as expected but guide non-classical enzymatic activities, such as the yeast orphan snR4 and snR45 that promote rRNA acetylation catalyzed by Kre33p [208]. In addition, whether orphan or not, many snoRNAs have the potential to hybridize with cellular RNAs, including mRNAs as suggested by the in silico identification of energetically stable complementarities [209,210]. In agreement, studies using human cell lines have confirmed that the downregulation of specific snoRNAs affects in various ways mRNA targets, e.g., the modulation of steady-state levels by snord83b [211], of splicing by snord115 [212] and snord27 [162], or editing modulation by snord115 [213,214]. However, there is still no clear demonstration of the physical interaction between endogenous snoRNAs and their mRNA targets. It has to be mentioned that this could be a difficult task as the interaction may be limited in space and time, e.g., if occurring co-transcriptionally during pre-mRNA synthesis. Mechanistically, it could be supposed that C/D snoRNP binding induces steric hindrance and/or perturbates co-transcriptional pre-mRNA maturation processes. However, other mechanisms closer to conventional functions, such as RNP chaperoning, could also be at work, which deserves to be clarified.
A growing list of proteins has been identified to associate with C/D snoRNPs and to modulate their activity. Several of these interactions concern nucleolar proteins. In the nucleolus, C/D snoRNPs interaction with rRNA is controlled by factors modulating the affinity and specificity of the interaction. The RNA helicases HAS1 [215], DHR1 [216], and DHX37 [217] trigger the release of the U14 and U3 snoRNPs, respectively, from the ribosomal particle. The yeast ATPases Dbp3p or Prp43p promote rRNA methylation likely by favoring the recycling efficiency of C/D snoRNPs [218,219]. Concerning scaRNAs, the RNA-binding protein LARP7 increases the efficiency of methylation-guided activity by bridging scaRNAs with U6 snRNA, and its deficiency is associated with splicing alterations in Alazami syndrome [220]. Very interestingly, many other interactions with functional outcomes have been identified in the last years. The SNORD50A-SNORD50B locus produces snoRNAs that interact with the FIP1 subunit of the cleavage and polyadenylation specificity factor (CPSF) to modulate mRNA 3′-end processing [221], with the proto-oncogene KRAS to inhibit its activity [222,223] or with the E3 ubiquitin ligase TRIM21 and its substrate Guanosine 5′-monophosphate synthase (GMPS) to increase their interaction [224]. Numerous C/D and H/ACA snoRNAs have been proposed to interact with the poly-ADP-ribosyltransferase PARP1 independently of DNA damage and to stimulate its catalytic activity in the nucleolus, leading to ADP-ribosylation of the RNA helicase DDX21 and increased rDNA transcription [225]. The RNA-binding protein FMRP has been shown to interact with a subset of C/D snoRNPs, and its downregulation affects rRNA methylation [226], which might also be due to its interaction with ribosomal proteins [227,228]. The interaction of the RNA-binding protein TDP43 with a subset of C/D scaRNAs is necessary for their localization to CBs instead of the nucleolus, and its downregulation induces defects in methylation of U1 and U2 snRNAs and alters splicing [112].
It is very likely that other molecular partners are waiting to be identified. Indeed, several functional analyses point to C/D snoRNAs, whereas the underlying mechanisms are not identified yet. As examples, cellular cholesterol homeostasis is affected by snord60 expression, while the analysis of rRNA modifications did not identify changes in methylation levels [229]. The expression and cytoplasmic localization of three snoRNAs hosted by the ribosomal Rpl13a gene is increased on cells exposed to fatty acids, and their knockdown protects against lipotoxic and oxidative stress [230]. If the mechanism used by these snoRNAs is unknown, it has been shown that their nuclear export involves the nuclear export factor NXF3 [231]. More largely, the export to the cytoplasm of snoRNAs might be a recurrent response to cell stress as it has also been observed in the Silkworm [232]. Even more surprisingly, it has been proposed that snoRNAs could be secreted through exosomes and absorbed by distant tissues where they could perform 2′-O-methylation of rRNA [233]. It is not known how this complex journey is achieved and regulated.
Despite this burst of data, numerous C/D snoRNAs have no identified role even if molecular associations have been identified. This is the case for several C/D snoRNAs that interact with splicing factors [234,235] or the spliceosome [236] in human cells or with the chromatin factor DF31 that localizes to euchromatic regions in Drosophila and tethers a heterogeneous pool of short RNAs that is enriched in snoRNAs [237]. This is also true for RNA partners, as the complex repertoire of snoRNA-C/D RNA interactions is beginning to be revealed by large-scale approaches [238]. In addition, future works are required to identify the functions of most orphan snoRNAs. This category concerns around 30–50% of C/D snoRNAs in Mammals, whereas orphan H/ACA snoRNAs are much rarer (for a review, see [239]). The leading example in this category is the repeated snord116 genes whose loss of expression is suspected of promoting the Prader Willi syndrome [240,241,242,243,244,245,246,247]. To note, the existence of orphan snoRNAs is not restricted to Mammals, e.g., if only a few snoRNAs are concerned in Saccharomyces cerevisiae, a quarter of snRNAs in the Archaea Pyrobaculum are orphans [248]. In a minority of cases, a molecular function has been found, such as eutherian snord115 that controls splicing and editing of a target mRNA [212,213,214] or the yeast snR4 and snR45 that guide rRNA acetylation [208]. The latter are also interesting as it shows that the role of orphan snoRNAs could rely to conventional target.
10. Conclusions
As it is true for other ncRNAs, the snoRNA category is currently subjected to numerous and exciting developments despite its early discovery. This is especially true for C/D snoRNAs that, in addition to be leading actors of rRNA and snRNA maturation, appear now as regulators of other cellular RNAs, including mRNAs. In parallel, it emerges that the biogenesis and function of C/D snoRNPs are controlled by regulatory systems that are poorly described. In the face of the bloom of C/D snoRNA fates and functions, the field is in need of molecular dissection as currently, too few mechanisms are in hands to understand this overt variety. Importantly enough, it has to be stressed that the great majority, not to say all, snoRNAs accumulate dramatically in the nucleolus at a steady-state (or in the CBs for scaRNAs), e.g., including orphan ones. Consequently, a rigorous analysis of C/D snoRNA functions should always question the existence of direct or indirect effects in these compartments. As a corollary, it is arguable that new snoRNA roles remain to be discovered in these compartments. Altogether, these important efforts are necessary to build a renewed understanding of C/D snoRNA functions.
Author Contributions
Conceptualization, B.C. and S.L.; literature investigation, writing, editing and review, L.B., B.C. and S.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from “La Ligue Contre le Cancer”. L.B. was a predoctoral fellow from the French “Ministère de l’Enseignement Supérieur et de la Recherche.”
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. Elegans Heterochronic Gene Lin-4 Encodes Small RNAs with Antisense Complementarity to Lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Ambros, V. MicroRNAs: Tiny Regulators with Great Potential. Cell 2001, 107, 823–826. [Google Scholar] [CrossRef] [Green Version]
- Weinberg, R.A.; Penman, S. Small Molecular Weight Monodisperse Nuclear RNA. J. Mol. Biol. 1968, 38, 289–304. [Google Scholar] [CrossRef]
- Nakamura, T.; Prestayko, A.W.; Busch, H. Studies on Nucleolar 4 to 6 S Ribonucleic Acid of Novikoff Hepatoma Cells. J. Biol. Chem. 1968, 243, 1368–1375. [Google Scholar] [CrossRef]
- Prestayko, A.W.; Tonato, M.; Busch, H. Low Molecular Weight RNA Associated with 28 s Nucleolar RNA. J. Mol. Biol. 1970, 47, 505–515. [Google Scholar] [CrossRef]
- Kass, S.; Tyc, K.; Steitz, J.A.; Sollner-Webb, B. The U3 Small Nucleolar Ribonucleoprotein Functions in the First Step of Preribosomal RNA Processing. Cell 1990, 60, 897–908. [Google Scholar] [CrossRef]
- Hughes, J.M. Functional Base-Pairing Interaction between Highly Conserved Elements of U3 Small Nucleolar RNA and the Small Ribosomal Subunit RNA. J. Mol. Biol. 1996, 259, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Tollervey, D. Base Pairing between U3 Small Nucleolar RNA and the 5′ End of 18S RRNA Is Required for Pre-RRNA Processing. Mol. Cell. Biol. 1999, 19, 6012–6019. [Google Scholar] [CrossRef] [Green Version]
- Dragon, F.; Gallagher, J.E.G.; Compagnone-Post, P.A.; Mitchell, B.M.; Porwancher, K.A.; Wehner, K.A.; Wormsley, S.; Settlage, R.E.; Shabanowitz, J.; Osheim, Y.; et al. A Large Nucleolar U3 Ribonucleoprotein Required for 18S Ribosomal RNA Biogenesis. Nature 2002, 417, 967–970. [Google Scholar] [CrossRef]
- Tyc, K.; Steitz, J.A. U3, U8 and U13 Comprise a New Class of Mammalian SnRNPs Localized in the Cell Nucleolus. EMBO J. 1989, 8, 3113–3119. [Google Scholar] [CrossRef]
- Kiss-László, Z.; Henry, Y.; Bachellerie, J.P.; Caizergues-Ferrer, M.; Kiss, T. Site-Specific Ribose Methylation of Preribosomal RNA: A Novel Function for Small Nucleolar RNAs. Cell 1996, 85, 1077–1088. [Google Scholar] [CrossRef] [Green Version]
- Balakin, A.G.; Smith, L.; Fournier, M.J. The RNA World of the Nucleolus: Two Major Families of Small RNAs Defined by Different Box Elements with Related Functions. Cell 1996, 86, 823–834. [Google Scholar] [CrossRef] [Green Version]
- Lafontaine, D.L.; Tollervey, D. Nop58p Is a Common Component of the Box C+D SnoRNPs That Is Required for SnoRNA Stability. RNA 1999, 5, 455–467. [Google Scholar] [CrossRef] [Green Version]
- Watkins, N.J.; Ségault, V.; Charpentier, B.; Nottrott, S.; Fabrizio, P.; Bachi, A.; Wilm, M.; Rosbash, M.; Branlant, C.; Lührmann, R. A Common Core RNP Structure Shared between the Small Nucleoar Box C/D RNPs and the Spliceosomal U4 SnRNP. Cell 2000, 103, 457–466. [Google Scholar] [CrossRef] [Green Version]
- Watkins, N.J.; Bohnsack, M.T. The Box C/D and H/ACA SnoRNPs: Key Players in the Modification, Processing and the Dynamic Folding of Ribosomal RNA. Wiley Interdiscip. Rev. RNA 2012, 3, 397–414. [Google Scholar] [CrossRef]
- Lafontaine, D.L.J. Noncoding RNAs in Eukaryotic Ribosome Biogenesis and Function. Nat. Struct. Mol. Biol. 2015, 22, 11–19. [Google Scholar] [CrossRef]
- Sloan, K.E.; Warda, A.S.; Sharma, S.; Entian, K.-D.; Lafontaine, D.L.J.; Bohnsack, M.T. Tuning the Ribosome: The Influence of RRNA Modification on Eukaryotic Ribosome Biogenesis and Function. RNA Biol. 2017, 14, 1138–1152. [Google Scholar] [CrossRef]
- Turowski, T.W.; Tollervey, D. Cotranscriptional Events in Eukaryotic Ribosome Synthesis. WIREs RNA 2015, 6, 129–139. [Google Scholar] [CrossRef]
- Henras, A.K.; Plisson-Chastang, C.; O’Donohue, M.-F.; Chakraborty, A.; Gleizes, P.-E. An Overview of Pre-Ribosomal RNA Processing in Eukaryotes. Wiley Interdiscip. Rev. RNA 2015, 6, 225–242. [Google Scholar] [CrossRef]
- Ganot, P.; Bortolin, M.L.; Kiss, T. Site-Specific Pseudouridine Formation in Preribosomal RNA Is Guided by Small Nucleolar RNAs. Cell 1997, 89, 799–809. [Google Scholar] [CrossRef] [Green Version]
- Ni, J.; Tien, A.L.; Fournier, M.J. Small Nucleolar RNAs Direct Site-Specific Synthesis of Pseudouridine in Ribosomal RNA. Cell 1997, 89, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Kiss, A.M.; Jády, B.E.; Bertrand, E.; Kiss, T. Human Box H/ACA Pseudouridylation Guide RNA Machinery. Mol. Cell. Biol. 2004, 24, 5797–5807. [Google Scholar] [CrossRef] [Green Version]
- Decatur, W.A.; Fournier, M.J. RNA-Guided Nucleotide Modification of Ribosomal and Other RNAs. J. Biol. Chem. 2003, 278, 695–698. [Google Scholar] [CrossRef] [Green Version]
- Morrissey, J.P.; Tollervey, D. Yeast SnR30 Is a Small Nucleolar RNA Required for 18S RRNA Synthesis. Mol. Cell. Biol. 1993, 13, 2469–2477. [Google Scholar] [CrossRef] [Green Version]
- Atzorn, V.; Fragapane, P.; Kiss, T. U17/SnR30 Is a Ubiquitous SnoRNA with Two Conserved Sequence Motifs Essential for 18S RRNA Production. Mol. Cell. Biol. 2004, 24, 1769–1778. [Google Scholar] [CrossRef] [Green Version]
- Meier, U.T. RNA Modification in Cajal Bodies. RNA Biol. 2017, 14, 693–700. [Google Scholar] [CrossRef]
- Bohnsack, M.T.; Sloan, K.E. Modifications in Small Nuclear RNAs and Their Roles in Spliceosome Assembly and Function. Biol. Chem. 2018, 399, 1265–1276. [Google Scholar] [CrossRef]
- Bratkovič, T.; Božič, J.; Rogelj, B. Functional Diversity of Small Nucleolar RNAs. Nucleic Acids Res. 2020, 48, 1627–1651. [Google Scholar] [CrossRef] [Green Version]
- Bergeron, D.; Fafard-Couture, É.; Scott, M.S. Small Nucleolar RNAs: Continuing Identification of Novel Members and Increasing Diversity of Their Molecular Mechanisms of Action. Biochem. Soc. Trans. 2020, 48, 645–656. [Google Scholar] [CrossRef] [Green Version]
- Kufel, J.; Grzechnik, P. Small Nucleolar RNAs Tell a Different Tale. Trends Genet. 2019, 35, 104–117. [Google Scholar] [CrossRef] [Green Version]
- Boivin, V.; Faucher-Giguère, L.; Scott, M.; Abou-Elela, S. The Cellular Landscape of Mid-Size Noncoding RNA. Wiley Interdiscip. Rev. RNA 2019, 10, e1530. [Google Scholar] [CrossRef] [Green Version]
- Dupuis-Sandoval, F.; Poirier, M.; Scott, M.S. The Emerging Landscape of Small Nucleolar RNAs in Cell Biology. Wiley Interdiscip. Rev. RNA 2015, 6, 381–397. [Google Scholar] [CrossRef]
- Massenet, S.; Bertrand, E.; Verheggen, C. Assembly and Trafficking of Box C/D and H/ACA SnoRNPs. RNA Biol. 2017, 14, 680–692. [Google Scholar] [CrossRef] [Green Version]
- Bachellerie, J.P.; Cavaillé, J.; Hüttenhofer, A. The Expanding SnoRNA World. Biochimie 2002, 84, 775–790. [Google Scholar] [CrossRef]
- Omer, A.D.; Lowe, T.M.; Russell, A.G.; Ebhardt, H.; Eddy, S.R.; Dennis, P.P. Homologs of Small Nucleolar RNAs in Archaea. Science 2000, 288, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Gaspin, C.; Cavaillé, J.; Erauso, G.; Bachellerie, J.P. Archaeal Homologs of Eukaryotic Methylation Guide Small Nucleolar RNAs: Lessons from the Pyrococcus Genomes. J. Mol. Biol. 2000, 297, 895–906. [Google Scholar] [CrossRef]
- Omer, A.D.; Ziesche, S.; Ebhardt, H.; Dennis, P.P. In Vitro Reconstitution and Activity of a C/D Box Methylation Guide Ribonucleoprotein Complex. Proc. Natl. Acad. Sci. USA 2002, 99, 5289–5294. [Google Scholar] [CrossRef] [Green Version]
- Rozhdestvensky, T.S.; Tang, T.H.; Tchirkova, I.V.; Brosius, J.; Bachellerie, J.-P.; Hüttenhofer, A. Binding of L7Ae Protein to the K-Turn of Archaeal SnoRNAs: A Shared RNA Binding Motif for C/D and H/ACA Box SnoRNAs in Archaea. Nucleic Acids Res. 2003, 31, 869–877. [Google Scholar] [CrossRef] [Green Version]
- Charpentier, B.; Muller, S.; Branlant, C. Reconstitution of Archaeal H/ACA Small Ribonucleoprotein Complexes Active in Pseudouridylation. Nucleic Acids Res. 2005, 33, 3133–3144. [Google Scholar] [CrossRef] [Green Version]
- Baker, D.L.; Youssef, O.A.; Chastkofsky, M.I.R.; Dy, D.A.; Terns, R.M.; Terns, M.P. RNA-Guided RNA Modification: Functional Organization of the Archaeal H/ACA RNP. Genes Dev. 2005, 19, 1238–1248. [Google Scholar] [CrossRef] [Green Version]
- Muller, S.; Leclerc, F.; Behm-Ansmant, I.; Fourmann, J.-B.; Charpentier, B.; Branlant, C. Combined in Silico and Experimental Identification of the Pyrococcus Abyssi H/ACA SRNAs and Their Target Sites in Ribosomal RNAs. Nucleic Acids Res. 2008, 36, 2459–2475. [Google Scholar] [CrossRef] [PubMed]
- Omer, A.D.; Ziesche, S.; Decatur, W.A.; Fournier, M.J.; Dennis, P.P. RNA-Modifying Machines in Archaea. Mol. Microbiol. 2003, 48, 617–629. [Google Scholar] [CrossRef]
- Dennis, P.P.; Omer, A. Small Non-Coding RNAs in Archaea. Curr. Opin. Microbiol. 2005, 8, 685–694. [Google Scholar] [CrossRef] [PubMed]
- van Nues, R.W.; Granneman, S.; Kudla, G.; Sloan, K.E.; Chicken, M.; Tollervey, D.; Watkins, N.J. Box C/D SnoRNP Catalysed Methylation Is Aided by Additional Pre-RRNA Base-Pairing. EMBO J. 2011, 30, 2420–2430. [Google Scholar] [CrossRef] [Green Version]
- Chagot, M.-E.; Quinternet, M.; Rothé, B.; Charpentier, B.; Coutant, J.; Manival, X.; Lebars, I. The Yeast C/D Box SnoRNA U14 Adopts a “Weak” K-Turn like Conformation Recognized by the Snu13 Core Protein in Solution. Biochimie 2019, 164, 70–82. [Google Scholar] [CrossRef]
- Klein, D.J.; Schmeing, T.M.; Moore, P.B.; Steitz, T.A. The Kink-Turn: A New RNA Secondary Structure Motif. EMBO J. 2001, 20, 4214–4221. [Google Scholar] [CrossRef]
- Moore, T.; Zhang, Y.; Fenley, M.O.; Li, H. Molecular Basis of Box C/D RNA-Protein Interactions; Cocrystal Structure of Archaeal L7Ae and a Box C/D RNA. Structure 2004, 12, 807–818. [Google Scholar] [CrossRef]
- Reichow, S.L.; Hamma, T.; Ferré-D’Amaré, A.R.; Varani, G. The Structure and Function of Small Nucleolar Ribonucleoproteins. Nucleic Acids Res. 2007, 35, 1452–1464. [Google Scholar] [CrossRef]
- Vidovic, I.; Nottrott, S.; Hartmuth, K.; Lührmann, R.; Ficner, R. Crystal Structure of the Spliceosomal 15.5 kD Protein Bound to a U4 SnRNA Fragment. Mol. Cell 2000, 6, 1331–1342. [Google Scholar] [CrossRef] [Green Version]
- Qu, G.; van Nues, R.W.; Watkins, N.J.; Maxwell, E.S. The Spatial-Functional Coupling of Box C/D and C’/D’ RNPs Is an Evolutionarily Conserved Feature of the Eukaryotic Box C/D SnoRNP Nucleotide Modification Complex. Mol. Cell. Biol. 2011, 31, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Hamma, T.; Ferré-D’Amaré, A.R. Structure of Protein L7Ae Bound to a K-Turn Derived from an Archaeal Box H/ACA SRNA at 1.8 A Resolution. Structure 2004, 12, 893–903. [Google Scholar] [CrossRef]
- Charron, C.; Manival, X.; Cléry, A.; Senty-Ségault, V.; Charpentier, B.; Marmier-Gourrier, N.; Branlant, C.; Aubry, A. The Archaeal SRNA Binding Protein L7Ae Has a 3D Structure Very Similar to That of Its Eukaryal Counterpart While Having a Broader RNA-Binding Specificity. J. Mol. Biol. 2004, 342, 757–773. [Google Scholar] [CrossRef] [PubMed]
- Nolivos, S.; Carpousis, A.J.; Clouet-d’Orval, B. The K-Loop, a General Feature of the Pyrococcus C/D Guide RNAs, Is an RNA Structural Motif Related to the K-Turn. Nucleic Acids Res. 2005, 33, 6507–6514. [Google Scholar] [CrossRef] [Green Version]
- Watkins, N.J.; Dickmanns, A.; Lührmann, R. Conserved Stem II of the Box C/D Motif Is Essential for Nucleolar Localization and Is Required, Along with the 15.5K Protein, for the Hierarchical Assembly of the Box C/D SnoRNP. Mol. Cell. Biol. 2002, 22, 8342–8352. [Google Scholar] [CrossRef] [Green Version]
- Szewczak, L.B.W.; Gabrielsen, J.S.; Degregorio, S.J.; Strobel, S.A.; Steitz, J.A. Molecular Basis for RNA Kink-Turn Recognition by the H15.5K Small RNP Protein. RNA 2005, 11, 1407–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cahill, N.M.; Friend, K.; Speckmann, W.; Li, Z.-H.; Terns, R.M.; Terns, M.P.; Steitz, J.A. Site-Specific Cross-Linking Analyses Reveal an Asymmetric Protein Distribution for a Box C/D SnoRNP. EMBO J. 2002, 21, 3816–3828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szewczak, L.B.W.; DeGregorio, S.J.; Strobel, S.A.; Steitz, J.A. Exclusive Interaction of the 15.5 KD Protein with the Terminal Box C/D Motif of a Methylation Guide SnoRNP. Chem. Biol. 2002, 9, 1095–1107. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Lai, S.; Jia, R.; Xu, A.; Zhang, L.; Lu, J.; Ye, K. Structural Basis for Site-Specific Ribose Methylation by Box C/D RNA Protein Complexes. Nature 2011, 469, 559–563. [Google Scholar] [CrossRef]
- Aittaleb, M.; Rashid, R.; Chen, Q.; Palmer, J.R.; Daniels, C.J.; Li, H. Structure and Function of Archaeal Box C/D SRNP Core Proteins. Nat. Struct. Mol. Biol. 2003, 10, 256–263. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, J.; Huang, L.; Lilley, D.M.J.; Ye, K. Functional Organization of Box C/D RNA-Guided RNA Methyltransferase. Nucleic Acids Res. 2020, 48, 5094–5105. [Google Scholar] [CrossRef]
- Barandun, J.; Chaker-Margot, M.; Hunziker, M.; Molloy, K.R.; Chait, B.T.; Klinge, S. The Complete Structure of the Small-Subunit Processome. Nat. Struct. Mol. Biol. 2017, 24, 944–953. [Google Scholar] [CrossRef]
- Clerget, G.; Bourguignon-Igel, V.; Marmier-Gourrier, N.; Rolland, N.; Wacheul, L.; Manival, X.; Charron, C.; Kufel, J.; Méreau, A.; Senty-Ségault, V.; et al. Synergistic Defects in Pre-RRNA Processing from Mutations in the U3-Specific Protein Rrp9 and U3 SnoRNA. Nucleic Acids Res. 2020, 48, 3848–3868. [Google Scholar] [CrossRef]
- Kiss-László, Z.; Henry, Y.; Kiss, T. Sequence and Structural Elements of Methylation Guide SnoRNAs Essential for Site-Specific Ribose Methylation of Pre-RRNA. EMBO J. 1998, 17, 797–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapinaite, A.; Simon, B.; Skjaerven, L.; Rakwalska-Bange, M.; Gabel, F.; Carlomagno, T. The Structure of the Box C/D Enzyme Reveals Regulation of RNA Methylation. Nature 2013, 502, 519–523. [Google Scholar] [CrossRef]
- Tran, E.; Zhang, X.; Lackey, L.; Maxwell, E.S. Conserved Spacing between the Box C/D and C′/D′ RNPs of the Archaeal Box C/D SRNP Complex Is Required for Efficient 2′-O-Methylation of Target RNAs. RNA 2005, 11, 285–293. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-L.; Perasso, R.; Qu, L.-H.; Amar, L. Exploration of Pairing Constraints Identifies a 9 Base-Pair Core within Box C/D SnoRNA-RRNA Duplexes. J. Mol. Biol. 2007, 369, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Lin, J.; Ye, K. Box C/D Guide RNAs Recognize a Maximum of 10 Nt of Substrates. Proc. Natl. Acad. Sci. USA 2016, 113, 10878–10883. [Google Scholar] [CrossRef] [Green Version]
- Graziadei, A.; Gabel, F.; Kirkpatrick, J.; Carlomagno, T. The Guide SRNA Sequence Determines the Activity Level of Box C/D RNPs. eLife 2020, 9. [Google Scholar] [CrossRef]
- Wu, S.; Wang, Y.; Wang, J.; Li, X.; Li, J.; Ye, K. Profiling of RNA Ribose Methylation in Arabidopsis thaliana. Nucleic Acids Res. 2021, 49. [Google Scholar] [CrossRef] [PubMed]
- Marmier-Gourrier, N.; Cléry, A.; Senty-Ségault, V.; Charpentier, B.; Schlotter, F.; Leclerc, F.; Fournier, R.; Branlant, C. A Structural, Phylogenetic, and Functional Study of 15.5-KD/Snu13 Protein Binding on U3 Small Nucleolar RNA. RNA 2003, 9, 821–838. [Google Scholar] [CrossRef] [Green Version]
- Rothé, B.; Back, R.; Quinternet, M.; Bizarro, J.; Robert, M.-C.; Blaud, M.; Romier, C.; Manival, X.; Charpentier, B.; Bertrand, E.; et al. Characterization of the Interaction between Protein Snu13p/15.5K and the Rsa1p/NUFIP Factor and Demonstration of Its Functional Importance for SnoRNP Assembly. Nucleic Acids Res. 2014, 42, 2015–2036. [Google Scholar] [CrossRef] [Green Version]
- Rothé, B.; Manival, X.; Rolland, N.; Charron, C.; Senty-Ségault, V.; Branlant, C.; Charpentier, B. Implication of the Box C/D SnoRNP Assembly Factor Rsa1p in U3 SnoRNP Assembly. Nucleic Acids Res. 2017, 45, 7455–7473. [Google Scholar] [CrossRef] [Green Version]
- Lübben, B.; Marshallsay, C.; Rottmann, N.; Lührmann, R. Isolation of U3 SnoRNP from CHO Cells: A Novel 55 KDa Protein Binds to the Central Part of U3 SnoRNA. Nucleic Acids Res. 1993, 21, 5377–5385. [Google Scholar] [CrossRef] [Green Version]
- Venema, J.; Vos, H.R.; Faber, A.W.; van Venrooij, W.J.; Raué, H.A. Yeast Rrp9p Is an Evolutionarily Conserved U3 SnoRNP Protein Essential for Early Pre-RRNA Processing Cleavages and Requires Box C for Its Association. RNA 2000, 6, 1660–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluk, H.; Soffner, J.; Lührmann, R.; van Venrooij, W.J. CDNA Cloning and Characterization of the Human U3 Small Nucleolar Ribonucleoprotein Complex-Associated 55-Kilodalton Protein. Mol. Cell. Biol. 1998, 18, 488–498. [Google Scholar] [CrossRef] [Green Version]
- Lukowiak, A.A.; Granneman, S.; Mattox, S.A.; Speckmann, W.A.; Jones, K.; Pluk, H.; Venrooij, W.J.; Terns, R.M.; Terns, M.P. Interaction of the U3-55k Protein with U3 SnoRNA Is Mediated by the Box B/C Motif of U3 and the WD Repeats of U3-55 k. Nucleic Acids Res. 2000, 28, 3462–3471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cléry, A.; Senty-Ségault, V.; Leclerc, F.; Raué, H.A.; Branlant, C. Analysis of Sequence and Structural Features That Identify the B/C Motif of U3 Small Nucleolar RNA as the Recognition Site for the Snu13p-Rrp9p Protein Pair. Mol. Cell. Biol. 2007, 27, 1191–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granneman, S.; Pruijn, G.J.M.; Horstman, W.; van Venrooij, W.J.; Luhrmann, R.; Watkins, N.J. The HU3-55K Protein Requires 15.5 K Binding to the Box B/C Motif as Well as Flanking RNA Elements for Its Association with the U3 Small Nucleolar RNA in Vitro. J. Biol. Chem. 2002, 277, 48490–48500. [Google Scholar] [CrossRef] [Green Version]
- Granneman, S.; Kudla, G.; Petfalski, E.; Tollervey, D. Identification of Protein Binding Sites on U3 SnoRNA and Pre-RRNA by UV Cross-Linking and High-Throughput Analysis of CDNAs. Proc. Natl. Acad. Sci. USA 2009, 106, 9613–9618. [Google Scholar] [CrossRef] [Green Version]
- Knox, A.A.; McKeegan, K.S.; Debieux, C.M.; Traynor, A.; Richardson, H.; Watkins, N.J. A Weak C’ Box Renders U3 SnoRNA Levels Dependent on HU3-55K Binding. Mol. Cell. Biol. 2011, 31, 2404–2412. [Google Scholar] [CrossRef] [Green Version]
- Watkins, N.J.; Lemm, I.; Ingelfinger, D.; Schneider, C.; Hossbach, M.; Urlaub, H.; Lührmann, R. Assembly and Maturation of the U3 SnoRNP in the Nucleoplasm in a Large Dynamic Multiprotein Complex. Mol. Cell 2004, 16, 789–798. [Google Scholar] [CrossRef]
- Chaker-Margot, M.; Barandun, J.; Hunziker, M.; Klinge, S. Architecture of the Yeast Small Subunit Processome. Science 2017, 355. [Google Scholar] [CrossRef] [PubMed]
- Kornprobst, M.; Turk, M.; Kellner, N.; Cheng, J.; Flemming, D.; Koš-Braun, I.; Koš, M.; Thoms, M.; Berninghausen, O.; Beckmann, R.; et al. Architecture of the 90S Pre-Ribosome: A Structural View on the Birth of the Eukaryotic Ribosome. Cell 2016, 166, 380–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Q.; Zhu, X.; Qi, J.; An, W.; Lan, P.; Tan, D.; Chen, R.; Wang, B.; Zheng, S.; Zhang, C.; et al. Molecular Architecture of the 90S Small Subunit Pre-Ribosome. eLife 2017, 6, e22086. [Google Scholar] [CrossRef] [PubMed]
- Méreau, A.; Fournier, R.; Grégoire, A.; Mougin, A.; Fabrizio, P.; Lührmann, R.; Branlant, C. An in Vivo and in Vitro Structure-Function Analysis of the Saccharomyces Cerevisiae U3A SnoRNP: Protein-RNA Contacts and Base-Pair Interaction with the Pre-Ribosomal RNA. J. Mol. Biol. 1997, 273, 552–571. [Google Scholar] [CrossRef]
- Borovjagin, A.V.; Gerbi, S.A. An Evolutionary Intra-Molecular Shift in the Preferred U3 SnoRNA Binding Site on Pre-Ribosomal RNA. Nucleic Acids Res. 2005, 33, 4995–5005. [Google Scholar] [CrossRef] [Green Version]
- Marmier-Gourrier, N.; Cléry, A.; Schlotter, F.; Senty-Ségault, V.; Branlant, C. A Second Base Pair Interaction between U3 Small Nucleolar RNA and the 5’-ETS Region Is Required for Early Cleavage of the Yeast Pre-Ribosomal RNA. Nucleic Acids Res. 2011, 39, 9731–9745. [Google Scholar] [CrossRef]
- Dutca, L.M.; Gallagher, J.E.G.; Baserga, S.J. The Initial U3 SnoRNA:Pre-RRNA Base Pairing Interaction Required for Pre-18S RRNA Folding Revealed by in Vivo Chemical Probing. Nucleic Acids Res. 2011, 39, 5164–5180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltrame, M.; Tollervey, D. Base Pairing between U3 and the Pre-Ribosomal RNA Is Required for 18S RRNA Synthesis. EMBO J. 1995, 14, 4350–4356. [Google Scholar] [CrossRef]
- Paul, A.; Tiotiu, D.; Bragantini, B.; Marty, H.; Charpentier, B.; Massenet, S.; Labialle, S. Bcd1p Controls RNA Loading of the Core Protein Nop58 during C/D Box SnoRNP Biogenesis. RNA 2019, 25, 496–506. [Google Scholar] [CrossRef]
- Hirose, T.; Steitz, J.A. Position within the Host Intron Is Critical for Efficient Processing of Box C/D SnoRNAs in Mammalian Cells. Proc. Natl. Acad. Sci. USA 2001, 98, 12914–12919. [Google Scholar] [CrossRef] [Green Version]
- Vincenti, S.; De Chiara, V.; Bozzoni, I.; Presutti, C. The Position of Yeast SnoRNA-Coding Regions within Host Introns Is Essential for Their Biosynthesis and for Efficient Splicing of the Host Pre-MRNA. RNA 2007, 13, 138–150. [Google Scholar] [CrossRef] [Green Version]
- Hirose, T.; Shu, M.-D.; Steitz, J.A. Splicing-Dependent and -Independent Modes of Assembly for Intron-Encoded Box C/D SnoRNPs in Mammalian Cells. Mol. Cell 2003, 12, 113–123. [Google Scholar] [CrossRef]
- Richard, P.; Kiss, T. Integrating SnoRNP Assembly with MRNA Biogenesis. EMBO Rep. 2006, 7, 590–592. [Google Scholar] [CrossRef] [Green Version]
- De, I.; Bessonov, S.; Hofele, R.; dos Santos, K.; Will, C.L.; Urlaub, H.; Lührmann, R.; Pena, V. The RNA Helicase Aquarius Exhibits Structural Adaptations Mediating Its Recruitment to Spliceosomes. Nat. Struct. Mol. Biol. 2015, 22, 138–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirose, T.; Ideue, T.; Nagai, M.; Hagiwara, M.; Shu, M.-D.; Steitz, J.A. A Spliceosomal Intron Binding Protein, IBP160, Links Position-Dependent Assembly of Intron-Encoded Box C/D SnoRNP to Pre-MRNA Splicing. Mol. Cell 2006, 23, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Morlando, M.; Ballarino, M.; Greco, P.; Caffarelli, E.; Dichtl, B.; Bozzoni, I. Coupling between SnoRNP Assembly and 3′ Processing Controls Box C/D SnoRNA Biosynthesis in Yeast. EMBO J. 2004, 23, 2392–2401. [Google Scholar] [CrossRef] [Green Version]
- Rothé, B.; Saliou, J.-M.; Quinternet, M.; Back, R.; Tiotiu, D.; Jacquemin, C.; Loegler, C.; Schlotter, F.; Peña, V.; Eckert, K.; et al. Protein Hit1, a Novel Box C/D SnoRNP Assembly Factor, Controls Cellular Concentration of the Scaffolding Protein Rsa1 by Direct Interaction. Nucleic Acids Res. 2014, 42, 10731–10747. [Google Scholar] [CrossRef] [PubMed]
- Bizarro, J.; Charron, C.; Boulon, S.; Westman, B.; Pradet-Balade, B.; Vandermoere, F.; Chagot, M.-E.; Hallais, M.; Ahmad, Y.; Leonhardt, H.; et al. Proteomic and 3D Structure Analyses Highlight the C/D Box SnoRNP Assembly Mechanism and Its Control. J. Cell Biol. 2014, 207, 463–480. [Google Scholar] [CrossRef] [PubMed]
- Boulon, S.; Marmier-Gourrier, N.; Pradet-Balade, B.; Wurth, L.; Verheggen, C.; Jády, B.E.; Rothé, B.; Pescia, C.; Robert, M.-C.; Kiss, T.; et al. The Hsp90 Chaperone Controls the Biogenesis of L7Ae RNPs through Conserved Machinery. J. Cell Biol. 2008, 180, 579–595. [Google Scholar] [CrossRef] [PubMed]
- McKeegan, K.S.; Debieux, C.M.; Boulon, S.; Bertrand, E.; Watkins, N.J. A Dynamic Scaffold of Pre-SnoRNP Factors Facilitates Human Box C/D SnoRNP Assembly. Mol. Cell. Biol. 2007, 27, 6782–6793. [Google Scholar] [CrossRef] [Green Version]
- McKeegan, K.S.; Debieux, C.M.; Watkins, N.J. Evidence That the AAA+ Proteins TIP48 and TIP49 Bridge Interactions between 15.5K and the Related NOP56 and NOP58 Proteins during Box C/D SnoRNP Biogenesis. Mol. Cell. Biol. 2009, 29, 4971–4981. [Google Scholar] [CrossRef] [Green Version]
- Bragantini, B.; Tiotiu, D.; Rothé, B.; Saliou, J.-M.; Marty, H.; Cianférani, S.; Charpentier, B.; Quinternet, M.; Manival, X. Functional and Structural Insights of the Zinc-Finger HIT Protein Family Members Involved in Box C/D SnoRNP Biogenesis. J. Mol. Biol. 2016, 428, 2488–2506. [Google Scholar] [CrossRef] [PubMed]
- Khoshnevis, S.; Dreggors, R.E.; Hoffmann, T.F.R.; Ghalei, H. A Conserved Bcd1 Interaction Essential for Box C/D SnoRNP Biogenesis. J. Biol. Chem. 2019, 294, 18360–18371. [Google Scholar] [CrossRef]
- Kiss, T.; Fayet, E.; Jády, B.E.; Richard, P.; Weber, M. Biogenesis and Intranuclear Trafficking of Human Box C/D and H/ACA RNPs. Cold Spring Harb. Symp. Quant. Biol. 2006, 71, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Quinternet, M.; Rothé, B.; Barbier, M.; Bobo, C.; Saliou, J.-M.; Jacquemin, C.; Back, R.; Chagot, M.-E.; Cianférani, S.; Meyer, P.; et al. Structure/Function Analysis of Protein-Protein Interactions Developed by the Yeast Pih1 Platform Protein and Its Partners in Box C/D SnoRNP Assembly. J. Mol. Biol. 2015, 427, 2816–2839. [Google Scholar] [CrossRef] [PubMed]
- Quinternet, M.; Chagot, M.-E.; Rothé, B.; Tiotiu, D.; Charpentier, B.; Manival, X. Structural Features of the Box C/D SnoRNP Pre-Assembly Process Are Conserved through Species. Structure 2016, 24, 1693–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henri, J.; Chagot, M.-E.; Bourguet, M.; Abel, Y.; Terral, G.; Maurizy, C.; Aigueperse, C.; Georgescauld, F.; Vandermoere, F.; Saint-Fort, R.; et al. Deep Structural Analysis of RPAP3 and PIH1D1, Two Components of the HSP90 Co-Chaperone R2TP Complex. Structure 2018, 26, 1196–1209.e8. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Kakihara, Y.; Gribun, A.; Huen, J.; Yang, G.; Khanna, M.; Costanzo, M.; Brost, R.L.; Boone, C.; Hughes, T.R.; et al. Molecular Chaperone Hsp90 Stabilizes Pih1/Nop17 to Maintain R2TP Complex Activity That Regulates SnoRNA Accumulation. J. Cell Biol. 2008, 180, 563–578. [Google Scholar] [CrossRef] [PubMed]
- Tycowski, K.T.; Shu, M.-D.; Kukoyi, A.; Steitz, J.A. A Conserved WD40 Protein Binds the Cajal Body Localization Signal of ScaRNP Particles. Mol. Cell 2009, 34, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marnef, A.; Richard, P.; Pinzón, N.; Kiss, T. Targeting Vertebrate Intron-Encoded Box C/D 2′-O-Methylation Guide RNAs into the Cajal Body. Nucleic Acids Res. 2014, 42, 6616–6629. [Google Scholar] [CrossRef] [Green Version]
- Izumikawa, K.; Nobe, Y.; Ishikawa, H.; Yamauchi, Y.; Taoka, M.; Sato, K.; Nakayama, H.; Simpson, R.J.; Isobe, T.; Takahashi, N. TDP-43 Regulates Site-Specific 2′-O-Methylation of U1 and U2 SnRNAs via Controlling the Cajal Body Localization of a Subset of C/D ScaRNAs. Nucleic Acids Res. 2019, 47, 2487–2505. [Google Scholar] [CrossRef] [Green Version]
- Boulon, S.; Verheggen, C.; Jady, B.E.; Girard, C.; Pescia, C.; Paul, C.; Ospina, J.K.; Kiss, T.; Matera, A.G.; Bordonné, R.; et al. PHAX and CRM1 Are Required Sequentially to Transport U3 SnoRNA to Nucleoli. Mol. Cell 2004, 16, 777–787. [Google Scholar] [CrossRef]
- Machyna, M.; Kehr, S.; Straube, K.; Kappei, D.; Buchholz, F.; Butter, F.; Ule, J.; Hertel, J.; Stadler, P.F.; Neugebauer, K.M. The Coilin Interactome Identifies Hundreds of Small Noncoding RNAs That Traffic through Cajal Bodies. Mol. Cell 2014, 56, 389–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pradet-Balade, B.; Girard, C.; Boulon, S.; Paul, C.; Azzag, K.; Bordonné, R.; Bertrand, E.; Verheggen, C. CRM1 Controls the Composition of Nucleoplasmic Pre-SnoRNA Complexes to Licence Them for Nucleolar Transport. EMBO J. 2011, 30, 2205–2218. [Google Scholar] [CrossRef] [Green Version]
- Verheggen, C.; Mouaikel, J.; Thiry, M.; Blanchard, J.M.; Tollervey, D.; Bordonné, R.; Lafontaine, D.L.; Bertrand, E. Box C/D Small Nucleolar RNA Trafficking Involves Small Nucleolar RNP Proteins, Nucleolar Factors and a Novel Nuclear Domain. EMBO J. 2001, 20, 5480–5490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verheggen, C.; Bertrand, E. CRM1 Plays a Nuclear Role in Transporting SnoRNPs to Nucleoli in Higher Eukaryotes. Nucleus 2012, 3, 132–137. [Google Scholar] [CrossRef] [Green Version]
- Bortolin-Cavaillé, M.-L.; Cavaillé, J. The SNORD115 (H/MBII-52) and SNORD116 (H/MBII-85) Gene Clusters at the Imprinted Prader-Willi Locus Generate Canonical Box C/D SnoRNAs. Nucleic Acids Res. 2012, 40, 6800–6807. [Google Scholar] [CrossRef] [PubMed]
- Vitali, P.; Royo, H.; Marty, V.; Bortolin-Cavaillé, M.-L.; Cavaillé, J. Long Nuclear-Retained Non-Coding RNAs and Allele-Specific Higher-Order Chromatin Organization at Imprinted SnoRNA Gene Arrays. J. Cell Sci. 2010, 123 Pt 1, 70–83. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-O.; Yin, Q.-F.; Wang, H.-B.; Zhang, Y.; Chen, T.; Zheng, P.; Lu, X.; Chen, L.-L.; Yang, L. Species-Specific Alternative Splicing Leads to Unique Expression of Sno-LncRNAs. BMC Genomics 2014, 15, 287. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Yin, Q.-F.; Luo, Z.; Yao, R.-W.; Zheng, C.-C.; Zhang, J.; Xiang, J.-F.; Yang, L.; Chen, L.-L. Unusual Processing Generates SPA LncRNAs That Sequester Multiple RNA Binding Proteins. Mol. Cell 2016, 64, 534–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Zhang, J.; Wang, M.; Li, X.; Gong, H.; Tang, H.; Chen, L.; Wan, L.; Liu, Q. Activity Dependent LoNA Regulates Translation by Coordinating RRNA Transcription and Methylation. Nat. Commun. 2018, 9, 1726. [Google Scholar] [CrossRef] [PubMed]
- Lykke-Andersen, S.; Ardal, B.K.; Hollensen, A.K.; Damgaard, C.K.; Jensen, T.H. Box C/D SnoRNP Autoregulation by a Cis-Acting SnoRNA in the NOP56 Pre-MRNA. Mol. Cell 2018, 72, 99–111.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deschamps-Francoeur, G.; Garneau, D.; Dupuis-Sandoval, F.; Roy, A.; Frappier, M.; Catala, M.; Couture, S.; Barbe-Marcoux, M.; Abou-Elela, S.; Scott, M.S. Identification of Discrete Classes of Small Nucleolar RNA Featuring Different Ends and RNA Binding Protein Dependency. Nucleic Acids Res. 2014, 42, 10073–10085. [Google Scholar] [CrossRef]
- Zywicki, M.; Bakowska-Zywicka, K.; Polacek, N. Revealing Stable Processing Products from Ribosome-Associated Small RNAs by Deep-Sequencing Data Analysis. Nucleic Acids Res. 2012, 40, 4013–4024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraiya, A.A.; Wang, C.C. SnoRNA, a Novel Precursor of MicroRNA in Giardia Lamblia. PLoS Pathog. 2008, 4, e10000224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, H.; Sun, Q.; Yao, Y. Characterization of Small RNAs Derived from TRNAs, RRNAs and SnoRNAs and Their Response to Heat Stress in Wheat Seedlings. PLoS ONE 2016, 11, e0150933. [Google Scholar] [CrossRef] [Green Version]
- Kishore, S.; Khanna, A.; Zhang, Z.; Hui, J.; Balwierz, P.J.; Stefan, M.; Beach, C.; Nicholls, R.D.; Zavolan, M.; Stamm, S. The SnoRNA MBII-52 (SNORD 115) Is Processed into Smaller RNAs and Regulates Alternative Splicing. Hum. Mol. Genet. 2010, 19, 1153–1164. [Google Scholar] [CrossRef] [Green Version]
- Ender, C.; Krek, A.; Friedländer, M.R.; Beitzinger, M.; Weinmann, L.; Chen, W.; Pfeffer, S.; Rajewsky, N.; Meister, G. A Human SnoRNA with MicroRNA-Like Functions. Mol. Cell 2008, 32, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Hutzinger, R.; Feederle, R.; Mrazek, J.; Schiefermeier, N.; Balwierz, P.J.; Zavolan, M.; Polacek, N.; Delecluse, H.-J.; Hüttenhofer, A. Expression and Processing of a Small Nucleolar RNA from the Epstein-Barr Virus Genome. PLoS Pathog. 2009, 5, e10000547. [Google Scholar] [CrossRef] [Green Version]
- Poole, A.R.; Vicino, I.; Adachi, H.; Yu, Y.-T.; Hebert, M.D. Regulatory RNPs: A Novel Class of Ribonucleoproteins That Potentially Contribute to Ribosome Heterogeneity. Biol. Open 2017, 6, 1342–1354. [Google Scholar] [CrossRef] [Green Version]
- Chow, R.D.; Chen, S. Sno-Derived RNAs Are Prevalent Molecular Markers of Cancer Immunity. Oncogene 2018, 37, 6442–6462. [Google Scholar] [CrossRef] [PubMed]
- Martens-Uzunova, E.S.; Hoogstrate, Y.; Kalsbeek, A.; Pigmans, B.; Vredenbregt-van den Berg, M.; Dits, N.; Nielsen, S.J.; Baker, A.; Visakorpi, T.; Bangma, C.; et al. C/D-Box SnoRNA-Derived RNA Production Is Associated with Malignant Transformation and Metastatic Progression in Prostate Cancer. Oncotarget 2015, 6, 17430–17444. [Google Scholar] [CrossRef] [Green Version]
- Mleczko, A.M.; Machtel, P.; Walkowiak, M.; Wasilewska, A.; Pietras, P.J.; Bąkowska-Żywicka, K. Levels of SdRNAs in Cytoplasm and Their Association with Ribosomes Are Dependent upon Stress Conditions but Independent from SnoRNA Expression. Sci. Rep. 2019, 9, 18397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, B.; Yegnasubramanian, S.; Wheelan, S.J.; Laiho, M. RNA-Seq of the Nucleolus Reveals Abundant SNORD44-Derived Small RNAs. PLoS ONE 2014, 9, e107519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taft, R.J.; Glazov, E.A.; Lassmann, T.; Hayashizaki, Y.; Carninci, P.; Mattick, J.S. Small RNAs Derived from SnoRNAs. RNA 2009, 15, 1233–1240. [Google Scholar] [CrossRef] [Green Version]
- Guillen-Chable, F.; Corona, U.R.; Pereira-Santana, A.; Bayona, A.; Rodríguez-Zapata, L.C.; Aquino, C.; Šebestová, L.; Vitale, N.; Hozak, P.; Castano, E. Fibrillarin Ribonuclease Activity Is Dependent on the GAR Domain and Modulated by Phospholipids. Cells 2020, 9, 1143. [Google Scholar] [CrossRef]
- Lemus-Diaz, N.; Ferreira, R.R.; Bohnsack, K.E.; Gruber, J.; Bohnsack, M.T. The Human Box C/D SnoRNA U3 Is a MiRNA Source and MiR-U3 Regulates Expression of Sortin Nexin 27. Nucleic Acids Res. 2020, 48, 8074–8089. [Google Scholar] [CrossRef]
- Zhong, F.; Zhou, N.; Wu, K.; Guo, Y.; Tan, W.; Zhang, H.; Zhang, X.; Geng, G.; Pan, T.; Luo, H.; et al. A SnoRNA-Derived PiRNA Interacts with Human Interleukin-4 Pre-MRNA and Induces Its Decay in Nuclear Exosomes. Nucleic Acids Res. 2015, 43, 10474–10491. [Google Scholar] [CrossRef] [Green Version]
- Langenberger, D.; Çakir, M.V.; Hoffmann, S.; Stadler, P.F. Dicer-Processed Small RNAs: Rules and Exceptions. J. Exp. Zool. B Mol. Dev. Evol. 2013, 320, 35–46. [Google Scholar] [CrossRef]
- Logan, M.K.; Burke, M.F.; Hebert, M.D. Altered Dynamics of ScaRNA2 and ScaRNA9 in Response to Stress Correlates with Disrupted Nuclear Organization. Biol. Open 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- McLaurin, D.M.; Logan, M.K.; Lett, K.E.; Hebert, M.D. Molecular Determinants That Govern ScaRNA Processing by Drosha/DGCR8. Biol. Open 2020, 9. [Google Scholar] [CrossRef]
- Kishore, S.; Gruber, A.R.; Jedlinski, D.J.; Syed, A.P.; Jorjani, H.; Zavolan, M. Insights into SnoRNA Biogenesis and Processing from PAR-CLIP of SnoRNA Core Proteins and Small RNA Sequencing. Genome Biol. 2013, 14, R45. [Google Scholar] [CrossRef] [Green Version]
- Lykke-Andersen, S.; Chen, Y.; Ardal, B.R.; Lilje, B.; Waage, J.; Sandelin, A.; Jensen, T.H. Human Nonsense-Mediated RNA Decay Initiates Widely by Endonucleolysis and Targets SnoRNA Host Genes. Genes Dev. 2014, 28, 2498–2517. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Ashraf, S.; Wang, J.; Lilley, D.M. Control of Box C/D SnoRNP Assembly by N6-Methylation of Adenine. EMBO Rep. 2017, 18, 1631–1645. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, S.; Huang, L.; Lilley, D.M.J. Effect of Methylation of Adenine N6 on Kink Turn Structure Depends on Location. RNA Biol. 2019, 16, 1377–1385. [Google Scholar] [CrossRef] [Green Version]
- Westman, B.J.; Verheggen, C.; Hutten, S.; Lam, Y.W.; Bertrand, E.; Lamond, A.I. A Proteomic Screen for Nucleolar SUMO Targets Shows SUMOylation Modulates the Function of Nop5/Nop58. Mol. Cell 2010, 39, 618–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westman, B.J.; Lamond, A.I. A Role for SUMOylation in SnoRNP Biogenesis Revealed by Quantitative Proteomics. Nucleus 2011, 2, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Qin, W.; Lv, P.; Fan, X.; Quan, B.; Zhu, Y.; Qin, K.; Chen, Y.; Wang, C.; Chen, X. Quantitative Time-Resolved Chemoproteomics Reveals That Stable O-GlcNAc Regulates Box C/D SnoRNP Biogenesis. Proc. Natl. Acad. Sci. USA 2017, 114, E6749–E6758. [Google Scholar] [CrossRef] [Green Version]
- Kakihara, Y.; Makhnevych, T.; Zhao, L.; Tang, W.; Houry, W.A. Nutritional Status Modulates Box C/D SnoRNP Biogenesis by Regulated Subcellular Relocalization of the R2TP Complex. Genome Biol. 2014, 15, 404. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Wu, Z.J.; Groner, A.C.; He, H.H.; Cai, C.; Lis, R.T.; Wu, X.; Stack, E.C.; Loda, M.; Liu, T.; et al. EZH2 Oncogenic Activity in Castration-Resistant Prostate Cancer Cells Is Polycomb-Independent. Science 2012, 338, 1465–1469. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.; Li, Y.; Meng, Q.; Li, Q.; Li, F.; Lu, B.; Shen, J.; Fazli, L.; Zhao, D.; Li, C.; et al. A PRC2-Independent Function for EZH2 in Regulating RRNA 2’-O Methylation and IRES-Dependent Translation. Nat. Cell Biol. 2021, 23. [Google Scholar] [CrossRef]
- Nachmani, D.; Bothmer, A.H.; Grisendi, S.; Mele, A.; Bothmer, D.; Lee, J.D.; Monteleone, E.; Cheng, K.; Zhang, Y.; Bester, A.C.; et al. Germline NPM1 Mutations Lead to Altered RRNA 2′-O-Methylation and Cause Dyskeratosis Congenita. Nat. Genet. 2019, 51, 1518–1529. [Google Scholar] [CrossRef]
- Bragantini, B.; Charron, C.; Bourguet, M.; Paul, A.; Tiotiu, D.; Rothé, B.; Marty, H.; Terral, G.; Hessmann, S.; Decourty, L.; et al. The Box C/D SnoRNP Assembly Factor Bcd1 Interacts with the Histone Chaperone Rtt106 and Controls Its Transcription Dependent Activity. Nat. Commun. 2021, 12, 1859. [Google Scholar] [CrossRef]
- de los Santos-Velázquez, A.I.; de Oya, I.G.; Manzano-López, J.; Monje-Casas, F. Late RDNA Condensation Ensures Timely Cdc14 Release and Coordination of Mitotic Exit Signaling with Nucleolar Segregation. Curr. Biol. 2017, 27, 3248–3263.e5. [Google Scholar] [CrossRef] [PubMed]
- Doyon, Y.; Selleck, W.; Lane, W.S.; Tan, S.; Côté, J. Structural and Functional Conservation of the NuA4 Histone Acetyltransferase Complex from Yeast to Humans. Mol. Cell. Biol. 2004, 24, 1884–1896. [Google Scholar] [CrossRef] [Green Version]
- Kobor, M.S.; Venkatasubrahmanyam, S.; Meneghini, M.D.; Gin, J.W.; Jennings, J.L.; Link, A.J.; Madhani, H.D.; Rine, J. A Protein Complex Containing the Conserved Swi2/Snf2-Related ATPase Swr1p Deposits Histone Variant H2A.Z into Euchromatin. PLoS Biol. 2004, 2, E131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Cai, Y.; Jin, J.; Florens, L.; Swanson, S.K.; Washburn, M.P.; Conaway, J.W.; Conaway, R.C. Subunit Organization of the Human INO80 Chromatin Remodeling Complex: An Evolutionarily Conserved Core Complex Catalyzes ATP-Dependent Nucleosome Remodeling. J. Biol. Chem. 2011, 286, 11283–11289. [Google Scholar] [CrossRef] [Green Version]
- Houry, W.A.; Bertrand, E.; Coulombe, B. The PAQosome, an R2TP-Based Chaperone for Quaternary Structure Formation. Trends Biochem. Sci. 2018, 43, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Kressler, D.; Doère, M.; Rojo, M.; Linder, P. Synthetic Lethality with Conditional Dbp6 Alleles Identifies Rsa1p, a Nucleoplasmic Protein Involved in the Assembly of 60S Ribosomal Subunits. Mol. Cell. Biol. 1999, 19, 8633–8645. [Google Scholar] [CrossRef] [Green Version]
- Soeno, Y.; Taya, Y.; Stasyk, T.; Huber, L.A.; Aoba, T.; Hüttenhofer, A. Identification of Novel Ribonucleo-Protein Complexes from the Brain-Specific SnoRNA MBII-52. RNA 2010, 16, 1293–1300. [Google Scholar] [CrossRef] [Green Version]
- Falaleeva, M.; Pages, A.; Matuszek, Z.; Hidmi, S.; Agranat-Tamir, L.; Korotkov, K.; Nevo, Y.; Eyras, E.; Sperling, R.; Stamm, S. Dual Function of C/D Box Small Nucleolar RNAs in RRNA Modification and Alternative Pre-MRNA Splicing. Proc. Natl. Acad. Sci. USA 2016, 113, E1625–E1634. [Google Scholar] [CrossRef] [Green Version]
- Diao, L.-T.; Xiao, Z.-D.; Leng, X.-M.; Li, B.; Li, J.-H.; Luo, Y.-P.; Li, S.-G.; Yu, C.-H.; Zhou, H.; Qu, L.-H. Conservation and Divergence of Transcriptional Coregulations between Box C/D SnoRNA and Ribosomal Protein Genes in Ascomycota. RNA 2014, 20, 1376–1385. [Google Scholar] [CrossRef] [Green Version]
- Nabavi, S.; Nazar, R.N. U3 SnoRNA Promoter Reflects the RNA’s Function in Ribosome Biogenesis. Curr. Genet. 2008, 54, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Herter, E.K.; Stauch, M.; Gallant, M.; Wolf, E.; Raabe, T.; Gallant, P. SnoRNAs Are a Novel Class of Biologically Relevant Myc Targets. BMC Biol. 2015, 13, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walz, S.; Lorenzin, F.; Morton, J.; Wiese, K.E.; von Eyss, B.; Herold, S.; Rycak, L.; Dumay-Odelot, H.; Karim, S.; Bartkuhn, M.; et al. Activation and Repression by Oncogenic MYC Shape Tumour-Specific Gene Expression Profiles. Nature 2014, 511, 483–487. [Google Scholar] [CrossRef]
- Dieci, G.; Preti, M.; Montanini, B. Eukaryotic SnoRNAs: A Paradigm for Gene Expression Flexibility. Genomics 2009, 94, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Pelczar, P.; Filipowicz, W. The Host Gene for Intronic U17 Small Nucleolar RNAs in Mammals Has No Protein-Coding Potential and Is a Member of the 5′-Terminal Oligopyrimidine Gene Family. Mol. Cell. Biol. 1998, 18, 4509–4518. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.M.; Steitz, J.A. Classification of Gas5 as a Multi-Small-Nucleolar-RNA (SnoRNA) Host Gene and a Member of the 5’-Terminal Oligopyrimidine Gene Family Reveals Common Features of SnoRNA Host Genes. Mol. Cell. Biol. 1998, 18, 6897–6909. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, R.; Suzuki, Y.; Takeuchi, N.; Wakaguri, H.; Ueda, T.; Sugano, S.; Nakai, K. Comprehensive Detection of Human Terminal Oligo-Pyrimidine (TOP) Genes and Analysis of Their Characteristics. Nucleic Acids Res. 2008, 36, 3707–3715. [Google Scholar] [CrossRef]
- Jalal, C.; Uhlmann-Schiffler, H.; Stahl, H. Redundant Role of DEAD Box Proteins P68 (Ddx5) and P72/P82 (Ddx17) in Ribosome Biogenesis and Cell Proliferation. Nucleic Acids Res. 2007, 35, 3590–3601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismael, H.; Altmeyer, S.; Stahl, H. Regulation of the U3-, U8-, and U13snoRNA Expression by the DEAD Box Proteins Ddx5/Ddx17 with Consequences for Cell Proliferation and Survival. Noncoding RNA 2016, 2, 11. [Google Scholar] [CrossRef]
- Gong, J.; Li, Y.; Liu, C.; Xiang, Y.; Li, C.; Ye, Y.; Zhang, Z.; Hawke, D.H.; Park, P.K.; Diao, L.; et al. A Pan-Cancer Analysis of the Expression and Clinical Relevance of Small Nucleolar RNAs in Human Cancer. Cell Rep. 2017, 21, 1968–1981. [Google Scholar] [CrossRef] [Green Version]
- Valleron, W.; Ysebaert, L.; Berquet, L.; Fataccioli, V.; Quelen, C.; Martin, A.; Parrens, M.; Lamant, L.; de Leval, L.; Gisselbrecht, C.; et al. Small Nucleolar RNA Expression Profiling Identifies Potential Prognostic Markers in Peripheral T-Cell Lymphoma. Blood 2012, 120, 3997–4005. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Xu, T.; Ganapathy, S.; Shadfan, M.; Long, M.; Huang, T.H.-M.; Thompson, I.; Yuan, Z.-M. Elevated SnoRNA Biogenesis Is Essential in Breast Cancer. Oncogene 2014, 33, 1348–1358. [Google Scholar] [CrossRef] [PubMed]
- Williams, G.T.; Farzaneh, F. Are SnoRNAs and SnoRNA Host Genes New Players in Cancer? Nat. Rev. Cancer 2012, 12, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.-Y.; Guo, P.; Boyd, J.; Sun, X.; Li, Q.; Zhou, W.; Dong, J.-T. Implication of SnoRNA U50 in Human Breast Cancer. J. Genet. Genomics 2009, 36, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.-Y.; Rodriguez, C.; Guo, P.; Sun, X.; Talbot, J.T.; Zhou, W.; Petros, J.; Li, Q.; Vessella, R.L.; Kibel, A.S.; et al. SnoRNA U50 Is a Candidate Tumor Suppressor Gene at 6q14.3 with a Mutation Associated with Clinically Significant Prostate Cancer. Hum. Mol. Genet. 2008, 17, 1031–1042. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Ye, M.-H.; Lv, S.-G.; Wang, Q.-X.; Wu, M.-J.; Xiao, B.; Kang, C.-S.; Zhu, X.-G. SNORD47, a Box C/D SnoRNA, Suppresses Tumorigenesis in Glioblastoma. Oncotarget 2017, 8, 43953–43966. [Google Scholar] [CrossRef]
- Zhu, W.; Niu, J.; He, M.; Zhang, L.; Lv, X.; Liu, F.; Jiang, L.; Zhang, J.; Yu, Z.; Zhao, L.; et al. SNORD89 Promotes Stemness Phenotype of Ovarian Cancer Cells by Regulating Notch1-c-Myc Pathway. J. Transl. Med. 2019, 17, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Zhao, L.; Wu, H.; Tian, G.; Dai, S.; Zhao, R.; Shan, B. C/D-Box Snord105b Promotes Tumorigenesis in Gastric Cancer via ALDOA/C-Myc Pathway. CPB 2018, 45, 2471–2482. [Google Scholar] [CrossRef] [Green Version]
- Cavaillé, J. Box C/D Small Nucleolar RNA Genes and the Prader-Willi Syndrome: A Complex Interplay. WIREs RNA 2017, 8, e1417. [Google Scholar] [CrossRef]
- Badrock, A.P.; Uggenti, C.; Wacheul, L.; Crilly, S.; Jenkinson, E.M.; Rice, G.I.; Kasher, P.R.; Lafontaine, D.L.J.; Crow, Y.J.; O’Keefe, R.T. Analysis of U8 SnoRNA Variants in Zebrafish Reveals How Bi-Allelic Variants Cause Leukoencephalopathy with Calcifications and Cysts. Am. J. Hum. Genet. 2020, 106, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Saxena, T.; Tandon, B.; Sharma, S.; Chameettachal, S.; Ray, P.; Ray, A.R.; Kulshreshtha, R. Combined MiRNA and MRNA Signature Identifies Key Molecular Players and Pathways Involved in Chikungunya Virus Infection in Human Cells. PLoS ONE 2013, 8, e79886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juranic Lisnic, V.; Babic Cac, M.; Lisnic, B.; Trsan, T.; Mefferd, A.; Das Mukhopadhyay, C.; Cook, C.H.; Jonjic, S.; Trgovcich, J. Dual Analysis of the Murine Cytomegalovirus and Host Cell Transcriptomes Reveal New Aspects of the Virus-Host Cell Interface. PLoS Pathog. 2013, 9, e1006311. [Google Scholar] [CrossRef] [PubMed]
- Fleming, D.S.; Miller, L.C. Identification of Small Non-Coding RNA Classes Expressed in Swine Whole Blood during HP-PRRSV Infection. Virology 2018, 517, 56–61. [Google Scholar] [CrossRef]
- Murray, J.L.; Sheng, J.; Rubin, D.H. A Role for H/ACA and C/D Small Nucleolar RNAs in Viral Replication. Mol. Biotechnol. 2014, 56, 429–437. [Google Scholar] [CrossRef]
- Stamm, S.; Lodmell, J.S. C/D Box SnoRNAs in Viral Infections: RNA Viruses Use Old Dogs for New Tricks. Noncoding RNA Res. 2019, 4, 46–53. [Google Scholar] [CrossRef]
- Deng, W.; Zhu, X.; Skogerbø, G.; Zhao, Y.; Fu, Z.; Wang, Y.; He, H.; Cai, L.; Sun, H.; Liu, C.; et al. Organization of the Caenorhabditis Elegans Small Non-Coding Transcriptome: Genomic Features, Biogenesis, and Expression. Genome Res. 2006, 16, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Hazen, S.P.; Naef, F.; Quisel, T.; Gendron, J.M.; Chen, H.; Ecker, J.R.; Borevitz, J.O.; Kay, S.A. Exploring the Transcriptional Landscape of Plant Circadian Rhythms Using Genome Tiling Arrays. Genome Biol. 2009, 10, R17. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, M.; Forné, I.; Ranjit, S.; Gratton, E.; Imhof, A.; Sassone-Corsi, P. BMAL1 Associates with NOP58 in the Nucleolus and Contributes to Pre-RRNA Processing. iScience 2020, 23, 101151. [Google Scholar] [CrossRef]
- Cavaillé, J.; Buiting, K.; Kiefmann, M.; Lalande, M.; Brannan, C.I.; Horsthemke, B.; Bachellerie, J.P.; Brosius, J.; Hüttenhofer, A. Identification of Brain-Specific and Imprinted Small Nucleolar RNA Genes Exhibiting an Unusual Genomic Organization. Proc. Natl. Acad. Sci. USA 2000, 97, 14311–14316. [Google Scholar] [CrossRef] [Green Version]
- Cavaillé, J.; Vitali, P.; Basyuk, E.; Hüttenhofer, A.; Bachellerie, J.P. A Novel Brain-Specific Box C/D Small Nucleolar RNA Processed from Tandemly Repeated Introns of a Noncoding RNA Gene in Rats. J. Biol. Chem. 2001, 276, 26374–26383. [Google Scholar] [CrossRef] [Green Version]
- Cavaillé, J.; Seitz, H.; Paulsen, M.; Ferguson-Smith, A.C.; Bachellerie, J.-P. Identification of Tandemly-Repeated C/D SnoRNA Genes at the Imprinted Human 14q32 Domain Reminiscent of Those at the Prader-Willi/Angelman Syndrome Region. Hum. Mol. Genet. 2002, 11, 1527–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erales, J.; Marchand, V.; Panthu, B.; Gillot, S.; Belin, S.; Ghayad, S.E.; Garcia, M.; Laforêts, F.; Marcel, V.; Baudin-Baillieu, A.; et al. Evidence for RRNA 2′-O-Methylation Plasticity: Control of Intrinsic Translational Capabilities of Human Ribosomes. Proc. Natl. Acad. Sci. USA 2017, 114, 12934–12939. [Google Scholar] [CrossRef] [Green Version]
- Hebras, J.; Krogh, N.; Marty, V.; Nielsen, H.; Cavaillé, J. Developmental Changes of RRNA Ribose Methylations in the Mouse. RNA Biol. 2020, 17, 150–164. [Google Scholar] [CrossRef]
- Belin, S.; Beghin, A.; Solano-Gonzàlez, E.; Bezin, L.; Brunet-Manquat, S.; Textoris, J.; Prats, A.-C.; Mertani, H.C.; Dumontet, C.; Diaz, J.-J. Dysregulation of Ribosome Biogenesis and Translational Capacity Is Associated with Tumor Progression of Human Breast Cancer Cells. PLoS ONE 2009, 4, e7147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcel, V.; Ghayad, S.E.; Belin, S.; Therizols, G.; Morel, A.-P.; Solano-Gonzàlez, E.; Vendrell, J.A.; Hacot, S.; Mertani, H.C.; Albaret, M.A.; et al. P53 Acts as a Safeguard of Translational Control by Regulating Fibrillarin and RRNA Methylation in Cancer. Cancer Cell 2013, 24, 318–330. [Google Scholar] [CrossRef] [Green Version]
- Krogh, N.; Jansson, M.D.; Häfner, S.J.; Tehler, D.; Birkedal, U.; Christensen-Dalsgaard, M.; Lund, A.H.; Nielsen, H. Profiling of 2′-O-Me in Human RRNA Reveals a Subset of Fractionally Modified Positions and Provides Evidence for Ribosome Heterogeneity. Nucleic Acids Res. 2016, 44, 7884–7895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janin, M.; Coll-SanMartin, L.; Esteller, M. Disruption of the RNA Modifications That Target the Ribosome Translation Machinery in Human Cancer. Mol. Cancer 2020, 19, 70. [Google Scholar] [CrossRef] [Green Version]
- Patil, P.; Kibiryeva, N.; Uechi, T.; Marshall, J.; O’Brien, J.E.; Artman, M.; Kenmochi, N.; Bittel, D.C. ScaRNAs Regulate Splicing and Vertebrate Heart Development. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2015, 1852, 1619–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitali, P.; Kiss, T. Cooperative 2′-O-Methylation of the Wobble Cytidine of Human Elongator TRNAMet(CAT) by a Nucleolar and a Cajal Body-Specific Box C/D RNP. Genes Dev. 2019, 33, 741–746. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Gurha, P.; Tran, E.J.; Maxwell, E.S.; Gupta, R. Sequential 2′-O-Methylation of Archaeal Pre-TRNATrp Nucleotides Is Guided by the Intron-Encoded but Trans-Acting Box C/D Ribonucleoprotein of Pre-TRNA. J. Biol. Chem. 2004, 279, 47661–47671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clouet d’Orval, B.; Bortolin, M.L.; Gaspin, C.; Bachellerie, J.P. Box C/D RNA Guides for the Ribose Methylation of Archaeal TRNAs. The TRNATrp Intron Guides the Formation of Two Ribose-Methylated Nucleosides in the Mature TRNATrp. Nucleic Acids Res. 2001, 29, 4518–4529. [Google Scholar] [CrossRef] [Green Version]
- Dennis, P.P.; Omer, A.; Lowe, T. A Guided Tour: Small RNA Function in Archaea. Mol. Microbiol. 2001, 40, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Elliott, B.A.; Ho, H.-T.; Ranganathan, S.V.; Vangaveti, S.; Ilkayeva, O.; Abou Assi, H.; Choi, A.K.; Agris, P.F.; Holley, C.L. Modification of Messenger RNA by 2′-O-Methylation Regulates Gene Expression in Vivo. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Deryusheva, S.; Gall, J.G. Small Cajal Body-Specific RNAs of Drosophila Function in the Absence of Cajal Bodies. Mol. Biol. Cell 2009, 20, 5250–5259. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Yang, J.; van Nues, R.; Watzinger, P.; Kötter, P.; Lafontaine, D.L.J.; Granneman, S.; Entian, K.-D. Specialized Box C/D SnoRNPs Act as Antisense Guides to Target RNA Base Acetylation. PLoS Genet. 2017, 13, e1006804. [Google Scholar] [CrossRef] [PubMed]
- Bazeley, P.S.; Shepelev, V.; Talebizadeh, Z.; Butler, M.G.; Fedorova, L.; Filatov, V.; Fedorov, A. SnoTARGET Shows That Human Orphan SnoRNA Targets Locate Close to Alternative Splice Junctions. Gene 2008, 408, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Kehr, S.; Bartschat, S.; Stadler, P.F.; Tafer, H. PLEXY: Efficient Target Prediction for Box C/D SnoRNAs. Bioinformatics 2011, 27, 279–280. [Google Scholar] [CrossRef] [Green Version]
- Sharma, E.; Sterne-Weiler, T.; O’Hanlon, D.; Blencowe, B.J. Global Mapping of Human RNA-RNA Interactions. Mol. Cell 2016, 62, 618–626. [Google Scholar] [CrossRef] [Green Version]
- Kishore, S.; Stamm, S. The SnoRNA HBII-52 Regulates Alternative Splicing of the Serotonin Receptor 2C. Science 2006, 311, 230–232. [Google Scholar] [CrossRef] [PubMed]
- Vitali, P.; Basyuk, E.; Le Meur, E.; Bertrand, E.; Muscatelli, F.; Cavaillé, J.; Huttenhofer, A. ADAR2-Mediated Editing of RNA Substrates in the Nucleolus Is Inhibited by C/D Small Nucleolar RNAs. J. Cell Biol. 2005, 169, 745–753. [Google Scholar] [CrossRef]
- Doe, C.M.; Relkovic, D.; Garfield, A.S.; Dalley, J.W.; Theobald, D.E.H.; Humby, T.; Wilkinson, L.S.; Isles, A.R. Loss of the Imprinted SnoRNA Mbii-52 Leads to Increased 5htr2c Pre-RNA Editing and Altered 5HT2CR-Mediated Behaviour. Hum. Mol. Genet. 2009, 18, 2140–2148. [Google Scholar] [CrossRef]
- Brüning, L.; Hackert, P.; Martin, R.; Davila Gallesio, J.; Aquino, G.R.R.; Urlaub, H.; Sloan, K.E.; Bohnsack, M.T. RNA Helicases Mediate Structural Transitions and Compositional Changes in Pre-Ribosomal Complexes. Nat. Commun. 2018, 9, 5383. [Google Scholar] [CrossRef]
- Sardana, R.; Liu, X.; Granneman, S.; Zhu, J.; Gill, M.; Papoulas, O.; Marcotte, E.M.; Tollervey, D.; Correll, C.C.; Johnson, A.W. The DEAH-Box Helicase Dhr1 Dissociates U3 from the Pre-RRNA to Promote Formation of the Central Pseudoknot. PLoS Biol. 2015, 13, e1002083. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, P.; Hackert, P.; Memet, I.; Sloan, K.E.; Bohnsack, M.T. The Human RNA Helicase DHX37 Is Required for Release of the U3 SnoRNP from Pre-Ribosomal Particles. RNA Biol. 2019, 16, 54–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aquino, G.R.R.; Krogh, N.; Hackert, P.; Martin, R.; Gallesio, J.D.; van Nues, R.W.; Schneider, C.; Watkins, N.J.; Nielsen, H.; Bohnsack, K.E.; et al. RNA Helicase-Mediated Regulation of SnoRNP Dynamics on Pre-Ribosomes and RRNA 2′-O-Methylation. Nucleic Acids Res. 2021, 49. [Google Scholar] [CrossRef]
- Leeds, N.B.; Small, E.C.; Hiley, S.L.; Hughes, T.R.; Staley, J.P. The Splicing Factor Prp43p, a DEAH Box ATPase, Functions in Ribosome Biogenesis. Mol. Cell. Biol. 2006, 26, 513–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasler, D.; Meduri, R.; Bąk, M.; Lehmann, G.; Heizinger, L.; Wang, X.; Li, Z.-T.; Sement, F.M.; Bruckmann, A.; Dock-Bregeon, A.-C.; et al. The Alazami Syndrome-Associated Protein LARP7 Guides U6 Small Nuclear RNA Modification and Contributes to Splicing Robustness. Mol. Cell 2020, 77, 1014–1031.e13. [Google Scholar] [CrossRef]
- Huang, C.; Shi, J.; Guo, Y.; Huang, W.; Huang, S.; Ming, S.; Wu, X.; Zhang, R.; Ding, J.; Zhao, W.; et al. A SnoRNA Modulates MRNA 3′ End Processing and Regulates the Expression of a Subset of MRNAs. Nucleic Acids Res. 2017, 45, 8647–8660. [Google Scholar] [CrossRef] [Green Version]
- Siprashvili, Z.; Webster, D.E.; Johnston, D.; Shenoy, R.M.; Ungewickell, A.J.; Bhaduri, A.; Flockhart, R.; Zarnegar, B.J.; Che, Y.; Meschi, F.; et al. The Noncoding RNAs SNORD50A and SNORD50B Bind K-Ras and Are Recurrently Deleted in Human Cancer. Nat. Genet. 2016, 48, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Che, Y.; Siprashvili, Z.; Kovalski, J.R.; Jiang, T.; Wozniak, G.; Elcavage, L.; Khavari, P.A. KRAS Regulation by Small Non-Coding RNAs and SNARE Proteins. Nat. Commun. 2019, 10, 5118. [Google Scholar] [CrossRef]
- Su, X.; Feng, C.; Wang, S.; Shi, L.; Gu, Q.; Zhang, H.; Lan, X.; Zhao, Y.; Qiang, W.; Ji, M.; et al. The Noncoding RNAs SNORD50A and SNORD50B-Mediated TRIM21-GMPS Interaction Promotes the Growth of P53 Wild-Type Breast Cancers by Degrading P53. Cell Death Differ. 2021. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-S.; Camacho, C.V.; Nagari, A.; Malladi, V.S.; Challa, S.; Kraus, W.L. Activation of PARP-1 by SnoRNAs Controls Ribosome Biogenesis and Cell Growth via the RNA Helicase DDX21. Mol. Cell 2019, 75, 1270–1285.e14. [Google Scholar] [CrossRef]
- D’Souza, M.N.; Gowda, N.K.C.; Tiwari, V.; Babu, R.O.; Anand, P.; Dastidar, S.G.; Singh, R.; Selvaraja, B.; Pal, R.; Ramesh, A.; et al. FMRP Interacts with C/D Box SnoRNA in the Nucleus and Regulates Ribosomal RNA Methylation. iScience 2018, 9, 399–411. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.; Sharma, M.R.; Shi, X.; Agrawal, R.K.; Joseph, S. Fragile X Mental Retardation Protein Regulates Translation by Binding Directly to the Ribosome. Mol. Cell 2014, 54, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Simsek, D.; Tiu, G.C.; Flynn, R.A.; Byeon, G.W.; Leppek, K.; Xu, A.F.; Chang, H.Y.; Barna, M. The Mammalian Ribo-Interactome Reveals Ribosome Functional Diversity and Heterogeneity. Cell 2017, 169, 1051–1065.e18. [Google Scholar] [CrossRef] [Green Version]
- Brandis, K.A.; Gale, S.; Jinn, S.; Langmade, S.J.; Dudley-Rucker, N.; Jiang, H.; Sidhu, R.; Ren, A.; Goldberg, A.; Schaffer, J.E.; et al. Box C/D Small Nucleolar RNA (SnoRNA) U60 Regulates Intracellular Cholesterol Trafficking. J. Biol. Chem. 2013, 288, 35703–35713. [Google Scholar] [CrossRef] [Green Version]
- Michel, C.I.; Holley, C.L.; Scruggs, B.S.; Sidhu, R.; Brookheart, R.T.; Listenberger, L.L.; Behlke, M.A.; Ory, D.S.; Schaffer, J.E. Small Nucleolar RNAs U32a, U33 and U35a Are Critical Mediators of Metabolic Stress. Cell Metab. 2011, 14, 33–44. [Google Scholar] [CrossRef] [Green Version]
- Li, M.W.; Sletten, A.C.; Lee, J.; Pyles, K.D.; Matkovich, S.J.; Ory, D.S.; Schaffer, J.E. Nuclear Export Factor 3 Regulates Localization of Small Nucleolar RNAs. J. Biol. Chem. 2017, 292, 20228–20239. [Google Scholar] [CrossRef] [Green Version]
- Huo, C.-Y.; Chang, M.-L.; Cheng, H.; Ma, T.-T.; Fu, Y.; Wang, Y.; Wang, Y.-Y.; Kan, Y.-C.; Li, D.-D. Small Nucleolar RNA of Silkworm Can Translocate from the Nucleolus to the Cytoplasm under Abiotic Stress. Cell Biol. Int. 2021, 45. [Google Scholar] [CrossRef] [PubMed]
- Rimer, J.M.; Lee, J.; Holley, C.L.; Crowder, R.J.; Chen, D.L.; Hanson, P.I.; Ory, D.S.; Schaffer, J.E. Long-Range Function of Secreted Small Nucleolar RNAs That Direct 2′-O-Methylation. J. Biol. Chem. 2018, 293, 13284–13296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanford, J.R.; Wang, X.; Mort, M.; Vanduyn, N.; Cooper, D.N.; Mooney, S.D.; Edenberg, H.J.; Liu, Y. Splicing Factor SFRS1 Recognizes a Functionally Diverse Landscape of RNA Transcripts. Genome Res. 2009, 19, 381–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Änkö, M.-L.; Müller-McNicoll, M.; Brandl, H.; Curk, T.; Gorup, C.; Henry, I.; Ule, J.; Neugebauer, K.M. The RNA-Binding Landscapes of Two SR Proteins Reveal Unique Functions and Binding to Diverse RNA Classes. Genome Biol. 2012, 13, R17. [Google Scholar] [CrossRef] [Green Version]
- Sperling, R. Small Non-Coding RNA within the Endogenous Spliceosome and Alternative Splicing Regulation. Biochim. Biophys. Acta Gene Regul. Mech. 2019, 1862, 194406. [Google Scholar] [CrossRef]
- Schubert, T.; Pusch, M.C.; Diermeier, S.; Benes, V.; Kremmer, E.; Imhof, A.; Längst, G. Df31 Protein and SnoRNAs Maintain Accessible Higher-Order Structures of Chromatin. Mol. Cell 2012, 48, 434–444. [Google Scholar] [CrossRef] [Green Version]
- Dudnakova, T.; Dunn-Davies, H.; Peters, R.; Tollervey, D. Mapping Targets for Small Nucleolar RNAs in Yeast. Wellcome Open Res. 2018, 3. [Google Scholar] [CrossRef]
- Labialle, S.; Cavaillé, J. Do Repeated Arrays of Regulatory Small-RNA Genes Elicit Genomic Imprinting? Concurrent Emergence of Large Clusters of Small Non-Coding RNAs and Genomic Imprinting at Four Evolutionarily Distinct Eutherian Chromosomal Loci. Bioessays 2011, 33, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, T.; del Gaudio, D.; German, J.R.; Shinawi, M.; Peters, S.U.; Person, R.E.; Garnica, A.; Cheung, S.W.; Beaudet, A.L. Prader-Willi Phenotype Caused by Paternal Deficiency for the HBII-85 C/D Box Small Nucleolar RNA Cluster. Nat. Genet. 2008, 40, 719–721. [Google Scholar] [CrossRef] [Green Version]
- de Smith, A.J.; Purmann, C.; Walters, R.G.; Ellis, R.J.; Holder, S.E.; Van Haelst, M.M.; Brady, A.F.; Fairbrother, U.L.; Dattani, M.; Keogh, J.M.; et al. A Deletion of the HBII-85 Class of Small Nucleolar RNAs (SnoRNAs) Is Associated with Hyperphagia, Obesity and Hypogonadism. Hum. Mol. Genet. 2009, 18, 3257–3265. [Google Scholar] [CrossRef]
- Duker, A.L.; Ballif, B.C.; Bawle, E.V.; Person, R.E.; Mahadevan, S.; Alliman, S.; Thompson, R.; Traylor, R.; Bejjani, B.A.; Shaffer, L.G.; et al. Paternally Inherited Microdeletion at 15q11.2 Confirms a Significant Role for the SNORD116 C/D Box SnoRNA Cluster in Prader-Willi Syndrome. Eur. J. Hum. Genet. 2010, 18, 1196–1201. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Potter, K.J.; Burnett, L.C.; Orsso, C.E.; Inman, M.; Ryman, D.C.; Haqq, A.M. Prader-Willi-Like Phenotype Caused by an Atypical 15q11.2 Microdeletion. Genes 2020, 11, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skryabin, B.V.; Gubar, L.V.; Seeger, B.; Pfeiffer, J.; Handel, S.; Robeck, T.; Karpova, E.; Rozhdestvensky, T.S.; Brosius, J. Deletion of the MBII-85 SnoRNA Gene Cluster in Mice Results in Postnatal Growth Retardation. PLoS Genet. 2007, 3, e235. [Google Scholar] [CrossRef]
- Ding, F.; Li, H.H.; Zhang, S.; Solomon, N.M.; Camper, S.A.; Cohen, P.; Francke, U. SnoRNA Snord116 (Pwcr1/MBII-85) Deletion Causes Growth Deficiency and Hyperphagia in Mice. PLoS ONE 2008, 3, e1709. [Google Scholar] [CrossRef] [Green Version]
- Polex-Wolf, J.; Lam, B.Y.; Larder, R.; Tadross, J.; Rimmington, D.; Bosch, F.; Cenzano, V.J.; Ayuso, E.; Ma, M.K.; Rainbow, K.; et al. Hypothalamic Loss of Snord116 Recapitulates the Hyperphagia of Prader-Willi Syndrome. J. Clin. Investig. 2018, 128, 960–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, A.; Copping, N.A.; Onaga, B.; Pride, M.C.; Coulson, R.L.; Yang, M.; Yasui, D.H.; LaSalle, J.M.; Silverman, J.L. Cognitive Deficits in the Snord116 Deletion Mouse Model for Prader-Willi Syndrome. Neurobiol. Learn. Mem. 2019, 165, 106874. [Google Scholar] [CrossRef]
- Lui, L.M.; Uzilov, A.V.; Bernick, D.L.; Corredor, A.; Lowe, T.M.; Dennis, P.P. Methylation Guide RNA Evolution in Archaea: Structure, Function and Genomic Organization of 110 C/D Box SRNA Families across Six Pyrobaculum Species. Nucleic Acids Res. 2018, 46, 5678–5691. [Google Scholar] [CrossRef]
Figure 1.
Schematic overview of guide box C/D small nucleolar ribonucleoproteins (C/D snoRNPs) and their target RNAs. (A) Box C/D small nucleolar RNAs (C/D snoRNAs) contain conserved sequences called C/D and C′/D′ boxes that fold into a kink-turn (K-turn) containing Watson–Crick pairings (solid blue lines) and non-canonical pairings U:U, G:A, and A:G (dotted blue lines). (B) Conventional C/D box snoRNPs associate a C/D snoRNA with the core proteins Nop1p/Fibrillarin (FBL), Nop58p/NOP58, Nop56p/NOP56 and Snu13p/SNU13 in Saccharomyces cerevisiae and Human, respectively. FBL is the methyltranferase that catalyzes the 2′-O-methylation of target RNAs. (C) Schematic representation of the bidimensional human SNORD24 structure (adapted from [50]). This snoRNA guides the 2′-O-methylation of 28S rRNA at positions C2338 and C2352. Classically, the methylated nucleotide (red star) hybridizes with the nucleotide located at the fifth position upstream of the D and D′ box sequences. (D) Schematic representation of the bidimensional U3 snoRNA structure and its association with pre-18S rRNA in Saccharomyces cerevisiae (adapted from [61,62]). U3 contains two C/D-like motifs called C′/D and B/C boxes that fold into a K-turn recognized by the Snu13p core protein [14]. In addition, the B/C motif recruits Rrp9p (U3-55K in humans) that is essential for 18S rRNA processing. The A/A′ motifs and the 5′/3′ hinges, located in the 5′ terminal domain of U3, hybridize with the 18S rRNA (solid red lines) and the 5′ ETS (external transcribed spacer, dotted red lines) of the 35S pre-rRNA, respectively. The base-pairing is located near the A0 and A1 pre-rRNA cleavage sites and is essential for the processing of 18S rRNA.
Figure 1.
Schematic overview of guide box C/D small nucleolar ribonucleoproteins (C/D snoRNPs) and their target RNAs. (A) Box C/D small nucleolar RNAs (C/D snoRNAs) contain conserved sequences called C/D and C′/D′ boxes that fold into a kink-turn (K-turn) containing Watson–Crick pairings (solid blue lines) and non-canonical pairings U:U, G:A, and A:G (dotted blue lines). (B) Conventional C/D box snoRNPs associate a C/D snoRNA with the core proteins Nop1p/Fibrillarin (FBL), Nop58p/NOP58, Nop56p/NOP56 and Snu13p/SNU13 in Saccharomyces cerevisiae and Human, respectively. FBL is the methyltranferase that catalyzes the 2′-O-methylation of target RNAs. (C) Schematic representation of the bidimensional human SNORD24 structure (adapted from [50]). This snoRNA guides the 2′-O-methylation of 28S rRNA at positions C2338 and C2352. Classically, the methylated nucleotide (red star) hybridizes with the nucleotide located at the fifth position upstream of the D and D′ box sequences. (D) Schematic representation of the bidimensional U3 snoRNA structure and its association with pre-18S rRNA in Saccharomyces cerevisiae (adapted from [61,62]). U3 contains two C/D-like motifs called C′/D and B/C boxes that fold into a K-turn recognized by the Snu13p core protein [14]. In addition, the B/C motif recruits Rrp9p (U3-55K in humans) that is essential for 18S rRNA processing. The A/A′ motifs and the 5′/3′ hinges, located in the 5′ terminal domain of U3, hybridize with the 18S rRNA (solid red lines) and the 5′ ETS (external transcribed spacer, dotted red lines) of the 35S pre-rRNA, respectively. The base-pairing is located near the A0 and A1 pre-rRNA cleavage sites and is essential for the processing of 18S rRNA.

Figure 2.
Model of eukaryotic C/D snoRNP biogenesis. The loading of the core proteins SNU13, NOP58, NOP56, and FBL does not occur autonomously in eukaryotic cells but is mediated by several assembly factors. The R2TP complex contains the proteins RUVBL1, RUVBL2, RPAP3, and PIH1D1 and the latter interacts directly with NOP58 to stabilize it. The assembly platform NUFIP1 and its stabilizing factor ZNHIT3 interact with SNU13 to form a trimer. The essential contribution of the assembly factor BCD1 is less clear; it is integrated in a network of interactions involving PIH1D1, NUFIP1, RUVBL1&2, NOP58, and SNU13, which suggests that these proteins form a protein-only pre-snoRNP complex that scaffolds SNU13 and NOP58 core protein assembly. The proteins are then loaded on nascent snoRNA molecules in a co-transcriptional way upon the direct binding of SNU13 to the K-turn structures formed by the conserved C/D motifs. The efficient RNP assembly of intronic snoRNAs is favored by the association of the splicing and biogenesis machineries via the splicing factor AQR. The nucleation of the pre-snoRNP protects the snoRNA from extensive trimming by exonuclease activities. When released, the assembly factors might be recycled in a new biogenesis process (dotted line). Finally, the C/D snoRNPs are transported to CBs and nucleoli for functional purposes. This model is valid in both Saccharomyces cerevisiae and Human.
Figure 2.
Model of eukaryotic C/D snoRNP biogenesis. The loading of the core proteins SNU13, NOP58, NOP56, and FBL does not occur autonomously in eukaryotic cells but is mediated by several assembly factors. The R2TP complex contains the proteins RUVBL1, RUVBL2, RPAP3, and PIH1D1 and the latter interacts directly with NOP58 to stabilize it. The assembly platform NUFIP1 and its stabilizing factor ZNHIT3 interact with SNU13 to form a trimer. The essential contribution of the assembly factor BCD1 is less clear; it is integrated in a network of interactions involving PIH1D1, NUFIP1, RUVBL1&2, NOP58, and SNU13, which suggests that these proteins form a protein-only pre-snoRNP complex that scaffolds SNU13 and NOP58 core protein assembly. The proteins are then loaded on nascent snoRNA molecules in a co-transcriptional way upon the direct binding of SNU13 to the K-turn structures formed by the conserved C/D motifs. The efficient RNP assembly of intronic snoRNAs is favored by the association of the splicing and biogenesis machineries via the splicing factor AQR. The nucleation of the pre-snoRNP protects the snoRNA from extensive trimming by exonuclease activities. When released, the assembly factors might be recycled in a new biogenesis process (dotted line). Finally, the C/D snoRNPs are transported to CBs and nucleoli for functional purposes. This model is valid in both Saccharomyces cerevisiae and Human.

Figure 3.
The emerging diversity of C/D box snoRNA processing and of C/D snoRNP biogenesis and function. Precursor C/D box snoRNAs are produced by both independent (1) and intronic (2 and 3) genes, then processed by the combined action of intron lariat debranching in the case of intronic snoRNAs and exonucleotidic activities acting at both the 5′ and 3′ ends. Co-transcriptional recruitment of snoRNP core proteins on the snoRNA allows the correct processing and biogenesis of the C/D snoRNP. Then, the particle reaches the Cajal body (CB) and the nucleolus to perform activities on snRNAs and rRNAs (in blue). In addition to these conventional mechanisms, C/D snoRNPs perform new activities (in gold italics) linked to the formation of lncRNAs flanked by snoRNA sequences at one or both ends generated from partially-processed, snoRNA-hosting introns (3), to the recruitment of additional proteins conferring new properties and/or enzymatic activities (4) or to the exchange of conventional core proteins by alternative proteins (5), to the trafficking to new subcellular locations (6), and to the generation of snoRNA-derived fragments (sdRNAs) or regulatory ribonucleoproteins (regRNPs; 7).
Figure 3.
The emerging diversity of C/D box snoRNA processing and of C/D snoRNP biogenesis and function. Precursor C/D box snoRNAs are produced by both independent (1) and intronic (2 and 3) genes, then processed by the combined action of intron lariat debranching in the case of intronic snoRNAs and exonucleotidic activities acting at both the 5′ and 3′ ends. Co-transcriptional recruitment of snoRNP core proteins on the snoRNA allows the correct processing and biogenesis of the C/D snoRNP. Then, the particle reaches the Cajal body (CB) and the nucleolus to perform activities on snRNAs and rRNAs (in blue). In addition to these conventional mechanisms, C/D snoRNPs perform new activities (in gold italics) linked to the formation of lncRNAs flanked by snoRNA sequences at one or both ends generated from partially-processed, snoRNA-hosting introns (3), to the recruitment of additional proteins conferring new properties and/or enzymatic activities (4) or to the exchange of conventional core proteins by alternative proteins (5), to the trafficking to new subcellular locations (6), and to the generation of snoRNA-derived fragments (sdRNAs) or regulatory ribonucleoproteins (regRNPs; 7).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Baldini, L.; Charpentier, B.; Labialle, S. Emerging Data on the Diversity of Molecular Mechanisms Involving C/D snoRNAs. Non-Coding RNA 2021, 7, 30. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7020030
AMA Style
Baldini L, Charpentier B, Labialle S. Emerging Data on the Diversity of Molecular Mechanisms Involving C/D snoRNAs. Non-Coding RNA. 2021; 7(2):30. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7020030
Chicago/Turabian StyleBaldini, Laeya, Bruno Charpentier, and Stéphane Labialle. 2021. "Emerging Data on the Diversity of Molecular Mechanisms Involving C/D snoRNAs" Non-Coding RNA 7, no. 2: 30. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7020030
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.