Nitrogen Sources Effect on Lactobacillus reuteri Growth and Performance Cultivated in Date Palm (Phoenix dactylifera L.) By-Products



Abstract
:1. Introduction
2. Materials and Methods
2.1. Date Extract Formulation
2.2. Preparation of Buffer Solution (BS)
2.3. Preparation of Protein Source
2.4. Preparation of MRS
2.5. Bacterial Culture Activation
2.6. Adaption Procedure
2.7. Inoculation Procedure
2.8. Determination of Bacterial Growth, Titratable Acidity, and Changing pH Values
2.9. Bacterial Enumeration
2.10. Statistical Analysis
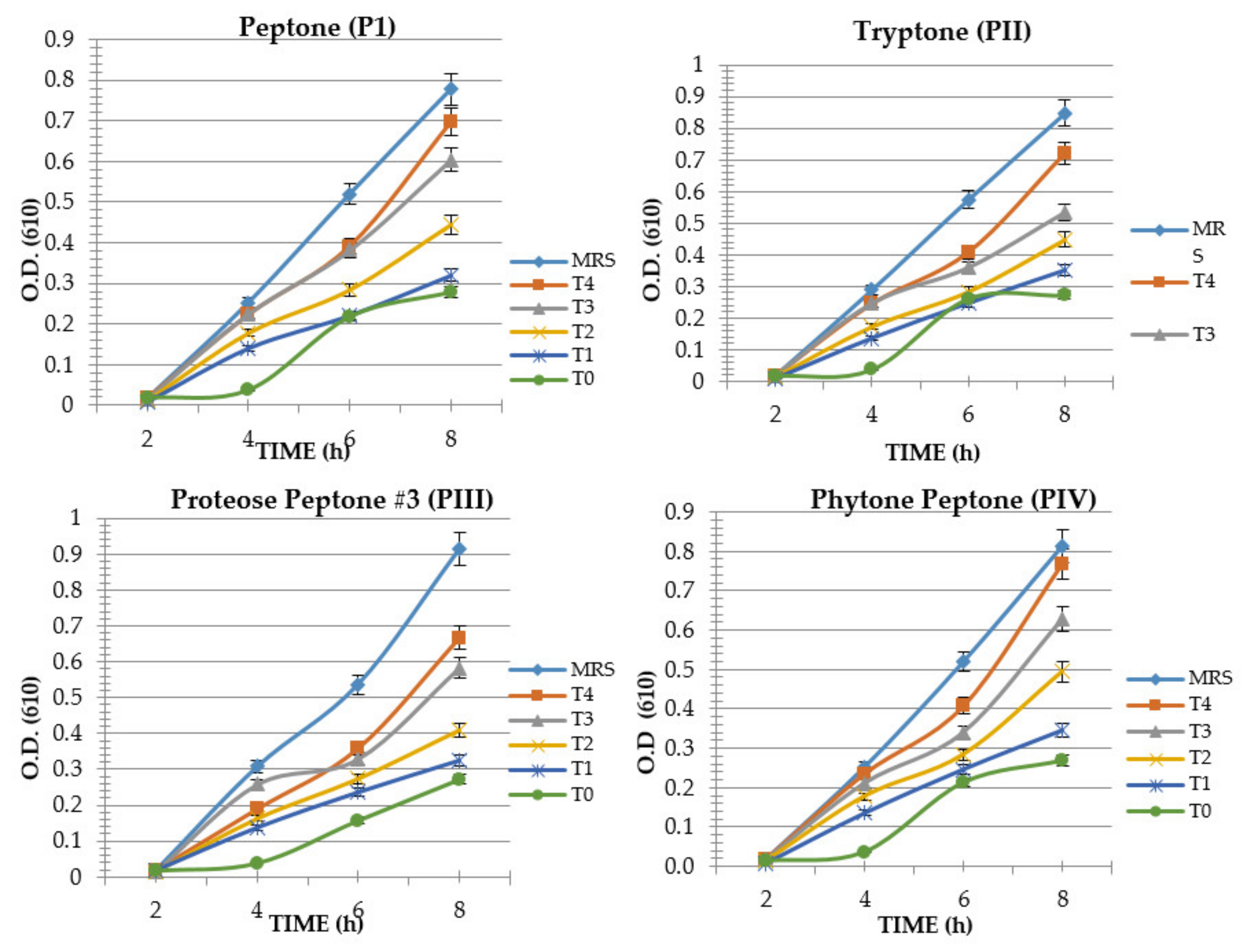
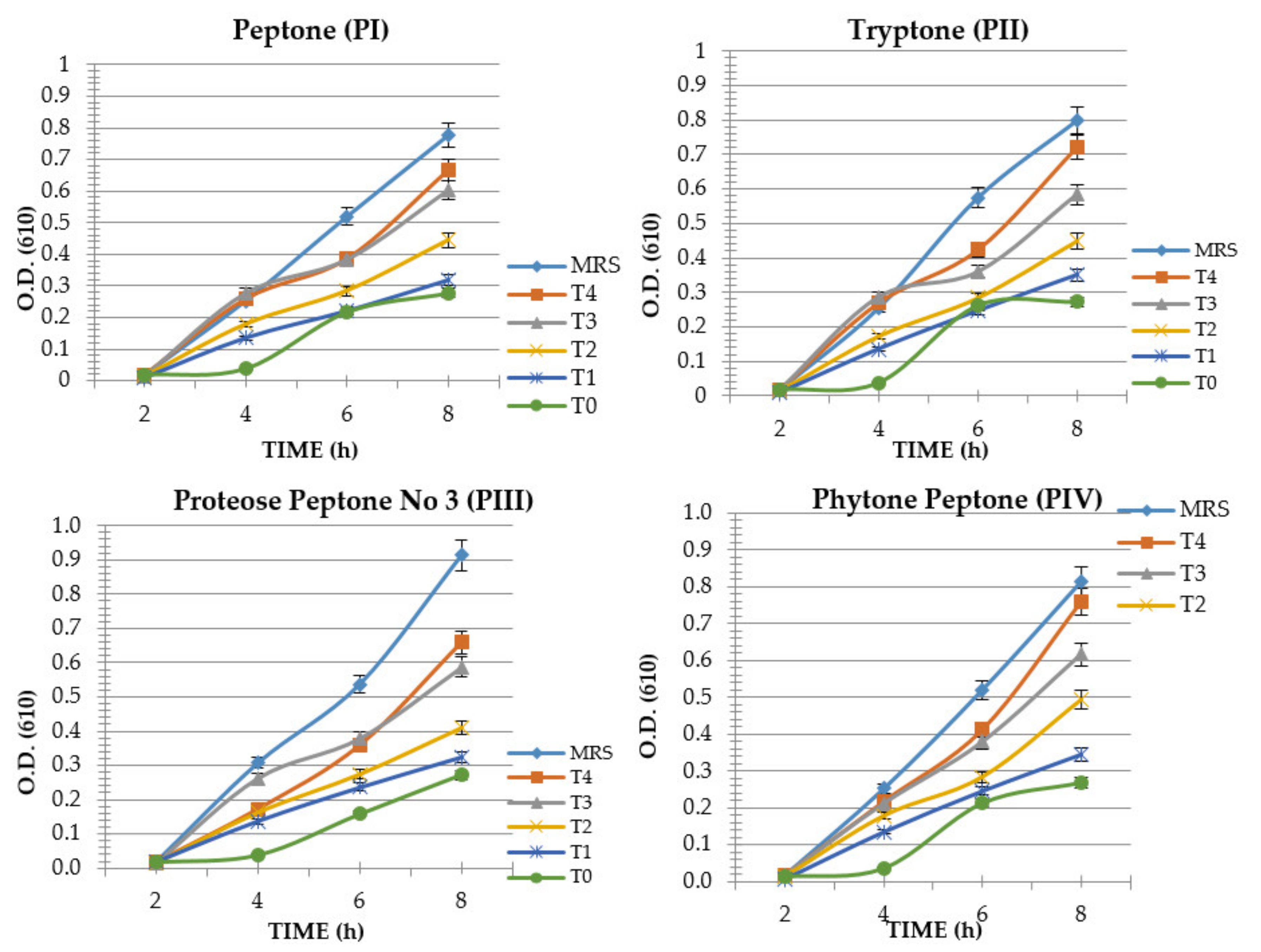
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ayad, A.A.; El-Rab, D.G.; El-Kherbawy, G.; Zaki, S.; Williams, L. Stimulating the Viability of Bifidobacterium spp. in Synbiotic Fermented Milk by Co-culturing with Lactobacillus paracasei 441 and Inulin. Int. J. Hortic. Agric. Food Sci. 2018, 2, 174–181. [Google Scholar] [CrossRef]
- Hébert, E.M.; Raya, R.R.; de Giori, G.S. Evaluation of Minimal Nutritional Requirements of Lactic acid Bacteria Used in Functional Foods, in Environmental Microbiology; Springer: Berlin, Germany, 2004; pp. 139–148. [Google Scholar]
- Ibrahim, G.; Mehanna, N.S.; El-Rab, D.G.; Abd-El-Salam, M.; Kholif, A.; Abdou, S.; El-Shibiny, S. Preparation and properties of set fermented milk containing inulin and different probiotics. In Proceedings of the 9th Egyptian Conference Dairy Science Technology, Cairo, Egypt, 9–11 October 2004; Egyptian Society of Dairy Science: Cairo, Egypt; pp. 117–132. [Google Scholar]
- Barakat, O.S.; Ibrahim, G.; Tawfik, N.; El-Kholy, W.; El-Rab, G.D. Identification and Probiotic Characteristics of Lactobacillus strains Isolated from Traditional Domiati Cheese. Int. J. Microbiol. Res. 2011, 3, 59–66. Available online: http://www.bioinfo.in/contents.php?id=27 (accessed on 27 May 2020).
- Lahtinen, S.; Ouwehand, A.C.; Salminen, S.; von Wright, A. Lactic Acid Bacteria: Microbiological and Functional Aspects; CRC Press: New York, NY, USA, 2011. [Google Scholar]
- Liu, M.; Bayjanov, J.R.; Renckens, B.; Nauta, A.; Siezen, R.J. The Proteolytic System of Lactic Acid Bacteria Revisited: A Genomic Comparison. BMC Genom. 2010, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Mills, D.A.; Block, D.E. Development of chemically defined media supporting high-cell-density growth of lactococci, enterococci, and streptococci. Appl. Environ. Microbiol. 2009, 75, 1080–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atilola, O.A.; Gyawali, R.; Aljaloud, S.O.; Ibrahim, S.A. Use of Phytone Peptone to Optimize Growth and Cell Density of Lactobacillus reuteri. Foods 2015, 4, 318–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomaa, E.Z. β-galactosidase from Lactobacillus delbrueckii and Lactobacillus reuteri: Optimization, characterization and formation of galactooligosaccharides. Ind. J. Biotechnol. 2018, 17, 407–415. [Google Scholar]
- Cotter, P.D.; Hill, C. Surviving the acid test: Responses of gram-positive bacteria to low pH. Microbiol. Mol. Biol. Rev. 2003, 67, 429–453. Available online: https://mmbr.asm.org/content/67/3/429.short (accessed on 27 May 2020). [CrossRef] [Green Version]
- Gao, C.; Ma, C.; Xu, P. Biotechnological routes based on lactic acid production from biomass. Biotechnol. Adv. 2011, 29, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Han, L.; Ji, Y.; Wang, X.; Tan, T. Fermentative production of L-lactic acid from hydrolysate of wheat bran by Lactobacillus rhamnosus. Biochem. Eng. J. 2010, 49, 138–142. [Google Scholar] [CrossRef]
- Coelho, F.; Augusto, M.; Lages, L.F. Contextual factors and the creativity of frontline employees: The mediating effects of role stress and intrinsic motivation. J. Retail. 2011, 87, 31–45. Available online: http://isiarticles.com/bundles/Article/pre/pdf/2248.pdf (accessed on 27 May 2020). [CrossRef]
- Khannous, L.; Souissi, N.; Ghorbel, B.; Jarboui, R.; Kallel, M.; Nasri, M.; Gharsallah, N. Treatment of saline wastewaters from marine-products processing factories by activated sludge reactor. Environ. Technol. 2003, 24, 1261–1268. [Google Scholar] [CrossRef]
- Hayek, S.A.; Shahbazi, A.; Awaisheh, S.S.; Shah, N.P.; Ibrahim, S.A. Sweet potatoes as a basic component in developing a medium for the cultivation of lactobacilli. Biosci. Biotechnol. Biochem. 2013, 77, 2248–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.X.; Shi, L.E.; Aleid, S.M. Date fruit: Chemical composition, nutritional and medicinal values, products. J. Sci. Food Agric. 2013, 93, 2351–2361. [Google Scholar] [CrossRef]
- Al-Farsi, M.A.; Lee, C.Y. Optimization of phenolics and dietary fibre extraction from date seeds. Food Chem. 2008, 108, 977–985. Available online: http://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/S0308814607012654 (accessed on 27 May 2020). [CrossRef]
- Juhaimi, F.A.; Ghafoor, K.; Özcan, M.M. Physical and chemical properties, antioxidant activity, total phenol and mineral profile of seeds of seven different date fruit (Phoenix dactylifera L.) varieties. Int. J. Food Sci. Nutr. 2012, 63, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, M.; Bahkali, A.H. Valorization of date palm (Phoenix dactylifera) fruit processing by-products and wastes using bioprocess technology—Review. Saudi J. Biol. Sci. 2013, 20, 105–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.-X.; Shi, L.-E.; Aleid, S.M. Date and their processing byproducts as substrates for bioactive compounds production. Braz. Arch. Biol. Technol. 2014, 57, 706–713. [Google Scholar] [CrossRef] [Green Version]
- Ayad, A.A. Survival and Viability of Freeze/Spray-Dried Lactobacillus ssp. Grown in Date Palm (Phoenix dactylifera L.) Base Medium; North Carolina Agricultural and Technical State University: Greensboro, NC, USA, 2016. [Google Scholar]
- Ayad, A.A.; El-Rab, D.G.; Shahbazi, A.; Worku, M.; Schimmel, K.; Ejimakor, G.; Zimmerman, T.; Ibrahim, S.A. Using Date Palm (Phoenix dactylifera L.) by-products to Cultivate Lactobacillus reuteri spp. J. Food Res. 2016, 5, 77. [Google Scholar] [CrossRef]
- Nancib, N.; Nancib, A.; Boudjelal, A.; Benslimane, C.; Blanchard, F.; Boudrant, J. The effect of supplementation by different nitrogen sources on the production of lactic acid from date juice by Lactobacillus casei subsp. rhamnosus. Bioresour. Technol. 2001, 78, 149–153. [Google Scholar] [CrossRef]
- Spinler, J.K.; Taweechotipatr, M.; Rognerud, C.L.; Ou, C.N.; Tumwasorn, S.; Versalovic, J. Human-derived probiotic Lactobacillus reuteri demonstrate antimicrobial activities targeting diverse enteric bacterial pathogens. Anaerobe 2008, 14, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Su, M.S.-W.; Oh, P.L.; Walter, J.; Gänzle, M.G. Intestinal origin of sourdough Lactobacillus reuteri isolates as revealed by phylogenetic, genetic, and physiological analysis. Appl. Environ. Microbiol. 2012, 78, 6777–6780. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Principle | Nutrient Value | |
|---|---|---|
| Energy | 307.12 Kcal | |
| Carbohydrates | 74.97 g | |
| Protein | 1.81g | |
| Total Fat | 0.15 g | |
| Dietary Fiber | 6.7 g | |
| Vitamins | Folates | 0.015 mg |
| Niacin | 1.61 mg | |
| Pantothenic acid | 0.805 mg | |
| Pyridoxine | 0.249 mg | |
| Riboflavin (B2) | 0.060 mg | |
| Thiamin (B1) | 0.050 mg | |
| Vitamin A | 0.16 mg | |
| Vitamin K | 0.0027 mg | |
| Minerals | Calcium | 64 mg |
| Copper | 0.36 mg | |
| Iron | 0.90 mg | |
| Magnesium | 54 mg | |
| Manganese | 0.296 mg | |
| Phosphorus | 62 mg | |
| Zinc | 0.44 mg | |
| Source | Description/Application | Suppliers |
|---|---|---|
| Peptone (PI) | Enzymatic digest of animal tissue, nitrogen in a form readily available to bacteria | Fisher Sci. |
| Tryptone (PII) | Pancreatic digest of casein is used as a nitrogen source for bacteria | Fisher Sci. |
| Proteose peptone No.3 (PIII) | Enzymatic digest of protein | BD Difco |
| Phytone peptone (PIV) | Papaic digest of soybean meal, designed specifically for cell culture applications, non-animal origin | BD Difco |
| Protein Source | ||||
|---|---|---|---|---|
| Growth Medium | PIV | PIII | PII | PI |
| DPM+0.0% (control) | 3.46 ± 0.1 E | |||
| DPM+0.2% | 3.45 ± 0.0 E | 3.51 ± 0.0 E | 3.35 ± 0.0 E | 3.42 ± 0.0 E |
| DPM+0.4% | 4.74 ± 0.0 D | 4.74 ± 0.0 C | 4.67 ± 0.0 C | 4.61 ± 0.0 D |
| DPM+0.6% | 6.79 ± 0.0 C | 5.64 ± 0.0 B | 5.78 ± 0.0 B | 5.27 ± 0.0 C |
| DPM+0.8% | 7.04 ± 0.0 B | 6.64 ± 0.0 B | 6.72 ± 0.0 B | 6.86 ± 0.0 B |
| Protein Source | ||||
|---|---|---|---|---|
| Growth Medium | PIV | PIII | PII | PI |
| DPM+0.0% (control) | 3.54 ± 0.1 D | |||
| DPM+0.2% | 3.72 ± 0.0 C | 3.53 ± 0.0 E | 3.51 ± 0.1 E | 3.52 ± 0.0 E |
| DPM+0.4% | 4.72 ± 0.1 C | 4.68 ± 0.1 D | 4.67 ± 0.1 D | 4.71 ± 0.1 D |
| DPM+0.6% | 6.83 ± 0.0 B | 6.58 ± 0.0 C | 5.48 ± 0.0 C | 6.78 ± 0.0 C |
| DPM+0.8% | 7.14 ± 0.0 B | 6.92 ± 0.0 B | 6.79 ± 0.1 B | 7.00 ± 0.0 B |
| Protein Source | ||||
|---|---|---|---|---|
| Growth Medium | PIV | PIII | PII | PI |
| DPM+0.0% (control) | 3.55± 0.1 E | |||
| DPM+0.2% | 3.48± 0.0 D | 3.57± 0.0 E | 4.51± 0.1 D | 3.52± 0.0 E |
| DPM+0.4% | 4.72± 0.1 D | 4.23± 0.1 D | 4.73± 0.1 C | 4.71± 0.1 D |
| DPM+0.6% | 6.78± 0.0 C | 6.58± 0.0 C | 5.64± 0.0 B | 5.6± 0.0 C |
| DPM+0.8% | 6.96± 0.0 B | 6.72± 0.0 B | 6.84± 0.1 B | 6.78± 0.0 B |
| Media | Nitrogen Sources g/L | Cost per Gram | Total Cost of Nitrogen Sources per Liter of Media |
|---|---|---|---|
| MRS | Proteose peptone No. 3 (10g) | $0.467/g | $4.67 |
| Beef Extract (10g) | $0.358/g | $3.58 | |
| Yeast extract (5g) | $0.194/g | $1.94 | |
| Total $10.19 | |||
| DPM | Phytone peptone (8g) | $0.186/g | Total $1.49 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayad, A.A.; Gad El-Rab, D.A.; Ibrahim, S.A.; Williams, L.L. Nitrogen Sources Effect on Lactobacillus reuteri Growth and Performance Cultivated in Date Palm (Phoenix dactylifera L.) By-Products. Fermentation 2020, 6, 64. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030064
Ayad AA, Gad El-Rab DA, Ibrahim SA, Williams LL. Nitrogen Sources Effect on Lactobacillus reuteri Growth and Performance Cultivated in Date Palm (Phoenix dactylifera L.) By-Products. Fermentation. 2020; 6(3):64. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030064
Chicago/Turabian StyleAyad, Amira A., Deiaa A. Gad El-Rab, Salam A. Ibrahim, and Leonard L. Williams. 2020. "Nitrogen Sources Effect on Lactobacillus reuteri Growth and Performance Cultivated in Date Palm (Phoenix dactylifera L.) By-Products" Fermentation 6, no. 3: 64. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030064