Exploring the Impact of Lipid-Rich Food Industry Waste Carbon Sources on the Growth of Candida cylindracea DSM 2031


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Materials
2.3. Determination of the Fatty Acid Composition of Oils and Fats Used
2.4. Media and Culture Conditions
2.5. Determination of Biomass Yield
2.6. Determination of Extracellular Lipase Activity
2.7. Determination of Selected Culture Parameters
2.8. Determination of the Growth Curve of C. cylindracea
2.9. Measurement of the Yeast Optical Density
2.10. Measurement of the pH of the Culture Medium
2.11. Statistical Analysis
3. Results and Discussion
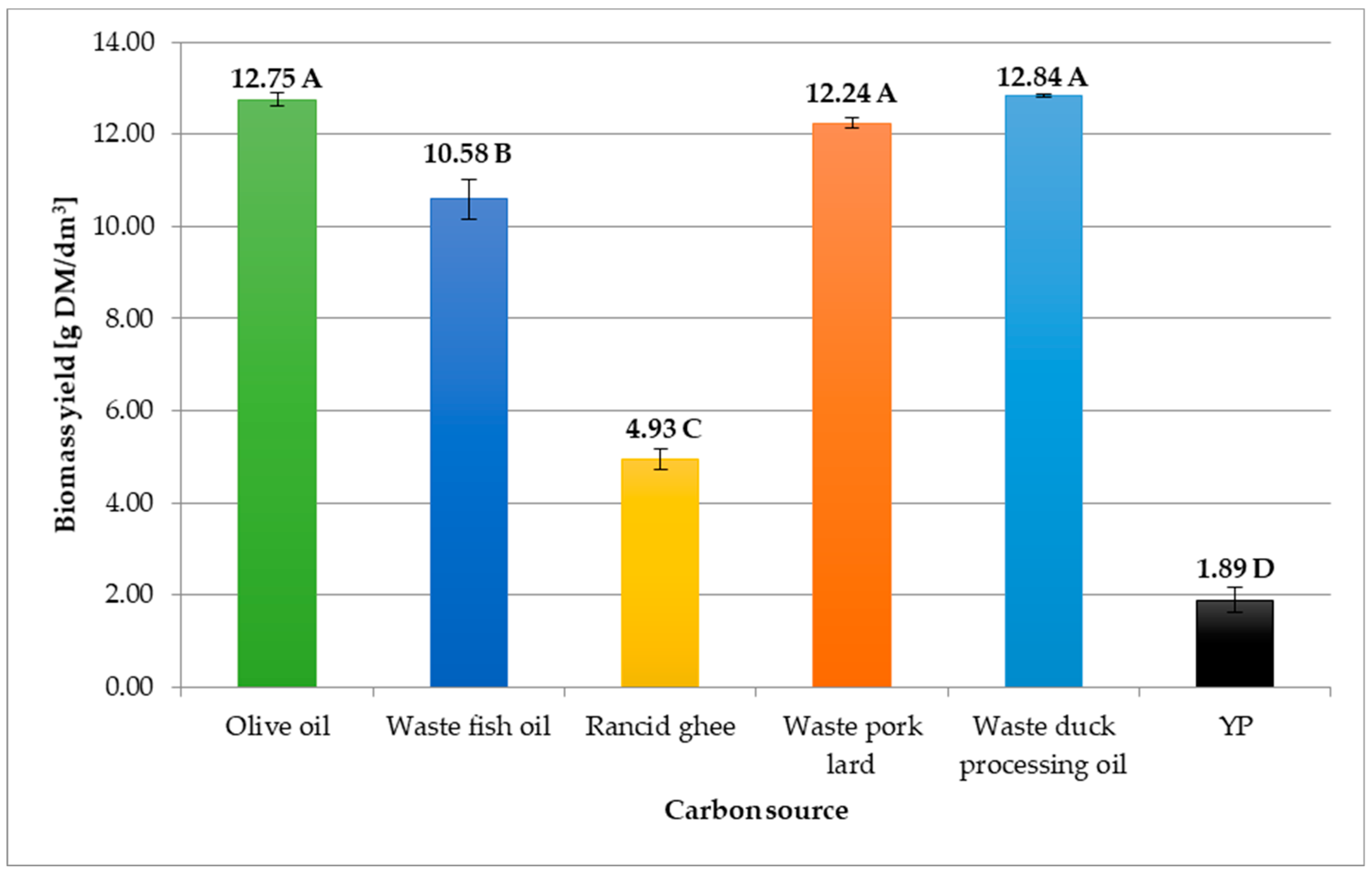
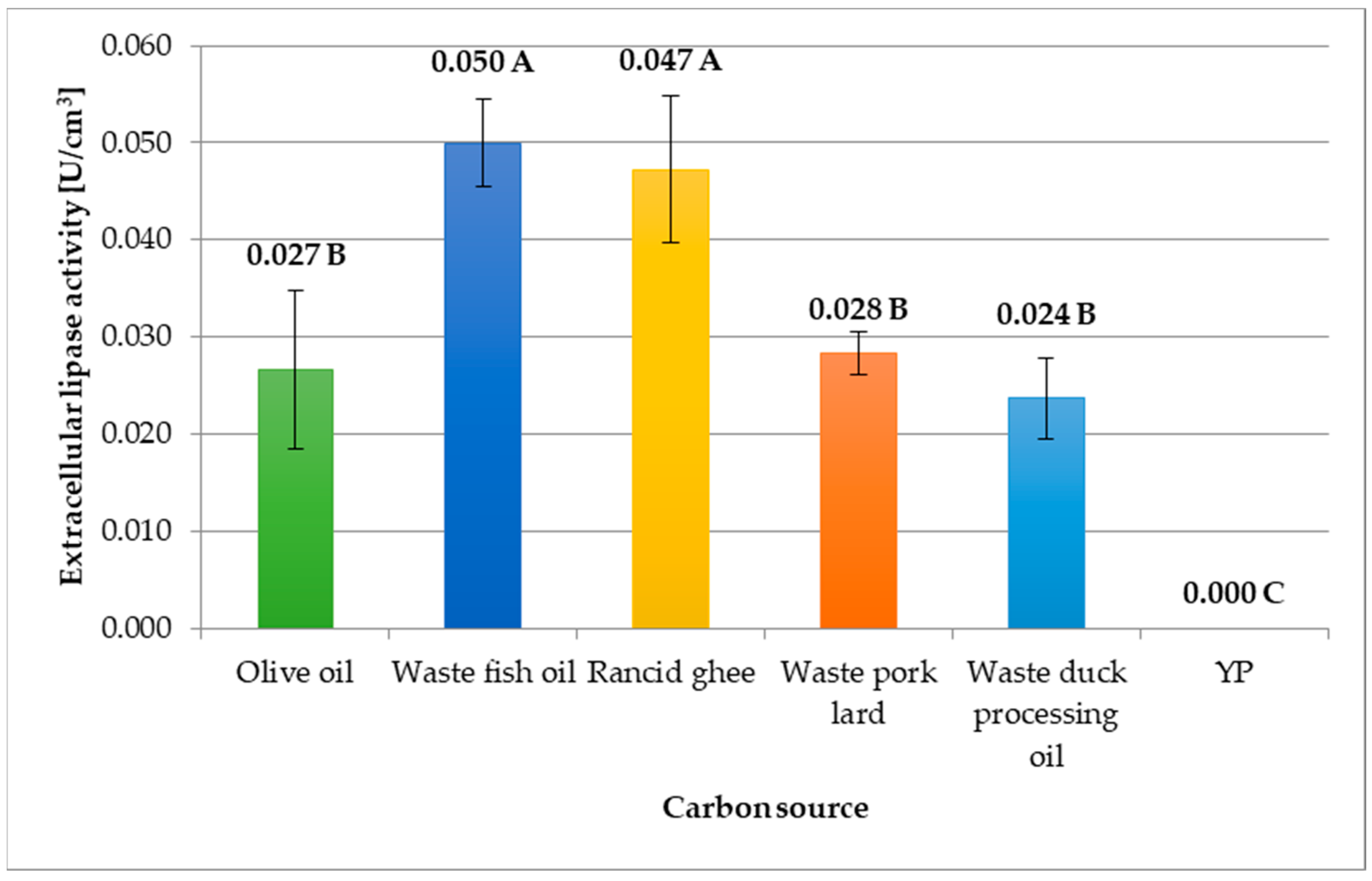
3.1. The Use of Lipid-Rich Food Industry Wastes in the Cultivation of C. cylindracea Yeast
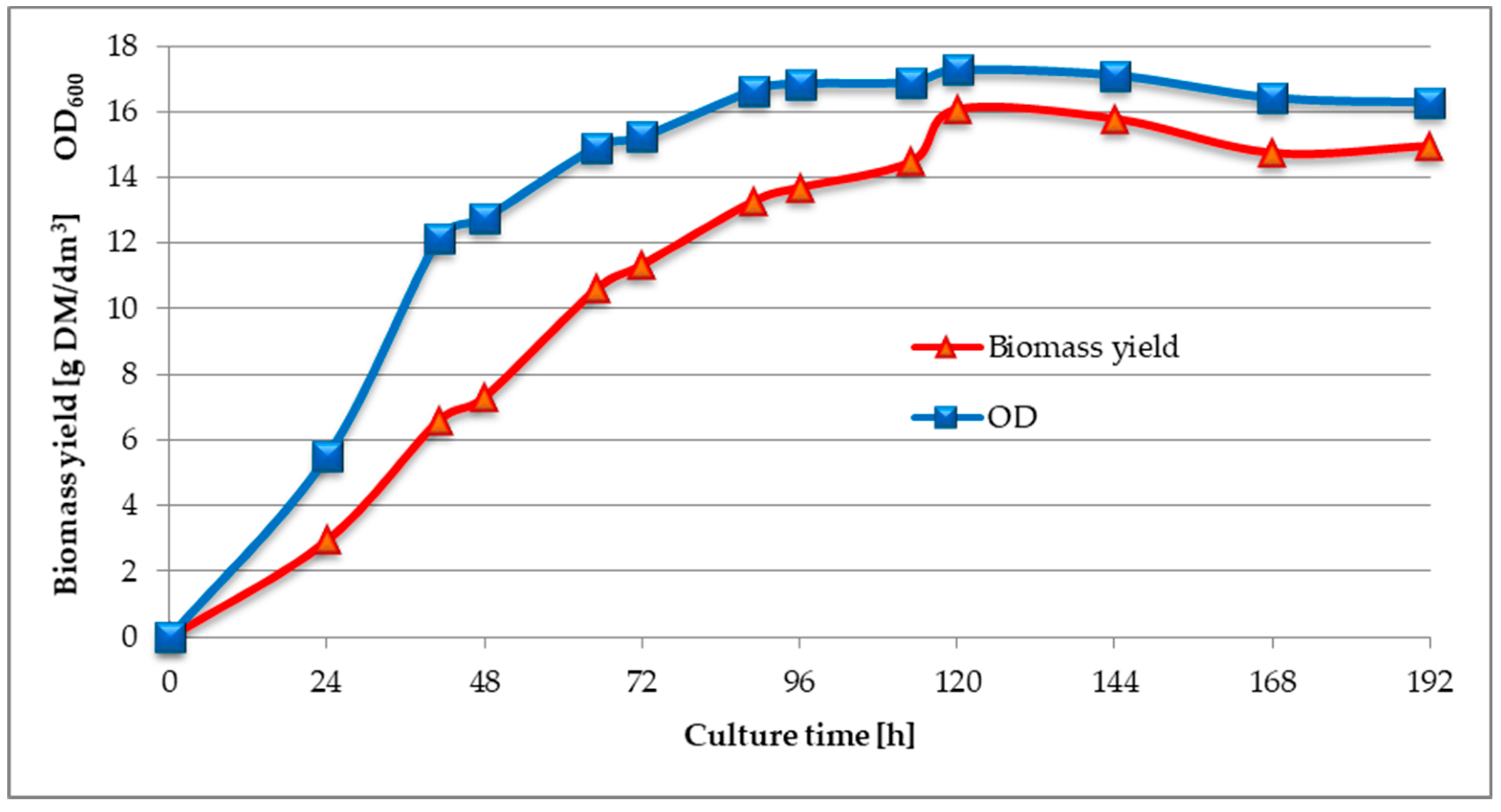
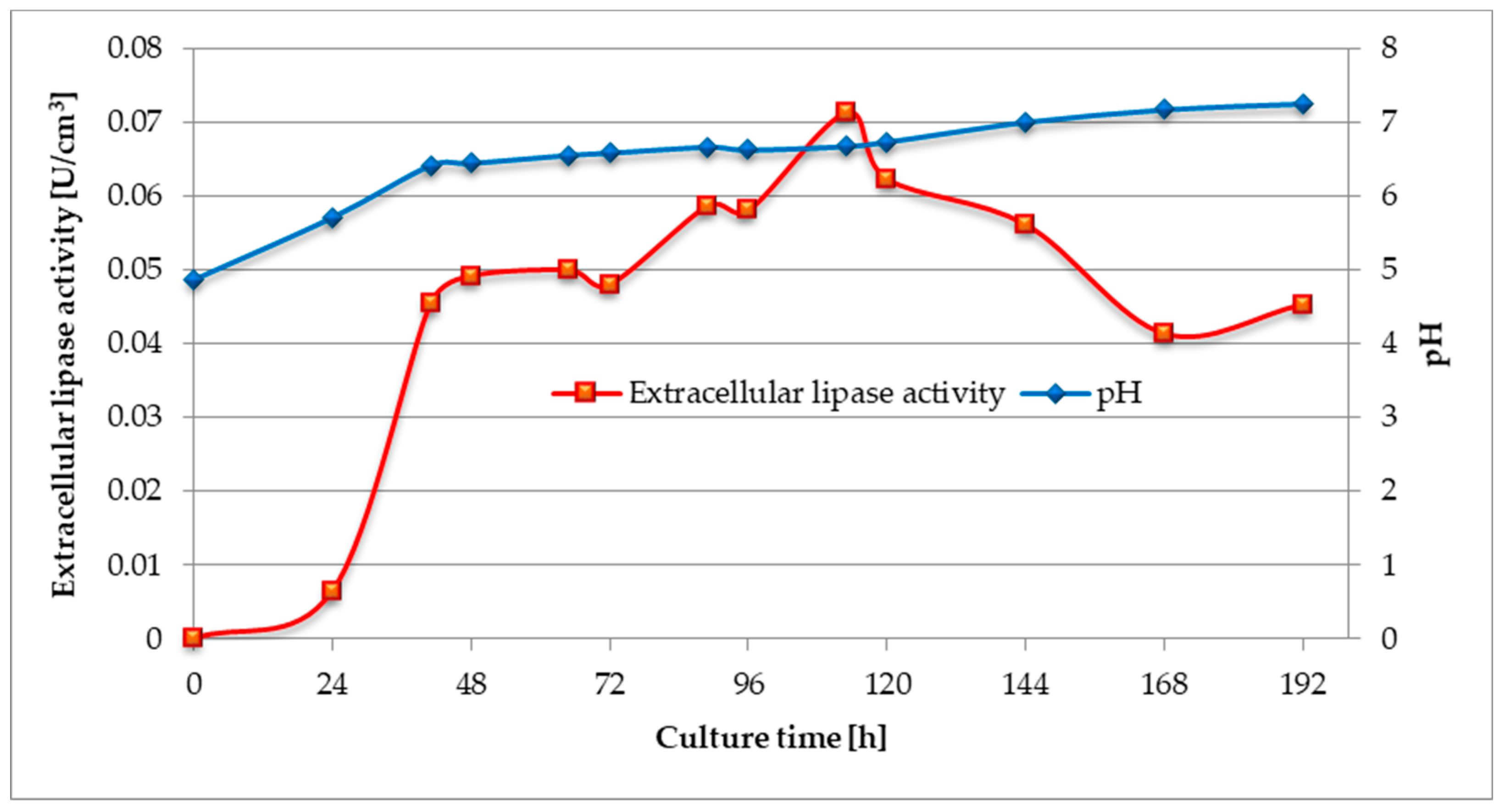
3.2. Investigation of Changes in Biomass Yield, Lipolytic Activity, and pH Value During the Culture of C. cylindracea in a Medium with Waste Fish Oil
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Domańska, W. Environment 2019; Statistics Poland: Warsaw, Poland, 2019; pp. 148–160.
- Hommel, R.K. CANDIDA Introduction. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 367–373. [Google Scholar]
- Lachance, M.A.; Boekhout, T.; Scorzetti, G.; Fell, J.W.; Kurtzman, C.P. Candida Berkhout (1923). In The Yeast, a Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 978–1278. [Google Scholar]
- Kurtzman, C.P.; Robnett, C.J. Indentification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Anton. Leeuw. 1998, 73, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Boontham, W.; Angchuan, J.; Boonmak, C.; Srisuk, N. Limtongozyma siamensis gen. nov., sp. nov., a yeast species in the Saccharomycetales and reassignment of Candida cylindracea to the genus Limtongozyma. Int. J. Syst. Evol. Microbiol. 2020, 70, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Machida, H. Studies on the production of lipase by microorganisms. Nippon Nogeikagaku Kaishi 1962, 36, 858–860. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.S.; Hou, C.T. Production of lipase by high density fed-batch culture of Candida cylindracea. Bioprocess Biosyst. Eng. 2006, 29, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Zalcain, I.; Zapalena, M.J.; Astiasaran, I.; Bello, J. Dry-fermented Sausages Elaborated with Lipase from Candida cylindracea. Comparison with Traditional Formulations. Meat Sci. 1995, 40, 55–61. [Google Scholar] [CrossRef]
- Belkacemi, F.Z.; Merabet-Khelassi, M.; Aribi-Zouioueche, L.; Riant, O. Productionof l-mentyl acetate through kinetic resolution by Candida cylindracea lipase: Effect of alkaloids as additives. Res. Chem. Intermed. 2018, 44, 6847–6860. [Google Scholar] [CrossRef]
- Alcântara, T.D.A.P.; Oliveira, J.M.; Evangelista-Barreto, N.S.; Marbac, P.A.S.; Cazetta, M.L. Aerobic decolorization of azo dye orange g by a new yeast isolate Candida cylindracea SJL6. Biosci. J. 2017, 33, 1340–1350. [Google Scholar] [CrossRef]
- Tang, A.; Zhang, Y.; Wei, T.; Wu, J.; Li, Q.; Liu, Y. Immobilization of Candida cylindracea Lipase by Covalent Attachment on Glu-Modified Bentonite. Appl. Biochem. Biotechnol. 2019, 187, 870–883. [Google Scholar] [CrossRef]
- Zieniuk, B.; Fabiszewska, A.; Białecka-Florjańczyk, E. Screening of solvents for favoring hydrolytic activity of Candida antarctica Lipase B. Bioproc. Biosyst. Eng. 2020, 43, 605–613. [Google Scholar] [CrossRef]
- Zieniuk, B.; Wołoszynowska, M.; Białecka-Florjańczyk, E. Enzymatic synthesis of biodiesel by direct transesterification of rapeseed cake. Int. J. Food Eng. 2020, 16, 3. [Google Scholar] [CrossRef]
- Kapturowska, A.; Stolarzewicz, I.; Krzyczkowska, J.; Białecka-Florjańczyk, E. Studies on lipolytic activity of sonicated enzymes from Yarrowia lipolytica. Ultrason. Sonochem. 2012, 19, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Sokolovska, I.; Albasi, C.; Riba, J.P.; Bales, V. Production of extracellular lipase by Candida cylindracea CBS 6330. Bioprocess Biosyst. Eng. 1998, 19, 179–186. [Google Scholar] [CrossRef]
- Fickers, P.; Marty, A.; Nicaud, J.M. The lipases from Yarrowia lipolytica: Genetics, production, regulation, biochemical characterization and biotechnological applications. Biotechnol. Adv. 2011, 29, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.H.; Gill, C.O. Batch growth of Saccharomycopsis lipolytica on animal fats. Appl. Microbiol. Biotechnol. 1985, 21, 292–298. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Selective uptake of fatty acids by the yeast Yarrowia lipolytica. Eur. J. Lipid Sci. Technol. 2003, 105, 651–655. [Google Scholar] [CrossRef]
- Lie, O.; Lambertsen, G. Fatty Acid Specificity of Candida cylindracea Lipase. Eur. J. Lipid Sci. Technol. 1986, 88, 365–367. [Google Scholar] [CrossRef]
- Fabiszewska, A.U.; Białecka-Florjańczyk, E. Factors influencing synthesis of extracellular lipases by Yarrowia lipolytica in medium containing vegetable oils. J. Microbiol. Biotechnol. Food Sci. 2014, 4, 231–237. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, C.; Lopes, M.; Ferreira, J.P.; Belo, I. Biological treatment of olive mill wastewater by non-conventional yeasts. Bioresour. Technol. 2009, 100, 3759–3763. [Google Scholar] [CrossRef] [Green Version]
- Vylkova, S.; Carmar, A.J.; Danhof, H.A.; Collete, J.R.; Zhou, H.; Lorenz, M.C. The fungal pathogen Candida albicans autoinduces hyphal morphogenesis by raising extracellular pH. mBio 2011, 2, e00055-11. [Google Scholar] [CrossRef] [Green Version]
- Krastanov, A.; Govindarajan, A.; Daniel, D. Studies on lipase fermentation using Candida cylindracea NRRL Y-175061N a stirred tank bioreactor. Bulg. J. Agric. Sci. 2008, 14, 290–299. [Google Scholar]
- Fabiszewska, A.U.; Stolarzewicz, I.; Zamojska, W.; Białecka-Florjańczyk, E. Carbon source impact on Yarrowia lipolytica KKP 379 lipase production. Appl. Biochem. Microbiol. 2014, 50, 404–410. [Google Scholar] [CrossRef]
- Salihu, A.; Alam, Z.M.; AbdulKarim, M.I.; Salleh, H.M. Optimization of lipase production by Candida cylindracea in palm oil mill effluent based medium using statistical experimental design. J. Mol. Catal. B Enzym. 2011, 69, 66–73. [Google Scholar] [CrossRef]
- Muralidhar, R.V.; Chirumamila, R.R.; Marchant, R.; Nigam, P. A response surface approach for the comparison of lipase production by Candida cylindracea using two different carbon sources. Biochem. Eng. J. 2001, 9, 17–23. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid Profiles [%] | |||||
|---|---|---|---|---|---|
| Olive Oil | Waste Fish Oil | Rancid Ghee | Waste Pork Lard | Waste Duck Processing Oil | |
| SFA1 | 14.0 | 24.3 | 68.2 | 42.3 | 36.6 |
| UFA | 85.0 | 75.6 | 29.8 | 55.7 | 59.4 |
| MUFA | 78.2 | 51.0 | 27.5 | 41.9 | 45.4 |
| PUFA | 6.8 | 24.6 | 2.3 | 13.8 | 14.0 |
| Y(P/X) [U/g] 1 | Y(X/S) [g/g] | qp [U/g × h] | vp [U/L × h] | |
|---|---|---|---|---|
| Olive oil | 2.09 | 0.64 | 0.03 | 0.41 |
| Waste fish oil | 4.72 | 0.53 | 0.07 | 0.77 |
| Rancid ghee | 9.57 | 0.25 | 0.15 | 0.73 |
| Waste pork lard | 2.31 | 0.61 | 0.04 | 0.44 |
| Waste duck processing oil | 1.85 | 0.64 | 0.03 | 0.36 |
| YP | 0.00 | 0.09 | 0.00 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zieniuk, B.; Mazurczak-Zieniuk, P.; Fabiszewska, A. Exploring the Impact of Lipid-Rich Food Industry Waste Carbon Sources on the Growth of Candida cylindracea DSM 2031. Fermentation 2020, 6, 122. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040122
Zieniuk B, Mazurczak-Zieniuk P, Fabiszewska A. Exploring the Impact of Lipid-Rich Food Industry Waste Carbon Sources on the Growth of Candida cylindracea DSM 2031. Fermentation. 2020; 6(4):122. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040122
Chicago/Turabian StyleZieniuk, Bartłomiej, Patrycja Mazurczak-Zieniuk, and Agata Fabiszewska. 2020. "Exploring the Impact of Lipid-Rich Food Industry Waste Carbon Sources on the Growth of Candida cylindracea DSM 2031" Fermentation 6, no. 4: 122. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040122