
Long Non-Coding RNA Regulation of Epigenetics in Vascular Cells
by
 , , , and
, , , and
Hester Garratt
1 ,
,
Robert Ashburn
1 ,
,
Miron Sopić
2,
Antonella Nogara
1,
Andrea Caporali
1,* and
Tijana Mitić
1,* 1
Centre for Cardiovascular Science, The Queen’s Medical Research Institute, Edinburgh BioQuarter, 47 Little France Crescent, Edinburgh EH16 4TJ, UK
2
Department of Medical Biochemistry, Faculty of Pharmacy, University of Belgrade, 11000 Belgrade, Serbia
*
Authors to whom correspondence should be addressed.
Non-Coding RNA 2021, 7(4), 62; https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7040062
Submission received: 30 June 2021
/
Revised: 2 September 2021
/
Accepted: 14 September 2021
/
Published: 23 September 2021
(This article belongs to the Special Issue Non-coding RNA: 5th Anniversary)
Abstract
:The vascular endothelium comprises the interface between the circulation and the vessel wall and, as such, is under the dynamic regulation of vascular signalling, nutrients, and hypoxia. Understanding the molecular drivers behind endothelial cell (EC) and vascular smooth muscle cell (VSMC) function and dysfunction remains a pivotal task for further clinical progress in tackling vascular disease. A newly emerging era in vascular biology with landmark deep sequencing approaches has provided us with the means to profile diverse layers of transcriptional regulation at a single cell, chromatin, and epigenetic level. This review describes the roles of major vascular long non-coding RNA (lncRNAs) in the epigenetic regulation of EC and VSMC function and discusses the recent progress in their discovery, detection, and functional characterisation. We summarise new findings regarding lncRNA-mediated epigenetic mechanisms—often regulated by hypoxia—within the vascular endothelium and smooth muscle to control vascular homeostasis in health and disease. Furthermore, we outline novel molecular techniques being used in the field to delineate the lncRNA subcellular localisation and interaction with proteins to unravel their biological roles in the epigenetic regulation of vascular genes.
1. The Role of the Vascular Endothelium in Health and Disease
The vascular endothelium comprises the layer of endothelial cells (ECs) that create the interface between the circulation and the vessel wall that is under dynamic regulation in accordance with its environment to maintain perfusion and the supply of oxygen and nutrients required for cellular metabolism [1,2]. A healthy vascular endothelium is key to maintaining vascular homeostasis, but little is known of what regulates its distinct and characteristic function. The angiogenic capacity of ECs is vital during embryonic and vascular tissue development and during wound healing in damaged tissues. Homeostatic mechanisms encourage hypoxia- or vascular endothelial growth factor (VEGF)-induced angiogenesis required for inflammatory and reproductive processes and post-ischaemic tissue regeneration (Figure 1A); and suppress vascular inflammation and thrombosis [3,4,5]. The vasculature is implicated in a range of pathological disease states, such as coronary and small vessel disease, peripheral arterial diseases, hypertension, aortic aneurysm, and diabetes mellitus—all of which are worldwide leading causes of morbidity and mortality [6]. Endothelial cell dysfunction can occur due to hypoxic or oxidative injury and predisposes to atherosclerotic and ischaemic disease by potentiating vascular inflammation and atherothrombosis (Figure 1B). Hence, an increasing understanding of endothelial homeostasis is continually sought as a primacy to the comprehension of many complex cardiovascular conditions. Increasingly, studies are being designed to elucidate molecular mechanisms behind EC and VSMC dysfunction and investigate drug therapies which induce revascularisation to facilitate repair of ischaemic sites.
2. The Era of lncRNAs in Vascular Biology
A new era for vascular biology has emerged with technological advancements and next-generation sequencing (NGS) approaches which have largely empowered our exploration of genomes over the past ten years [8]. Hence, the focus has shifted from the coding to the non-coding part of the genome, comprising non-coding RNA genes, regulatory DNA sequences, introns, and interspersed nuclear elements. The large number of non-coding RNAs named short (microRNA, miRNA) and long non-coding RNAs (lncRNAs) regulate important cellular functions—including within vascular cells. Beside the ENCODE database (https://github.com/ENCODE-DCC/encoded/releases/tag/v121.0, accessed on 4 May 2021), there is now NONCODE (http://www.noncode.org/, accessed on 8 August 2021), an integrated knowledge database dedicated especially to lncRNAs [9].
The lncRNAs are a heterogeneous group of RNAs defined as transcripts >200 nucleotides long. LncRNAs can be categorised by their origin of transcription and position of neighbouring genes into sense, natural antisense, intronic, intergenic, bidirectional promoter, and enhancer lncRNAs. Their biogenesis is distinct from that of mRNA as they are poorly evolutionarily conserved, often less abundant, more tissue-specific, display distinct spatiotemporal expression and specific subcellular localisation and function, and tend to form secondary structural domains folded into specific scaffolds [10,11]. Indeed, despite significant differences in the primary sequence, lncRNA homology across species can be identified through the analysis of their secondary and tertiary structures key to their specific function [12]. The use of NGS combined with high-resolution methods to detect lncRNA sites of interaction on a genome or with other RNA species (via enzymatic or chemical probing) has provided insight into the structural folding modules, domains, and loops for several vascular lncRNAs like Braveheart, HOTAIR, MEG3, RepA, Xist, and SRA [13]. Their structural conservation, as opposed to nucleotide sequence conservation, appears to be crucial for maintaining their function. For example, maternally expressed gene 3 (MEG3) is a highly structured endothelial-enriched lncRNA whose complex secondary and tertiary molecular architecture permits its key interactions with epigenetic regulators and tumour suppressor genes [14,15,16]. The tertiary structure of MEG3—possibly owing to conserved motifs with a high GC (guanine-cytosine) content (57%)—enables the formation of pseudoknots that are crucial to its regulation of the p53 pathway, DNA damage response, and associated apoptosis to protect EC function [14,17,18]. Understanding of lncRNA functions in vivo is additionally complicated by the fact that, due to alternative splicing, lncRNAs can exist in multiple functional isoforms in the cell. For example, different transcripts of antisense ncRNA in the INK4 locus (ANRIL) have opposite expression patterns in coronary arteries and even exert antagonising functions in ECs [17].
Recently, greater focus has been placed on identifying the subcellular localisation of vascular lncRNAs in order to functionally characterise nuclear-, cytoplasmic-, and chromatin-enriched ones [18]. Indeed, MEG3, MALAT1 (metastasis associated lung adenocarcinoma transcript 1), NEAT1 (nuclear enriched abundant transcript 1 or nuclear paraspeckle assembly transcript 1), LEENE (lncRNA that enhances eNOS expression), and GATA-6AS (antisense transcript of GATA6) in endothelial cells, and ANRIL, SMILR (smooth muscle enriched long noncoding RNA), and SENCR (smooth muscle and endothelial cell enriched migration/differentiation-associated lncRNA) in vascular smooth muscle cells have been reported as regulators of endothelial and smooth muscle differentiation [1,8]; vascular cell proliferation, apoptosis, and vascular remodelling [19,20]; and endothelial function and angiogenesis [5,21,22,23,24,25] through affecting gene transcription to mediate endothelial homeostasis or lead to vascular pathologies (Table 1).
ENCODE RNA sequencing datasets suggest that subcellular lncRNA nuclear/cytoplasmic ratios may be mechanised by factors including slow nuclear export, nuclear or cytoplasmic degradation, and splicing efficiency [32]. Increasingly, lncRNAs are reported to be retained for activity in the nucleus (nucleolus and chromatin) and associated with local functions including mediating inter- and intra-chromosomal interactions and sequestering RNA complexes at chromatin [33]. Through their primary sequence or secondary structures, nuclear lncRNAs can transcriptionally or post-transcriptionally moderate vascular gene expression through cis- or trans-regulatory mechanisms. LncRNAs may regulate interactions with DNA/RNA-binding proteins at specific loci through acting as guides, scaffolds, or decoys, may mediate chromatin looping and higher-order chromatin interactions between promoters and distal enhancers, and may control alternative splicing, whilst lncRNA transcription itself may favourably re-shape chromatin confirmation, aid transcription factor recruitment, or impose enhancer-like effects on the neighbouring genes through the transcription from enhancer regions (aka elncRNAs) [34,35].
Cytoplasmic localisation, however, allows for lncRNA modulation of mRNA stability and translation, microRNA activity, protein turnover, and signal transduction pathways through acting as cytosolic scaffolds, degradation tags, sponges, and molecular decoys—ultimately mediating physiological and pathological vascular processes, including angiogenesis and atherosclerosis in diseases such as hypertension, retinopathy, and malignancy [36,37]. MEG3, for example, has been observed to translocate to the cytoplasm upon hypoxia in pulmonary artery smooth muscle cells (PASMCs), subsequently sequestering and inducing the degradation of microRNA miR-328. This ultimately upregulates IGF1R and regulates hypoxia-induced PASMC proliferation and pulmonary hypertension [38].
In addition to intracellular localization, lncRNAs have been detected extracellularly in several body fluids, including whole blood, plasma, urine, saliva, and gastric juice [39,40,41]. Extracellular lncRNAs are relatively resistant to degradation, reflect dynamically intracellular changes during disease, and are easily detected in body fluids; thus, showing high biomarker potential [38]. Diagnostic and prognostic properties of lncRNAs have been demonstrated in cardiometabolic diseases. The plasma levels of mitochondria-derived lncRNA long intergenic non-coding RNA predicting cardiac remodelling (LIPCAR) are associated with left ventricular remodelling following myocardial infarction and an increased risk of developing heart disease [42]. ANRIL and QSOX1 (quiescin sulfhydryl oxidase 1) have also been detected in plasma of such individuals and thereby serve as potential biomarkers—although it is unclear which cellular compartment or cell type they arise from. Moreover, circulating SENCR levels have been shown to discriminate pioglitazone therapy responders from non-responders to a better extent than traditional clinical markers [43,44]. Many other circulating lncRNAs may prove to be powerful biomarkers for diabetic and cardiovascular disease states, yet their diagnostic and prognostic values remain an open field of investigation [40].
Remarkably, some recent studies have even questioned the non-coding nature of lncRNAs. Novel in silico approaches have suggested the existence of small open reading frames (ORFs) within a small number of lncRNAs, whilst ribosomal profiling has revealed their potential to be translated [45]. Mass spectrometry-based proteomics approaches coupled with transcriptome sequencing have effectively been used to identify micro-peptides encoded by putative lncRNAs. The validation of ORFs and post-translational function is required to establish any roles these micro-peptides may have in vascular biology [46].
One resource that has helped vascular researchers explore the layers of gene regulation to assess direct and controlled comparison of gene expression between different cell types is the Tabula Muris, available here https://tabula-muris.ds.czbiohub.org/, accessed on 6 June 2020) [47]. Similarly, the single-cell transcriptional profiles of fibroblasts and the vascular mural cell are valuable resource serving as a “postal code” to provide molecular information to vascular researchers, available here http://betsholtzlab.org/Publications/databases.html, accessed on 2 September 2020) [48]. RNA sequencing data of the brain, lung, and heart endothelial translatome from adult Cdh5CreERT2/+; Rpl22HA/+ mice at baseline and during inflammation can be visualised at http://rehmanlab.org/ribo [2] (accessed on 4 May 2021); the human heart translatome visualised at https://shiny.mdc-berlin.de/cardiac-translatome/; [49], (accessed on 4 May 2021) and the hypoxia-induced rat cardiomyocytes translatome visualized in reference [50].
3. Long Non-Coding RNAs as Remodellers of the Vascular Epigenome
Epigenetics imply heritable changes to the genome that do not involve changes to the underlying nucleotide sequence. Although the biological relevance for the vast majority of lncRNAs remains elusive, lncRNAs are known to influence gene expression at a transcriptional, post-transcriptional, translational, and post-translational level; and to represent a new layer of epigenetic control in vascular homeostasis that is dysregulated in disease [6,20,47].
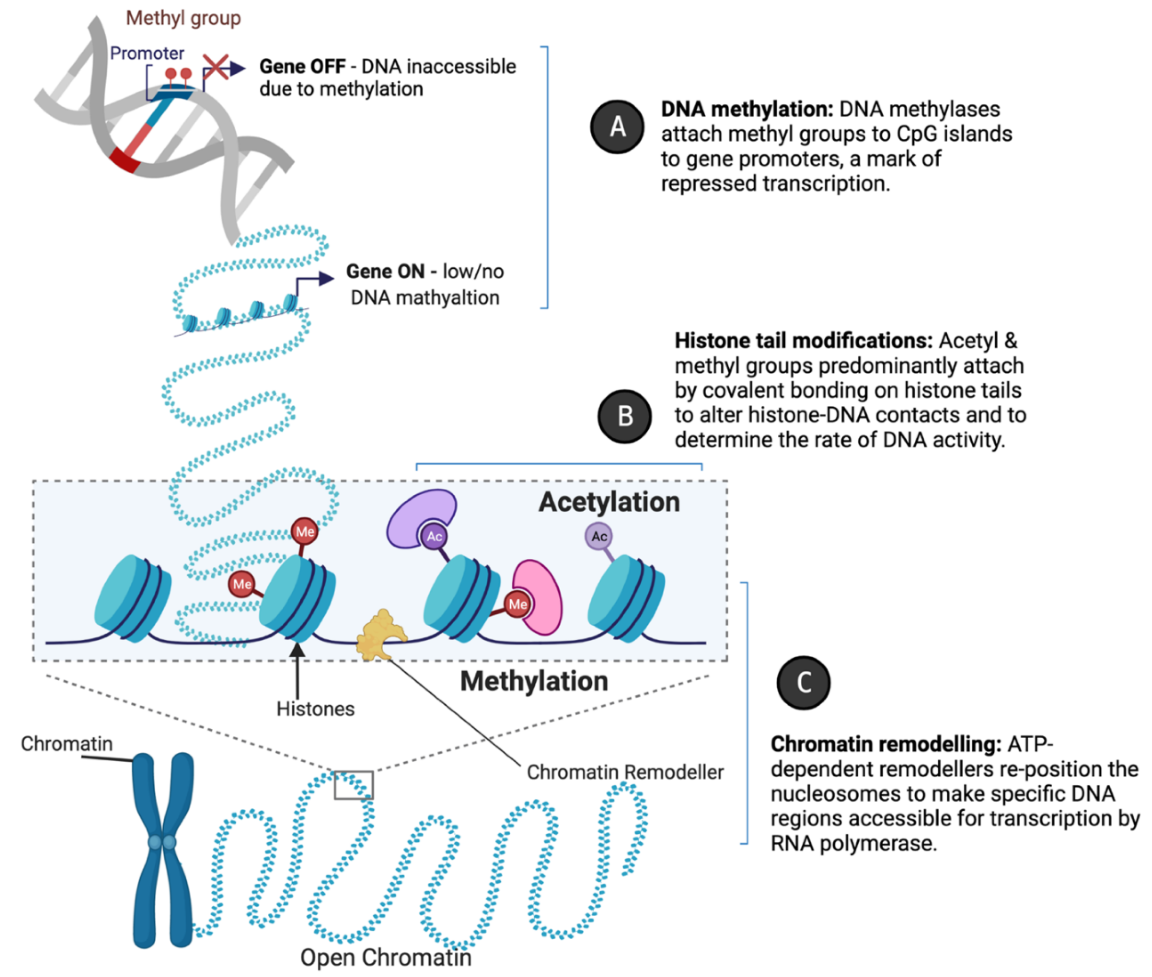
Epigenetic changes fine-tune vascular gene expression in a temporal, tissue-specific fashion through functional chromatin alterations which alter DNA accessibility at specific loci for replicative enzymes or transcription factors. The mechanisms include (a) DNA methylation, (b) histone tail modification, and (c) chromatin remodelling. DNA methylation typically acts to repress gene expression through impeding transcription factor binding; histone-modifying enzymes (“writers” and “erasers”) catalyse the addition and removal of acetylation and methylation marks to alter chromatin architecture; whilst chromatin remodellers re-position nucleosomes to condense or decondense chromatin, restricting or enabling access for transcriptional machinery, respectively (Figure 2A–C).
Paragons of endothelial-enriched genes crucial to endothelial homeostasis, such as eNOS (endothelial Nitric Oxide Synthase) and many pro-angiogenic genes implicated in the revascularisation response to hypoxia, are known to be dynamically epigenetically regulated by DNA methylation and changes in the histone density [51]. Moreover, histone methylation has been shown to epigenetically regulate gene expression in accordance with a hypoxic environment. Jumonji C domain-containing histone demethylases are part of the 2-oxoglutarate-dependent dioxygenase (2-OGDD) enzyme family which depend on oxygen as a co-factor for their activity. In the absence of oxygen, dioxygenase activity, and therefore lysine demethylase activity, is inhibited—inducing histone tail hypermethylation and altering the expression of certain genes to control hypoxia-dependent cell fate [52,53].
LncRNAs can act via multiple mechanisms to epigenetically regulate vascular gene transcription—including acting to recruit, scaffold, or sequester such chromatin remodellers and chromatin modifiers to influence chromatin reorganisation (Table 1) [6]. Some lncRNAs can be taken up by extracellular vesicles (EVs) as exosomes that are secreted by many cell types as cell–cell communicators in response to stress conditions. Hence, lncRNA-enriched EVs are readily being reported as biomarkers [54]. The active process of lncRNA sorting into exosomes can be facilitated by RNA-binding proteins (RBPs) with a growing understanding of their mechanism of loading and action involving epigenetic modifications, through which exosomal lncRNAs can alleviate the progression of some diseases [55,56].
Further examples of lncRNA-mediated epigenetic regulation in vascular cells, including those interacting with one of the major transcriptional repressors, polycomb repressive complex 2 (PRC2), are discussed below.
4. Polycomb Repressive Complex 2 and lncRNAs
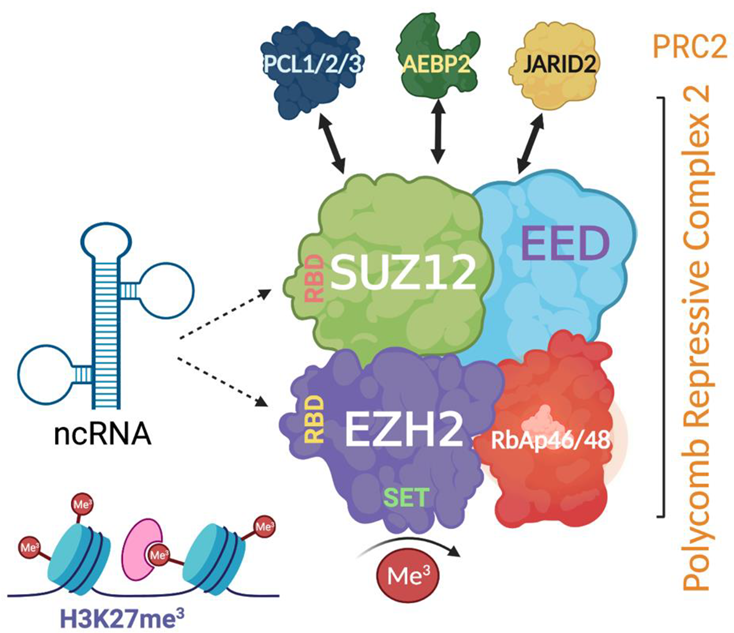
The polycomb repressive complex 2 (PRC2) is a major histone methyltransferase in vascular biology. Through the deposition of histone H3 lysine K27 trimethylation (H3K27me3) modification, PRC2 compacts chromatin and represses gene expression. PRC2 comprises four major subunits including EZH2 (enhancer of zeste homolog 2) which possesses the catalytic histone methyltransferase SET domain; EED (embryonic ectoderm development) which binds the H3K27me3 mark and is responsible for its propagation; SUZ12 (suppressor of zeste 12 homolog); and RbAp46/48 (retinoblastoma protein associated protein 46/48) (Figure 3) [57]. The mechanisms behind PRC2 engagement with native chromatin, as well as its recruitment in a context-dependent manner, remain to be fully understood [58]. EZH2-mediated epigenomic landscapes guide cardiovascular and endothelial differentiation and maturation, maintain tissue-specific genetic blueprints established during development, and are implicated in cardiovascular disease [55,57,58]. Moreover, PRC2 histone methyltransferase activity appears to be a key regulator of angiogenesis. An increased EZH2 expression negatively regulates the revascularisation response to hypoxia through the deposition of H3K27me3 onto pro-angiogenic genes such as eNOS, thus worsening outcomes in ischaemia, and similarly halts the endothelial lineage commitment in pro-angiogenic haematopoietic and endothelial progenitor cells [20,48,59].
It is now recognised that in addition to its chromatin binding capacity, PRC2 displays a tissue-specific lncRNA-binding capacity, including an ability to bind lncRNAs involved in vascular endothelial homeostasis such as MEG3—elucidating a model for PRC2 recruitment to specific loci. Both EZH2 and SUZ12 subunits bind lncRNA through conserved sequences and interact with chromatin in a mutually antagonistic fashion, with subsequent lncRNA-mediated target gene recognition through triplex formation—representing a general mechanism to guide transcriptional regulators [59]. As well as serving as a scaffold to mediate H3K27me3 deposition on chromatin, motif-driven RNA binding may fine-tune transcriptional repression by recruiting PRC2 to stall the activity of RNA polymerase-II [60].
5. Multi-Omics Approaches to Detect Long Non-Coding RNAs in Vascular Cells
The use of novel technologies has immensely improved the detection of lncRNAs and the identification of their in vivo targets [61]. Chromatin immunoprecipitation (ChIP) and assay for transposon-accessible chromatin (ATAC-seq) allow the mapping of protein-DNA interaction in vivo on a genome scale [62]. RNA immunoprecipitation (RIP), chromatin isolation by RNA purification (ChIRP), individual-nucleotide-resolution UV cross-linking and immunoprecipitation (iCLIP), and cross-linking, ligation and sequencing of hybrids (CLASH) use glutaraldehyde-, formaldehyde- and UV-mediated cross-linking approaches in order to preserve RNA–RNA and RNA–protein interactions and to capture specific lncRNAs and their distribution between intracellular fractions using biotinylated antisense DNA probes [63,64,65].
In Table 1, we have outlined the experimental approaches used to delineate epigenetic functions of some nuclear lncRNAs in a vascular setting, whilst Table 2 reports the experimental use of these techniques in the vascular field between 2018 and present. The techniques for mapping the RNA interactome have also been comprehensively reviewed recently (see reference [33]).
5.1. MEG3
Maternally expressed gene 3 (MEG3) is a major hypoxia-sensitive lncRNA which interacts with PRC2 and has complex roles in EC function. Hypoxia-inducible factor 1α (HIF1α) upregulates the MEG3 expression in response to hypoxia [66] and under hypoxic conditions, MEG3 has been observed to translocate to the cytoplasm [38]. MEG3 contains a 356-nucleotide element which associates with U1 small nuclear ribonucleoprotein particles (snRNP) and is responsible for its nuclear retention [67]. MEG3 is a negative regulator of angiogenesis in hypoxia, with MEG3 knock-out mice showing enhanced angiogenesis in the brain and the augmented expression of pro-angiogenic genes [68]. MEG3 is also, however, required for VEGF-induced EC migration and angiogenesis in human umbilical vein endothelial cells (HUVECs) [64]. It has been proposed that MEG3 both negatively regulates pro-angiogenic genes and contributes to age-associated endothelial dysfunction through its interaction with EZH2 or JARID2 and the recruitment of PRC2 to epigenetically regulate chromatin—yet these mechanisms are yet to be fully elucidated [24]. Indeed, it is known that MEG3 modulates the activity of genes in the transforming growth factor-b (TGF-b) pathway through its interaction with PRC2 and formation of RNA–DNA triplexes [26].
5.2. MALAT1
LncRNA-metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) has been implicated in the development of diabetic nephropathy through its epigenetic regulation of renal glomerular EC (HRGEC) gene expression [27]. It was demonstrated that MALAT1 was upregulated in patients with diabetic nephropathy and inversely correlated with the expression of klotho—a gene known to improve the function of hyperglycaemia (HG)-induced glomerular EC injury. Authors used RIP and ChIP approaches in HRGECs following HG-induced injury to define the underlying mechanism. MALAT1 mediates HG-induced HRGEC injury through the binding and recruitment of G9a methyltransferase to deposit the H3K9me1 repressive signature to epigenetically suppress klotho expression. Such research not only demonstrates the role of MALAT1 in devising the chromatin status of klotho but implies the prospect of targeting MALAT1 to ameliorate glomerular EC function in hyperglycaemia to impede the progression of diabetic nephropathy.
Epigenetic pathways regulated by MALAT1 are also implicated in the development of thoracic aortic aneurysm (TAA) and stenotic disease. The dysfunction of the vascular smooth muscle cell (VSMC) through deficiencies in contractile proteins predisposes to the pathogenesis of these conditions. Histone deacetylase complex, HDAC9, is known to recruit EZH2 to epigenetically modify chromatin through the deposition of repressive H3K27me3. EZH2 was identified as a crucial player in TAA development, with its inhibition-enhancing expression of contractile protein genes such as SM22a and improving aortic performance in mice [29]. Moreover, pathological recruitment of a nuclear HDAC9–MALAT1–BRG1 (brahma-related gene-1) complex to deposit H3K27me3 on gene promoters and downregulate contractile protein gene expression has been highlighted as a pathway implicated in VSMC dysfunction in TAA. Indeed, Malat1 or Hdac9 disruption restored a contractile phenotype, ameliorated aortic wall architecture, and prevented experimental TAA growth [28]. MALAT1-mediated nuclear targeting of HDAC9 to transcriptionally silence genes encoding contractile proteins also mediates stenotic vascular disease, with MALAT1 or HDAC9 deficiency or EZH2 inhibition attenuating neointimal formation and preserving the beneficial VSMC contractile phenotypes [29].
5.3. NEAT1
Although nuclear enriched abundant transcript 1 (NEAT1) is nuclear lncRNA, the findings have shown that its localisation is dispersed within the nucleus and is necessary for the formation of paraspeckles [69]. Ahmed et al. demonstrated that lncRNA-NEAT1 expression was upregulated in VSMCs in response to vascular injury to promote a proliferative phenotype and blood vessel repair [22]. NEAT1 silencing in human coronary artery smooth muscle cells (HCASMCs) increased SM-specific gene expression and reduced VSMC proliferation and migration; whilst NEAT1 knockout (KO) mice displayed decreased neointima formation and vascular repair. Moreover, ChIP assays showed that NEAT1 KO in HCASMCs enriched active chromatin marks (H3K4me3 and H3K9ac) and depleted inactive modifications [22]. Mechanistically, RIP showed binding of NEAT1 to the WDR5 chromatin modifier which has a scaffolding role for H3K4 methyltransferase complexes to deposit the H3K4 mark and contribute to an open chromatin state. Upon stimulation of a proliferative, anti-contractile phenotype, the NEAT1 binding to WDR5 was enhanced. Overall, it appears that NEAT1 operates by a mechanism of sequestering WDR5 from SM contractile genes to initiate epigenetic repression of SM contractile gene expression and modulate phenotypic switching towards a proliferative phenotype.
5.4. MANTIS
MANTIS is an intronic lncRNA-located antisense to Annexin A4 (ANXA4) that acts as an important regulator of angiogenic sprouting and alignment of ECs [23]. The downregulation of MANTIS expression, controlled by histone demethylase JARID1B, was observed in the lungs of patients with idiopathic pulmonary arterial hypertension, and the upregulation was observed in carotid arteries of Macaca fascicularis under an atherosclerosis regression diet, and in endothelial cells isolated from human glioblastoma patients. Knocking out MANTIS with LNA-GapmeRs led to the downregulation of key endothelial genes important for angiogenesis including SOX18, SMAD6, and COUP-TFII, and the upregulation of stress-induced genes contributory to atherosclerosis such as interleukin 6 (IL-6) and superoxide dismutase 2 (SOD2). Genes in proximity to the MANTIS locus in the genome were not influenced by the MANTIS knockout, suggesting that it acts in a trans- rather than cis-regulatory manner. Mechanistically, MANTIS appears to influence epigenetic gene regulation through acting via the BRG1 component of the chromatin remodelling complex, SWI/SNF (SWItch/Sucrose non-fermentable), to facilitate access by RNA polymerase II machinery to the specific genes which counteract atherosclerotic processes [23].
MANTIS appears to further impede atherosclerosis development by limiting pro-inflammatory intercellular adhesion molecule 1 (ICAM-1)-mediated endothelial monocyte adhesion by diminishing binding of BRG1 at the ICAM-1 promoter to repress its transcription. Indeed, MANTIS silencing has been shown to increase endothelial monocyte adhesion via ICAM-1. Moreover, statins and laminar flow ameliorate atherosclerotic vascular disease by inducing MANTIS expression through epigenetic mechanisms and Krüppel-like factor (KLF) 2 and 4 transcription factors [70]. Since MANTIS appears to both limit atherosclerosis development in a multitude of ways and mediate the pleiotropic effects of statins, it may be an effective therapeutic target for ameliorating vascular disease.
5.5. LEENE
LncRNA that enhances eNOS expression (LEENE) is an enhancer-associated RNA that plays an important role in regulating eNOS expression and endothelial barrier function [21,34]. Like many athero-protective genes, LEENE expression is regulated by two transcription factors, KLF2 and KLF4. Miao et al. used the chromatin conformation capture methods to demonstrate that LEENE targets the distal enhancer region located near the eNOS promoter in ECs, acting as a facilitator of RNA polymerase II recruitment to the eNOS promoter to enhance its transcription. LEENE RNA alone was not sufficient to enhance the eNOS transcription, yet genomic or transcriptional inhibition of the LEENE enhancer region suppressed the eNOS transcription and increased the transcription of pro-inflammatory and pro-atherosclerotic ICAM1 and VCAM1 [21]. Moreover, a cytoplasmic function has been described for LEENE (linc00520) comprising the regulation of pathways central for VEGF signalling and cell adhesion during the endothelial response to shear stress, resulting in its additional name, shear stress-induced long non-coding RNA (LASSIE) [71]. LASSIE associates with junctional proteins, such as PECAM-1, and the intermediate filament protein nestin, serving as a link between the cytoskeleton and adherent junctions to regulate EC alignment and vascular barrier function [71].
5.6. GATA6-AS
It has previously been shown that HIF1α-dependent signalling pathways promote the hypoxia-induced, pathology-associated cellular process of endothelial–mesenchymal transition (endMT) through mediating epigenetic changes via methyltransferases in endothelial cells [10]. Since a plethora of lncRNAs are known to respond to HIF [66] and together regulate methylase enzymes [72,73], it is unsurprising that further studies have illuminated a role for lncRNAs in the regulation of endMT. A study by Neumann et al. recently characterised the participation of the nuclear-enriched, hypoxia-regulated lncRNA antisense transcript of GATA6 (GATA6-AS) in the epigenetic regulation of endothelial gene expression underlying endMT [30]. Using an RNA deep sequencing approach, the authors identified that lncRNA GATA6-AS was upregulated under hypoxia and regulated endMT as well as sprouting angiogenesis and endothelial cell migration. GATA6-AS silencing impaired the endMT in vitro and augmented angiogenesis in vivo in a HUVEC-based xenograft model. Mechanistically, using the RIP and iCLIP techniques in HUVECs, the authors demonstrated that GATA6-AS associated with the LOXL2 deaminase enzyme, known to remove activating H3K4me3 marks, regulates the angiogenesis-related genes periostin and cyclooxygenase-1. Since GATA6-AS silencing reduced H3K4me3 of genes, this suggested that lncRNA GATA6-AS negatively regulates LOXL2 the demethylase function—promoting H3K4me3 mark deposition and the activation of angiogenesis-related genes.
5.7. ANRIL
Studies are increasingly revealing the implication of lncRNA-mediated histone methylation in the progression of atherosclerosis. LncRNA-ANRIL (antisense ncRNA in the INK4 locus), otherwise known as cyclin-dependent kinase inhibitor 2B antisense transcript 1 (CDKN2B-AS1), is known to contribute to atherosclerotic disease states including stroke, coronary heart disease, and peripheral vascular disease through epigenetically regulating the endothelial inflammatory response as well as cell proliferation, adhesion, and apoptosis [31]. ANRIL promotes plaque formation due to its epigenetic regulation of chromatin regions in cis by recruitment of PRC1 and PRC2, and its ability to bind specific sequences on regulators such as CCCTC-binding factor (CTCF) to regulate distant genes in trans [31]. For example, ANRIL has been found to contribute to the formation of atherosclerotic plaques through epigenetically regulating the CDKN2B promoter [74]. ANRIL expression was found to be upregulated in atherosclerotic compared to non-atherosclerotic tissues and in macrophage-derived foam cells. In contrast, CDKN2B was down regulated. To elucidate the molecular mechanism of ANRIL, authors used a variety of techniques. RNA–DNA pull down and capture was used to identify RNA–DNA triplexes between nuclear-localised ANRIL and the CDKN2B promoter. RIP and ChIP approaches were used to determine interactions between ANRIL, CCCTC-binding factor (CTCF), EZH2, and CDKN2B. It was found that ANRIL regulated CDKN2B transcriptional repression through EZH2 recruitment and H3K27me3 deposition onto the CDKN2B promoter, under the regulation of CTCF. ANRIL silencing attenuated atherosclerosis in macrophage-derived foam cells and in ApoE−/− mouse models of atherosclerosis and promoted the reversal by macrophage reverse cholesterol transport (mRCT)—suggesting a potential therapeutic modality in targeting ANRIL for treating atherosclerosis.
6. Application and RNA Therapeutics
Our evolving understanding of lncRNAs and their role in epigenetic regulation of vascular cells may have a profound translational impact. Epigenetic therapies have already shown therapeutic potential in the context of cancer and PRC2 inhibitor agents such as EED226, EPZ, CPI-1205, and GSK126, which have also shown effectiveness in other settings. EZH2 inhibition led to the increased expression of pro-angiogenic genes and enhanced angiogenic functions and EC proliferation in vitro, whilst promoting blood flow recovery in mouse ischaemic muscles in vivo [51]. In a mouse model of TAA, EZH2 inhibition improved the outcomes due to the augmented expression of pathologically inactivated genes encoding contractile proteins [29]. The inhibition of EZH2 can also facilitate the activation of cardiac genes, indicating a promising way to generate or reprogramme human induced cardiomyocyte-like cells (hiCMs) [75].
Previously, when designing such PRC2 inhibitors, the RNA binding capacity of PRC2 was not taken into consideration. Disruption of lncRNA-mediated PRC2 recruitment has, however, been shown to reduce the severity of spinal muscular atrophy due to reduced H3K27me3 occupancy at the SMN gene, increasing its expression [76]. Moreover, another study indicated the use of a selective compound to manipulate the interaction between lncRNA-HOTAIR and EZH2 to mediate the downstream epigenetic mechanisms in the context of cancer therapy [77]. New therapeutic opportunities are therefore arising to block PRC2 recruitment effectively by dissociating lncRNAs using targeted therapy. With this novel class of pharmacological PRC2 inhibitors there is a potential to study the lncRNA epigenetic modulation of vascular gene expression and assess the therapeutic effectiveness of selective interference with the lncRNA-PRC2/EZH2 interaction [78].
Publications are increasingly noting the potential for lncRNA therapeutics in cardiovascular disease, with multiple vascular lncRNAs being identified as promising targets for the inhibition by antisense oligonucleotides and short hairpin RNAs in the treatment of atherosclerotic disease and in the induction of angiogenesis and vascular remodelling and repair [79,80]. In vivo GapmeR-mediated inhibition of MEG3, for example, reduced cardiac fibrosis, indicating its potential as a novel target for the prevention of cardiac remodelling, whilst a similar inhibition of MALAT1 attenuated the endothelial revascularisation response following ischaemia [25,80]. HOTAIR, guided by the transcriptional factor SNAIL, acts as a site-specific scaffold for PRC2 to mediate the repression of epithelial genes. Garbo et al. effectively counteracted HOTAIR function in the context of hepatocellular carcinoma cells with few off-target effects to impair the epithelial to mesenchymal transition (EMT) responsible for driving vascular inflammation [81]. Such research, in combination with our growing understanding of lncRNA-mediated epigenetic mechanisms in vascular cells, implies great potential for lncRNAs as targets to mitigate endothelial or vascular smooth muscle cell dysfunction, initiate therapeutic angiogenesis and vascular regeneration, and ultimately treat ischaemic cardiovascular diseases.
RNA molecules are an attractive therapeutic concept due to their low immunogenicity, effective cell and nuclear penetration, low cost to manufacture, and ease of chemical modification [81]. Multiple challenges in the development, however, have been identified, which may limit the clinical application of RNA therapeutics. These include the difficulty in delivering high enough doses of inhibitors, including GapmeRs and siRNAs without the risks of dose-dependent toxicity such as hepatotoxicity, and the difficulty in targeting less accessible molecules predominantly localised to the nucleus [80]. Nevertheless, novel techniques to enable efficient delivery and tissue-specific enrichment of lncRNA-targeting molecules have been proposed, such as the use of viral vectors, exosomes, nanoparticles, cell-penetrating peptides, antibodies, and engineered extracellular vesicles [79,80,81,82]. Making use of on-target sequence specificity to enhance the efficient ncRNA delivery and reduce the unwanted off-target side effects has also been proposed [82], with the reported tools paving the way for disruption of lncRNAs with minimal off-target effects [81]. Moreover, poor evolutionary conservation of lncRNA represents an additional difficulty in their therapeutic development, requiring the critical evaluation of animal models used to ensure efficacy, and to avoid undesirable adverse effects [80].
7. Future Perspective and Directions
Thus far, the advancements in research methods have significantly contributed to our understanding of lncRNA actions within the vasculature. Increasingly effective and sophisticated mechanistic studies are providing insight into the molecular mechanisms by which lncRNAs epigenetically regulate endothelial and VSMC gene expression in vascular homeostasis, ultimately enabling a more profound understanding of the pathological mechanisms involved in vascular disease and elucidating the potential therapeutic targets. The modulation of gene expression via targeting the epigenetic pathways implicated in angiogenesis, endothelial dysfunction, and vascular disease is a novel therapeutic avenue to explore in the future—however, greater characterisation of lncRNA interactions with the vascular gene promoters and epigenetic regulators such as PRC2 are required. Further advancements in the field largely depend on a better understanding of the structural organisation of lncRNAs and structure-to-function relations. Due to the poor conservation across species, the currently available tools for functional prediction based on a primary sequence similarity or structural predictions are not of great use [11]. However, the use of CRISPR (clustered regularly interspaced short palindromic repeats)-mediated gene editing to introduce targeted modifications into lncRNAs, together with novel technologies such as genome-wide chromatin interrogation, locked nucleic acids, bridged nucleic acids, and novel approaches to RNA purification in vitro and in vivo, will provide a more comprehensive understanding of the lncRNA structural domains, their folding patterns, and undiscovered mechanisms of action.
Author Contributions
Conceptualization and resources, T.M. and A.C.; data curation, A.N.; writing—original draft preparation, H.G.; writing—review and editing, H.G., M.S., T.M. and R.A.; visualization, supervision, project administration and funding acquisition, T.M. All authors have read and agreed to the published version of the manuscript.
Funding
Tijana Mitic is supported by the British Heart Foundation (BHF) Career Re-entry Fellowship (FS/16/38/32351).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
No conflict of interest for all the authors.
References
- Garg, A.; Gupta, S.K.; Thum, T. Long non-coding RNAs: A crucial part of the vasculature puzzle. Vasc. Pharmacol. 2019, 114, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Jambusaria, A.; Hong, Z.; Zhang, L.; Srivastava, S.; Jana, A.; Toth, P.; Dai, Y.; Malik, A.B.; Rehman, J. Endothelial heterogeneity across distinct vascular beds during homeostasis and inflammation. eLife 2020, 9, 9. [Google Scholar] [CrossRef] [PubMed]
- Kok, F.O.; Baker, A.H. The function of long non-coding RNAs in vascular biology and disease. Vasc. Pharmacol. 2019, 114, 23–30. [Google Scholar] [CrossRef]
- Aryal, B.; Suárez, Y. Non-coding RNA regulation of endothelial and macrophage functions during atherosclerosis. Vasc. Pharmacol. 2019, 114, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Wang, S. Angio-LncRs: LncRNAs that regulate angiogenesis and vascular disease. Theranostics 2018, 8, 3654–3675. [Google Scholar] [CrossRef] [PubMed]
- Rosa-Garrido, M.; Chapski, D.; Vondriska, T.M. Epigenomes in Cardiovascular Disease. Circ. Res. 2018, 122, 1586–1607. [Google Scholar] [CrossRef] [PubMed]
- Annex, B.H. Therapeutic angiogenesis for critical limb ischaemia. Nat. Rev. Cardiol. 2013, 10, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Jaé, N.; Heumüller, A.W.; Fouani, Y.; Dimmeler, S. Long non-coding RNAs in vascular biology and disease. Vasc. Pharmacol. 2019, 114, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Fang, S.; Zhang, L.; Guo, J.; Niu, Y.; Wu, Y.; Li, H.; Zhao, L.; Li, X.; Teng, X.; Sun, X.; et al. NONCODEV5: A comprehensive annotation database for long non-coding RNAs. Nucleic Acids Res. 2018, 46, D308–D314. [Google Scholar] [CrossRef]
- Zhang, X.; Li, D.Y.; Reilly, M.P. Long intergenic noncoding RNAs in cardiovascular diseases: Challenges and strategies for physiological studies and translation. Atherosclerosis 2019, 281, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Graf, J.; Kretz, M. From structure to function: Route to understanding lncRNA mechanism. BioEssays 2020, 42, e2000027. [Google Scholar] [CrossRef] [PubMed]
- Ulitsky, I. Evolution to the rescue: Using comparative genomics to understand long non-coding RNAs. Nat. Rev. Genet. 2016, 17, 601–614. [Google Scholar] [CrossRef]
- Zampetaki, A.; Albrecht, A.; Steinhofel, K. Long Non-coding RNA Structure and Function: Is There a Link? Front. Physiol. 2018, 9, 1201. [Google Scholar] [CrossRef]
- Uroda, T.; Anastasakou, E.; Rossi, A.; Teulon, J.-M.; Pellequer, J.-L.; Annibale, P.; Pessey, O.; Inga, A.; Chillón, I.; Marcia, M. Conserved Pseudoknots in lncRNA MEG3 Are Essential for Stimulation of the p53 Pathway. Mol. Cell 2019, 75, 982–995.e9. [Google Scholar] [CrossRef] [Green Version]
- Sherpa, C.; Rausch, J.W.; Le Grice, S.F.J. Structural characterization of maternally expressed gene 3 RNA reveals conserved motifs and potential sites of interaction with polycomb repressive complex 2. Nucleic Acids Res. 2018, 46, 10432–10447. [Google Scholar] [CrossRef] [Green Version]
- Chillón, I.; Marcia, M. The molecular structure of long non-coding RNAs: Emerging patterns and functional implications. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 662–690. [Google Scholar] [CrossRef]
- Cho, H.; Li, Y.; Archacki, S.; Wang, F.; Yu, G.; Chakrabarti, S.; Guo, Y.; Chen, Q.; Wang, Q.K. Splice variants of lncRNA RNA ANRIL exert opposing effects on endothelial cell activities associated with coronary artery disease. RNA Biol. 2020, 17, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- Man, H.-S.J.; A Marsden, P. LncRNAs and epigenetic regulation of vascular endothelium: Genome positioning system and regulators of chromatin modifiers. Curr. Opin. Pharmacol. 2019, 45, 72–80. [Google Scholar] [CrossRef]
- Ballantyne, M.D.; Pinel, K.; Dakin, R.; Vesey, A.T.; Diver, L.; MacKenzie, R.; Garcia, R.; Welsh, P.; Sattar, N.; Hamilton, G.; et al. Smooth Muscle Enriched Long Noncoding RNA (SMILR) Regulates Cell Proliferation. Circulation 2016, 133, 2050–2065. [Google Scholar] [CrossRef]
- Li, Y.; Maegdefessel, L. Non-coding RNA Contribution to Thoracic and Abdominal Aortic Aneurysm Disease Development and Progression. Front. Physiol. 2017, 8, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Y.; Ajami, N.E.; Huang, T.-S.; Lin, F.-M.; Lou, C.-H.; Wang, Y.-T.; Li, S.; Kang, J.; Munkacsi, H.; Maurya, M.R.; et al. Enhancer-associated long non-coding RNA LEENE regulates endothelial nitric oxide synthase and endothelial function. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Ahmed, A.S.I.; Dong, K.; Liu, J.; Wen, T.; Yu, L.; Xu, F.; Kang, X.; Osman, I.; Hu, G.; Bunting, K.M.; et al. Long noncoding RNA NEAT1 (nuclear paraspeckle assembly transcript 1) is critical for phenotypic switching of vascular smooth muscle cells. Proc. Natl. Acad. Sci. USA 2018, 115, E8660–E8667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leisegang, M.S.; Fork, C.; Josipovic, I.; Richter, F.M.; Preussner, J.; Hu, J.; Miller, M.J.; Epah, J.; Hofmann, P.; Günther, S.; et al. Long Noncoding RNA MANTIS Facilitates Endothelial Angiogenic Function. Circulation 2017, 136, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Boon, R.; Hofmann, P.; Michalik, K.M.; Lozano-Vidal, N.; Berghäuser, D.; Fischer, A.; Knau, A.; Jaé, N.; Schürmann, C.; Dimmeler, S. Long Noncoding RNA Meg3 Controls Endothelial Cell Aging and Function. J. Am. Coll. Cardiol. 2016, 68, 2589–2591. [Google Scholar] [CrossRef]
- Michalik, K.M.; You, X.; Manavski, Y.; Doddaballapur, A.; Zörnig, M.; Braun, T.; John, D.; Ponomareva, Y.; Chen, W.; Uchida, S.; et al. Long Noncoding RNA MALAT1 Regulates Endothelial Cell Function and Vessel Growth. Circ. Res. 2014, 114, 1389–1397. [Google Scholar] [CrossRef] [PubMed]
- Mondal, T.; Subhash, S.; Vaid, R.; Enroth, S.; Uday, S.; Reinius, B.; Mitra, S.; Mohammed, A.; James, A.R.; Hoberg, E.; et al. MEG3 long noncoding RNA regulates the TGF-β pathway genes through formation of RNA–DNA triplex structures. Nat. Commun. 2015, 6, 7743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Ren, D.; Xu, G. Long noncoding RNA MALAT1 mediates high glucose-induced glomerular endothelial cell injury by epigenetically inhibiting klotho via methyltransferase G9a. IUBMB Life 2019, 71, 873–881. [Google Scholar] [CrossRef]
- Cardenas, C.L.L.; Kessinger, C.W.; Cheng, Y.; Macdonald, C.; MacGillivray, T.; Ghoshhajra, B.; Huleihel, L.; Nuri, S.; Yeri, A.S.; Jaffer, F.A.; et al. An HDAC9-MALAT1-BRG1 complex mediates smooth muscle dysfunction in thoracic aortic aneurysm. Nat. Commun. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Cardenas, C.L.; Kessinger, C.W.; MacDonald, C.; Jassar, A.S.; Isselbacher, E.M.; Jaffer, F.A.; Lindsay, M.E. Inhibition of the methyltranferase EZH2 improves aortic performance in experimental thoracic aortic aneurysm. JCI Insight 2018, 3, 97493. [Google Scholar] [CrossRef] [Green Version]
- Neumann, P.; Jaé, N.; Knau, A.; Glaser, S.F.; Fouani, Y.; Rossbach, O.; Krüger, M.; John, D.; Bindereif, A.; Grote, P.; et al. The lncRNA GATA6-AS epigenetically regulates endothelial gene expression via interaction with LOXL2. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chi, J.-S.; Li, J.-Z.; Jia, J.-J.; Zhang, T.; Liu, X.-M.; Yi, L. Long non-coding RNA ANRIL in gene regulation and its duality in atherosclerosis. Curr. Med. Sci. 2017, 37, 816–822. [Google Scholar] [CrossRef]
- Zuckerman, B.; Ulitsky, I. Predictive models of subcellular localization of long RNAs. RNA 2019, 25, 557–572. [Google Scholar] [CrossRef]
- Guh, C.-Y.; Hsieh, Y.-H.; Chu, H.-P. Functions and properties of nuclear lncRNAs-from systematically mapping the interactomes of lncRNAs. J. Biomed. Sci. 2020, 27, 44. [Google Scholar] [CrossRef]
- Zhang, C.; Han, B.; Xu, T.; Li, D. The biological function and potential mechanism of long non-coding RNAs in cardiovascular disease. J. Cell. Mol. Med. 2020, 24, 12900–12909. [Google Scholar] [CrossRef]
- Engreitz, J.M.; Haines, J.E.; Perez, E.; Munson, G.; Chen, J.; Kane, M.; McDonel, P.E.; Guttman, M.; Lander, E.S. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nat. Cell Biol. 2016, 539, 452–455. [Google Scholar] [CrossRef]
- Noh, J.H.; Kim, K.M.; McClusky, W.G.; Abdelmohsen, K.; Gorospe, M. Cytoplasmic functions of long noncoding RNAs. Wiley Interdiscip. Rev. RNA 2018, 9, e1471. [Google Scholar] [CrossRef]
- Zhang, C.; Niu, K.; Lian, P.; Hu, Y.; Shuai, Z.; Gao, S.; Ge, S.; Xu, T.; Xiao, Q.; Chen, Z. Pathological Bases and Clinical Application of Long Noncoding RNAs in Cardiovascular Diseases. Hypertension 2021, 78, 16–29. [Google Scholar] [CrossRef]
- Xing, Y.; Zheng, X.; Fu, Y.; Qi, J.; Li, M.; Ma, M.; Wang, S.; Li, S.; Zhu, D. Long Noncoding RNA-Maternally Expressed Gene 3 Contributes to Hypoxic Pulmonary Hypertension. Mol. Ther. 2019, 27, 2166–2181. [Google Scholar] [CrossRef]
- Martignano, F.; Rossi, L.; Maugeri, A.; Gallà, V.; Conteduca, V.; De Giorgi, U.; Casadio, V.; Schepisi, G. Urinary RNA-based biomarkers for prostate cancer detection. Clin. Chim. Acta 2017, 473, 96–105. [Google Scholar] [CrossRef]
- De Gonzalo-Calvo, D.; Kenneweg, F.; Bang, C.; Toro, R.; Van Der Meer, R.W.; Rijzewijk, L.J.; Smit, J.W.; Lamb, H.J.; Llorente-Cortes, V.; Thum, T. Circulating long-non coding RNAs as biomarkers of left ventricular diastolic function and remodelling in patients with well-controlled type 2 diabetes. Sci. Rep. 2016, 6, 37354. [Google Scholar] [CrossRef]
- Sartori, D.A.; Chan, D.W. Biomarkers in prostate cancer. Curr. Opin. Oncol. 2014, 26, 259–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumarswamy, R.; Bauters, C.; Volkmann, I.; Maury, F.; Fetisch, J.; Holzmann, A.; Lemesle, G.; de Groote, P.; Pinet, F.; Thum, T. Circulating Long Noncoding RNA, LIPCAR, Predicts Survival in Patients With Heart Failure. Circ. Res. 2014, 114, 1569–1575. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Zhao, D.; Dong, P.; Wang, H.; Li, D.; Lai, L. Effects of ANRIL polymorphisms on the likelihood of coronary artery disease: A meta-analysis. J. Cell. Biochem. 2019, 120, 6113–6119. [Google Scholar] [CrossRef] [PubMed]
- Vanhaverbeke, M.; Vausort, M.; Veltman, D.; Zhang, L.; Wu, M.; Laenen, G.; Gillijns, H.; Moreau, Y.; Bartunek, J.; Van De Werf, F.; et al. Peripheral Blood RNA Levels of QSOX1 and PLBD1 Are New Independent Predictors of Left Ventricular Dysfunction After Acute Myocardial Infarction. Circ. Genom. Precis. Med. 2019, 12, e002656. [Google Scholar] [CrossRef] [PubMed]
- Calviello, L.; Mukherjee, N.; Wyler, E.; Zauber, H.; Hirsekorn, A.; Selbach, M.; Landthaler, M.; Obermayer, B.; Ohler, U. Detecting actively translated open reading frames in ribosome profiling data. Nat. Methods 2015, 13, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Hartford, C.C.R.; Lal, A. When Long Noncoding Becomes Protein Coding. Mol. Cell. Biol. 2020, 40, e00528-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Tabula Muris Consortium; Overall Coordination; Logistical Coordination; Organ Collection and Processing; Library Preparation and Sequencing; Computational Data Analysis; Cell Type Annotation; Writing Group; Supplemental Text Writing Group; Principal Investigators. Single-cell transcriptomics of 20 mouse organs creates a Tabula Muris. Nature 2018, 562, 367–372. [Google Scholar] [CrossRef]
- Muhl, L.; Genové, G.; Leptidis, S.; Liu, J.; He, L.; Mocci, G.; Sun, Y.; Gustafsson, S.; Buyandelger, B.; Chivukula, I.V.; et al. Single-cell analysis uncovers fibroblast heterogeneity and criteria for fibroblast and mural cell identification and discrimination. Nat. Commun. 2020, 11, 1–18. [Google Scholar] [CrossRef]
- van Heesch, S.; Witte, F.; Schneider-Lunitz, V.; Schulz, J.F.; Adami, E.; Faber, A.B.; Kirchner, M.; Maatz, H.; Blachut, S.; Sandmann, C.-L.; et al. The Translational Landscape of the Human Heart. Cell 2019, 178, 242–260.e29. [Google Scholar] [CrossRef] [Green Version]
- Shen, Z.; Zeng, L.; Zhang, Z. Translatome and Transcriptome Profiling of Hypoxic-Induced Rat Cardiomyocytes. Mol. Ther. -Nucleic Acids 2020, 22, 1016–1024. [Google Scholar] [CrossRef]
- Mitić, T.; Caporali, A.; Floris, I.; Meloni, M.; Marchetti, M.; Urrutia, R.; Angelini, G.; Emanueli, C. EZH2 Modulates Angiogenesis In Vitro and in a Mouse Model of Limb Ischemia. Mol. Ther. 2015, 23, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Batie, M.; Frost, J.; Frost, M.; Wilson, J.W.; Schofield, P.; Rocha, S. Hypoxia induces rapid changes to histone methylation and reprograms chromatin. Science 2019, 363, 1222–1226. [Google Scholar] [CrossRef]
- Chakraborty, A.A.; Laukka, T.; Myllykoski, M.; Ringel, A.E.; Booker, M.A.; Tolstorukov, M.Y.; Meng, Y.J.; Meier, S.R.; Jennings, R.B.; Creech, A.L.; et al. Histone demethylase KDM6A directly senses oxygen to control chromatin and cell fate. Science 2019, 363, 1217–1222. [Google Scholar] [CrossRef] [Green Version]
- Kenneweg, F.; Bang, C.; Xiao, K.; Boulanger, C.M.; Loyer, X.; Mazlan, S.; Schroen, B.; Hermans-Beijnsberger, S.; Foinquinos, A.; Hirt, M.N.; et al. Long Noncoding RNA-Enriched Vesicles Secreted by Hypoxic Cardiomyocytes Drive Cardiac Fibrosis. Mol. Ther. -Nucleic Acids 2019, 18, 363–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.; Huang, W. New Developments in Exosomal lncRNAs in Cardiovascular Diseases. Front. Cardiovasc. Med. 2021, 8, 709169. [Google Scholar] [CrossRef] [PubMed]
- Fabbiano, F.; Corsi, J.; Gurrieri, E.; Trevisan, C.; Notarangelo, M.; D’Agostino, V.G. RNA packaging into extracellular vesicles: An orchestra of RNA-binding proteins? J. Extracell. Vesicles 2020, 10, 12043. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, G.G. No Easy Way Out for EZH2: Its Pleiotropic, Noncanonical Effects on Gene Regulation and Cellular Function. Int. J. Mol. Sci. 2020, 21, 9501. [Google Scholar] [CrossRef]
- Yuan, J.-L.; Yin, C.-Y.; Li, Y.-Z.; Song, S.; Fang, G.-J.; Wang, Q.-S. EZH2 as an Epigenetic Regulator of Cardiovascular Development and Diseases. J. Cardiovasc. Pharmacol. 2021, 78, 192–201. [Google Scholar] [CrossRef]
- Beltran, M.; Yates, C.M.; Skalska, L.; Dawson, M.; Reis, F.P.; Viiri, K.; Fisher, C.; Sibley, C.; Foster, B.; Bartke, T.; et al. The interaction of PRC2 with RNA or chromatin is mutually antagonistic. Genome Res. 2016, 26, 896–907. [Google Scholar] [CrossRef]
- Rosenberg, M.; Blum, R.; Kesner, B.; Aeby, E.; Garant, J.-M.; Szanto, A.; Lee, J.T. Motif-driven interactions between RNA and PRC2 are rheostats that regulate transcription elongation. Nat. Struct. Mol. Biol. 2021, 28, 103–117. [Google Scholar] [CrossRef]
- Machyna, M.; Simon, M.D. Catching RNAs on chromatin using hybridization capture methods. Brief. Funct. Genom. 2018, 17, 96–103. [Google Scholar] [CrossRef]
- Sahu, S.K.; Basu, A.; Tiwari, V.K. Measuring Chromatin Accessibility. Rab GTPases 2021, 105–121. [Google Scholar]
- Ramanathan, M.; Porter, D.F.; Khavari, P.A. Methods to study RNA—Protein interactions. Nat. Methods 2019, 16, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Zhang, Q.C.; da Rocha, S.T.; Flynn, R.A.; Bharadwaj, M.; Calabrese, J.M.; Magnuson, T.; Heard, E.; Chang, H.Y. Systematic Discovery of Xist RNA Binding Proteins. Cell 2015, 161, 404–416. [Google Scholar] [CrossRef] [Green Version]
- Paraskevopoulou, M.D.; Hatzigeorgiou, A.G. Analyzing MiRNA–LncRNA Interactions. Cell. Quiesc. 2016, 1402, 271–286. [Google Scholar]
- Choudhry, H.; Harris, A.L. Advances in Hypoxia-Inducible Factor Biology. Cell Metab. 2018, 27, 281–298. [Google Scholar] [CrossRef]
- Azam, S.; Hou, S.; Zhu, B.; Wang, W.; Hao, T.; Bu, X.; Khan, M.; Lei, H. Nuclear retention element recruits U1 snRNP components to restrain spliced lncRNAs in the nucleus. RNA Biol. 2019, 16, 1001–1009. [Google Scholar] [CrossRef]
- He, C.; Yang, W.; Yang, J.; Ding, J.; Li, S.; Wu, H.; Zhou, F.; Jiang, Y.; Teng, L.; Yang, J. Long Noncoding RNA MEG3 Negatively Regulates Proliferation and Angiogenesis in Vascular Endothelial Cells. DNA Cell Biol. 2017, 36, 475–481. [Google Scholar] [CrossRef]
- Clemson, C.M.; Hutchinson, J.N.; Sara, S.A.; Ensminger, A.; Fox, A.; Chess, A.; Lawrence, J.B. An Architectural Role for a Nuclear Noncoding RNA: NEAT1 RNA Is Essential for the Structure of Paraspeckles. Mol. Cell 2009, 33, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Leisegang, M.S.; Bibli, S.-I.; Günther, S.; Pflüger-Müller, B.; A Oo, J.; Höper, C.; Seredinski, S.; Yekelchyk, M.; Schmitz-Rixen, T.; Schürmann, C.; et al. Pleiotropic effects of laminar flow and statins depend on the Krüppel-like factor-induced lncRNA MANTIS. Eur. Hear. J. 2019, 40, 2523–2533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanicek, L.; Lozano-Vidal, N.; Bink, D.I.; Hooglugt, A.; Yao, W.; Wittig, I.; Van Rijssel, J.; Van Buul, J.D.; Van Bergen, A.; Klems, A.; et al. Long non-coding RNA LASSIE regulates shear stress sensing and endothelial barrier function. Commun. Biol. 2020, 3, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.L.; Dunne, K.; Walport, L.J.; Flashman, E.; Kawamura, A. Epigenetic regulation by histone demethylases in hypoxia. Epigenomics 2015, 7, 791–811. [Google Scholar] [CrossRef] [Green Version]
- Strzyz, P. Hypoxia makes its mark on histones. Nat. Rev. Mol. Cell Biol. 2019, 20, 324–325. [Google Scholar] [CrossRef]
- Ou, M.; Li, X.; Zhao, S.; Cui, S.; Tu, J. Long non-coding RNA CDKN2B-AS1 contributes to atherosclerotic plaque formation by forming RNA-DNA triplex in the CDKN2B promoter. EBioMedicine 2020, 55, 102694. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhao, L.; Yu, X.; Zhang, J.; Qian, L.; Jin, J.; Lu, R.; Zhou, Y. Inhibition of EZH2 primes the cardiac gene activation via removal of epigenetic repression during human direct cardiac reprogramming. Stem Cell Res. 2021, 53, 102365. [Google Scholar] [CrossRef]
- Woo, C.J.; Maier, V.K.; Davey, R.; Brennan, J.; Li, G.; Brothers, J.; Schwartz, B.; Gordo, S.; Kasper, A.; Okamoto, T.R.; et al. Gene activation of SMN by selective disruption of lncRNA-mediated recruitment of PRC2 for the treatment of spinal muscular atrophy. Proc. Natl. Acad. Sci. USA 2017, 114, E1509–E1518. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ren, Y.; Wang, Y.; Tan, Y.; Wang, Q.; Cai, J.; Zhou, J.; Yang, C.; Zhao, K.; Yi, K.; et al. A Compound AC1Q3QWB Selectively Disrupts HOTAIR-Mediated Recruitment of PRC2 and Enhances Cancer Therapy of DZNep. Theranostics 2019, 9, 4608–4623. [Google Scholar] [CrossRef]
- Hsu, J.H.-R.; Rasmusson, T.; Robinson, J.; Pachl, F.; Read, J.; Kawatkar, S.; O’Donovan, D.H.; Bagal, S.; Code, E.; Rawlins, P.; et al. EED-Targeted PROTACs Degrade EED, EZH2, and SUZ12 in the PRC2 Complex. Cell Chem. Biol. 2020, 27, 41–46.e17. [Google Scholar] [CrossRef]
- Born, L.J.; Harmon, J.W.; Jay, S.M. Therapeutic potential of extracellular vesicle-associated long noncoding RNA. Bioeng. Transl. Med. 2020, 5, 10172. [Google Scholar] [CrossRef] [PubMed]
- Lucas, T.; Bonauer, A.; Dimmeler, S. RNA Therapeutics in Cardiovascular Disease. Circ. Res. 2018, 123, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Garbo, S.; Tripodi, M.; Battistelli, C. A novel RNA- based approach to counteract EMT. Oncoscience 2021, 8, 53–54. [Google Scholar] [CrossRef] [PubMed]
- Winkle, M.; El-Daly, S.M.; Fabbri, M.; Calin, G.A. Noncoding RNA therapeutics—Challenges and potential solutions. Nat. Rev. Drug Discov. 2021, 20, 629–651. [Google Scholar] [CrossRef] [PubMed]
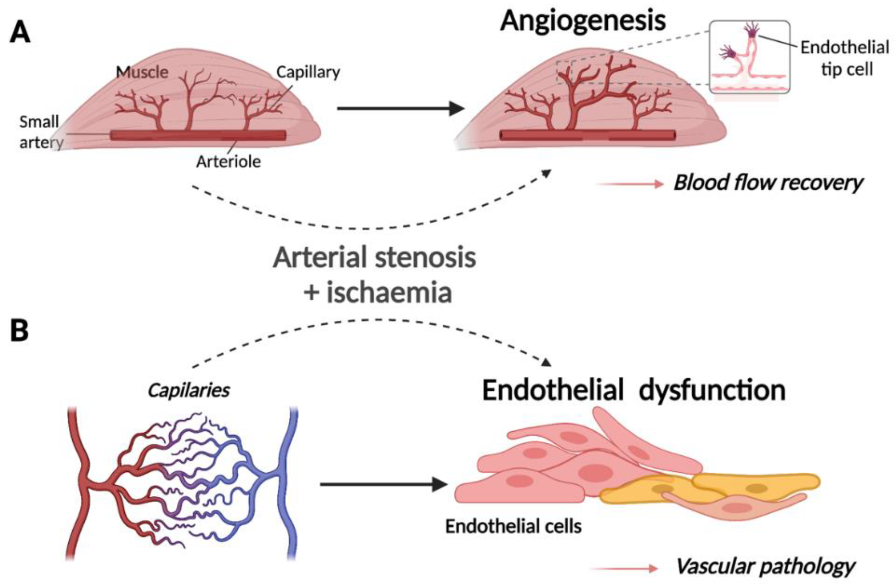
Figure 1.
Schematic representation of (A) the normal physiological response to ischaemia in muscle tissue and (B) the pathogenesis of ischaemic vascular disease in endothelial cells (modified from [7]). Created with BioRender.com. (A) Upon restricted blood supply to the musculature, capillary beds within the ischaemic tissue undergo physiological angiogenesis whereby new vessels branch from the arterioles and infiltrate muscle fibres to increase the delivery of oxygen to metabolically active cells. (B) Upstream arterial stenosis or occlusion due to atherosclerosis may result in ischaemia of capillary ECs, promoting endothelial dysfunction and contributing to pathogenesis of occlusive arterial disease.
Figure 1.
Schematic representation of (A) the normal physiological response to ischaemia in muscle tissue and (B) the pathogenesis of ischaemic vascular disease in endothelial cells (modified from [7]). Created with BioRender.com. (A) Upon restricted blood supply to the musculature, capillary beds within the ischaemic tissue undergo physiological angiogenesis whereby new vessels branch from the arterioles and infiltrate muscle fibres to increase the delivery of oxygen to metabolically active cells. (B) Upstream arterial stenosis or occlusion due to atherosclerosis may result in ischaemia of capillary ECs, promoting endothelial dysfunction and contributing to pathogenesis of occlusive arterial disease.

Figure 2.
Epigenetic mechanisms of gene regulation. Created with BioRender.com. Epigenetic regulators such as (A) DNA methylases, (B) histone-modifying enzymes, and (C) chromatin remodellers modulate gene expression by mediating accessibility to DNA sequences for replicative enzymes or transcription factors. This is achieved through altering the compaction of chromatin by the enzymatic modification of DNA or histones, or by nucleosome repositioning.
Figure 2.
Epigenetic mechanisms of gene regulation. Created with BioRender.com. Epigenetic regulators such as (A) DNA methylases, (B) histone-modifying enzymes, and (C) chromatin remodellers modulate gene expression by mediating accessibility to DNA sequences for replicative enzymes or transcription factors. This is achieved through altering the compaction of chromatin by the enzymatic modification of DNA or histones, or by nucleosome repositioning.

Figure 3.
Schematic representation of the structure of polycomb repressive complex 2 (PRC2) and its interaction with non-coding RNA (ncRNA) via RNA binding sites. Created with BioRender.com. PRC2 has three major proteins including EZH2 (enhancer of zeste homolog 2), EED (embryonic ectoderm development), and SUZ12 (suppressor of zeste 12 homolog). EZH2 and SUZ12 have RNA-binding capacity. Accessory proteins such as JARID2 (jumonji and AT-rich interaction domain containing 2) associate and often co-purify with PRC2.
Figure 3.
Schematic representation of the structure of polycomb repressive complex 2 (PRC2) and its interaction with non-coding RNA (ncRNA) via RNA binding sites. Created with BioRender.com. PRC2 has three major proteins including EZH2 (enhancer of zeste homolog 2), EED (embryonic ectoderm development), and SUZ12 (suppressor of zeste 12 homolog). EZH2 and SUZ12 have RNA-binding capacity. Accessory proteins such as JARID2 (jumonji and AT-rich interaction domain containing 2) associate and often co-purify with PRC2.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Epigenetic mechanisms of nuclear vascular lncRNAs and approaches used to explore their function.
Table 1.
Epigenetic mechanisms of nuclear vascular lncRNAs and approaches used to explore their function.
| lncRNA | Function | Epigenetic Mechanism | Cell Type | Ref |
|---|---|---|---|---|
| Nucleus | ||||
| Chromatin isolation by RNA purification (ChIRP) | ||||
| MEG3 | Modulates activity of TGFb pathway genes by RNA–DNA triplex formation | Recruits PRC2 complex to repress TGFb by H3K27me3 methylation | BT-549 | [26] |
| Chromatin immunoprecipitation (ChIP) | ||||
| MALAT1 | Mediates hyperglycaemia-induced glomerular EC injury and diabetic nephropathy | Recruits G9a methyltransferase to increase H3K9me1 levels on the klotho promoter to repress transcription | HRGECs | [27] |
| ChIP following lncRNA knockdown | ||||
| NEAT1 | Represses VSMC contractile genes to promote a proliferative phenotype and vascular repair in response to injury | Sequesters WDR5 modifier to decrease transcriptionally active chromatin marks (H3K4me3, H3K9ac) and increase inactive modifications, condensing chromatin structure and facilitating gene repression | HCASMCs | [22] |
| RNA immunoprecipitation (RIP) | ||||
| LEENE | Promotes eNOS expression and endothelial cell function | Recruits and enhances binding of RNA polymerase II, initiating pro-transcriptional chromatin remodelling at the eNOS promoter | HUVECs | [21] |
| Individual nucleotide resolution UV cross-linking and immunoprecipitation (iCLIP) | ||||
| MALAT1 | Contributes to VSMC dysfunction in thoracic aortic aneurysm (TAA) | Binds BRG1 and histone deacetylase HDAC9 to pathologically target the complex to the nucleus where HDAC9 recruits EZH2 to transcriptionally silence VSMC-specific genes encoding contractile proteins | VSMCs | [28,29] |
| GATA6-AS | Regulates hypoxia-induced endothelial to mesenchymal transition (EndMT), angiogenesis and EC migration | Binds and negatively regulates H3K4me3 demethylase activity of LOXL2 chromatin modifier to increase activity of target genes | HUVECs | [30] |
| ANRIL | Contributes to development of atherosclerotic plaque through inhibiting macrophage reverse cholesterol transport (mRCT) | Recruits EZH2 and acts as a scaffold by forming RNA–DNA triplexes with the CDKN2B promoter to enrich H3K27me3 and induce repressive chromatin remodelling under regulation by CTCF protein to silence CDKN2B transcription | THP-1 macrophage-derived foam cells | [17,31] |
BT-549—Human breast epithelial carcinoma cell line, HRGECs—Human renal glomerular endothelial cells, HCASMCs—Human coronary artery smooth muscle cells, HUVECs—Human umbilical vein endothelial cells, VSMCs—Vascular smooth muscle cells, BRG1—Brahma-related gene-1.
Table 2.
Total number of Gene Expression Omnibus (GEO) series reporting the use of named techniques in vascular field between 2018–present.
Table 2.
Total number of Gene Expression Omnibus (GEO) series reporting the use of named techniques in vascular field between 2018–present.
| Search Terms | Reported | |
|---|---|---|
| Technique Used | Cell Type | Number of GEO Series |
| RIP | Vascular cells | 1 (GSE142386) |
| Endothelial Cells | 2 | |
| ATAC-seq | Endothelial Cells | 13 |
| HAEC | 4 | |
| CLASH | Endothelial Cells | 1 (GSE101978) |
| iCLIP | Endothelial Cells | 2 |
| HUVEC | 1 (GSE99686) | |
| ChIP-seq | HUVEC | 23 |
| HAEC | 2 | |
| VSMC | 2 | |
| HPAEC | 3 | |
| RNA-seq | Endothelial Cells | 99 |
| HUVEC | 44 | |
| VSMC | 3 |
RIP—RNA immunoprecipitation, ATAC-seq—Assay for transposase-accessible chromatin using sequencing, CLASH—Cross-linking ligation and sequencing of hybrids, iCLIP—Individual-nucleotide-resolution UV cross-linking and immunoprecipitation, ChIP-seq—Chromatin immunoprecipitation with sequencing, RNA-seq—RNA sequencing, HAEC—Human aortic endothelial cells, HPAEC—Primary human pulmonary artery endothelial cells.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Garratt, H.; Ashburn, R.; Sopić, M.; Nogara, A.; Caporali, A.; Mitić, T. Long Non-Coding RNA Regulation of Epigenetics in Vascular Cells. Non-Coding RNA 2021, 7, 62. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7040062
AMA Style
Garratt H, Ashburn R, Sopić M, Nogara A, Caporali A, Mitić T. Long Non-Coding RNA Regulation of Epigenetics in Vascular Cells. Non-Coding RNA. 2021; 7(4):62. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7040062
Chicago/Turabian StyleGarratt, Hester, Robert Ashburn, Miron Sopić, Antonella Nogara, Andrea Caporali, and Tijana Mitić. 2021. "Long Non-Coding RNA Regulation of Epigenetics in Vascular Cells" Non-Coding RNA 7, no. 4: 62. https://0-doi-org.brum.beds.ac.uk/10.3390/ncrna7040062
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.