The Importance of Yeasts on Fermentation Quality and Human Health-Promoting Compounds †


CQ-VR, Chemistry Research Centre, University of Trás-os-Montes and Alto Douro (UTAD), School of Life Sciences and Environment, Dep. of Biology and Environment, Enology building, 5000-801 Vila Real, Portugal
†
I dedicate this small review article to my daughter, Ana Margarida, a 1st-year student of Pharmaceutical Sciences. I would like to encourage her to study these subjects in the search for more natural health-promoting compounds, of natural origin, in detrimental of “laboratory made” ones.
Fermentation 2019, 5(2), 46; https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation5020046
Submission received: 19 April 2019
/
Revised: 27 May 2019
/
Accepted: 29 May 2019
/
Published: 31 May 2019
(This article belongs to the Special Issue Modern Technologies and Their Influence in Fermentation Quality)
Abstract
:Non-Saccharomyces are important during wine fermentation once they influence wine composition. In the early stages of wine fermentation, and together with indigenous or commercial strains of Saccharomyces cerevisiae, non-Saccharomyces are able to transform grape-must sugars into ethanol, CO2, and other important secondary metabolites. A better understanding of yeast biochemistry will allow the selection of yeast strains that have defined specific influences on fermentation efficiency, wine quality, and the production of human health-promoting compounds. Yeast metabolism produces compounds derived from tryptophan, melatonin, and serotonin, which are found in fermented beverages, such as wine and beer. Melatonin is a neurohormone secreted from the pineal gland and has a wide-ranging regulatory and neuroprotective role, while serotonin, as well as being a precursor of melatonin synthesis, is also a neurotransmitter. This review summarizes the importance of some conventional and nonconventional yeast strains’ alcoholic fermentations, especially in the production of metabolites that promote human health and thus, attract consumers attention towards fermented beverages. A brief reference is also made on fermented beverages containing probiotics, namely kombucha, also known as kombucha tea, and its interesting microorganism’s symbiotic relationships named SCOBY.
Keywords:
Yeasts; alcoholic beverages; resveratrol; glutathione; trehalose; tryptophan; melatonin; serotonin; tyrosol; tryptophol; hydroxytyrosol; IAA; probiotics
1. Introduction
The term “fermentation” comes from the Latin word “fermentum” (meaning, to ferment). The science of fermentation is called “zymology” and the first zymologist was Louis Pasteur who was able to identify and apply yeast in fermentation [1]. Food fermentations date back at least 6000 years. In the 16th century, the beginning of industrialization initiated technological interventions in food and beverage production [2]. However, it was in the last two centuries that significant changes in the world’s food system have occurred. In olden days, fermentation of food was meant for food preservation and flavor improvement [3], nowadays, in food and beverages fermentation, various technologies and operations are used with the aim of converting fairly perishable and indigestible raw materials into pleasant foods and drinkable beverages with added value and high stability [4]. The assurance of the quality and safety of the final product is the main goal of the technologies applied [5].
Biotechnology plays a radical role in the production, conservation, nutritional enrichment, and value addition of foods. To be able to understand the science of microbiology in food and beverage applications with identification of new-fermenting species is an advantage to enhance the quality of our food products.
Food and beverage processing using microorganisms is the most suitable technology for the development of innovative fermented food products. Solid state fermentation is used for processing of vinegar, soy sauce, tea, and cheese [6]. Wine, beer, distilled beverages, and yogurt are developed by submerged fermentation. Both methods of fermentation are influenced by numerous factors, including temperature, pH, nature, and composition of the medium, dissolved O2, dissolved CO2, operational system, and feeding with precursors, among others. Variation in these factors may affect the rate of fermentation, the product spectrum and yield, the sensory properties of the product (appearances, taste, smell, texture), physic-chemical properties, nutritional quality, and the production of metabolites that promote human health attracting consumers attention towards fermented products, namely beverages.
2. Alcoholic Beverages Consumption and Health-Promoting Compounds
The prevention of diseases by altering lifestyle and dietary conducts may present more benefits than medical care. Up till now, adjusting individual dietary habits is a challenge. Most often, consumers must choose between nutrition, taste, price, convenience, and cost [7]. Nowadays, the nutritional value appears to be the health benefit that has the most impact on a consumer’s purchase [8].
In the last 30 years, the effects of wine on human health have been studied by many scientists. In 1992, Renaud and de Lorgeril [9] published a study about the higher wine consumption in the French population, in comparison with other industrialized countries. According to these authors, wine caused a lower incidence of coronary heart disease, notwithstanding the intake of the traditional French diet, rich in saturated fatty acids. This finding constituted the so-called “French paradox”. Since then, many studies have been carried out on wine indicating that persons consuming daily moderate amounts of wine display a reduction of cardiovascular mortality and an improvement of antioxidant parameters, when compared with individuals who abstain or who drink alcohol to excess [10,11]. Moreover, Poli et al. [11] also mentioned that in healthy adults, spontaneous consumption of alcoholic beverages (30 g ethanol/day for men and 15 g/day for women) is to be considered acceptable. However, the possible interactions between alcohol and acute or chronic drug use must be discussed with the primary care physician.
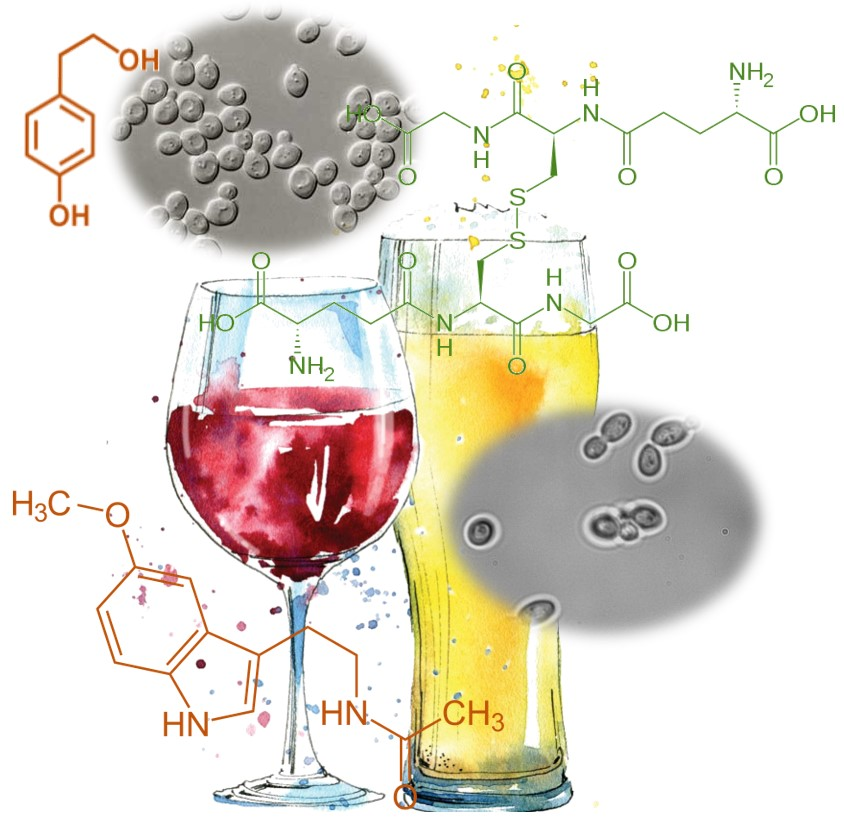
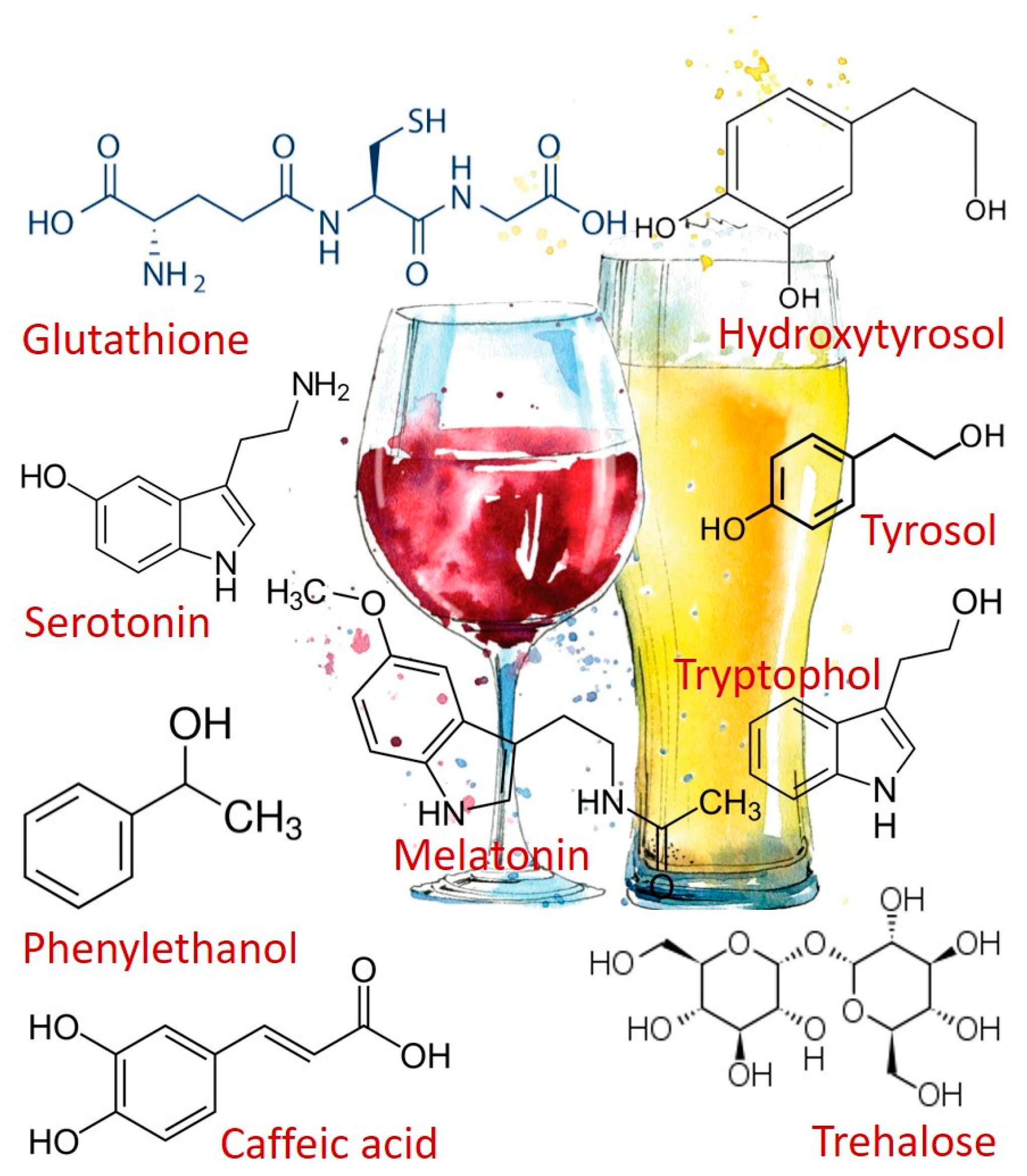
Oxidative stress and antioxidant deficiency have been implicated in the pathogenesis of many diseases and conditions, including atherosclerosis, cancer, aging, and respiratory disease. Glutathione (L-g-glutamyl-L-cysteinyl-glycine, GSH) (Figure 1) is a major antioxidant acting as a free radical scavenger that protects the cell from ROS (reactive oxygen species). In addition, GSH is involved in nutrient metabolism and regulation of cellular metabolic functions ranging from DNA and protein synthesis to signal transduction, cell proliferation, and apoptosis [12,13,14].
Another important molecule is trehalose (Figure 1). This sugar possesses inflammatory properties [15] presenting, also, the ability to protect cellular membranes and labile proteins against denaturation as a result of desiccation and oxidative stress [16].
Yeast metabolism produces compounds derived from tryptophan, which are found in fermented beverages, such as wine and beer. In particular, melatonin and serotonin (Figure 1). Serotonin is a neurohormone that regulates circadian rhythms, and also has an alleged protective effect against neurodegenerative and degenerative diseases (Alzheimer’s, Parkinson’s and Angiogenesis) [17]. Moreover, serotonin is a neurotransmitter itself, and a precursor of melatonin synthesis.
In humans, melatonin (N-acetyl-5-methoxytryptamine) is a hormone that modulates several physiological processes. This molecule is an indole-amine found in many living organisms like plants, microorganisms, and humans. Melatonin modulates many human physiological processes including the sleep/wake cycle and the reproductive physiology via a receptor-mediated mechanism [18,19] acting, also, as an antioxidant via nonreceptor processes [20]. It is well known that the intake of foods containing melatonin increases its level in plasma and the number of melatonin-derived metabolites [21]. Studies have been carried out to identify melatonin in grapes [22] and beverages such as beer and wine [23,24]. Interesting is the reported concentrations of melatonin in grapes (Vitis vinifera L.) and wines: 150 µg/g in Merlot grapes [25]; 130 ng/mL in Tempranillo wine [26].
Tyrosol and tryptophol (Figure 1) are produced by yeasts during alcoholic fermentation from the catabolism of amino acids tyrosine and tryptophan, respectively, whereas hydroxytyrosol is produced by hydroxylation of its precursor tyrosol. Tyrosol, hydroxytyrosol, tryptophol are reported to possess several health-enhancing activities, deriving from their free radical scavenging, anticarcinogenic, cardioprotective (induces myocardial protection against ischemia-related stress [27]) and antimicrobial properties [28].
It´s due to the presence of tyrosol and caffeic acids (Figure 1), that white wine has been reported as having cardioprotective benefits. Tyrosol and caffeic acids are able to activate the cell survival signaling pathway and the FOXO3a longevity-associated gene [29,30]. Moreover, tyrosol has been shown to have an important role in the taste of some alcoholic beverages, such as sake [31] and wine [32] by exhibiting a bitter taste above the sensory threshold, but below the recognition threshold.
Tryptophol is also used as a precursor in the synthesis of the drug Indoramin, an α-adrenoreceptor blocking drug used to treat hypertension [33], and in the treatment of benign prostatic hyperplasia [34].
Phenylethanol (Figure 1), also produced by Candida albicans as an auto-antibiotic [35] is an aromatic compound that is commonly found in plants, such as roses, possessing pleasant floral rose-like odor. Due to its preservative properties, phenylethanol is often used in soap-based detergents because of its stability in basic conditions. Phenylethanol can also act as a natural preservative in wine and beer to prevent spoilage [35].
3. Mechanisms of Microbial Resistance to Environment Changes that Produce Health-Promoting Compounds
Conservation and commercialization of yeast cultures in fresh liquid or pressed forms are not economically advantageous. Thus, dehydrated yeasts present numerous advantages, such as lower cost, convenient for transport and storage, and ease of handling [36]. However, the drying of the yeasts signifies highly sensitive transformation processes for microorganism’s which can lead to cell death or a significant decrease in cell activity potential [37]. The final water volume of the cells, induced by dehydration-rehydration cycles, influence the cells survival [38], and the modification of plasma membrane fluidity during the dehydration-rehydration cycles affects the plasma membrane structure and may induce cell mortality [39].
Increase of contact surface of the cells with air during dehydration also induces accumulation of ROS (reactive oxygen species)—[O2•− (superoxide anion), •OH (hydroxyl radical), H2O2 (hydrogen peroxide) and ReOOH (hydroperoxides)]—and may contribute to inactivation of several enzymes, leading, also, to cell death [40]. In the presence of these stress conditions yeasts are able to synthesize compounds such as glutathione and trehalose [41].
Glutathione (GSH) is a ubiquitous low molecular weight thiol tripeptide containing glutamate, cysteine, and glycine (Glu-Cys-Gly), it is present in large amounts in yeasts and it can be found in the reduced or oxidized forms (GSH and GSSG, respectively). Glutathione plays a crucial role in redox equilibrium reactions, protecting the cell from oxidative stress, by allowing the formation of native disulfide bonds and by scavenging free radicals present in the cytosol; reactions mediated via glutathione reductase and glutathione peroxidase [12,42].
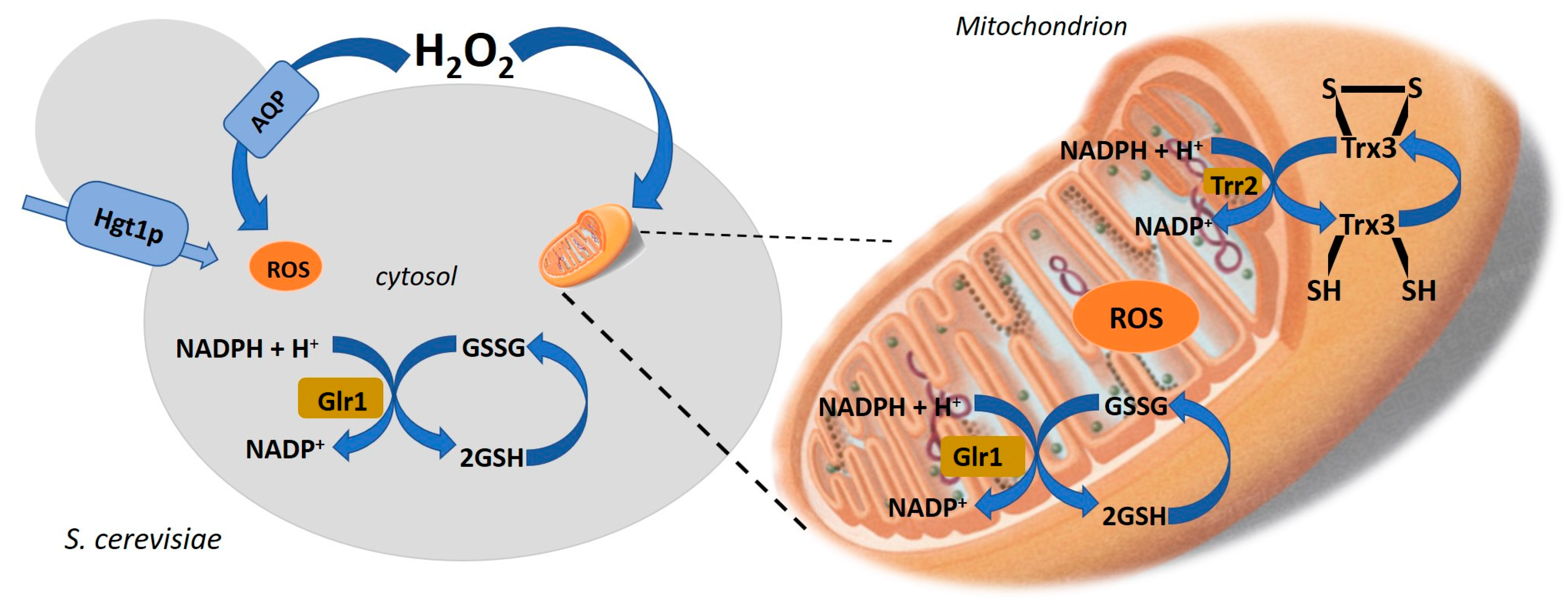
Hgt1p in yeast S. cerevisiae was the first identified high-specificity and high-affinity glutathione transporter (Km 54 mM) [43]. Hgt1p belongs to oligopeptide transporter family which was also found in fungi, plants, and prokaryotes. Genetic and physiological investigations revealed that gene HGT1 (open reading frame YJL212c) as encoding a high-affinity glutathione transporter. Yeast strains deleted in HGT1 gene did not show any detectable plasma membrane glutathione transport. This transporter is required for the uptake of glutathione from the extracellular medium (Figure 2) [43]. Moreover, mitochondria are a primary source of ROS in cells and mitochondrial thiols are therefore major ROS targets. This phenomenon is exacerbated by the relatively alkaline pH of mitochondria. Therefore, redox regulation is critical for numerous mitochondrial functions, and yeast strains lacking GSH are unable to grow by respiration due to an accumulation of oxidative damage to mitochondrial DNA. Transport of H2O2 across yeast cell membranes can be facilitated by transporters such as aquaporins. Hydrogen peroxide causes oxidative stress but also plays important roles as a signaling molecule in the regulation of many biological processes (Figure 2) [44].
Thiol redox regulation plays a role in the response of cells to oxidative stress conditions. Gostimskaya and Grant [45] emphasize the importance of compartmentalized redox regulation when cells are subjected to oxidative stress conditions (Figure 2). At the same time as cytosolic glutathione represents the first major pool of thiols, which would be a target of oxidation in response to exposure to an exogenous oxidant, it is the mitochondrial glutathione pool which is crucial for oxidant tolerance.
Has mentioned before, one interesting disaccharide with potential for medical application, namely in ophthalmology, is trehalose [15]. This sugar, constituted by two glucose residues linked by an α-1-1 glycosidic bond, is widespread in many species of plants, animals and microorganisms, including wine yeasts, where its function is to protect cells against desiccation. However, this sugar is no naturally found in mammals. Nowadays trehalose is used in the biopharmaceutical preservation of labile protein drugs and in the cryopreservation of human cells and is under investigation for the treatment of Huntington’s chorea and Alzheimer’s disease. It can also be used as a preventive drug to treat dryness in mammalian eyes, a common tear and ocular surface multifactorial disease that can lead to inflammatory reaction [15]. Trehalose also acts as a storage carbohydrate for the cell and it plays a very important role as a protectant against osmotic stress and ethanol stress, in yeast cells [47]. Trehalose acts as a stabilizing effect on the plasma membrane, providing it with increased tolerance to desiccation, dehydration, temperature changes, and high temperature [16]. It can also act as an antioxidant component by reducing oxidation reactions rates while enhancing the viscosity of cell cytoplasm [48].
Câmara et al. [49] studied the effects of glutathione and trehalose biosynthesis in the dehydration stress responses of three non-Saccharomyces yeasts strains (Torulaspora delbrueckii CBS4865, Metschnikowia pulcherrima CBS5833 and Lachancea thermotolerans CBS6340). The results obtained will help to better understand certain physiological responses of non-Saccharomyces yeasts to dehydration, leading to the promotion and production of new high performance dehydrated non-Saccharomyces yeasts strains to be used in food and beverages elaboration. According to the mentioned work [49], yeasts grown in nutrient-rich medium accumulated glutathione leading to a higher resistance to dehydration, whilst the nutrient-poor medium induced the cells to accumulate large amounts of trehalose, which partially protected them from GSSG accumulation.
4. Melatonin and Other Tryptophan Metabolites
In the scientific world, the theme of “wine and health” topics have been focused mainly on polyphenols, once these bioactive compounds are present in plants and are released into fermented products. However, yeast also transforms other molecules into biologically active compounds [19]. Since the pioneering work of Sprenger and co-workers [50] that melatonin molecule, has been reported as being present in wine, and its presence has been related to the activity of the yeast involved in the fermentation process. Originally, seen as a unique product of the pineal gland of vertebrates, called a neurohormone, at the present, it is considered a ubiquitous molecule present in most living organisms [51].
Rodriguez-Naranjo and co-workers [26] studied the capacity of different yeasts to produce melatonin during alcoholic fermentation. Different Saccharomyces yeast strains, used for industrial fermentation of beer or as nutritional complements, and non-Saccharomyces yeast strains (Metschnikowia pulcherrima and Starmerella bacillaris) were tested by the referred authors to analyze intracellular and extracellular melatonin production in synthetic grape must. Interestingly, at the beginning of fermentation melatonin was detected, in the intracellular compartment, either in Saccharomyces or in non-Saccharomyces strains. Production levels differed among strains, being Starmerella bacillaris the non-Saccharomyces yeast that presented the highest concentration. Nevertheless, extracellular melatonin was detected at different time-points over the fermentation process, depending on the yeast strain. However, the same authors [26] also reported that the presence of tryptophan is essential for melatonin production since it is its principal precursor, it increases final melatonin content and it accelerates its formation. Moreover, the synthesis of melatonin largely depends on the growth phase of the yeast and the concentration of the reducing sugars.
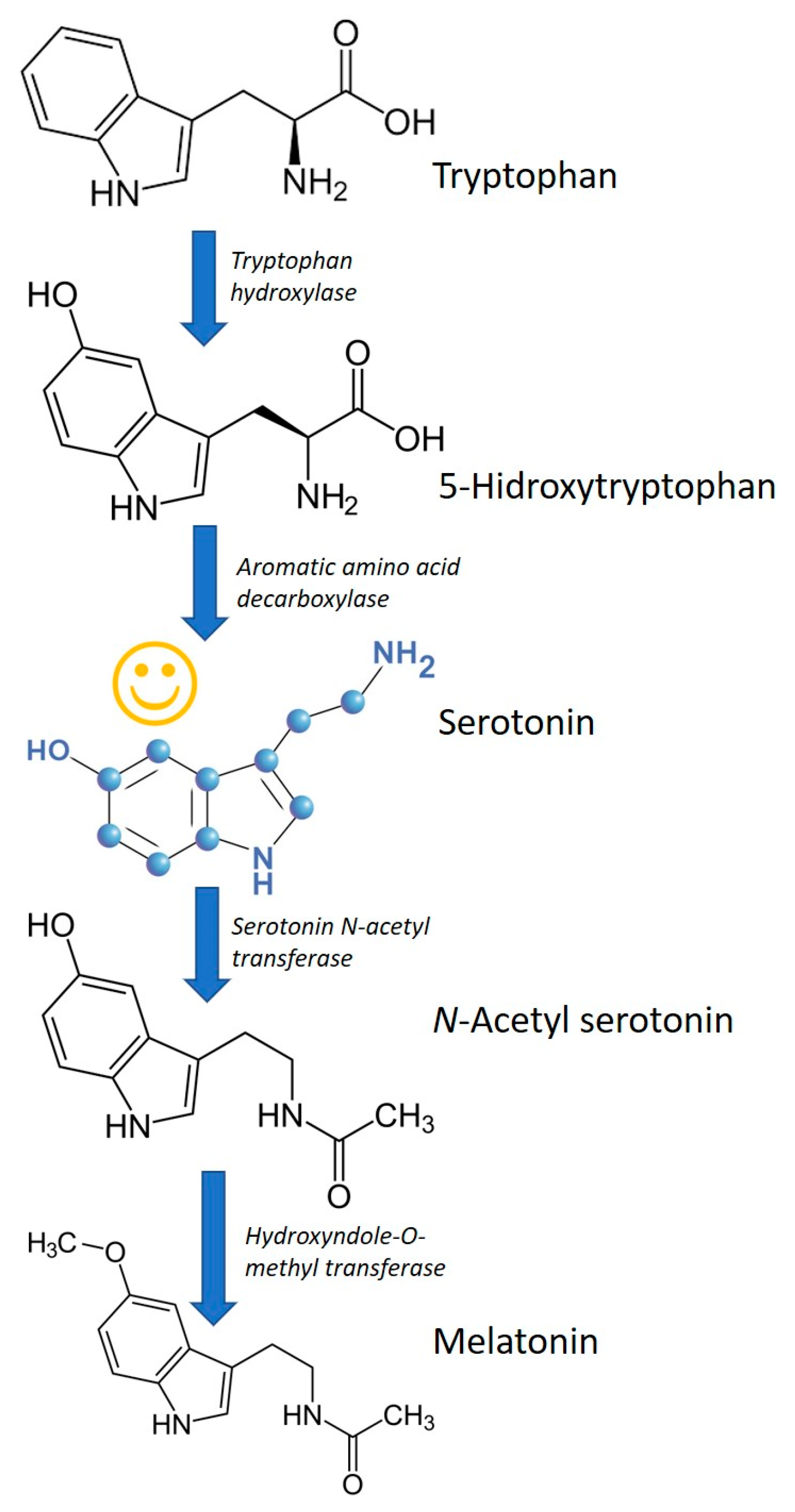
The metabolic pathway for melatonin production in yeast is not completely clarified, nevertheless, the formation of serotonin might be an intermediate metabolite in the pathway [19], Figure 3.
Germann et al. [52], studied de novo melatonin biosynthesis from glucose, by genetically modifying Saccharomyces cerevisiae strains that comprise heterologous genes encoding one or more variants of L-tryptophan hydroxylase, 5-hydroxy-L-tryptophan decarboxylase, serotonin acetyltransferase, acetylserotonin O-methyltransferase, and means for providing the cofactor tetrahydrobiopterin via heterologous biosynthesis and recycling pathways. At the end of the process, yeast strain produced melatonin in concentrations of 14.50 ± 0.57 mg L−1 in a 76 hours fermentation, using glucose as sole carbon source.
Although is thought to be an intermediate for melatonin production, there is no evidence for the production of serotonin by S. cerevisiae. Serotonin has been found in wines at levels ranging from 2 to 23 mg L−1, mainly as a result of the malolactic fermentation performed by Lactobacillus plantarum strains [53,54].
In 2016, Tan and co-workers [55], proposed an alternative pathway for synthesizing melatonin from L-tryptophan via 5-methoxytryptamine with the formation of the intermediate compound N-acetyl-5-hydroxytryptamine [56]. Different Saccharomyces strains (Lalvin QA23, Enartis ES488, Lallemand ICV-GRE, and Uvaferm) and non-Saccharomyces yeast (Torulaspora delbrueckii and Metschnikowia pulcherrima) have been reported to produce melatonin during alcoholic fermentation in synthetic must [57,58]. Moreover, T. delbrueckii is considered an innovative biotechnological tool, of great commercial interest, to be used in pure culture or in sequential inoculation with S. cerevisiae, for bio-modulating wines acidity [59] among other interesting enological features [60].
Tryptophan metabolism includes, in addition to 5-hydroxytryptamine and melatonin, other important metabolites such as indolic compounds like 3-indoleacetic acid (3-IAA) which is the most common plant hormone, of the auxin class, and is known to have many effects including cell proliferation enhancement and antioxidant properties [61]. Kim and co-workers [61] investigated the effects of IAA on H2O2 induced oxidative toxicity in human dental pulp stem cells and verified that H2O2-induced cytotoxicity was attenuated after IAA treatment. Moreover, according to Fernández-Cruz and co-workers [56], besides IAA, tryptophan metabolites (tryptophol, tryptamine, and l-tryptophan ethyl ester) also present potential as antioxidants and neuroprotective agents. The mentioned authors [56] examined the occurrence of these compounds during the alcoholic fermentation of musts from seven grape cultivars. Fermentations were performed with three S. cerevisiae strains and, in two cases, a sequential inoculation with Torulaspora delbrueckii. Interestingly, they found that the profile of indolic compounds during alcoholic fermentation depended on the cultivar and not on the yeast strain used. Nonetheless, fermentation time was found to be a more influential factor [56].
5. Fusel Alcohols Formed Via the Ehrlich Pathway
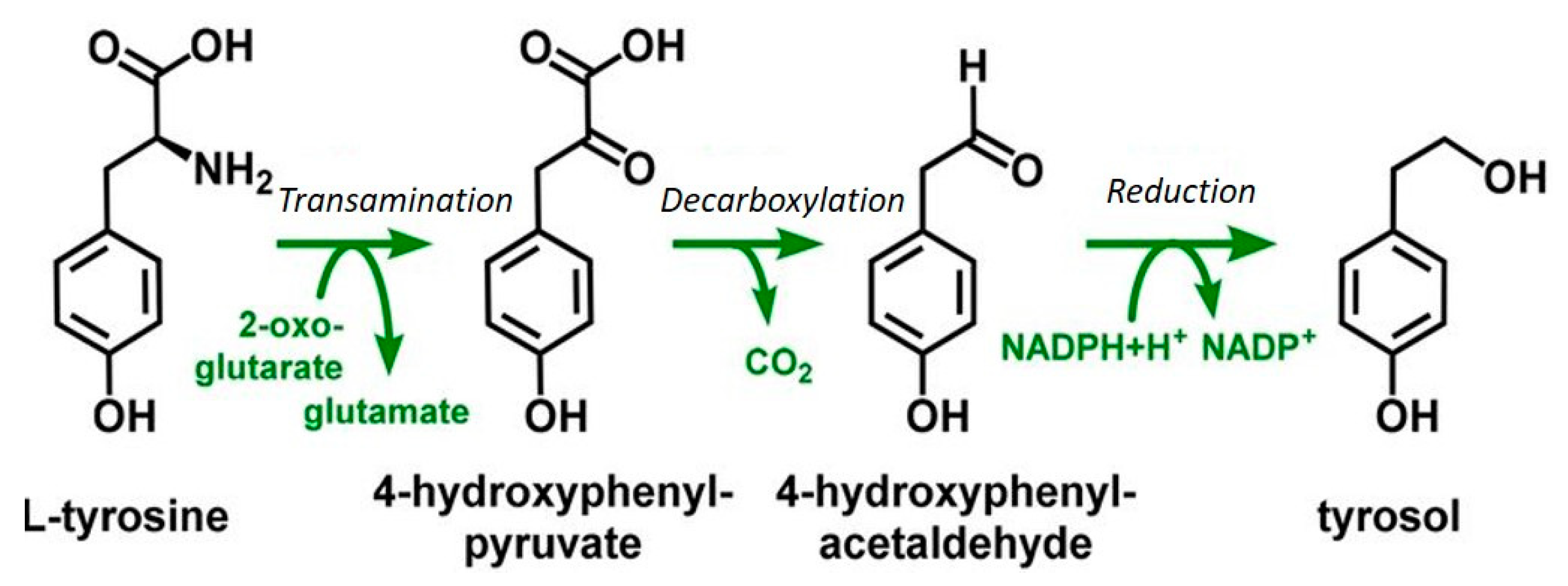
The synthesis of tryptophol by yeast was first described by Felix Ehrlich in 1912 [62,63] as the metabolic conversion of amino acids via the successive steps of transamination, decarboxylation, and reduction [64], Figure 4.
Similarly, to tryptophol, phenylethanol, and tyrosol, are phenolic compounds or fusel alcohols formed via the Ehrlich pathway by yeast metabolism. These compounds can yield health benefits as well as contribute to the flavors and aromas of fermented food and beverages [63,65].
Banach and Ooi [65] investigated the possibility of increasing the yield of tyrosol, tryptophol, and phenylethanol in wine (Alexander’s Pinot Chardonnay grape juice) and beer [modification of the English Ale recipe composed of chocolate malt barley grain, dried malt extract (DME) and liquid malt extract (LME), supplemented with either the equivalent volume of malt-kiwi purée or with amino acids] using two different yeast strains, and supplementing the substrate with the relevant amino acid precursors or fruits high in these amino acids. At the end of the work, they found that flavor enhancement and enrichment of antioxidants, in wine and beer, could be achieved through supplementing the fermentation (in the case of beer-fruit-supplemented beer) media with precursor amino acids as well as careful choices of the appropriate yeast strain.
6. Fermented Beverages Containing Probiotics
It is common knowledge that most of the fermented milk contains probiotic microorganisms (live microorganisms, which when administered in adequate amounts, confer a health benefit on the host). Yogurt, the most common product of milk lactose fermentation, has on its constitution several lactic acid bacteria. So, the domination of milk-based beverages fermented by LAB, mainly Leuconostoc, lactobacilli, and lactococci, is clear. Milk fermentation in colder climates promotes the growth of mesophilic bacteria such as Lactococcus and Leuconostoc, whereas beverages produced at higher temperatures usually have greater counts of thermophilic bacteria such as Lactobacillus and Streptococcus [66,67]. Most often the probiotic bacteria come from Lactobacillus or Bifidobacterium or a cocktail of both [68].
Another class of fermented beverages is those made from cereals (maize, millet, barley, oats, rye, wheat, rice and sorghum), were the natural microbial component is used to ferment grains. The microbial populations responsible for the fermentation of these beverages are not, yet, well characterized. Of several blends, it has been suggested that fermentation by S. cerevisiae, Leuconostoc mesenteroides, and Lactobacillus confusus produce the most palatable beverages [66].
One example of a known fermented beverage, with probiotic characteristics, is kombucha. It is a fermented sweetened tea, originally from China, but, enjoyed worldwide. Medusomyces gisevii Lindau represents a symbiotic microbial community known as “tea fungus” or kombucha tea. During tea fermentation, the added sugar is converted into organic acids and ethanol by yeast and bacteria. The microorganisms co-exist in interdependent symbiotic relationships named SCOBY (symbiotic culture of bacteria and yeast). Organic acids and ethanol formed during tea fermentation protect SCOBY from the colonization of other microorganisms. The yeast present in SCOBY can vary and may include Brettanomyces/Dekkera, Schizosaccharomyces, Torulaspora, Zygosaccharomyces, and Pichia [69]. The bacteria that we can find in kombucha are species of Gluconacetobacter xylinus, G. kombuchae sp. nov., Acetobacter nitrogenifigens sp. nov, Acetobacter intermedius, sp. nov. [70,71].
Nondairy fermented beverage from a blend of cassava and rice based on Brazilian indigenous beverage “cauim” using probiotic lactic acid bacteria and yeast was studied by Freire and co-workers in 2017 [72]. A triple group of microorganisms was used to ferment the blend: Lactobacillus plantarum CCMA 0743 (from cauim), Torulaspora delbrueckii CCMA 0235 (from tarubá), and the commercial probiotic L. acidophilus LAC-04. According to the authors [72], the bacteria populations were around 8.0 log (CFU mL−1) at the end of all fermentations as recommended for probiotic products. The final beverage obtained was considered a non-alcoholic drink since the ethanol degree was lower than 0.5% (v/v).
7. Final Remarks
The choice to consume alcohol should be based on individual considerations, taking into account the influence on health and diet, the risk of alcoholism and abuse, the effect on behavior, and other factors that may vary with age and lifestyle.
Fermentation remains the oldest but most prevalent means of food and drinks processing and preservation. Important studies have been carried out aiming to understand certain physiological responses of Saccharomyces and non-Saccharomyces yeasts during the fermentation process, in order to promote the production and the marketing of new high-performance dehydrated yeasts strains for food and beverages elaboration, that will also be able to produce health-promoting compounds.
Fermented drinks, namely wine and beer, can definitely serve as vehicles for beneficial compounds that play an important role in human health, namely in the prevention of some 21st-century diseases. However, further studies are required to ascertain the combination of grape cultivar and inoculation strain or inoculation strategy that could optimize the concentration of health-promoting compounds so as to realize their potential bioactivity in wine.
In what concerns probiotics, the credibility of specific health claims, and their safety must be established through science-based clinical studies. Maybe, in the future, probiotics will be used as approved drugs that will be prescribed together with, or instead of, antibiotics.
The role of traditional beverages, in the future of the fermented beverage industry, maybe to inspire the expansion of new products and, of course, assess a country’s disposition to accept them. Undeniably, with the availability and improvements in technology, and consumers’ increasing interest in functional foods, the positioning in the market for fermented beverages is more promising than ever.
Funding
We appreciate the financial support provided to the Research Unit in Vila Real (PEst-OE/QUI/UI0616/2014) by FCT—Portugal and COMPETE.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Manchester, K.L. Louis Pasteur (1822–1895)—chance and the prepared mind. Trends Biotechnol. 1995, 13, 511–515. [Google Scholar] [CrossRef]
- Truninger, M. The historical development of industrial and domestic food technologies. In The Handbook of Food Research; Murcott, A., Belasco, W., Jackson, P., Eds.; Bloomsbury: London, UK, 2013; pp. 82–102. [Google Scholar]
- Mishra, S.S.; Ray, R.C.; Panda, S.K.; Montet, D. Technological Innovations in Processing of Fermented Foods. An Overview. In Fermented Foods, Part II: Technological Intervention, 1st ed.; Ray, R.C., Montet, D., Eds.; Taylors and Francis, CRC Press: London, UK, 2017; p. 525. [Google Scholar]
- Ray, R.C.; Joshi, V.K. Fermented Foods: Past, present, and future scenario. In Microorganisms and Fermentation of Traditional Foods; Ray, R.C., Montet, D., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 1–36. [Google Scholar]
- Ray, R.C. Fermented foods in health-related issues. Int. J. Food Ferment. Technol. 2013, 3, 1. [Google Scholar]
- Ghosh, J.S. Solid state fermentation and food processing: a short review. J. Nutr. Food Sci. 2015, 6, 453. [Google Scholar] [CrossRef]
- Blaylock, J.; Smallwood, D.; Kassel, K.; Variyam, J.; Aldrich, L. Economics, food choices, and nutrition. Food Policy 1999, 24, 269–286. [Google Scholar] [CrossRef] [Green Version]
- Lähteenmäki, L. Claiming health in food products. Food Qual. Prefer. 2013, 27, 196–201. [Google Scholar] [CrossRef]
- Renaud, S.D.; de Lorgeril, M. Wine, alcohol, platelets, and the French paradox for coronary heart disease. Lancet 1992, 339, 1523–1526. [Google Scholar] [CrossRef]
- Guilford, J.M.; Pezzuto, J.M. Wine and health: A review. Am. J. Enol. Vitic. 2011, 62, 471–486. [Google Scholar] [CrossRef]
- Poli, A.; Marangoni, F.; Avogaro, A.; Barba, G.; Bellentani, S.; Bucci, M.; Cambieri, R.; Catapano, A.L.; Costanzo, S.; Cricelli, C.; et al. Moderate alcohol use and health: A consensus document. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 487–504. [Google Scholar] [CrossRef]
- Chakravarthi, S.; Jessop, C.E.; Bulleid, N.J. The role of glutathione in disulfide bond formation and endoplasmic-reticulum-generated oxidative stress. EMBO Rep. 2006, 7, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Fang, Y.-Z.; Yang, S.; Lupton, J.R.; Turner, N.D. Glutathione Metabolism and Its Implications for Health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosnan, J.T.; Brosnan, M.E. Glutathione and The Sulfur-Containing Amino Acids: An Overview. In Glutathione and Sulfur Amino Acids in Human Health and Disease; Masella, R., Mazza, G., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2009; ISBN 978-0-470-17085-4. [Google Scholar]
- Luyckx, J.; Baudouin, C. Trehalose: An intriguing disaccharide with potential for medical application in ophthalmology. Clin. Ophthalmol. 2011, 5, 577–581. [Google Scholar] [CrossRef]
- Eleutherio, E.; Panek, A.; De Mesquita, J.F.; Trevisol, E.; Magalhães, R. Revisiting. yeast trehalose metabolism. Curr. Genet. 2015, 61, 263–274. [Google Scholar] [CrossRef]
- Hornedo-Ortega, R.; Cerezo, A.B.; Troncoso, A.M.; Garcia-Parrilla, M.C.; Mas, A. Melatonin and Other Tryptophan Metabolites Produced by Yeasts: Implications in Cardiovascular and Neurodegenerative Diseases. Front. Microbiol. 2016, 6, 1565. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Manchester, L.C.; Pilar-Terron, M.; Flores, L.J.; Koppisepi, S. Medical implications of melatonin: receptor-mediated and receptor-independent actions. Adv. Med. Sci. 2007, 52, 11–28. [Google Scholar]
- Mas, A.; Guillamon, J.M.; Torija, M.J.; Beltran, G.; Cerezo, A.B.; Troncoso, A.M.; Garcia-Parrilla, M.C. Bioactive compounds derived from the yeast metabolism of aromatic amino acids during alcoholic fermentation. BioMed. Res. Int. 2014, 898045. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: a physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Garrido, M.; Paredes, S.D.; Cubero, J.; Lozano, M.; Toribio-Delgado, A.F.; Muñoz, J.L.; Reiter, R.J.; Barriga, C.; Rodríguez, A.B. Jerte valley cherry-enriched diets improve nocturnal rest and increase 6-sulfatoxymelatonin and total antioxidant capacity in the urine of middle-aged and elderly humans. J. Gerontol. A Biol. Sci. Med. Sci. 2010, 65A, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Vitalini, S.; Gardana, C.; Zanzotto, A.; Simonetti, P.; Faoro, F.; Fico, G.; Iriti, M. The presence of melatonin in grapevine (Vitis vinifera L.) berry tissues. J. Pineal Res. 2011, 51, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.D.; Moreno, H.; Calvo, J.R. Melatonin present in beer contributes to increase the levels of melatonin and antioxidant capacity of the human serum. Clin. Nutr. 2009, 28, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Naranjo, M.I.; Torija, M.J.; Mas, A.; Cantos-Villar, E.; Garcia-Parrilla, M.D. Production of melatonin by Saccharomyces strains undergrowth and fermentation conditions. J. Pineal Res. 2012, 53, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Hall, B.A.; Le, C.H.; Saxena, P.K. Changes in the levels of indoleamine phytochemicals during véraison and ripening of wine grapes. J. Pineal Res. 2010, 49, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Naranjo, M.I.; Gil-Izquierdo, A.; Troncoso, A.M.; Cantos, E.C.; Garcia-Parrilla, M.C. Melatonin: A new bioactive compound present in wine. J. Food Compost. Anal. 2011, 24, 603–608. [Google Scholar] [CrossRef]
- Samuel, S.M.; Thirunavukkarasu, M.; Penumathsa, S.V.; Paul, D.; Maulik, N. Akt/FOXO3a/SIRT1-Mediated Cardioprotection by n-Tyrosolagainst Ischemic Stress in Rat in Vivo Model of Myocardial Infarction: Switching Gears toward Survival and Longevity. J. Agric. Food Chem. 2008, 56, 9692–9698. [Google Scholar] [CrossRef]
- Dudley, J.I.; Lekli, I.; Mukherjee, S.; Das, M.; Bertelli, A.A.; Das, D.K. Does white wine qualify for French paradox? Comparison of the cardioprotective effects of red and white wines and their constituents: resveratrol, tyrosol, and hydroxytyrosol. J. Agric. Food Chem. 2008, 56, 9362–9373. [Google Scholar] [CrossRef]
- Willcox, B.J.; Donlon, T.A.; He, Q.; Chen, R.; Grove, J.S.; Yano, K.; Masaki, K.H.; Willcox, D.C.; Rodriguez, B.; Curb, J.D. FOXO3a Genotype Is Strongly Associated with Human Longevity. Proc. Natl. Acad. Sci. USA 2008, 105, 13987–13992. [Google Scholar] [CrossRef]
- Thirunavukkarasu, M.; Penumathsa, S.V.; Samuel, S.M.; Akita, Y.; Zhan, L.; Bertelli, A.A.; Maulik, G.; Maulik, N. White Wine-induced Cardio Protection against Ischemia-Reperfusion Injury is Mediated by Life-Extending Akt/FOXO3a/NFκB Survival Pathway. J. Agric. Food Chem. 2008, 56, 6733–6739. [Google Scholar] [CrossRef]
- Utsunomiya, H. Flavor terminology and reference standards for sensory analysis of sake. J. Brew. Soc. Jpn. 2006, 101, 730–739. [Google Scholar] [CrossRef]
- Luís, R.S.; Paula, B.A.; Patrícia, V.; Rosa, M.S.; Martha, E.T.; Encarna, V. Analysis of non-colored phenolics in red wine: Effect of Dekkera bruxellensis. Yeast 2005, 89, 185–189. [Google Scholar] [CrossRef]
- Gould, B.A.; Mann, S.; Davies, A.B.; Altman, D.G.; Raftery, E.B. α-Adrenoreceptor Blockade with Indoramin in Hypertension. J. Cardiovasc. Pharmacol. 1983, 5, 343–348. [Google Scholar] [CrossRef]
- Kirby, R.S.; Pool, J.L. Alpha adrenoceptor blockade in the treatment of benign prostatic hyperplasia: Past, present, and future. Br. J. Urol. 1997, 80, 521–532. [Google Scholar] [CrossRef]
- Lingappa, B.T.; Prasad, M.; Lingappa, Y.; Hunt, D.F.; Biemann, K. Phenethyl Alcohol, and Tryptophol: Auto-antibiotics Produced by the Fungus Candida albicans. Science 1969, 163, 192–194. [Google Scholar] [CrossRef] [PubMed]
- Luna-Solano, G.; Salgado-Cervantes, M.A.; Ramirez-Lepe, M.; Garcia-Alvarado, M.A.; Rodriguez-Jimenes, G.C. Effect of drying type and drying conditions over the fermentative ability of brewer’s yeast. J. Food Process. Eng. 2003, 26, 135–147. [Google Scholar] [CrossRef]
- Rapoport, A. Anhydrobiosis and Dehydration of Yeasts. In Biotechnology of Yeasts and Filamentous Fungi; Sibirny, A.A., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 87–116. [Google Scholar]
- Gervais, P.; Beney, L. Osmotic mass transfer in the yeast Saccharomyces cerevisiae. Cell. Mol. Biol. 2001, 47, 831–840. [Google Scholar] [PubMed]
- Dupont, S.; Beney, L.; Ritt, J.-F.; Lherminier, J.; Gervais, P. Lateral reorganization of the plasma membrane is involved in the yeast resistance to severe dehydration. Biochim. Biophys. Acta Biomembr. 2010, 1798, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Garre, E.; Raginel, F.; Palacios, A.; Julien, A.; Matallana, E. Oxidative stress responses, and lipid peroxidation damage are induced during dehydration in the production of dry active wine yeasts. Int. J. Food Microbiol. 2010, 136, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Câmara, A.A., Jr.; Nguyen, T.D.; Jossier, A.; Endrizzi, A.; Saurel, R.; Simonin, H.; Husson, F. Improving total glutathione and trehalose contents in Saccharomyces cerevisiae cells to enhance their resistance to fluidized bed drying. Process. Biochem. 2018, 69, 45–51. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for the quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat. Protoc. 2006, 1, 3159–3165. [Google Scholar] [CrossRef]
- Bourboulou, A.; Shahi, P.; Chakladar, A.; Delrot, S.; Bachhawat, A.K. Hgt1p, a High-Affinity Glutathione Transporter from the Yeast Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 13259–13265. [Google Scholar] [CrossRef]
- Veal, E.A.; Day, A.M.; Morgan, B.A. Hydrogen peroxide sensing and signaling. Mol. Cell 2007, 26, 1–14. [Google Scholar] [CrossRef]
- Gostimskaya, I.; Grant, C.M. Yeast mitochondrial glutathione is an essential antioxidant with mitochondrial thioredoxin providing a back-up system. Free Radic. Biol. Med. 2016, 94, 55–65. [Google Scholar] [CrossRef]
- Bachhawat, A.K.; Thakur, A.; Kaur, J.; Zulkifli, M. Glutathione transporters. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 3154–3164. [Google Scholar] [CrossRef]
- Russell, I. Chapter 4—Yeast. In Brewing Materials and Processes; Bamforth, C.W., Ed.; Academic Press: London, UK, 2016; pp. 77–96. [Google Scholar]
- Herdeiro, R.S.; Pereira, M.D.; Panek, A.D.; Eleutherio, E.C.A. Trehalose protects Saccharomyces cerevisiae from lipid peroxidation during oxidative stress. Biochim. Biophys. Acta Gen. Subj. 2006, 1760, 340–346. [Google Scholar] [CrossRef]
- Câmara Jr., A.A.; Maréchal, P.-A.; Tourdot-Maréchal, R.; Husson, F. Dehydration stress responses of yeasts Torulaspora delbrueckii, Metschnikowia pulcherrima and Lachancea thermotolerans: Effects of glutathione and trehalose biosynthesis. Food Microbiol. 2019, 79, 137–146. [Google Scholar] [CrossRef]
- Sprenger, J.; Hardeland, R.; Fuhrberg, B.; Han, S.-Z. Melatonin and other 5-methoxylated indoles in yeast: presence in high concentrations and dependence on tryptophan availability. Cytologia 1999, 64, 209–213. [Google Scholar] [CrossRef]
- Tan, D.-X.; Hardeland, R.; Manchester, L.C.; Korkmaz, A.; Ma, S.; Rosales-Corral, S.; Reiter, R.J. Functional roles of melatonin in plants, and perspectives in nutritional and agricultural science. J. Exp. Bot. 2012, 63, 577–597. [Google Scholar] [CrossRef]
- Germann, S.M.; Jacobsen, S.A.; Schneider, K.; Harrison, S.J.; Jensen, N.B.; Stahlhut, S.G.; Borodina, I.; Luo, H.; Zhu, J.; Maury, J. Glucose-based microbial production of the hormone melatonin in yeast Saccharomyces cerevisiae. Biotech. J. 2016, 11, 717–724. [Google Scholar] [CrossRef]
- Manfroi, L.; Silva, P.H.A.; Rizzon, L.A.; Sabaini, P.S.; Glória, M.B.A. Influence of alcoholic and malolactic starter cultures on bioactive amines in Merlot wines. Food Chem. 2009, 116, 208–213. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Ye, D.O.; Zhu, B.Q.; Wu, G.F.; Duan, C.Q. Rapid HPLC analysis of amino acids and biogenic amines in wines during fermentation and evaluation of matrix effect. Food Chem. 2014, 163, 6–15. [Google Scholar] [CrossRef]
- Tan, D.-X.; Hardeland, R.; Back, K.; Manchester, L.C.; Alatorre-Jimenez, M.A.; Reiter, R.J. On the significance of an alternate pathway of melatonin synthesis via 5-methoxytryptamine: comparisons across species. J. Pineal Res. 2016, 61, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Cruz, E.; Cerezo, A.; Cantos-Villar, E.; Troncoso, A.; García-Parrilla, M. Time course of l-tryptophan metabolites when fermenting natural grape musts: effect of inoculation treatments and cultivar on the occurrence of melatonin and related indolic compounds. Aust. J. Grape Wine R. 2019, 25, 92–100. [Google Scholar] [CrossRef]
- Rodriguez-Naranjo, M.I.; Gil-Izquierdo, A.; Troncoso, A.M.; Cantos-Villar, E.; Garcia-Parrilla, M.C. Melatonin is synthesised by yeast during alcoholic fermentation in wines. Food Chem. 2011, 126, 1608–1613. [Google Scholar] [CrossRef]
- Fernández-Cruz, E.; Alvarez-Fernández, M.A.; Valero, E.; Troncoso, A.M.; García-Parrilla, M.C. Melatonin and derived tryptophan metabolites produced during alcoholic fermentation by different yeast strains. Food Chem. 2017, 217, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Vilela, A. Use of Nonconventional Yeasts for Modulating Wine Acidity. Fermentation 2019, 5, 27. [Google Scholar] [CrossRef]
- Puertas, B.; Jiménez, M.J.; Cantos-Villar, E.; Cantoral, J.M.; Rodríguez, M.E. Use of Torulaspora delbrueckii and Saccharomyces cerevisiae in semi-industrial sequential inoculation to improve quality of Palomino and Chardonnay wines in warm climates. J. Appl. Microbiol. 2017, 122, 733–746. [Google Scholar] [CrossRef]
- Kim, D.; Kim, H.; Kim, K.; Roh, S. The Protective Effect of Indole-3-Acetic Acid (IAA) on H2O2-Damaged Human Dental Pulp Stem Cells Is Mediated by the AKT Pathway and Involves Increased Expression of the Transcription Factor Nuclear Factor-Erythroid 2-Related Factor 2 (Nrf2) and Its Downstream Target Heme Oxygenase 1 (HO-1). Oxid. Med. Cell Longev. 2017, 2017, 8639485. [Google Scholar] [CrossRef]
- Ehrlich, F. Uber Tryptophol (β-Indolyl-Athylalkohol), Ein Neues Gar Produkt der Hefe Aus Aminosäuren. Ber. Dtsch. Chem. Ges. 1912, 45, 883–889. [Google Scholar] [CrossRef]
- Dickinson, J.R. Nitrogen metabolism. In The Metabolism and Molecular Physiology of Saccharomyces cerevisiae, 2nd ed.; Dickinson, J.R., Schweizer, M., Eds.; CRC Press: London, UK, 2004; ISBN 0-415-29900-490000. [Google Scholar]
- Hazelwood, L.A.; Jean-Marc, D.; van Maris, A.J.A.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway fuel alcohol production: a century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Banach, A.; Ooi, B. Enhancing the Yields of Phenolic Compounds during Fermentation Using Saccharomyces cerevisiae Strain 96581. Food Nut. Sci. 2014, 5, 2063–2070. [Google Scholar] [CrossRef]
- Marsh, A.J.; Hill, C.; Ross, R.P.; Cotter, P.D. Fermented beverages with health-promoting potential: Past and future perspectives. Trends Food Sci Technol. 2014, 38, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Fijan, S. Microorganisms with claimed probiotic properties: an overview of recent literature. Int J. Environ. Res. Public Health. 2014, 11, 4745–4767. [Google Scholar] [CrossRef]
- Kozyrovska, N.O.; Reva, O.N.; Goginyan, V.B.; De Vera, J.-P. Kombucha microbiome as a probiotic: A view from the perspective of post-genomics and synthetic ecology. Biopolym. Cell. 2012, 28, 103–113. [Google Scholar] [CrossRef]
- Teoh, A.L.; Heard, G.; Cox, J. Yeast ecology of Kombucha fermentation. Int. J. Food Microbiol. 2004, 95, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Gachhui, R. Nitrogen-fixing, and cellulose-producing Gluconacetobacter kombuchae sp. nov., isolated from Kombucha tea. Int. J. Syst. Evol. Microbiol. 2007, 57, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Gachhui, R. Novel nitrogen-fixing Acetobacter nitrogenifigens sp. nov., isolated from Kombucha tea. Int. J. Syst. Evol. Microbiol. 2006, 56, 1899–1903. [Google Scholar] [CrossRef] [PubMed]
- Freire, A.L.; Ramos, C.L.; da Costa Souza, P.N.; Cardoso, M.G.B.; Schwan, R.F. Nondairy beverage produced by controlled fermentation with potential probiotic starter cultures of lactic acid bacteria and yeast. Int. J. Food Microbiol. 2017, 248, 39–46. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical Structures of the health-promoting compounds mentioned.

Figure 2.
Thiol redox regulation in the response of cells to oxidative stress conditions. Hgt1p (Glutathione transporters in the yeast S. cerevisiae [43,46]); AQP (aquaporin-mediated H2O2 diffusion transport); Glrl (glutathione reductase); Trr2 (Thioredoxin reductase 2, mitochondrial); Trx3 (Thioredoxin-3, mitochondrial, disulfide oxidoreductase activity) Adapted from Gostimskaya and Grant [45].
Figure 2.
Thiol redox regulation in the response of cells to oxidative stress conditions. Hgt1p (Glutathione transporters in the yeast S. cerevisiae [43,46]); AQP (aquaporin-mediated H2O2 diffusion transport); Glrl (glutathione reductase); Trr2 (Thioredoxin reductase 2, mitochondrial); Trx3 (Thioredoxin-3, mitochondrial, disulfide oxidoreductase activity) Adapted from Gostimskaya and Grant [45].

Figure 3.
Synthesis of melatonin and serotonin, as an intermediate compound, from tryptophan in yeast. Adapted from Mas et al. [19].
Figure 3.
Synthesis of melatonin and serotonin, as an intermediate compound, from tryptophan in yeast. Adapted from Mas et al. [19].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vilela, A. The Importance of Yeasts on Fermentation Quality and Human Health-Promoting Compounds. Fermentation 2019, 5, 46. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation5020046
AMA Style
Vilela A. The Importance of Yeasts on Fermentation Quality and Human Health-Promoting Compounds. Fermentation. 2019; 5(2):46. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation5020046
Chicago/Turabian StyleVilela, Alice. 2019. "The Importance of Yeasts on Fermentation Quality and Human Health-Promoting Compounds" Fermentation 5, no. 2: 46. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation5020046
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.