The Effect of Non-Saccharomyces and Saccharomyces Non-Cerevisiae Yeasts on Ethanol and Glycerol Levels in Wine



1
Perennia Food and Agriculture Inc., 32 Main Street, Kentville, NS B4N 1J5, Canada
2
Office of Industry and Community Engagement, Acadia University, 210 Horton Hall, 18 University Ave, Wolfville, NS B4P 2R6, Canada
3
Tasmanian Institute of Agriculture, University of Tasmania, Prospect, TAS 7249, Australia
4
Cool Climate Oenology and Viticulture Institute (CCOVI), Brock University, 1812 Sir Isaac Brock Way, St. Catharines, ON L2S 3A1, Canada
5
Department of Biological Science, Faculty of Maths and Science, Brock University, 1812 Sir Isaac Brock Way, St. Catharines, ON L2S 3A1, Canada
*
Author to whom correspondence should be addressed.
Fermentation 2020, 6(3), 77; https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030077
Submission received: 21 May 2020
/
Revised: 20 July 2020
/
Accepted: 28 July 2020
/
Published: 30 July 2020
(This article belongs to the Special Issue Enological Repercussions of Non-Saccharomyces Species 2.0)
Abstract
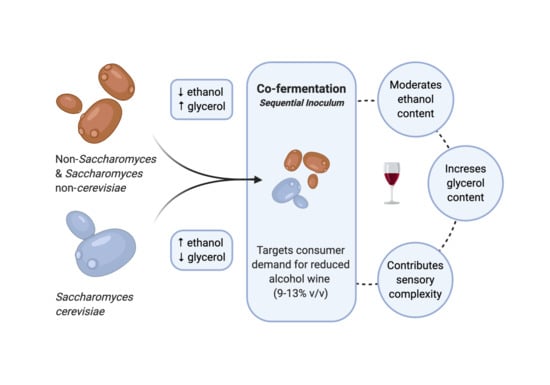
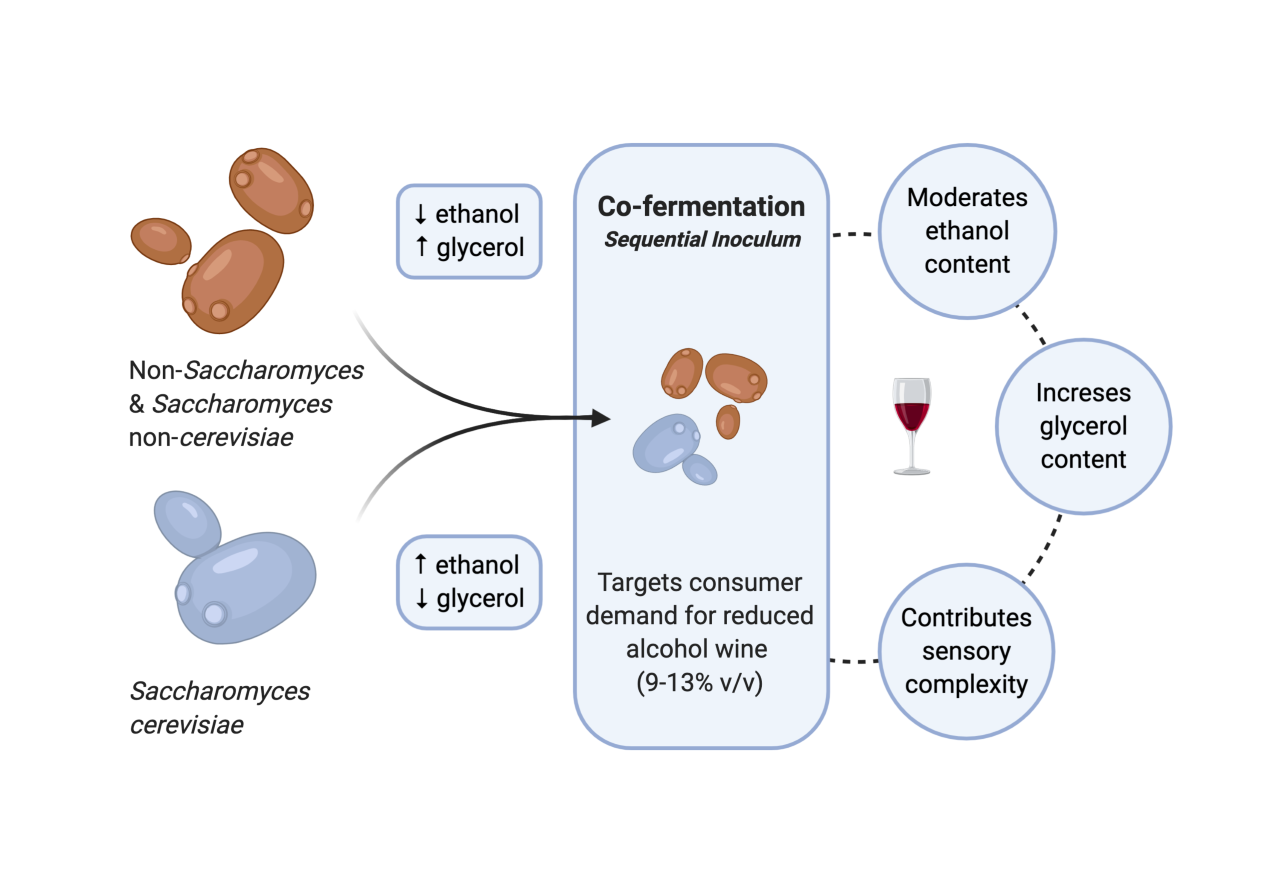
:Non-Saccharomyces and Saccharomyces non-cerevisiae studies have increased in recent years due to an interest in uninoculated fermentations, consumer preferences, wine technology, and the effect of climate change on the chemical composition of grapes, juice, and wine. The use of these yeasts to reduce alcohol levels in wines has garnered the attention of researchers and winemakers alike. This review critically analyses recent studies concerning the impact of non-Saccharomyces and Saccharomyces non-cerevisiae on two important parameters in wine: ethanol and glycerol. The influence they have in sequential, co-fermentations, and solo fermentations on ethanol and glycerol content is examined. This review highlights the need for further studies concerning inoculum rates, aeration techniques (amount and flow rate), and the length of time before Saccharomyces cerevisiae sequential inoculation occurs. Challenges include the application of such sequential inoculations in commercial wineries during harvest time.
1. Introduction
After carbon dioxide (CO2), ethanol and glycerol are the most abundant compounds produced during alcoholic fermentation. The levels of ethanol and glycerol in wine depend upon many factors, such as seasonal events affecting the concentration of grape sugar, and winemaking decisions, including fermentation conditions and fermenting yeasts [1].
At a commercial scale, inoculations with Saccharomyces cerevisiae strains are often preferred over those with non-Saccharomyces or S. non-cerevisiae yeasts, because the latter is considered responsible for incomplete fermentations (and consequently high levels of residual sugar in wine), and they produce high concentrations of acetic acid and ethyl acetate [2,3]. Nevertheless, non-Saccharomyces or S. non-cerevisiae yeasts are important to winemakers, particularly to those who target wines with unique sensory characters that are popularly recognised as typical of their geographical origin or variety [4,5,6,7]. These yeasts are also popular among winemakers who choose to produce less alcoholic wines [8].
Although non-Saccharomyces and S. non-cerevisiae yeasts are sought after for their specific oenological characteristics, it is a challenge for some of these yeasts to conduct a complete fermentation to a desired level of dryness. This is very important to winemakers, in part because finished wines with higher levels of residual sugars above 0.5 g/L require high doses of sulfur dioxide (SO2) to ensure their microbial stability to prevent wine spoilage. Therefore, inoculations of non-Saccharomyces/S. non-cerevisiae in mixed cultures with S. cerevisiae strains, which have higher fermentation rates, have been studied to ensure complete fermentation.
The strategy of using selected mixed cultures for alcoholic fermentation is believed to be the key to produce wines with desirable characteristics that meet changing market demands with less ethanol but still with flavours comparable to standard wines [9]. This strategy is carried out by two different methods of inoculation: (1) co-inoculation, which involves concurrent inoculations of non-Saccharomyces/S. non-cerevisiae yeasts at high cell concentration (e.g., 107 cell/mL) with S. cerevisiae; and/or (2) sequential inoculation, which involves inoculating non-Saccharomyces/S. non-cerevisiae yeasts to start the fermentation and continue for a determined amount of time alone, and inoculating S. cerevisiae to take over and complete the fermentation [9,10]. The time period before carrying out the sequential S. cerevisiae inoculation and the Saccharomyces/non-Saccharomyces or Saccharomyces non-cerevisiae inoculum ratio are both important parameters that affect the fermentation kinetics and oenological outcomes, and the former generally varies between 1 and 3 days [11,12,13,14].
Many reviews have studied different perspectives of non-Saccharomyces/S. non-cerevisiae yeasts for modern winemaking practices, including their influence on different wine quality parameters with an emphasis on traits such as the primary (or varietal) and secondary (or fermentative) aromas of wines, acidity, freshness, as well as specific styles of wines (such as traditional method sparkling wines and red table wines) [3,9,15,16,17,18,19,20,21].
The aim of this review is to highlight those studies that have shown a direct link between the use of non-Saccharomyces or S. non-cerevisiae and the concentration of ethanol and glycerol in wine or synthetic media. The first part of this review provides an overview of ethanol and glycerol as contributors to wine sensory characteristics, and a general overview of non-S. cerevisiae or S. non-cerevisiae yeasts. The second part of this review provides more specific details of individual non-S. cerevisiae or S. non-cerevisiae species that are relevant to the wine industry. We conclude this review by suggesting what additional research might help winemakers have greater control over wine quality outcomes.
2. Ethanol Reduction
Ethanol is produced by yeast during the alcoholic fermentation and is generally found in the range of 11.5–15% v/v in wines. It is an important wine component that directly effects organoleptic properties, aging, and wine stability [22]. The impact of ethanol on the sensory profile of wines and other alcoholic beverages has been recently reviewed [23]. Ethanol influences taste and mouthfeel sensations, alters the sensation of sweetness, increases bitterness, decreases sourness, and contributes to the hotness sensation and body of the wine [24,25,26,27]. Ethanol can also decrease the volatility of aroma compounds by increasing their solubility in the wine [28], making small compounds such as fruity-driven ethyl esters and acetates less recognisable by human senses.
According to the International Organisation of Vine and Wine (OIV), the alcohol strength of wines must be a minimum of 8.5% v/v, although in cool climate wine regions, this value can be lowered to 7% v/v [29]. Over the past two decades, ethanol content in wines has been noticeably increasing in some regions by 0.1–1% per year [30,31]. Apart from hotter climates leading to higher sugar berry levels at harvest and therefore, higher alcohol contents in wine [32], one of the main reasons behind this progressive increase is consumer demand for specific wine styles, which are described as rich, well-structured, with a flavour profile dominated by dark, ripe fruits [33]. This style requires optimal grape maturity and higher sugar content of 240 g/L or more [34].
Nonetheless, an increasing trend for reduced alcohol in beverages (broadly defined as containing 9% to 13% v/v ethanol), and low-alcohol (0.5–2% v/v) wines by consumers has been recently observed [35,36]. Increasing health and safety consciousness and global initiatives towards moderating alcohol consumption are reasons for producing lower alcohol wines that appeal to wine drinkers [37]. Wines may also be subjected to higher taxation depending on their alcohol content, which increases the final cost of wines to the consumer [38]. Since ethanol is the main source of caloric content in wine, there is also a risk of a negative impact on wine export to countries where health labeling of foods and beverages served at restaurants is voluntary or mandatory [39].
During winemaking, high sugar and therefore ethanol can cause sluggish and stuck alcoholic fermentations and can be challenging for successful malolactic fermentations [40,41]. As a means to address these issues, methods have been studied that include lowering the final ethanol content of wine using a wide selection of interventions. These can be grouped into (1) pre- (e.g., viticultural, juice dilution, and fermentation of early harvest fruit); (2) concurrent (e.g., non-Saccharomyces yeasts, modified yeasts, and arrested fermentation) [42,43]; and (3) post-fermentation (e.g., non-membrane and membrane ethanol removal) [43,44,45]. The use of microbiological approaches such as inoculation with non-Saccharomyces yeasts for producing wines with less ethanol is a promising alternative to the removal of ethanol by membrane based-approaches [11,46,47,48]. The advantages associated with the use of low-ethanol/high-glycerol yielding yeasts include their relatively easy application and lower costs when compared to more expensive and less eco-friendly approaches, such as membrane contactors, nanofiltration, or the spinning cone column [49,50]. However, it is important to acknowledge that low-ethanol/high-glycerol yielding yeasts are much less effective than the membrane-based processes in terms of ethanol reductions [44], and they are perhaps only suitable when winemakers want to achieve a reduction in ethanol content by up to 3.0% v/v [49].
Inoculations with non-Saccharomyces yeasts can be used as a strategy to produce lower alcohol wines due to the yeasts′ different sugar utilisation pathways, including respiration, alcoholic fermentation, and glycerol–pyruvic metabolisms, and different regulatory mechanisms, in comparison to S. cerevisiae [51]. While the theoretical sugar-to-ethanol yield for a complete fermentation by S. cerevisiae generally ranges from 90% to 95%, the residual sugar is consumed via alternative metabolic pathways and biomass biosynthesis [52]. On the other hand, ethanol yield and the by-products formed vary immensely amongst non-Saccharomyces yeasts [23]. For example, due to the Crabtree effect, S. cerevisiae prefers fermentation metabolism rather than respiration when the sugar amount exceeds 10 g/L [40]. In contrast, among non-Saccharomyces yeasts, there are strains and species that can consume sugar with aerobic respiration regardless of sugar concentration [53,54] without contributing significantly to the final ethanol level of the wine. Therefore, non-Saccharomyces yeasts have been studied under partial and controlled aeration strategies during fermentation to achieve lower ethanol by allowing part of the sugar to be consumed via respiration rather than alcoholic fermentation [47,55]. However, an increase in undesirable volatile compounds, such as acetic acid and ethyl acetate, are the main limiting factors of the aeration strategies that require the application of a proper aeration regime [55,56,57].
As a response to interest in reduced alcohol levels in wines, researchers studied non-S. cerevisiae and S. non-cerevisiae yeast species [49,56,58,59,60] (Table 1). Several non-Saccharomyces yeast strains were identified, including Metschnikowia pulcherrima, and two species of Kluyveromyces, which have the capacity to decrease ethanol yields by respiration [59]. M. pulcherrima AWRI 1149 was identified as a potential yeast to produce wine with a reduced ethanol concentration, having been identified following the evaluation of 50 non-Saccharomyces isolates under limited aeration conditions, and in sequential inoculations with S. cerevisiae [49]. A similar study with 48 non-Saccharomyces yeast isolates identified Torulaspora delbrueckii AWRI 1152 and Zygosaccharomyces bailii AWRI 1578 yeasts as suitable for reducing ethanol [56]. More recently, the respiratory, fermentative, and physiological characteristics of 114 non-Saccharomyces yeasts were evaluated [60]. Taking into account their ability to reduce ethanol content Hanseniaspora uvarum BHu9 and BHu11, Hanseniaspora osmophila BHo51, Starmerella bacillaris (synonym. Candida zemplinina) BSb55, and Candida membranaefaciens BCm71 were selected as candidates for co-fermentations.
3. Glycerol
Glycerol is the most abundant yeast metabolism by-product after ethanol and CO2. It is produced from dihydrodroxyacetone phosphate, which is first reduced to glycerol-3-phosphate via glycerol-3-phosphate dehydrogenase (GPDH), and then converted into glycerol by a specific phosphatase. This is a non-volatile 3-hydroxy alcohol, which is a polyol also known as a sugar alcohol. Glycerol is a viscous liquid at room temperature and appears to contribute to mouthfeel and viscosity at, or above 28 g/L and sweetness in the range of 5–12 g/L [42,61,62,63]. However, Nieuwoudt, et al. [64] did not find any link between wine quality (expressed as the number of medals received at a wine competition) and concentrations of glycerol in wine, and Goold et al. [42] concluded in their review that glycerol had only a minimal influence on the viscosity of wine.
The synthesis of glycerol and acetic acid, in addition to ethanol, are both linked to redox balance [22,42]. The significance of glycerol synthesis to redox balance has been suggested to be due to the inability of mutants (unable to synthesise glycerol) to grow in anaerobic conditions [22,65]. During the stationary phase of yeast during fermentation, glycerol synthesis has been found to be associated with redox balance by removing excess reducing power [22,66].
Many factors can influence the production of glycerol, which is in general more abundant in wines fermented with non-Saccharomyces than those fermented with S. cerevisiae [67,68,69], and in red wine (approximately 10.5 g/L) compared to white wines (approximately 7 g/L) [64]. Glycerol is generally more abundant in red wines in part because red juice typically ferments at higher temperatures (20–25 °C) than white wines (< 20 °C). Yet, fermenting temperatures positively influence the production of glycerol by yeasts, and non-Saccharomyces and S. non-cerevisiae are not an exception. For example, increasing the fermentation temperature from 16 to 20 °C increased the glycerol content from 1.69 to 3.04 g/L in co-fermentations of Candida stellata and S. cerevisiae [52]. A significant increase in the glycerol content of a grape juice was also observed after increasing the temperature from 12 to 25 °C, with fermentations carried out by pure Saccharomyces paradoxus reporting an increment of approximately 2.5 g/L, for example [12]. The sugar level of grapes at harvest (and therefore in the juice) also influences the production of glycerol, because this compound is accumulated by yeast to combat dehydration by balancing the intracellular osmolarity with that of the medium [1]. This effect has become even more evident in recent times due to hotter seasonal temperatures compressing the ripening windows of different grape varieties in warm climates. This has meant that winemakers delay the harvest date because of wineries operating at full capacity, thereby causing a part of the crop to overripe in the vineyard [70]. Hranilovic, et al. [71] reported that the glycerol content of a Shiraz wine from early harvest grapes (approximately 265 g/L TSS) was much higher than those from the late harvest (approximately 325 g/L TSS), with a M. pulcherrima strain (followed by a S. cerevisiae inoculum), producing early and late harvest wines with 10.51 and 12.59 g/L glycerol, respectively. Juice with high sugar concentration also leads to an excess in acetic acid, which can be explained by yeasts trying to maintain redox balances by using surplus NAD(P)+ accumulated during the synthesis of fermentation metabolites [62,72,73,74].
The growth of non-Saccharomyces yeast species such as Lachancea thermotolerans, T. delbrueckii, and M. pulcherrima strictly depends upon oxygen availability [57,75,76]. When the effect of oxygen availability on glycerol production by non-Saccharomyces was evaluated, oxygenation at three dissolved oxygen levels of 0.08, 0.41, and 1.71 mg/L resulted in glycerol reduction [76]. This was evident for a T. delbrueckii strain in co-fermentation with S. cerevisiae, with the glycerol content decreasing from 6.79 g/L in the T. delbrueckii anaerobic treatment up to 1.09 g/L in the co-inoculation treatment with 1.71 mg/L of dissolved oxygen. Different results were reported by Morales et al. [57], who observed increased glycerol yields for M. pulcherrima/S. cerevisiae mixed cultures under controlled oxygenation conditions (sparged with pure air, nitrogen, or mixtures of both) during the first 48 h of fermentation, and anaerobic conditions thereafter.
The non-Saccharomyces yeasts, which have the capacity to redirect the sugar consumption for the production of alternative compounds, rather than ethanol, have been studied in wines with reduced ethanol content. These alternative compounds could be glycerol and pyruvic acid produced via glycerol–pyruvic metabolisms. Alternatively, before sugar is utilised during alcoholic fermentation, sugars can be consumed via respiratory metabolism [40], which is the case with various non-Saccharomyces yeasts with lower Crabtree effect.
Other factors can have an impact on the formation of glycerol by non-Saccharomyces. These include the concentration of nitrogen and sulfites [8]. Limited nitrogen concentrations in the must (in the form of amino acids and ammonium) can lead to a significant increase of glycerol production. By contrast, higher levels of sulfur dioxide lead to higher levels of glycerol [30]. Increased glycerol production has been found to be linked to increased acetic acid, which is easily detected due to its vinegar smell [3,18,77].
Upon a thorough literature review, we identified five non-Saccharomyces yeasts that have been widely studied due to their different oenological traits; Schizosaccharomyces pombe, Metschnikowia pulcherrima, Lachancea thermotolerans, Candida stellata, and Torulaspora delbrueckii. These five non-Saccharomyces yeasts have different pathways that result in ethanol reduction and glycerol production while influencing different parameters of the chemical composition of wines. The following sections of this review present current knowledge regarding the effect of alternative yeasts that influence ethanol and glycerol concentrations while highlighting gaps in our knowledge that require further research.
4. Schizosaccharomyces Pombe
Schizosaccharomyces pombe is a widely studied yeast due to its particular ability to moderate wine acidity via malic acid degradation [78]. Among its other promising traits is the ability to enhance the color of red wines and reduce Ochratoxin A, biogenic amines, and ethyl carbamate [79,80,81]. The most detrimental metabolites produced by spoilage yeasts in pure culture or spoiled juices have been found to decease in mixed fermentations carried out at the laboratory scale [2,82]. The main characteristics of S. pombe and its application in winemaking were reviewed recently [83,84]. S. pombe is commercially available as an alternative method to de-acidity wine [19].
Unlike some non-Saccharomyces yeast species, S. pombe is capable of fermenting wines up to comparable concentrations of Saccharomyces, in the range of 10–15% v/v ethanol, depending on the strain and presence of aeration [85]. Malo-alcoholic fermentation [86] and the glycerol–pyruvic pathway [87] observed in S. pombe inoculations have different impacts on the final ethanol content of the wines. Therefore, while some studies reported an ethanol reduction with inoculations involving S. pombe [87], others have reported no difference or even an increase [88,89,90,91].
The use of S. pombe (strain 938) for white winemaking was investigated by Benito et al. [87], with sole, mixed, and sequential fermentations in conjunction with S. cerevisiae (Cru Blanc). All the strains in the study were able to ferment the wines to dryness. The sole fermentation of S. pombe showed 0.65% v/v lower ethanol compared to S. cerevisiae alone, values being 13.18% v/v and 12.53% v/v, respectively. Similar results were obtained with the sequential inoculation, with 0.4% v/v lower ethanol value in comparison to the control treatment [87]. Loira et al. [88] studied the effect on Syrah wine sensory quality of S. pombe strains (938, V1 and 4.2) in mixed and sequential fermentations with S. cerevisiae strain 7VA. S. pombe was not used as a sole inoculum in this study. Ethanol reduction did not occur in the mixed or sequential fermentations with S. pombe and S. cerevisiae compared to treatments with a sole inoculum of S. cerevisiae. In this study, slightly higher ethanol values that were not statistically significant were reported for mixed and sequential fermentations with S. pombe strains (13.2% to 13.5% v/v) compared to S. cerevisiae (13.2% v/v) [88]. Although the same strains of S. pombe (938) were used in these studies, different delay times were applied for the sequential inoculation of S. cerevisiae (48 h versus 7 days), and different strains of S. cerevisiae were used for the comparison.
Increased ethanol levels have been reported in studies that used S. pombe where juices had high malic acid content. S. pombe (Y0119) was used in a sequential inoculation with S. cerevisiae (NT116) to ferment Kei-apple (Dovyalis caffra L.) juice, which contained high malic acid (45 g/L). An increase in ethanol level (6.08% v/v) was reported compared to sole inoculation with S. cerevisiae (4.67% v/v) [91]. Similarly, slightly higher ethanol values (approximately 0.2% to 0.5% v/v) were reported in a study where S. pombe was used in sequential inoculation with S. cerevisiae for fermenting plum juice (Prunus domestica L.), compared to S. cerevisiae as a control [90]. Studies that included S. pombe that measured glycerol reported that the yeast produced lower levels of glycerol than the other yeasts in the study (Table 2) [20].
5. Metschnikowia Pulcherrima
Metschnikowia pulcherrima is a non-Saccharomyces yeast that is commercially available from many manufacturers [10,19]. Its ability to enhance varietal aroma compounds [50,92] and reduce the ethanol content of wines has raised interest in its commercial use [93,94]. Recently, the impact of M. pulcherrima in winemaking has been reviewed [95].
The ability of M. pulcherrima for withstanding ethanol concentrations of up to 7% v/v has been reported by Combina et al. [96]. Recently, four strains of M. pulcherrima were identified as being able to actively grow at a higher ethanol concentration (9% v/v) [97]. Sixty-two of the 65 strains isolated from the Douro region of Portugal in this study were able to tolerate a 6% v/v ethanol level. On the other hand, its ability to ferment was reported up to levels of 4% v/v in micro-fermentations conducted in pasteurised grape must [13]. Consequently, different co-inoculation and sequential inoculation strategies have been studied [14,57,93].
Contreras et al. [49] identified a M. pulcherrima strain that can reduce the ethanol content of wine through part of a sequential inoculation with S. cerevisiae. Lower ethanol levels of 0.9% and 1.6% v/v were achieved in Chardonnay and Shiraz wines, respectively, compared to a sole inoculum of S. cerevisiae, which found ethanol levels of 15.1% v/v in Chardonnay and 13.8% v/v in Shiraz wines. A similar result of 0.9% v/v ethanol reduction was reported in a study carried out with the sequential inoculation of M. pulcherrima with S. cerevisiae compared to a sole inoculum of S. cerevisiae that produced ethanol levels of 13.2% v/v [93]. Furthermore, a mixed inoculum of M. pulcherrima and S. uvarum with sequential inoculation of S. cerevisiae was conducted. Two different M. pulcherrima/S. uvarum inoculum ratios were applied (1 × 106 cells/mL to 1 × 105 cells/mL, and 1 × 106 cells/mL to 1 × 104 cells/mL). Both resulted in an ethanol reduction of 1.7% v/v, along with higher concentrations of succinic acid and glycerol, compared to wine fermented with S. cerevisiae [93]. Varela et al. [50] studied M. pulcherrima and S. uvarum, both of which were found to be able to produce wines with reduced ethanol content. Along with the ethanol reduction, the sensory profile and volatile aromatic composition of Merlot wines were studied. Fermentation was conducted via co-inoculation using M. pulcherrima (1 × 106 cells/mL) and S. cerevisiae (1 × 105 cells/mL). An ethanol reduction of 1.0% v/v, along with higher concentrations of ethyl acetate, total esters, total higher alcohols, and total sulfur compounds were detected in wines fermented with a co-inoculation of M. pulcherrima and S. cerevisiae, compared to wines fermented with S. cerevisiae. Although a higher production of ethyl acetate and total sulfur-containing compounds was detected, the sensory panel did not detect associated negative attributes in the wines [50].
The effect of different aeration regimes and immobilisation on the ethanol reduction with selected strains of M. pulcherrima has been studied [98]. An ethanol reduction of 1.38% v/v was achieved in first 72 h of fermentation of Verdicchio must with M. pulcherrima under an aeration flow of 20 mL/L/min, compared to the control that used S. cerevisiae [98]. A blend of Malvasia and Viura (Macabeo) must was fermented in a study by Morales et al. [57] using a mixed culture of M. pulcherrima and S. cerevisiae, and using different aeration regimes (sparged with air or nitrogen). The lowest ethanol values were reported for the treatments sparged with air and fermented with the mixed culture of M. pulcherrima and S. cerevisiae, regardless of the inoculum level of S. cerevisiae (1% or 10%). In these treatments, 11% v/v ethanol was obtained, compared to 14.7% v/v and 12.9% v/v ethanol in the treatments fermented with S. cerevisiae sparged with nitrogen and air, respectively. However, high acetic acid values (higher than 0.65 g/L) obtained under air-sparged treatment produced wines that would have been unacceptable for consumers and do not meet market regulations were reported. In contrast, treatments sparged with nitrogen and fermented with the mixed culture of M. pulcherrima and S. cerevisiae had an ethanol reduction of 0.8% v/v compared to those fermented with S. cerevisiae, with acceptable levels of acetic acid (lower than 0.1 g/L) [57]. The same level of ethanol reduction was achieved (0.8% v/v) using a sequential fermentation with Viura/Macabeo and Malvasia varieties using M. pulcherrima with S. cerevisiae compared to the control (S. cerevisiae) under non-aerated conditions [99].
The ability of selected immobilised non-Saccharomyces yeasts (Starmerella bombicola (formerly named Candida stellata), Metschnikowia pulcherrima, Hanseniaspora osmophila, and Hanseniaspora uvarum) to reduce the ethanol content in wine via sequential fermentation has been also studied [14]. In synthetic grape juice, the sequential inoculation of M. pulcherrima had 1.14% to 1.35% v/v, with 48 and 72 h delays in inoculation of S. cerevisiae, respectively. Ethanol concentration was reduced by 1.10% to 1.46% v/v in natural grape juice in the same study [14]. Similarly, Röcker et al. [47] studied five non-Saccharomyces yeast strains including M. pulcherrima for a sequential inoculation with S. cerevisiae var. bayanus strain under three different aeration conditions (aeration with sterile pressured air during 15 and 5 days, and under regulated oxygen content of 20% dissolved oxygen), to ferment Riesling must. Ethanol concentration was reduced by up to 3.8% v/v within 3 days of aeration, although the authors reported an increase in vinegar (associated with acetic acid) and oxidation sensory attributes [47].
Three non-Saccharomyces yeast strains (M. pulcherrima, T. delbrueckii, and Zygosaccharomyces bailii) fermented Chardonnay using sequential inoculation with S. cerevisiae under three different aeration conditions (no air addition, 5 mL/min aeration (0.025 VVM), 10 mL/min aeration (0.05 VVM)). The authors reported that the sequential inoculation with M. pulcherrima reduced alcohol by 1.6% v/v, which was the highest ethanol reduction among all the non-Saccharomyces yeast strains in the study, compared to S. cerevisiae [43]. In this study, applied aeration conditions did not cause an increase in the acetic acid production. However, in the wines produced with a sequential inoculation of M. pulcherrima and S. cerevisiae, an over-production of ethyl acetate (280 mg/L) was detected under the aeration regime of 0.05 VVM. This indicates an oxygen threshold for the over-production of this compound [43], which causes unpleasant odors such as nail polish remover and vinegar [100].
The ability of M. pulcherrima to reduce the final ethanol content via its respiratory characteristic has been shown with various studies (Table 1). Recently, M. pulcherrima has been reported to produce lower levels of glycerol under semi-anaerobic conditions than when the air flow into the fermentations was 1 mL/L/min and 20 mL/L/min (Table 2). Therefore, taken together in respect to ethanol and glycerol content, future research with M. pulcherrima (solo, sequential, or co-fermentations) should include a range of aeration strategies, a range of inoculum rates, and an investigation into the length of time before inoculation of S. cerevisiae yeast, to render its application feasible at a commercial winemaking scale to achieve ethanol reduction without compromising or enhancing sensory characteristics [14,43,47,59,93,97].
6. Lachancea Thermotolerans
Lachancea thermotolerans (previously Kluyveromyces thermotolerans) is available commercially from yeast manufacturers [19,101]. This yeast has specific oenological traits including a positive influence on wines’ sensory profile [102] and total acidity [13,103]. The main characteristics of L. thermotolerans and its effects on winemaking were recently reviewed [101,104,105,106].
L. thermotolerans in pure culture was able to reach 10.46% v/v ethanol levels in micro-fermentations of pasteurised commercial white wine must, conducted at 25 °C, by leaving more than 50 g/L residual sugar. The control wine inoculated with a pure culture of S. cerevisiae EC-1118 fermented the must to dryness (less than 6 g/L residual sugar), reaching an ethanol level of 13.04% v/v [102]. The study continued at industrial scale, and an ethanol reduction of 0.7% v/v was achieved with a Sangiovese must [102]. In the study by Del Fresno et al. [89], a higher ethanol reduction of 1.2% v/v was achieved. This was compared to a different S. cerevisiae strain (7VA), which was sequentially inoculated later than the previous study (6 days), with a higher inoculum amount (both at 108 cell/mL). The laboratory-scale fermentation using Tempranillo must conducted at 25 °C and 12.66% v/v ethanol level used sequential inoculation, while sole inoculation of S. cerevisiae 7VA was 13.84% v/v [89].
As well as its ability to decrease the pH of wine, the metabolic pathway of L. thermotolerans that can turn sugars into lactic acid is also described as a way to reduce the level of alcohol in wines [18]. A study conducted using micro-vinifications of Airén must conducted at 25 °C [80] with L. thermotolerans 617 (100 mL containing 2.27 × 107 CFU/mL) followed by the sequential inoculation of S. cerevisiae 87 (107 CFU/mL of) with 96 h of delay, an ethanol reduction of 0.4% v/v was achieved, compared to the sole inoculation of S. cerevisiae 87. This sequential inoculation also showed a higher lactic acid production (3.18 g/L) and lower pH (3.52, compared to 3.74 in control wine). The sensory panel perceived this treatment to have better sensorial properties, with higher scores for sweetness, despite similar levels of residual sugar compared to control. This was explained by the possible effect of higher L-lactic acid concentration produced by the effect of L. thermotolerans.
Other beneficial compositional effects reported in L. thermotolerans treated wines include increases in glycerol at concentrations high enough (>5 g/L) that they could be of sensory relevance. As shown by Kapsopoulou et al. [107] for grape must containing 160 g/L sugar (with 7.4 g/L titratable acidity, at pH 3.5), glycerol formation was significantly higher (5.75 g/L) when S. cerevisiae TH941 (5 × 105 cell/mL) was inoculated 3 days after the inoculation of L. thermotolerans SCM952 (5 × 105 cell/mL). This was compared to sole S. cerevisiae fermentations (4.82 g/L) and other sequential inoculations (after 1 and 2 days). The authors attributed this to the increased survival of the L. thermotolerans strain in the co-fermentation treatment in comparison to the other treatments. Likewise, a very high glycerol level (11.22 g/L) was reported in Sangiovese red wines obtained at a commercial scale by the sequential inoculation of S. cerevisiae EC1118 (106 cell/mL) 2 days after that of L. thermotolerans 101 (107 cell/mL) [102]. This value was significantly higher compared to the control (9.02 g/L) and the co-inoculated L. thermotolerans and S. cerevisiae treatments (9.68 g/L). Comitini et al. [13] inoculated non-Saccharomyces yeasts in combination with the S. cerevisiae EC1118 strain as a starter. All non-Saccharomyces strains were inoculated at 107 cell/mL, while the EC1118 starter strain was inoculated at three different concentrations: 107,105, and 103 cell/mL. Of all the inoculations tested, those of L. thermotolerans with S. cerevisiae produced higher levels of glycerol (6.95–7.58 g/L) at all three ratios than when S. cerevisiae was inoculated on its own (6.23–6.56 g/L). Contrary to the C. zemplinina and M. pulcherrima inoculations, no significant differences were found between the three different inoculum ratios (1:1, 100:1, 10,000:1) of L. thermotolerans with S. cerevisiae. This suggests that glycerol production may not correlate with cell concentration and persistence of the L. thermotolerans yeast, at least according to this study. Where the effects of sequential and co-inoculations of L. thermotolerans with S. cerevisiae yeasts on glycerol concentration were evaluated, the glycerol content in sequentially inoculated fermentations (7.55 g/L) was higher than those observed in the co-inoculated treatments (7.18 g/L) [80].
These findings suggest that sequential fermentation with L. thermotolerans and S. cerevisiae remains a viable option for winemakers. Further investigations on the sensory quality of the wines could help identify if these variations in glycerol are discernable by consumers and preserve the wines from the formation of off-flavours.
7. Candida Stellata/Starmerella Bombicola
Candida stellata is a widely studied yeast due to its positive contributions to wine, including its capacity to produce desirable metabolites such as glycerol [72], and its ability to carry out enzymatic activities that have positive effects on wine sensory attributes. An extensive review of C. stellata was carried out by García et al. [108]. Recent studies uncovered the mistake of referring to C. zemplinina instead of C. stellata, which may explain the disputable characterisations of the oenological traits of these species [109]. For instance, recently, a commonly used strain of C. stellata, DBVPG 3827, was reclassified as Starmella bombicola [108].
Traditionally dominating overripe, infected, or botrytised grape berries, C. stellata is reported to tolerate at least 9% v/v ethanol concentration, while at 15 °C, its growth was recorded at 11% v/v ethanol concentration, with decreased tolerance at both 10 °C and 30 °C [22,108,110].
Soden et al. [72] studied the effect of inoculations with C. stellata and S. cerevisiae on Chardonnay juice. The treatments included sole yeast fermentations of C. stellata and S. cerevisiae, as well as co-inoculation and sequential inoculation conducted at a room temperature of 18 °C. C. stellata in sole fermentation was not able to consume all the sugar, reaching an ethanol concentration of only 5.8% v/v. Therefore, S. cerevisiae was used for the sequential inoculation, and added after the fermentation activity of C. stellata had ceased, with an inoculation density of 5 × 106 cells/mL for both yeasts. The resulting wine was dry and had a significantly lower ethanol concentration (11.8% v/v) in comparison to the control with a sole inoculation of S. cerevisiae (12.5% v/v) [72]. The same authors also highlighted in their study the increases in glycerol concentration in the wines fermented with C. stellata in comparison to sole S. cerevisiae fermentations. The glycerol levels varied from 5.2 g/L in the co-inoculation treatment up to 15.7 g/L in the sequential inoculation trial. Similar results for ethanol reduction were found by Ferraro et al. [111] but using immobilised cells of C. stellata and S. cerevisiae on Trebbiano Toscano grape must fermented at 20 °C. Inoculum of S. cerevisiae at 5 × 106 cells/mL was added after 3 days of fermentation. The sequential fermentation of immobilised cells of C. stellata and S. cerevisiae reached an ethanol level of 10.6% v/v compared to the control (11.24% v/v), which was inoculated with S. cerevisiae only [111]. The ethanol reduction was explained by the significant increase (approximately 70%) in glycerol as a consequence of low fermentation rate and reduced production of ethanol with immobilised cells of C. stellata [112].
Immobilised cells of Starmerella bombicola (formerly referred to as Candida stellata) were used in sequential inoculation with S. cerevisiae on Verdicchio grape must. The fermentation was conducted at 25 °C and the inoculation of S. cerevisiae (1 × 106 cell/mL) with 72 h of delay. The effect of the removal of the immobilised S. bombicola cells was investigated [14]. The 1.07% and 1.64% v/v less ethanol achieved with and without the removal of S. bombicola beads, respectively, was notable compared to inoculation with S. cerevisiae. A significant increase was seen in the concentration of by-products such as glycerol and succinic acid. The ethanol reduction was explained by the production of by-products from glycerol–pyruvic fermentation or other metabolic pathways [14].
Both growing in similar environmental conditions, including high sugar-containing musts, and possessing similar taxonomic and oenological profile characteristics, C. zemplinina (synonym Starmerella bacillaris [113]) and C. stellata have produced contrasting results in previous literature. Magyar and Tóth [114] evaluated the oenological characteristics of some yeast strains, including four strains of both C. stellata and C. zemplinina. While both C. stellata and C. zemplinina showed a strong fructophilic character, C. stellata showed higher ethanol and glycerol yield and the same level of volatile acidity compared to C. zemplinina [114].
The role of Candida zemplinina (synonym Starmerella bacillaris) as a tool to produce wines with less ethanol levels but higher glycerol concentrations has been extensively studied and recently reviewed [115]. Candida isolates were obtained from Sicilian musts by Di Maio et al. [116] and sequential inoculations with three different C. zemplinina strains and S. cerevisiae (NDA21) were conducted on Nero d’Avola must. The highest ethanol reduction was 0.3% v/v obtained with the sequential inoculation of C. zemplinina Cz3 strain, compared to a pure inoculation of S. cerevisiae (NDA21), along with higher glycerol content [116]. A similar level of ethanol reduction (0.3% v/v) and higher glycerol production was recorded in the study conducted by Rolle et al. [117] on Barbera must. In this study, two different C. zemplinina strains were used (FC54 and C.z03). The inoculation of C. zemplinina (106 cells/mL) was followed by the inoculation of S. cerevisiae Uvaferm BC (106 cells/mL) after 2 days. Fermentations of natural grape must by sequential inoculations of C. zemplinina with S. cerevisiae EC1118 increased the level of glycerol (5.45–6.30 g/L in the final wine), and remarkably produced less acetaldehyde and total SO2 compared to the other yeasts [118].
8. Torulaspora Delbrueckii
Torulaspora delbrueckii was one of the first commercially available non-Saccharomyces yeasts and is currently available in dry or frozen form from many yeast manufacturers and suppliers [3,10,19]. Some of the oenological traits of T. delbrueckii reported in studies include optimising wine quality parameters i.e., enhancing aroma composition, and positively impacting the foam properties for traditional methods of sparkling wine [119,120,121,122]. Studies have been conducted using this yeast on different wine styles, and its effect on the winemaking practices was recently reviewed [123,124,125].
Significant ethanol reductions due to the utilisation of T. delbrueckii in sequential fermentations have been reported when compared to pure inoculations with S. cerevisiae (Table 1). Nevertheless, some studies reported slight ethanol reductions (lower than 0.2% v/v) [125] or none [88]. Additional, higher ethanol reduction levels were achieved when aeration was integrated during the fermentation process, which stimulates aerobic metabolism [56]. Contreras et al. [56] carried out a sequential inoculation with T. delbrueckii AWRI1152, followed by inoculation of S. cerevisiae AWRI1631 when 50% of sugar was consumed. Four different aeration regimes were applied (air or nitrogen), and fermentations were conducted at 22 °C with agitation (200 rpm) in a chemically defined grape juice medium. With the aeration at 5 mL/min (0.025 VVM) for the first 24 h of the fermentation, an ethanol reduction of 1.5% v/v was achieved, compared to the control of S. cerevisiae under anaerobic conditions. No increase in acetic acid levels occurred; however, the impact on the flavour profile of the wine requires further investigation [56]. To answer this question, Canonico et al. [43] studied three non-Saccharomyces yeast strains (M. pulcherrima, T. delbrueckii, and Zygosaccharomyces bailii) and their ability to reduce ethanol under limited aeration conditions. In this study, sequential inoculation with T. delbrueckii resulted in 0.9% to 1% v/v lower ethanol, depending on the aeration strategy (0.025 VVM and 0.05 VVM) compared to control of S. cerevisiae under anaerobic conditions. Furthermore, wines fermented with T. delbrueckii under aerobic conditions showed a favorable balance between ethanol reduction and volatile profile [43].
Recent studies showed that the nutrient supplementation has a positive correlation on the ethanol yield of T. delbrueckii [126,127]. Additionally, Mecca et al. [127] studied three commercially available T. delbrueckii strains and compared some oenological characteristics. Significant differences were reported in ethanol yields, as well as in the volatile aroma compounds [127]. Concerning the glycerol content of wines made using T. delbrueckii, a range of 4.1–8.9 g/L has been reported in wines (Table 2) [125,128,129]. It has been suggested that T. delbrueckii has a more developed glycerol–pyruvic pathway than other yeasts, although some studies have found no differences in glycerol production [91,124,130].
9. Other Non-Saccharomyces and Saccharomyces Non-cerevisiae Yeasts
In addition to the yeasts mentioned in the sections above, new yeasts are constantly being investigated for their ability to ferment wines and their role in uninoculated fermentations. Contreras et al. [56] studied Zygosaccharomyces bailii in sequential inoculation under different aeration conditions in chemically defined grape juice medium. With the aeration at 5 mL/min (0.025 VVM) throughout the fermentation, an ethanol reduction of 2% (v/v) was achieved, as well as a significantly lower acetic acid compared to the control of S. cerevisiae under anaerobic conditions. Using the same Z. bailii strain in sequential inoculation, in Chardonnay must, Canonico et al. [43] reported a significant ethanol reduction (1% v/v) under aeration (0.05 VVM). In this study, a lower ethanol reduction (0.8% v/v) was achieved with a lower aeration (0.025 VVM); however, the resultant wine had a promising volatile profile including individual esters, higher alcohols, and volatile acidity.
Various studies have reported ethanol reduction with non-Saccharomyces yeasts from different genus, including Pichia and Hanseniaspora (Table 1). Maturano et al. [11] studied two non-Saccharomyces yeasts, H. uvarum and C. membranaefaciens, with the sequential inoculation with S. cerevisiae on Malbec must. Taking a step further, three fermentation factors—inoculum size, time prior to inoculation of Saccharomyces cerevisiae, and fermentation temperature—were optimised using a Box–Behnken experimental design [11]. By applying the optimised factors, the highest ethanol reduction with H. uvarum was achieved with an inoculum size of 5 × 106 cells/mL and a delay of 48 h 37 min before S. cerevisiae inoculation with the fermentation temperature of 25 °C. However, for C. membranaefaciens, the optimised factors were different: an inoculum size of 2.72 × 106 cells/mL, delay of 24 h 15 min before S. cerevisiae inoculation, and fermentation temperature of 25 °C was used. The study showed that the time before the inoculation of S. cerevisiae affected the ethanol production of the non-Saccharomyces yeasts. The high sugar consumption ability of H. uvarum via oxidative metabolism was reported to be the reason for this [11].
The application of sequential inoculations with long delays prior to S. cerevisiae inoculation in winery environments could be challenging. Competitive native or wild S. cerevisiae species present in the winery environment can dominate the fermentation before achieving the expected effect from the inoculated non-Saccharomyces yeasts [14]. A recent study by Canonico et al. [43] reported the volatile profile of reduced ethanol wines. Finding an acceptable balance between ethanol reduction, volatile aroma profile, and sensory characteristics of the wines is crucial. Therefore, studies conducted on a pilot scale that includes sensory analysis of the reduced alcohol wines made from non-Saccharomyces yeasts is lacking. Nitrogen management is an important factor to achieve ethanol reduction, which has been highlighted by authors in previous studies [126,127,131]. The specific nutrient needs of non-Saccharomyces yeasts used for ethanol reduction purposes should be further studied.

{kind=link}
Table 1.
Ethanol reduction in wines produced from mixed fermentations with non-Saccharomyces and Saccharomyces non-cerevisiae with S. cerevisiae yeast.
Table 1.
Ethanol reduction in wines produced from mixed fermentations with non-Saccharomyces and Saccharomyces non-cerevisiae with S. cerevisiae yeast.
| Grape Variety | Wine Style | Ethanol Reduction % (v/v) | Inoculation | Reference |
|---|---|---|---|---|
| Schizosaccharomyces pombe | ||||
| Airén | White still | 0.4 | Sequential inoculation with | [87] |
| S. cerevisiae | ||||
| Airén | White still | 0.65 | Pure inoculation | [87] |
| Schizosaccharomyces japonicus | ||||
| Trebbiano | White still | 2.4 | Sequential inoculation | [132] |
| S. japonicus (immobilised)+ | ||||
| S. cerevisiae | ||||
| Trebbiano | White still | 1.7 | Co-inoculation | [132] |
| S. japonicus (immobilised)+ | ||||
| S. cerevisiae | ||||
| Metschnikowia pulcherrima | ||||
| Malvasia/Viura | White still | 0.8 | Sequential inoculation with | [99] |
| S. cerevisiae (aeration) | ||||
| Chardonnay | White still | 0.9 | Sequential inoculation with | [49] |
| S. cerevisiae | ||||
| Shiraz | Red still | 0.9 | Sequential inoculation with | [93] |
| S. cerevisiae | ||||
| Merlot | Red still | 1 | Co-inoculation with | [50] |
| S. cerevisiae | ||||
| Synthetic grape juice | – | 1.1–1.3 | Sequential inoculation | [14] |
| M. pulcherrima (immobilised)+ | ||||
| S. cerevisiae | ||||
| Verdicchio | White still | 1.2–1.6 | Sequential inoculation | [14] |
| M. pulcherrima (immobilised)+ | ||||
| S. cerevisiae | ||||
| Chardonnay | White still | 0.7–1.6 | Sequential inoculation | [43] |
| (aeration) | ||||
| Shiraz | Red still | 1.6 | Sequential inoculation with | [49] |
| S. cerevisiae | ||||
| Malvasia/Viura | White still | 3.7 | Sequential inoculation with | [57] |
| S. cerevisiae (aeration) | ||||
| Riesling | White still | 3.8 | Sequential inoculation | [47] |
| (aeration) | ||||
| Lachancea thermotolerans | ||||
| Shiraz | Red still | 0.4 | Sequential inoculation with | [71] |
| S. cerevisiae | ||||
| Airén | White still | 0.4 | Sequential inoculation with | [80] |
| S. cerevisiae | ||||
| Sangiovese | Red still | 0.7 | Sequential inoculation with | [102] |
| S. cerevisiae | ||||
| Tempranillo | Red still | 1.2 | Sequential inoculation with | [89] |
| S. cerevisiae | ||||
| Candida stellata/Starmerella bombicola | ||||
| Trebbiano | White still | 0.6 | Sequential inoculation | [111] |
| C. stellata (immobilised) + | ||||
| S. cerevisiae | ||||
| Chardonnay | White still | 0.7 | Sequential inoculation with | [72] |
| S. cerevisiae | ||||
| Verdicchio | White still | 1.6 | Sequential inoculation with S. | [14] |
| bombicola (immobilised) + | ||||
| S. cerevisiae | ||||
| Candida zemplinina/Starmerella bacillaris | ||||
| Nero d’Avola | Rosé still | 0.3 | Sequential inoculation with | [116] |
| S. cerevisiae | ||||
| Barbera | Red still | 0.3 | Sequential inoculation with | [117] |
| S. cerevisiae | ||||
| Barbera | Red still | 0.7 | Sequential inoculation with | [133] |
| S. cerevisiae | ||||
| Riesling | White still | 0.8 | Sequential inoculation with | [47] |
| S. cerevisiae (aeration) | ||||
| Torulaspora delbrueckii | ||||
| Airén | White still | 0.3 | Sequential inoculation with | [134] |
| S. cerevisiae | ||||
| Corvina, Rondinella, Corvinone | Red still | 0.45 | Sequential inoculation with | [135] |
| S. cerevisiae (aeration) | ||||
| Tempranillo | Red still | 0.5 | Sequential inoculation with | [130] |
| S. cerevisiae | ||||
| Chardonnay | White still | 0.9–1.0 | Sequential inoculation with | [43] |
| S. cerevisiae (aeration) | ||||
| Chemically defined grape juice medium | – | 1.5 | Sequential inoculation with | [56] |
| S. cerevisiae (aeration) | ||||
| Malvasia/Viura | White still | 0.5 | Sequential inoculation with | [99] |
| S. cerevisiae (aeration) | ||||
| Zygosaccharomyces bailii | ||||
| Chardonnay | White still | 1.0 | Sequential inoculation with | [43] |
| S. cerevisiae (aeration) | ||||
| Chemically defined grape juice medium | – | 2.0 | Sequential inoculation with | [56] |
| S. cerevisiae (aeration) | ||||
| Pichia kluvyeri | ||||
| Riesling | White still | 0.25 | Sequential inoculation with | [136] |
| S. cerevisiae | ||||
| Riesling | White still | 3.0 | Sequential inoculation with | [47] |
| S. cerevisiae (aeration) | ||||
| Pichia guilliermodii | ||||
| Riesling | White still | 2.0 | Sequential inoculation with | [47] |
| S. cerevisiae (aeration) | ||||
| Hanseniaspora uvarum | ||||
| Pinotage | Red still | 0.8 | Sequential inoculation with | [137] |
| S. cerevisiae | ||||
| Synthetic grape juice | – | 0.8–1.0 | Sequential inoculation | [14] |
| H. osmophila (immobilised)+ | ||||
| S. cerevisiae | ||||
| Verdicchio | White still | 1.0–1.2 | Sequential inoculation | [14] |
| H. osmophila (immobilised)+ | ||||
| S. cerevisiae | ||||
| Sauvignon blanc | White still | 1.3 | Sequential inoculation with | [137] |
| S. cerevisiae | ||||
| Hanseniaspora opuntiae | ||||
| Pinotage | Red still | 0.6 | Sequential inoculation with | [137] |
| S. cerevisiae | ||||
| Sauvignon blanc | White still | 1.3 | Sequential inoculation with | [137] |
| S. cerevisiae | ||||
| Hanseniaspora osmophila | ||||
| Synthetic grape juice | – | 0.8–1.3 | Sequential inoculation | [14] |
| H. osmophila (immobilised)+ | ||||
| S. cerevisiae | ||||
| Verdicchio | White still | 1 | Sequential inoculation | [14] |
| H. osmophila (immobilised)+ | ||||
| S. cerevisiae | ||||
| Saccharomyces uvarum | ||||
| Shiraz | Red still | 0.8 | Sequential inoculation with | [93] |
| S. cerevisiae | ||||
| Merlot | Red still | 1.7 | Pure inoculation | [50] |
Recent studies were conducted with the aim of determining the combinations of two non-Saccharomyces yeast species (S. pombe and L. thermotolerans) [138]. The combination of these yeasts was studied at a laboratory scale using micro-vinifications of Tempranillo grape must [79,89,139,140]. Higher levels of glycerol concentrations were reported in mixed fermentations with L. thermotolerans (CONCERTO™) and S. pombe (inoculated simultaneously at 106 cfu/mL), compared to a control with S. cerevisiae [79,139,140]. Glycerol increase varied between 0.27 and 0.71 g/L, which may be related to the strain of S. pombe used (V2 and 4.5) and control S. cerevisiae (88 and CECT 87). A different strain of S. pombe (938) was studied in combination with L. thermotolerans (CONCERTO™) [89]. Two inoculum ratios of S. pombe/L. thermotolerans (1:1 and 1:3) were used for the laboratory-scale fermentations with Tempranillo must. The authors reported significantly lower glycerol values in mixed fermentation treatments (5.02 g/L and 6.78 g/L) compared to the control fermentation with S. cerevisiae 7VA (7.42 g/L) [89]. Therefore, more studies need to be conducted to understand the effect of the interaction between these two non-Saccharomyces yeasts.
Combinations of non-Saccharomyces and Saccharomyces non-cerevisiae yeasts for wine fermentations have been investigated, including M. pulcherrima and S. uvarum [93,141]. M. pulcherrima and S. uvarum were used in mixed inoculum in the study [93]. Glycerol levels of 12.30 and 12.48 g/L were reported from two different inoculum ratios of M. pulcherrima/S. uvarum (1 × 106 cells/mL to 1 × 105 cells/mL, and 1 × 106 cells/mL to 1 × 104 cells/mL, respectively) in laboratory-scale fermentations of Shiraz must. These values were significantly higher compared to the control wine fermented with S. cerevisiae, where 7.91 g/L glycerol concentration was reported. Although the highest level of glycerol (14.55 g/L) was reported in sequential inoculation with S. uvarum and S. cerevisiae, considering the values of ethanol reduction, a mixed inoculum of M. pulcherrima and S. uvarum could be a promising combination [93]. The same treatments were applied to Chardonnay must in the study by Varela et al. [141]. Similarly, significantly higher glycerol levels were reported in both inoculum ratios of mixed fermentation of M. pulcherrima/S. uvarum (11.90 g/L and 12.63 g/L) compared to control wine fermented with S. cerevisiae (8.20 g/L). Unlike the study of Contreras et al. [93], the reported values in the mixed fermentations were not significantly different from the sequential inoculation of S. uvarum and S. cerevisiae, in which a glycerol level of 11.19 g/L was reported [141]. Considering the ethanol decrease and glycerol levels together, the combined inoculation of M. pulcherrima and S. uvarum shows promising results.
Applying different non-Saccharomyces and Saccharomyces non-cerevisiae yeasts, a wide range of glycerol concentrations has been reported in studies (Table 2) of still and sparkling wines, as well as white and red wines (3.5 g/L to 15.9 g/L). Glycerol levels above 5.2 g/L may directly affect the style of the wine by influencing sensory characteristics including the sweetness, body, and structure of the wines [56]. Therefore, the selection criteria for the use of these yeasts should always consider the sensory characteristics of the final wines. In the case of sparkling wines, glycerol levels have an impact on the viscosity, volatile aroma compounds, and foaming [142]. High levels of glycerol produced by some Saccharomyces non-cerevisiae yeasts, such as Saccharomyces kudriavzevii [143] or Schizosaccharomyces japonicus [132] during base wine fermentation, may not make them suitable candidates for achieving a complete second alcoholic fermentation in sparkling winemaking [8]. Additionally, studies have reported an increase of acetic acid when glycerol is produced by some yeasts [8,73].
Table 2.
Glycerol concentrations in wines from non-Saccharomyces and Saccharomyces non-cerevisiae yeast studies.
Table 2.
Glycerol concentrations in wines from non-Saccharomyces and Saccharomyces non-cerevisiae yeast studies.
| Grape Variety | Wine Style | Glycerol Concentration (g/L) | Method of Detection | Reference |
|---|---|---|---|---|
| Torulaspora delbrueckii | ||||
| Tempranillo | Red still | * 8.6–8.9 | Enzymatically with MIURA One oenological analyser | [129] |
| Chemically defined grape juice medium | N/A | 9.3 | HPLC | [56] |
| Tempranillo | Red still | 6.7 | Y15 enzymatic analyser | [125] |
| Viura/Macabeo | White still | 4.1 | Enzymatically with MIURA One oenological analyser | [128] |
| Hanseniaspora uvarum | ||||
| Negromaro | Red still | 5.5 | HPLC | [144] |
| Chemically defined grape juice medium | N/A | 3.5 | HPLC | [56] |
| Metschnikowia pulcherrima | ||||
| Tempranillo | Red still | * 8.2–8.6 | Enzymatically with MIURA One oenological analyser | [129] |
| Viura/Macabeo | White still | 4.8 | Enzymatically with MIURA One oenological analyser | [128] |
| Chardonnay | White still | 5.5–7.8 | HPLC | [43] |
| Schizosaccharomyces pombe‡ | ||||
| Airén | Sparkling wine | 4.7 | Y15 enzymatic analyser | [145] |
| Tempranillo | Red sparkling wine | 5.0 | Y15 enzymatic analyser | [145] |
| Schizosaccharomyces japonicus | ||||
| Trebbiano | White still | 15.9 | HPLC | [132] |
| Saccharomycodes ludwigii‡ | ||||
| Airén | Sparkling wine | 5.0 | Y15 enzymatic analyser | [145] |
| Tempranillo | Red sparkling wine | 5.1 | Y15 enzymatic analyser | [145] |
| Saccharomyces uvarum‡ | ||||
| Synthetic grape must | N/A | 5.2 | HPLC | [12] |
| Cabernet franc | Red wine | 10–12 | Enzymatically using Megazyme International assay kit | [146] |
| Lachancea thermotolerans | ||||
| Tempranillo | Red still | * 8.2–8.3 | Enzymatically with MIURA One oenological analyser | [129] |
| Viura/Macabeo | White still | 4.7 | Enzymatically with MIURA One oenological analyser | [128] |
| Starmerella bacillaris | ||||
| Synthetic grape must | N/A | * 7.7–8.2 | HPLC | [128] |
| Williopsis pratensis | ||||
| Tempranillo | Red still | 8.0 | Enzymatically with MIURA One oenological analyser | [129] |
| Zygosaccharomyces bailii | ||||
| Tempranillo | Red still | 7.8 | Enzymatically with MIURA One oenological analyser | [129] |
| Viura/Macabeo | White still | 5.6 | Enzymatically with MIURA One oenological analyser | [128] |
| Candida vini | ||||
| Tempranillo | Red still | 7.9 | Enzymatically with MIURA One oenological analyser | [129] |
* Two or more products of the same yeast strain used in the study. ‡ Saccharomyces non-cerevisiae yeast.
10. Conclusions
This review focused on studies concerning non-Saccharomyces and Saccharomyces non-cerevisiae yeasts, and their effect in solo, sequential, and co-fermentation with S. cerevisiae or other non-Saccharomyces yeasts, on the ethanol and glycerol content of wines. The application of large-scale sequential inoculations of yeast in a commercial scale winery could be challenging during a busy harvest period, especially if the length of time varies between yeast additions. There is a distinct lack of studies concerning the long-term effect of these yeasts on wine from aging on lees and in bottles, as well as the effect of high glycerol levels in sparkling wines on sensory characteristics. Further research should include the inoculum rate of the yeasts, aging ability of the wines on lees, aeration techniques including flow rate, and time before the sequential yeast is added.
Author Contributions
B.K. conceived the idea, supervised the writing, contributed to the glycerol section and produced Table 2. N.N.I. contributed to the introduction and yeast sections, and produced Table 1. R.L. wrote the alcohol section and contributed to other section within the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors wish to thank Hannah Charnock, Brock University, St. Catharines, Ontario, Canada for designing the graphic abstract that accompanies this article using Biorender Software, (Toronto, Canada).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Scanes, K.; Hohrnann, S.; Prior, B. Glycerol production by the yeast Saccharomyces cerevisiae and its relevance to wine: A review. S. Afr. J. Enol. Vitic. 1998, 19, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francesca, N.; Gaglio, R.; Alfonzo, A.; Settanni, L.; Corona, O.; Mazzei, P.; Romano, R.; Piccolo, A.; Moschetti, G. The wine: Typicality or mere diversity? The effect of spontaneous fermentations and biotic factors on the characteristics of wine. Agric. Agric. Sci. Procedia 2016, 8, 769–773. [Google Scholar] [CrossRef] [Green Version]
- Vigentini, I.; Barrera Cardenas, S.; Valdetara, F.; Faccincani, M.; Panont, C.A.; Picozzi, C.; Foschino, R. Use of native yeast strains for in-bottle fermentation to face the uniformity in sparkling wine production. Front. Microbiol. 2017, 8, 1225. [Google Scholar] [CrossRef]
- Ciani, M.; Canonico, L.; Oro, L.; Comitini, F. Footprint of Nonconventional Yeasts and Their Contribution in Alcoholic Fermentations. In Biotechnological Progress and Beverage Consumption; Academic Press: Cambridge, MA, USA, 2020; pp. 435–465. [Google Scholar]
- Ciani, M.; Capece, A.; Comitini, F.; Canonico, L.; Siesto, G.; Romano, P. Yeast interactions in inoculated wine fermentation. Front. Microbiol. 2016, 7, 555. [Google Scholar] [CrossRef] [Green Version]
- García, M.; Esteve-Zarzoso, B.; Cabellos, J.M.; Arroyo, T. Sequential non-Saccharomyces and Saccharomyces cerevisiae fermentations to reduce the alcohol content in wine. Fermentation 2020, 6, 60. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [Green Version]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial resources and enological significance: Opportunities and benefits. Front. Microbiol. 2017, 8, 995. [Google Scholar] [CrossRef] [Green Version]
- Maturano, Y.P.; Mestre, M.V.; Kuchen, B.; Toro, M.E.; Mercado, L.A.; Vazquez, F.; Combina, M. Optimization of fermentation-relevant factors: A strategy to reduce ethanol in red wine by sequential culture of native yeasts. Int. J. Food Microbiol. 2019, 289, 40–48. [Google Scholar] [CrossRef]
- Alonso-del-Real, J.; Lairón-Peris, M.; Barrio, E.; Querol, A. Effect of temperature on the prevalence of Saccharomyces non-cerevisiae species against a S. cerevisiae wine strain in wine fermentation: Competition, physiological fitness, and influence in final wine composition. Front. Microbiol. 2017, 8, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Canonico, L.; Comitini, F.; Oro, L.; Ciani, M. Sequential fermentation with selected immobilized non-Saccharomyces yeast for reduction of ethanol content in wine. Front. Microbiol. 2016, 7, 278. [Google Scholar] [CrossRef] [Green Version]
- Pretorius, I.S. Conducting wine symphonics with the aid of yeast genomics. Beverages 2016, 2, 36. [Google Scholar] [CrossRef] [Green Version]
- Benito, Á.; Calderón, F.; Benito, S. The influence of non-Saccharomyces species on wine fermentation quality parameters. Fermentation 2019, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Torrado, R.; Barrio, E.; Querol, A. Alternative yeasts for winemaking: Saccharomyces non-cerevisiae and its hybrids. Crit. Rev. Food Sci. Nutr. 2018, 58, 1780–1790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilela, A. Use of nonconventional yeasts for modulating wine acidity. Fermentation 2019, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Escott, C.; Bañuelos, M.A.; Loira, I.; del Fresno, J.M.; González, C.; Suárez-Lepe, J.A. Contribution of non-Saccharomyces yeasts to wine freshness. A review. Biomolecules 2020, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Ivit, N.N.; Kemp, B. The impact of non-Saccharomyces yeast on traditional method sparkling wine. Fermentation 2018, 4, 73. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Escott, C.; Loira, I.; Del Fresno, J.M.; González, C.; Suárez-Lepe, J.A. Influence of Saccharomyces and non-Saccharomyces yeasts in the formation of pyranoanthocyanins and polymeric pigments during red wine making. Molecules 2019, 24, 4490. [Google Scholar] [CrossRef] [Green Version]
- Jackson, R. Fermentation. In Wine Science: Principles and Applications (Food Science and Technology); Academic Press: Amsterdam, The Netherlands, 2014; Chapter 7; p. 493. [Google Scholar]
- Ickes, C.M.; Cadwallader, K.R. Effects of ethanol on flavor perception in alcoholic beverages. Chemosens. Percept. 2017, 10, 119–134. [Google Scholar] [CrossRef]
- King, E.S.; Dunn, R.L.; Heymann, H. The influence of alcohol on the sensory perception of red wines. Food Qual. Pref. 2013, 28, 235–243. [Google Scholar] [CrossRef]
- Nurgel, C.; Pickering, G. Modeling of sweet, bitter and irritant sensations and their interactions elicited by model ice wines. J. Sens. Stud. 2006, 21, 505–519. [Google Scholar] [CrossRef]
- Fischer, U.; Noble, C.A. The effect of ethanol, catechin concentration, and ph on sourness and bitterness of wine. Am. J. Enol. Vitic. 1994, 45, 6–10. [Google Scholar]
- Gawel, R.; Sluyter, S.V.; Waters, E.J. The effects of ethanol and glycerol on the body and other sensory characteristics of Riesling wines. Aust. J. Grape Wine Res. 2007, 13, 38–45. [Google Scholar] [CrossRef]
- Saha, B.; Longo, R.; Torley, P.; Saliba, A.; Schmidtke, L. SPME method optimized by Box-Behnken design for impact odorants in reduced alcohol wines. Foods 2018, 7, 127. [Google Scholar] [CrossRef] [Green Version]
- OIV. Oiv International Standard for the Labelling of Wines 2015; OIV: Paris, France, 2015. [Google Scholar]
- Godden, P.; Wilkes, E.; Johnson, D. Trends in the composition of Australian wine 1984–2014. Aust. J. Grape Wine Res. 2015, 21, 741–753. [Google Scholar] [CrossRef]
- Alston, J.M.; Fuller, K.B.; Lapsley, J.T.; Soleas, G.; Tumber, K.P. Splendide mendax: False label claims about high and rising alcohol content of wine. J. Wine Econ. 2015, 10, 275–313. [Google Scholar] [CrossRef]
- Van Leeuwen, C.; Darriet, P. The impact of climate change on viticulture and wine quality. J. Wine Econ. 2016, 11, 150–167. [Google Scholar] [CrossRef] [Green Version]
- Williamson, P.O.; Robichaud, J.; Francis, I.L. Comparison of Chinese and Australian consumers’ liking responses for red wines. Aust. J. Grape Wine Res. 2012, 18, 256–267. [Google Scholar] [CrossRef]
- Bindon, K.; Varela, C.; Kennedy, J.; Holt, H.; Herderich, M. Relationships between harvest time and wine composition in Vitis vinifera L. cv. Cabernet sauvignon 1. Grape and wine chemistry. Food Chem 2013, 138, 1696–1705. [Google Scholar] [CrossRef] [PubMed]
- Jordão, A.; Vilela, A.; Cosme, F. From sugar of grape to alcohol of wine: Sensorial impact of alcohol in wine. Beverages 2015, 1, 292–310. [Google Scholar] [CrossRef] [Green Version]
- Bucher, T.; Deroover, K.; Stockley, C. Low-alcohol wine: A narrative review on consumer perception and behaviour. Beverages 2018, 4, 82. [Google Scholar] [CrossRef] [Green Version]
- Saliba, A.; Ovington, L.; Moran, C. Consumer demand for low-alcohol wine in an Australian sample. Int. J. Wine Res. 2013, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.; Sinha, K.; Vandenberg, B. Pricing as a means of controlling alcohol consumption. Br. Med Bull. 2017, 123, 149–158. [Google Scholar] [CrossRef]
- Annunziata, A.; Pomarici, E.; Vecchio, R.; Mariani, A. Do consumers want more nutritional and health information on wine labels? Insights from the eu and USA. Nutrients 2016, 8, 416. [Google Scholar] [CrossRef] [Green Version]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. Conditions of Yeast Development. In Handbook of Enology, Volume 1: The Microbiology of Wine and Vinifications; John Wiley & Sons: Hoboken, NJ, USA, 2006; pp. 79–114. [Google Scholar]
- Zapparoli, G.; Tosi, E.; Azzolini, M.; Vagnoli, P.; Krieger, S. Bacterial inoculation strategies for the achievement of malolactic fermentation in high-alcohol wines. S. Afr. J. Enol. Vitic. 2009, 30, 49–55. [Google Scholar] [CrossRef]
- Goold, H.D.; Kroukamp, H.; Williams, T.C.; Paulsen, I.T.; Varela, C.; Pretorius, I.S. Yeast’s balancing act between ethanol and glycerol production in low-alcohol wines. Microb. Biotechnol. 2017, 10, 264–278. [Google Scholar] [CrossRef]
- Canonico, L.; Solomon, M.; Comitini, F.; Ciani, M.; Varela, C. Volatile profile of reduced alcohol wines fermented with selected non-Saccharomyces yeasts under different aeration conditions. Food Microbiol. 2019, 84, 103247. [Google Scholar] [CrossRef]
- Longo, R.; Blackman, J.W.; Torley, P.J.; Rogiers, S.Y.; Schmidtke, L.M. Changes in volatile composition and sensory attributes of wines during alcohol content reduction. J. Sci. Food Agric. 2017, 97, 8–16. [Google Scholar] [CrossRef]
- Longo, R.; Blackman, J.W.; Antalick, G.; Torley, P.J.; Rogiers, S.Y.; Schmidtke, L.M. A comparative study of partial dealcoholisation versus early harvest: Effects on wine volatile and sensory profiles. Food Chem. 2018, 261, 21–29. [Google Scholar] [CrossRef]
- Varela, C.; Dry, P.; Kutyna, D.; Francis, I.; Henschke, P.; Curtin, C.; Chambers, P. Strategies for reducing alcohol concentration in wine. Aust. J. Grape Wine Res. 2015, 21, 670–679. [Google Scholar] [CrossRef]
- Röcker, J.; Strub, S.; Ebert, K.; Grossmann, M. Usage of different aerobic non-Saccharomyces yeasts and experimental conditions as a tool for reducing the potential ethanol content in wines. Eur. Food Res. Technol. 2016, 242, 2051–2070. [Google Scholar] [CrossRef]
- Ciani, M.; Morales, P.; Comitini, F.; Tronchoni, J.; Canonico, L.; Curiel, J.A.; Oro, L.; Rodrigues, A.J.; Gonzalez, R. Non-conventional yeast species for lowering ethanol content of wines. Front. Microbiol. 2016, 7, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras, A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin, C.; Varela, C. Evaluation of non-Saccharomyces yeasts for the reduction of alcohol content in wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, C.; Barker, A.; Tran, T.; Borneman, A.; Curtin, C. Sensory profile and volatile aroma composition of reduced alcohol Merlot wines fermented with Metschnikowia pulcherrima and Saccharomyces uvarum. Int. J. Food Microbiol. 2017, 252, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Comitini, F. Use of non-Saccharomyces yeasts in red winemaking. In Red Wine Technology; Antonio, M., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 51–68. [Google Scholar]
- Ciani, M.; Comitini, F. Influence of temperature and oxygen concentration on the fermentation behaviour of Candida stellata in mixed fermentation with Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2006, 22, 619–623. [Google Scholar] [CrossRef]
- Alexander, M.; Jeffries, T. Respiratory efficiency and metabolite partitioning as regulatory phenomena in yeasts. Enzym. Microb. Technol. 1990, 12, 2–19. [Google Scholar] [CrossRef]
- De Deken, R. The crabtree effect: A regulatory system in yeast. Microbiology 1966, 44, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canonico, L.; Galli, E.; Ciani, E.; Comitini, F.; Ciani, M. Exploitation of three non-conventional yeast species in the brewing process. Microorganisms 2019, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.A.; Curtin, C.; Varela, C. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.; Rojas, V.; Quirós, M.; Gonzalez, R. The impact of oxygen on the final alcohol content of wine fermented by a mixed starter culture. Appl. Microbiol. Biotechnol. 2015, 99, 3993–4003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, R.; Quirós, M.; Morales, P. Yeast respiration of sugars by non-Saccharomyces yeast species: A promising and barely explored approach to lowering alcohol content of wines. Trends Food Sci. Technol. 2013, 29, 55–61. [Google Scholar] [CrossRef]
- Quirós, M.; Rojas, V.; Gonzalez, R.; Morales, P. Selection of non-Saccharomyces yeast strains for reducing alcohol levels in wine by sugar respiration. Int. J. Food Microbiol. 2014, 181, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Mestre Furlani, M.V.; Maturano, Y.P.; Combina, M.; Mercado, L.A.; Toro, M.E.; Vazquez, F. Selection of non-Saccharomyces yeasts to be used in grape musts with high alcoholic potential: A strategy to obtain wines with reduced ethanol content. FEMS Yeast Res. 2017, 17. [Google Scholar] [CrossRef] [Green Version]
- Ballester-Tomás, L.; Prieto, J.A.; Gil, J.V.; Baeza, M.; Randez-Gil, F. The Antarctic yeast Candida sake: Understanding cold metabolism impact on wine. Int. J. Food Microbiol. 2017, 245, 59–65. [Google Scholar] [CrossRef]
- Mbuyane, L.L.; de Kock, M.; Bauer, F.F.; Divol, B. Torulaspora delbrueckii produces high levels of c5 and c6 polyols during wine fermentations. FEMS Yeast Res. 2018, 18, foy084. [Google Scholar] [CrossRef]
- Puškaš, V.S.; Miljić, U.D.; Djuran, J.J.; Vučurović, V.M. The aptitude of commercial yeast strains for lowering the ethanol content of wine. Food Sci. Nutr. 2020, 8, 1489–1498. [Google Scholar] [CrossRef]
- Nieuwoudt, H.; Prior, B.; Pretorius, L.; Bauer, F. Glycerol in South African table wines: An assessment of its relationship to wine quality. S. Afr. J. Enol. Vitic. 2002, 23, 22–30. [Google Scholar] [CrossRef]
- Nissen, T.L.; Hamann, C.W.; Kielland-Brandt, M.C.; Nielsen, J.; Villadsen, J. Anaerobic and aerobic batch cultivations of Saccharomyces cerevisiae mutants impaired in glycerol synthesis. Yeast 2000, 16, 463–474. [Google Scholar] [CrossRef]
- Roustan, J.; Sablayrolles, J. Impact of the addition of electron acceptors on the by-products of alcoholic fermentation. Enzym. Microb. Technol. 2002, 31, 142–152. [Google Scholar] [CrossRef]
- Rantsiou, K.; Dolci, P.; Giacosa, S.; Torchio, F.; Tofalo, R.; Torriani, S.; Suzzi, G.; Rolle, L.; Cocolin, L. Candida zemplinina can reduce acetic acid produced by Saccharomyces cerevisiae in sweet wine fermentations. Appl. Environ. Microbiol. 2012, 78, 1987–1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domizio, P.; Liu, Y.; Bisson, L.; Barile, D. Cell wall polysaccharides released during the alcoholic fermentation by Schizosaccharomyces pombe and S. Japonicus: Quantification and characterization. Food Microbiol. 2017, 61, 136–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Gianvito, P.; Perpetuini, G.; Tittarelli, F.; Schirone, M.; Arfelli, G.; Piva, A.; Patrignani, F.; Lanciotti, R.; Olivastri, L.; Suzzi, G. Impact of Saccharomyces cerevisiae strains on traditional sparkling wines production. Food Res. Int. 2018, 109, 552–560. [Google Scholar] [CrossRef]
- Longo, R.; Blackman, J.W.; Antalick, G.; Torley, P.J.; Rogiers, S.Y.; Schmidtke, L.M. Volatile and sensory profiling of Shiraz wine in response to alcohol management: Comparison of harvest timing versus technological approaches. Food Res. Int. 2018, 109, 561–571. [Google Scholar] [CrossRef]
- Hranilovic, A.; Li, S.; Boss, P.K.; Bindon, K.; Ristic, R.; Grbin, P.R.; Van der Westhuizen, T.; Jiranek, V. Chemical and sensory profiling of Shiraz wines co-fermented with commercial non-Saccharomyces inocula. Aust. J. Grape Wine Res. 2018, 24, 166–180. [Google Scholar] [CrossRef]
- Soden, A.; Francis, I.; Oakey, H.; Henschke, P. Effects of co-fermentation with Candida stellata and Saccharomyces cerevisiae on the aroma and composition of chardonnay wine. Aust. J. Grape Wine Res. 2000, 6, 21–30. [Google Scholar] [CrossRef]
- Eglinton, J.M.; Heinrich, A.J.; Pollnitz, A.P.; Langridge, P.; Henschke, P.A.; de Barros Lopes, M. Decreasing acetic acid accumulation by a glycerol overproducing strain of Saccharomyces cerevisiae by deleting the ald6 aldehyde dehydrogenase gene. Yeast 2002, 19, 295–301. [Google Scholar] [CrossRef]
- Li, H.; Du, G.; Li, H.-L.; Wang, H.-L.; Yan, G.-L.; Zhan, J.-C.; Huang, W.-D. Physiological response of different wine yeasts to hyperosmotic stress. Am. J. Enol. Vitic. 2010, 61, 529–535. [Google Scholar] [CrossRef]
- Pérez-Nevado, F.; Albergaria, H.; Hogg, T.; Girio, F. Cellular death of two non-Saccharomyces wine-related yeasts during mixed fermentations with Saccharomyces cerevisiae. Int. J. Food Microbiol. 2006, 108, 336–345. [Google Scholar]
- Shekhawat, K.; Bauer, F.F.; Setati, M.E. Impact of oxygenation on the performance of three non-Saccharomyces yeasts in co-fermentation with Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2017, 101, 2479–2491. [Google Scholar] [CrossRef]
- Prior, B.; Toh, T.; Jolly, N.; Baccari, C.; Mortimer, R. Impact of yeast breeding for elevated glycerol production on fermentative activity and metabolite formation in Chardonnay wine. S. Afr. J. Enol. Vitic. 2000, 21, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.; Ramón-Portugal, F.; Andrade, P.; Abreu, S.; de Fatima Texeira, M.; Strehaiano, P. Malic acid consumption by dry immobilized cells of Schizosaccharomyces pombe. Am. J. Enol. Vitic. 2003, 54, 50–55. [Google Scholar]
- Benito, Á.; Calderón, F.; Benito, S. The combined use of Schizosaccharomyces pombe and Lachancea thermotolerans—Effect on the anthocyanin wine composition. Molecules 2017, 22, 739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benito, Á.; Calderón, F.; Palomero, F.; Benito, S. Quality and composition of Airén wines fermented by sequential inoculation of Lachancea thermotolerans and Saccharomyces cerevisiae. Food Technol. Biotechnol. 2016, 54, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, F.; Morassut, M.; Moruno, E.G.; Di Stefano, R. Influence of yeast strain on Ochratoxin A content during fermentation of white and red must. Food Microbiol. 2006, 23, 411–417. [Google Scholar] [CrossRef]
- Escott, C.; Del Fresno, J.M.; Loira, I.; Morata, A.; Suárez-Lepe, J.A. Zygosaccharomyces rouxii: Control strategies and applications in food and winemaking. Fermentation 2018, 4, 69. [Google Scholar] [CrossRef] [Green Version]
- Loira, I.; Morata, A.; Palomero, F.; González, C.; Suárez-Lepe, J.A. Schizosaccharomyces pombe: A promising biotechnology for modulating wine composition. Fermentation 2018, 4, 70. [Google Scholar] [CrossRef] [Green Version]
- Benito, S. The impacts of Schizosaccharomyces on winemaking. Appl. Microbiol. Biotechnol. 2019, 103, 4291–4312. [Google Scholar] [CrossRef]
- Suárez-Lepe, J.; Palomero, F.; Benito, S.; Calderón, F.; Morata, A. Oenological versatility of Schizosaccharomyces spp. Eur. Food Res. Technol. 2012, 235, 375–383. [Google Scholar] [CrossRef]
- Palomero, F.; Morata, A.; Benito, S.; Calderón, F.; Suárez-Lepe, J. New genera of yeasts for over-lees aging of red wine. Food Chem. 2009, 112, 432–441. [Google Scholar] [CrossRef]
- Benito, S.; Palomero, F.; Morata, A.; Calderon, F.; Palmero, D.; Suárez-Lepe, J. Physiological features of Schizosaccharomyces pombe of interest in making of white wines. Eur. Food Res. Technol. 2013, 236, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Loira, I.; Morata, A.; Comuzzo, P.; Callejo, M.J.; González, C.; Calderón, F.; Suárez-Lepe, J.A. Use of Schizosaccharomyces pombe and Torulaspora delbrueckii strains in mixed and sequential fermentations to improve red wine sensory quality. Food Res. Int. 2015, 76, 325–333. [Google Scholar] [CrossRef]
- Del Fresno, J.M.; Morata, A.; Loira, I.; Bañuelos, M.A.; Escott, C.; Benito, S.; Chamorro, C.G.; Suárez-Lepe, J.A. Use of non-Saccharomyces in single-culture, mixed and sequential fermentation to improve red wine quality. Eur. Food Res. Technol. 2017, 243, 2175–2185. [Google Scholar] [CrossRef]
- Miljić, U.; Puškaš, V.; Vučurović, V.; Muzalevski, A. Fermentation characteristics and aromatic profile of plum wines produced with indigenous microbiota and pure cultures of selected yeast. J. Food Sci. 2017, 82, 1443–1450. [Google Scholar] [CrossRef]
- Minnaar, P.; Jolly, N.; Paulsen, V.; Du Plessis, H.; Van Der Rijst, M. Schizosaccharomyces pombe and Saccharomyces cerevisiae yeasts in sequential fermentations: Effect on phenolic acids of fermented kei-apple (Dovyalis caffra L.) juice. Int. J. Food Microbiol. 2017, 257, 232–237. [Google Scholar] [CrossRef]
- Sadoudi, M.; Tourdot-Maréchal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacón, J.-J.; Ballester, J.; Vichi, S.; Guérin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast–yeast interactions revealed by aromatic profile analysis of sauvignon blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef]
- Contreras, A.; Curtin, C.; Varela, C. Yeast population dynamics reveal a potential collaboration between Metschnikowia pulcherrima and Saccharomyces uvarum for the production of reduced alcohol wines during Shiraz fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 1885–1895. [Google Scholar] [CrossRef]
- Duarte, F.L.; Egipto, R.; Baleiras-Couto, M.M. Mixed fermentation with Metschnikowia pulcherrima using different grape varieties. Fermentation 2019, 5, 59. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Loira, I.; Escott, C.; del Fresno, J.M.; Bañuelos, M.A.; Suárez-Lepe, J.A. Applications of Metschnikowia pulcherrima in wine biotechnology. Fermentation 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Combina, M.; Elía, A.; Mercado, L.; Catania, C.; Ganga, A.; Martinez, C. Dynamics of indigenous yeast populations during spontaneous fermentation of wines from Mendoza, Argentina. Int. J. Food Microbiol. 2005, 99, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, C.; Lage, P.; Esteves, M.; Chambel, L.; Mendes-Faia, A.; Mendes-Ferreira, A. Molecular and phenotypic characterization of Metschnikowia pulcherrima strains from douro wine region. Fermentation 2018, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Canonico, L.; Comitini, F.; Ciani, M. Metschnikowia pulcherrima Selected Strain for Ethanol Reduction in Wine: Influence of Cell Immobilization and Aeration Condition. Foods 2019, 8, 378. [Google Scholar] [CrossRef] [Green Version]
- Tronchoni, J.; Curiel, J.A.; Sáenz-Navajas, M.P.; Morales, P.; De-La-Fuente-Blanco, A.; Fernández-Zurbano, P.; Ferreira, V.; Gonzalez, R. Aroma profiling of an aerated fermentation of natural grape must with selected yeast strains at pilot scale. Food Microbiol. 2018, 70, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Ribéreau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. Alcohols and Other Volatile Compounds. In Handbook of Enology, Volume 2: The Chemistry of Wine Stabilization and Treatments, 2nd ed.; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2006; pp. 51–64. [Google Scholar]
- Benito, S. The impacts of Lachancea thermotolerans yeast strains on winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 6775–6790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Balikci, E.K.; Tanguler, H.; Jolly, N.P.; Erten, H. Influence of Lachancea thermotolerans on cv. Emir wine fermentation. Yeast 2016, 33, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Loira, I.; Tesfaye, W.; Bañuelos, M.A.; González, C.; Suárez Lepe, J.A. Lachancea thermotolerans applications in wine technology. Fermentation 2018, 4, 53. [Google Scholar] [CrossRef] [Green Version]
- Porter, T.J.; Divol, B.; Setati, M.E. Lachancea yeast species: Origin, biochemical characteristics and oenological significance. Food Res. Int. 2019, 119, 378–389. [Google Scholar] [CrossRef]
- Vilela, A. Lachancea thermotolerans, the Non-Saccharomyces Yeast that Reduces the Volatile Acidity of Wines. Fermentation 2018, 4, 56. [Google Scholar] [CrossRef] [Green Version]
- Kapsopoulou, K.; Mourtzini, A.; Anthoulas, M.; Nerantzis, E. Biological acidification during grape must fermentation using mixed cultures of Kluyveromyces thermotolerans and Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2007, 23, 735–739. [Google Scholar] [CrossRef]
- García, M.; Esteve-Zarzoso, B.; Cabellos, J.M.; Arroyo, T. Advances in the study of Candida stellata. Fermentation 2018, 4, 74. [Google Scholar] [CrossRef] [Green Version]
- Csoma, H.; Sipiczki, M. Taxonomic reclassification of Candida stellata strains reveals frequent occurrence of Candida zemplinina in wine fermentation. FEMS Yeast Res. 2008, 8, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.; Fleet, G. The effects of temperature and pH on the ethanol tolerance of the wine yeasts, Saccharomyces cerevisiae, Candida stellata and Kloeckera apiculata. J. Appl. Bacteriol. 1988, 65, 405–409. [Google Scholar] [CrossRef]
- Ferraro, L.; Fatichenti, F.; Ciani, M. Pilot scale vinification process using immobilized Candida stellata cells and Saccharomyces cerevisiae. Process Biochem. 2000, 35, 1125–1129. [Google Scholar] [CrossRef]
- Ciani, M.; Ferraro, L. Enhanced glycerol content in wines made with immobilized Candida stellata cells. Appl. Environ. Microbiol. 1996, 62, 128–132. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Esteve-Zarzoso, B.; Cocolin, L.; Mas, A.; Rantsiou, K. Viable and culturable populations of Saccharomyces cerevisiae, Hanseniaspora uvarum and Starmerella bacillaris (synonym Candida zemplinina) during Barbera must fermentation. Food Res. Int. 2015, 78, 195–200. [Google Scholar] [CrossRef]
- Magyar, I.; Tóth, T. Comparative evaluation of some oenological properties in wine strains of Candida stellata, Candida zemplinina, Saccharomyces uvarum and Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 94–100. [Google Scholar] [CrossRef]
- Englezos, V.; Giacosa, S.; Rantsiou, K.; Rolle, L.; Cocolin, L. Starmerella bacillaris in winemaking: Opportunities and risks. Curr. Opin. Food Sci. 2017, 17, 30–35. [Google Scholar] [CrossRef]
- Di Maio, S.; Genna, G.; Gandolfo, V.; Amore, G.; Ciaccio, M.; Oliva, D. Presence of Candida zemplinina in Sicilian musts and selection of a strain for wine mixed fermentations. S. Afr. J. Enol. Vitic. 2012, 33, 80–87. [Google Scholar] [CrossRef]
- Rolle, L.; Englezos, V.; Torchio, F.; Cravero, F.; Río Segade, S.; Rantsiou, K.; Giacosa, S.; Gambuti, A.; Gerbi, V.; Cocolin, L. Alcohol reduction in red wines by technological and microbiological approaches: A comparative study. Aust. J. Grape Wine Res. 2017, 24, 62–74. [Google Scholar] [CrossRef]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of non-Saccharomyces yeasts to wine volatile and sensory diversity: A study on Lachancea thermotolerans, Metschnikowia spp. and Starmerella bacillaris strains isolated in Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef] [PubMed]
- Loira, I.; Vejarano, R.; Bañuelos, M.; Morata, A.; Tesfaye, W.; Uthurry, C.; Villa, A.; Cintora, I.; Suárez-Lepe, J. Influence of sequential fermentation with Torulaspora delbrueckii and Saccharomyces cerevisiae on wine quality. LWT-Food Sci. Technol. 2014, 59, 915–922. [Google Scholar] [CrossRef] [Green Version]
- Arslan, E.; Çelik, Z.D.; Cabaroğlu, T. Effects of pure and mixed autochthonous Torulaspora delbrueckii and Saccharomyces cerevisiae on fermentation and volatile compounds of narince wines. Foods 2018, 7, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Royo, E.; Pascual, O.; Kontoudakis, N.; Esteruelas, M.; Esteve-Zarzoso, B.; Mas, A.; Canals, J.M.; Zamora, F. Oenological consequences of sequential inoculation with non-Saccharomyces yeasts (Torulaspora delbrueckii or Metschnikowia pulcherrima) and Saccharomyces cerevisiae in base wine for sparkling wine production. Eur. Food Res. Technol. 2015, 240, 999–1012. [Google Scholar] [CrossRef]
- Canonico, L.; Comitini, F.; Ciani, M. Torulaspora delbrueckii for secondary fermentation in sparkling wine production. Food Microbiol. 2018, 74, 100–106. [Google Scholar] [CrossRef]
- Ramírez, M.; Velázquez, R. The yeast Torulaspora delbrueckii: An interesting but difficult-to-use tool for winemaking. Fermentation 2018, 4, 94. [Google Scholar] [CrossRef] [Green Version]
- Benito, S. The impact of Torulaspora delbrueckii yeast in winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 3081–3094. [Google Scholar] [CrossRef]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderon, F.; Benito, S. Dynamic analysis of physiological properties of Torulaspora delbrueckii in wine fermentations and its incidence on wine quality. Appl. Microbiol. Biotechnol. 2015, 99, 1911–1922. [Google Scholar] [CrossRef]
- Taillandier, P.; Lai, Q.P.; Julien-Ortiz, A.; Brandam, C. Interactions between Torulaspora delbrueckii and Saccharomyces cerevisiae in wine fermentation: Influence of inoculation and nitrogen content. World J. Microbiol. Biotechnol. 2014, 30, 1959–1967. [Google Scholar] [CrossRef] [Green Version]
- Mecca, D.; Benito, S.; Beisert, B.; Brezina, S.; Fritsch, S.; Semmler, H.; Rauhut, D. Influence of nutrient supplementation on Torulaspora delbrueckii wine fermentation aroma. Fermentation 2020, 6, 35. [Google Scholar] [CrossRef] [Green Version]
- Escribano, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A. Wine aromatic compound production and fermentative behaviour within different non-Saccharomyces species and clones. J. Appl. Microbiol. 2018, 124, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- Escribano-Viana, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aroma evolution throughout alcoholic fermentation sequentially inoculated with non-Saccharomyces/Saccharomyces yeasts. Food Res. Int. 2018, 112, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Ruiz, J.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Rauhut, D.; Benito, S.; Santos, A. Influence of Torulaspora delbrueckii in varietal thiol (3-SH and 4-MSP) release in wine sequential fermentations. Int. J. Food Microbiol. 2017, 257, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.J.; Raimbourg, T.; Gonzalez, R.; Morales, P. Environmental factors influencing the efficacy of different yeast strains for alcohol level reduction in wine by respiration. LWT-Food Sci. Technol. 2016, 65, 1038–1043. [Google Scholar] [CrossRef]
- Domizio, P.; Lencioni, L.; Calamai, L.; Portaro, L.; Bisson, L.F. Evaluation of the yeast Schizosaccharomyces japonicus for use in wine production. Am. J. Enol. Vitic. 2018, 69, 266–277. [Google Scholar] [CrossRef] [Green Version]
- Englezos, V.; Rantsiou, K.; Cravero, F.; Torchio, F.; Ortiz-Julien, A.; Gerbi, V.; Rolle, L.; Cocolin, L. Starmerella bacillaris and Saccharomyces cerevisiae mixed fermentations to reduce ethanol content in wine. Appl. Microbiol. Biotechnol. 2016, 12, 5515–5526. [Google Scholar] [CrossRef]
- Izquierdo Canas, P.M.; Palacios Garcia, A.T.; Garcia Romero, E. Enhancement of flavour properties in wines using sequential inoculations of non-Saccharomyces (Hansenula and Torulaspora) and Saccharomyces yeast starter. VITIS-J. Grapevine Res. 2011, 50, 177–182. [Google Scholar]
- Azzolini, M.; Fedrizzi, B.; Tosi, E.; Finato, F.; Vagnoli, P.; Scrinzi, C.; Zapparoli, G. Effects of Torulaspora delbrueckii and Saccharomyces cerevisiae mixed cultures on fermentation and aroma of Amarone wine. Eur. Food Res. Technol. 2012, 235, 303–313. [Google Scholar] [CrossRef]
- Benito, A.; Hofmann, T.; Laier, M.; Lochbüler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Rossouw, D.; Bauer, F.F. Exploring the phenotypic space of non-Saccharomyces wine yeast biodiversity. Food Microbiol. 2016, 55, 32–46. [Google Scholar] [CrossRef]
- Benito, S. Combined use of Lachancea thermotolerans and Schizosaccharomyces pombe in winemaking: A review. Microorganisms 2020, 8, 655. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Benito, S. Combined use of S. pombe and L. thermotolerans in winemaking. Beneficial effects determined through the study of wines’ analytical characteristics. Molecules 2016, 21, 1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benito, Á.; Calderón, F.; Benito, S. Mixed alcoholic fermentation of Schizosaccharomyces pombe and Lachancea thermotolerans and its influence on mannose-containing polysaccharides wine composition. AMB Express 2019, 9, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, C.; Sengler, F.; Solomon, M.; Curtin, C. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016, 209, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Kemp, B.; Alexandre, H.; Robillard, B.; Marchal, R. Effect of production phase on bottle-fermented sparkling wine quality. J. Agric. Food Chem. 2015, 63, 19–38. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-López, F.N.; Pérez-Torrado, R.; Querol, A.; Barrio, E. Modulation of the glycerol and ethanol syntheses in the yeast Saccharomyces kudriavzevii differs from that exhibited by Saccharomyces cerevisiae and their hybrid. Food Microbiol. 2010, 27, 628–637. [Google Scholar] [CrossRef] [PubMed]
- De Benedictis, M.; Bleve, G.; Grieco, F.; Tristezza, M.; Tufariello, M. An optimized procedure for the enological selection of non-Saccharomyces starter cultures. Antonie Van Leeuwenhoek 2011, 99, 189–200. [Google Scholar] [CrossRef]
- Ivit, N.N.; Loira, I.; Morata, A.; Benito, S.; Palomero, F.; Suárez-Lepe, J.A. Making natural sparkling wines with non-Saccharomyces yeasts. Eur. Food Res. Technol. 2018, 244, 925–935. [Google Scholar] [CrossRef]
- Kelly, J.M.; van Dyk, S.A.; Dowling, L.K.; Pickering, G.J.; Kemp, B.; Inglis, D.L. Saccharomyces uvarum yeast isolate consumes acetic acid during fermentation of high sugar juice and juice with high starting volatile acidity. OENO ONE Int. J. Vine Wine Sci. 2020, 54, 199–211. [Google Scholar]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ivit, N.N.; Longo, R.; Kemp, B. The Effect of Non-Saccharomyces and Saccharomyces Non-Cerevisiae Yeasts on Ethanol and Glycerol Levels in Wine. Fermentation 2020, 6, 77. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030077
AMA Style
Ivit NN, Longo R, Kemp B. The Effect of Non-Saccharomyces and Saccharomyces Non-Cerevisiae Yeasts on Ethanol and Glycerol Levels in Wine. Fermentation. 2020; 6(3):77. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030077
Chicago/Turabian StyleIvit, Nedret Neslihan, Rocco Longo, and Belinda Kemp. 2020. "The Effect of Non-Saccharomyces and Saccharomyces Non-Cerevisiae Yeasts on Ethanol and Glycerol Levels in Wine" Fermentation 6, no. 3: 77. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6030077
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.