Hydrolyzed Agricultural Residues—Low-Cost Nutrient Sources for l-Lactic Acid Production

Thünen-Institute of Agricultural Technology, Bundesallee 47, 38116 Braunschweig, Germany
*
Author to whom correspondence should be addressed.
Fermentation 2020, 6(4), 97; https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040097
Submission received: 31 August 2020
/
Revised: 25 September 2020
/
Accepted: 27 September 2020
/
Published: 6 October 2020
(This article belongs to the Special Issue Lactic Acid Fermentation and the Colours of Biotechnology 2.0)
Abstract
:Lactic acid is a building block for polylactic acid, which is one of the most promising polymers based on renewable resources and is used mainly in packaging industry. This bio-based polymer is biodegradable and provides an ecological and economical alternative to petrochemical plastics. The largest cost blocks of biotechnological lactic acid production, accounting for up to 38% of the total costs, are substrate and nutrient sources, such as peptone, meat, and yeast extract. Based on a systematic analysis of nutritional requirements, the substitution of yeast extract by low-cost protein-rich agricultural hydrolysates was estimated for the production of l-lactic acid with Lactobacillus casei. Cultivations in 24-well microtiter plates enabled analysis of nutrient requirements and the usage of various hydrolysates with a high parallel throughput and repeated sampling. Rapeseed meal (RM) and distillers’ dried grains with solubles (DDGS) were tested as low-cost protein-rich agricultural residues. By using chemically or enzymatically hydrolyzed rapeseed meal or DDGS, 70% of the nutrient sources was replaced in the fermentation process at identical productivity and product yields. All in all, the total costs of l-lactic acid production with Lactobacillus casei could potentially be reduced by up to 23%.
1. Introduction
Bio-based building blocks for the polymer and packaging industries compete with low-priced petrochemical plastics. To provide an ecological and economical alternative to petrochemical plastics, not only yield, productivity, and the final titers of biotechnological processes have to be optimized for monomer production but also the costs of production, especially medium costs, must be reduced. Besides the substrate, nutrient sources like yeast extract, with a price of 9–10 €/kg, incur the highest production costs [1].
Polylactic acid (PLA), a bio-based and biodegradable polyester derived from lactic acid, is one of the most promising alternatives to petrochemical plastics [2]. Ninety percent of lactic acid is produced by bacterial fermentation [3], and the nutritional requirements of lactic acid bacteria are high because of their auxotrophy towards amino acids and vitamins [4,5,6,7,8]. In 2016, 1220 kt of lactic acid was required, and the demand of the global market is predicted to increase to around 1960 kt by 2025 [8]. Optical pure l- or d-lactic acids can be produced by the appropriate lactic acid bacteria, which is advantageous over chemical production wherein a racemic mix of dl-lactic acid is formed [2]. There is a need for optically pure l- or d-lactic acids, since the properties of PLA depend on the enantiomeric ratio of l- to d-lactic acid. [6,9]. Moreover, pure l-lactic acid is used in the pharmaceutical and food industries [10].
Techno-economic evaluations have indicated yeast extract to be a high-cost factor of lactic acid production, contributing up to 38% of the total costs [11,12,13]. Yeast extract and other complex nutrient sources are used in fermentations because they provide besides organic nitrogen, like amino acids, vitamins, salts, trace elements, or nucleic acids [14]. Alternative sources of nutrients in the form of low-cost and protein-rich agricultural residues [15,16,17], such as rapeseed meal (RM) or distillers’ dried grains with solubles (DDGS), can be used to replace yeast extract. However, the high molecular structures of the agricultural residues have to be disintegrated by enzymatic or chemical hydrolysis prior to use.
The major problem of the common hydrolysates is the insufficient availability of free amino acids and small peptides, a problem caused by an insufficient degree of hydrolysis, which results in a lower productivity of lactic acid [16]. A hydrolysis method with 3m H2SO4 was developed in a previous study, and yeast extract was successfully substituted by hydrolysates of low-cost protein-rich agricultural residues based on a systematic analysis of the nutritional requirements for the production of d-lactic acid using Sporolactobacillus inulinus [16,18]. In the present study, this knowledge is transferred to l-lactic acid production using Lactobacillus casei, and the nutrient requirements of the strain are identified. The aim of this study is to minimize the use of expensive complex nutrient sources and to supplement them with low-cost hydrolysates of agricultural residues resulting an equal productivity, yield, and final titer of l-lactic acid.
2. Materials and Methods
Lactobacillus casei ATCC 393 was purchased from the American Type Culture Collection (Manassas, USA). The preculture was inoculated with a cryo stock culture that was stored at –80 °C in 50% (v/v) glycerol and incubated in 100 mL shaking flasks for 17 h at 38 °C and 0 rpm. For preculture and cultivation, MRS (deMan, Rogosa, und Sharpe) medium was used: 20 g·L−1 (preculture) or 120 g·L−1 (cultivation) glucose; 10 g·L−1 meat extract (Carl Roth GmbH and Co. KG, Karlsruhe, Germany); 5 g·L−1 Fermtech yeast extract (Merck, Darmstadt, Germany); 10 g·L−1 peptone (Fluka, Buchs, Switzerland); and 2 g·L−1 K2HPO4, 2 g·L−1 (NH4)2HCitrat, 5 g·L−1 sodium acetate, 0.2 g·L−1 MgSO4, 50 mg·L−1 MnSO4, and 90 g·L−1 pulverized CaCO3 (Carl Roth, Karlsruhe, Germany). In the modified MRS medium, 5 g·L−1 meat extract and 5 g·L−1 yeast extract and, in limited MRS medium, 3 g·L−1 yeast extract were used instead of the mixture of meat extract, yeast extract, and peptone. A total nitrogen content of 1.11 g·L−1 was used for cultivation with hydrolysates and combinations of yeast extract and hydrolysate, which corresponded to a total nitrogen content of 5 g·L−1 meat extract and 5 g·L−1 yeast extract. The cultivations were inoculated with 2% (v/v) and processed in 24-well plates with 1.5-mL culture volumes (Sarstedt, Nümbrecht, Germany) at 37 °C and 400 rpm in minimum duplicates. The mean values calculated from these duplicate experiments and the corresponding standard deviations (error bars) are presented in the figures. All solutions were sterilized by autoclaving for 20 min at 121 °C.
The B-vitamin solution and amino acid solution were calculated for the modified MRS medium with 5 g·L−1 meat extract and 5 g·L−1 yeast extract based on the results of [18]. The B-vitamin mix contained 1.25 mg·L−1 thiamine∙HCl (B1), 500 µg·L−1 riboflavin (B2), 10 mg·L−1 niacin (B3), 2.5 mg·L−1 Ca-pantothenate (B5), 1 mg·L−1 pyridoxine ∙ HCl (B6), 20 µg·L−1 biotin (B7), 750 µg·L−1 folic acid (B9), 1 µg·L−1 cyanocobalamin (B12), and 75 mg·L−1 inositol. The amino acid mix was prepared with 375 µg·L−1 alanine (Ala), 115 µg·L−1 asparagine (Asn), 790 µg·L−1 aspartic acid (Asp), 1395 µg·L−1 glutamic acid (Glu), 360 µg·L−1 glycine (Gly), 235 µg·L−1 histidine (His), 316 µg·L−1 isoleucine (Ile), 563 µg·L−1 leucine (Leu), 560 µg·L−1 lysine (Lys), 133 µg·L−1 methionine (Met), 323 µg·L−1 phenylalanine (Phe), 536 µg·L−1 proline (Pro), 412 µg·L−1 serine (Ser), 370 µg·L−1 threonine (Thr), 81 µg·L−1 tryptophan (Trp), 148 µg·L−1 tyrosine (Tyr), and 447 µg·L−1 valine (Val).
Distillers’ dried grains with solubles (DDGS) was purchased from Crop Energies AG (Mannheim, Germany), and rapeseed meal (RM) was purchased from Archer Daniels Midland AG (Chicago, IL, USA). The total nitrogen content of DGGS was 26.6 mg∙g−1 and of RM was 48.8 mg∙g−1. The agricultural residues are characterized in detail in [18].
For hydrolysis, the agricultural residues were ground with an ultra-centrifugal mill (750 µm, ZM 200, Retsch, Haan, Germany) and sieved to a particle size of less than 710 µm. In cases of chemical hydrolysis, 400 g of the agricultural residue was continuously mixed with 1.2 L of 3 M H2SO4 in a 2-L triple-necked round-bottom flask with a reflux condenser for 24 h at 110 °C. After cooling down to room temperature, the pH value of the hydrolysate was adjusted to 6 by adding Ca(OH)2. The slurry was centrifuged (4600× g, 20 min, and 10 °C), and the pellet was washed twice with 400 mL water. The supernatants were collected and filtered by a pleated filter (MN 615 ¼ Ø 150 mm, Macherey-Nagel, Düren, Germany). The pH value of the hydrolysate was adjusted to 6.5 using 1 M NaOH, the hydrolysate was sterilized for 20 min at 121 °C, and the total nitrogen content was determined.
For enzymatic hydrolysis, Protex™ 6L (DuPont™ Genencor, Rochester, NY, USA) was used; 4.4 g agricultural residue was weighed into a 250-mL bottle and mixed with 70 mL of water, and the pH was adjusted to pH 9.5. A 1% enzyme solution, referred to as the protein concentration, was added to the suspended agricultural residue and incubated at 60 °C with 100 rpm. After 48 h of incubation, the pH value of the hydrolysate was adjusted to 6.5 with 0.5 M H2SO4 and autoclaved for 20 min at 121 °C and the total nitrogen content was determined.
Lactic acid and glucose concentrations were measured by high-performance liquid chromatography (HPLC). A Dionex ICS-5000 system from Thermo scientific (Sunnyvale, CA, USA) equipped with an Aminex HPX-87H column (300 × 7.8 mm) (Bio-Rad, Hercules, CA, USA) and a refraction index detector (RI-101, Shodex Tokyo, Japan) was used at 60 °C. The mobile phase was 5 mM H2SO4 at a flow rate of 0.7 mL min−1. Fifty microliters of the sample were first dissolved in 950 µL of 50 mM H2SO4 and centrifuged at 20,800× g for 10 min at room temperature (RT), and the supernatant was diluted with 50 mM H2SO4.
The total nitrogen content (TN) was determined by Kjeldahl method. A 150 mg sample in a Kjeldahl reaction tube with boiling stones was mixed with an antifoam tablet (Carl Roth, Karlsruhe, Germany) and a Kjeltab S (5 g K2SO4, 5 mg Se) as well as 10 mL 96% H2SO4. The thermal degradation was performed in a Kjeldathrem (Gerhardt GmbH & Co. KG, Königswinter, Germany) with the following temperature gradient: 100 °C (30 min), 200 °C (60 min), 300 °C (60 min), 420 °C (60 min), and RT (30 min). Distillation and titration were carried out with a Vapodest (Gerhardt GmbH & Co. KG, Königswinter, Germany) using 33% NaOH, 2% B(OH)3, and 0.05 M H2SO4.
For the determination of free amino acids, an analysis kit (EZ:faas GC-FID Physiological) from Phenomenex (Torrance, CA, USA) was used. For peptide-bounded amino acids, 50 mg of a dried sample was hydrolyzed with 1 mL 6 M HCl + 0.02% phenol in a headspace vial (cleaned with concentrated HCl and water). The vial was sealed with a septum, deep-frozen in liquid nitrogen, evacuated, and heated to 110 °C for 48 h. The hydrolysate was diluted to 100 mL and filtered. Free amino acids were analyzed according to the manufacturer’s instructions. A gas chromatograph with a flame ionization detector GC-17A from Shimadzu (Kyoto, Japan) with a Zebron ZB-AAA column (10 m × 0.25 mm × 0.25 μm) from Phenomenex (Torrance, USA) was used for the measurement with H2 as carrier gas with a flow rate of 2.15 mL min−1. The temperature of 110 °C was heated up to 250 °C with a gradient of 20 °C min−1, further up to 320 °C with 10 °C min−1, and was held for 1 min.
3. Results
3.1. Reference Cultivation and Variation of the Complex Nutrient Sources in MRS Medium
To compare different compositions and concentrations of the complex nutrient sources for yeast extract, peptone, and meat extract in MRS medium, combinations of 0, 3, 5, and 10 g·L−1 of the different complex medium components were tested. L. casei was cultivated in 24-well plates, and the final l-lactic acid titer after 70.5 h and the maximum productivity were determined and listed in Table 1. The combination of 5 g·L−1 yeast extract, 10 g·L−1 peptone, and 10 g·L−1 meat extract was defined as first reference cultivation and resulted in a final titer of 106.3 g·L−1 l-lactic acid, and a maximum productivity of 4.63 g·L−1·h−1 was obtained. Comparable results of final titer (>100 g·L−1 l-lactic acid) and maximum productivity (>4.3 g·L−1·h−1) were also achieved with a combination of 5 g·L−1 yeast extract and 5 g·L−1 meat extract with and without addition of peptone, whereas the total nitrogen concentration was reduced by 42% and 67% compared to the reference. A complete conversion of glucose to l-lactic acid could not be achieved with the single use of peptone, yeast extract, or meat extract in concentrations ≤ 5 g·L−1. Based on these results, the modified MRS medium with 5 g·L−1 yeast extract and 5 g·L−1 meat extract was chosen as a new reference for further cultivations (in the following, simply called reference).
To examine nutrition requirements by nutrition limitation, 3 g·L−1 yeast extract was used in limited MRS medium (Table 1).
3.2. Nutrient Requirements
To reduce the complex nutrient source and to use low-cost hydrolysates of agricultural residues instead in order to reach an equal productivity, yield, and final titer of L-lactic acid, the nutrient requirements of Lactobacillus casei need to be determined first. Among others, yeast extract is comprised of important nutrients, like amino acids and vitamins. Therefore, their influence was examined in more detail.
3.2.1. B-Vitamin Requirements
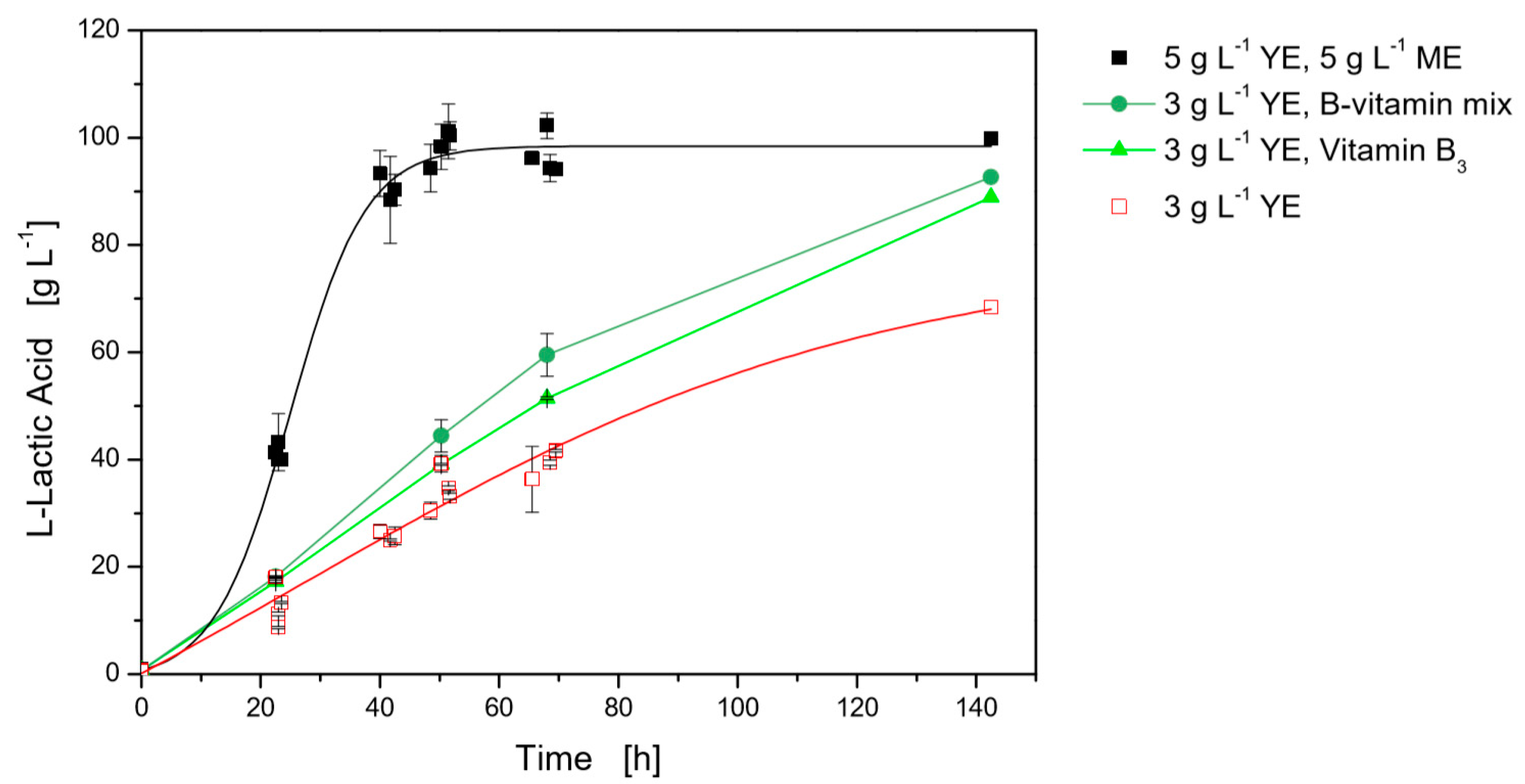
Based on a study of Klotz et al. [18], that analyzed the composition of the used yeast and meat extract, a synthetic B-vitamin mix was designed. The tested concentration of each vitamin corresponds to a mixture of 5 g·L−1 yeast extract and 5 g·L−1 meat extract. To identify B-vitamin requirements for L. casei, each vitamin was added separately to limited MRS medium with 3 g·L−1 yeast extract (Figure 1). With the nutrition limitation, an l-lactic acid concentration of 68.4 g·L−1 was reached after 140 h. All single vitamin addition, except niacin (Vitamin B3), resulted in 67.3 g·L−1 ± 5.5 g·L−1 l-lactic acid with a remaining glucose concentration of 52.4 g·L−1 ± 5.5 g·L−1, and the added vitamins did not influence cultivation in the tested concentrations. The addition of niacin had a considerable positive impact on the cultivation, resulting in a final titer of 88.9 g·L−1 l-lactic acid and a yield of 0.90 (w/w) ± 0.01 (w/w). This titer was slightly lower than the cultivation with the B-vitamin mix, with 92.6 g·L−1 l-lactic acid and a yield of 0.93 (w/w) ± 0.02 (w/w). Also, the tests with addition of niacin and the total B-vitamin mix did not result in a total conversion of the substrate in 140 h, and 18.6 g·L−1 ± 2.6 g·L−1 glucose still remained. In total, the maximum productivity was clearly below the maximum productivity of the reference cultivation with modified MRS medium (5 g·L−1 yeast extract and 5 g·L−1 meat extract), which is on average 3.9 g·L−1·h−1, whereas the yield was constant at 0.93 (w/w) ± 0.05 (w/w). On average, the max. productivity of the single tested vitamins was 0.93 g·L−1·h−1, using niacin the value increased to 0.99 g·L−1·h−1 and for the B-vitamin mix to 1.15 g·L−1·h−1.
3.2.2. Amino Acid Requirements and the Influence of Different Medium Components
Similar to the investigation of B-vitamin requirements, amino acid (AA) requirements were evaluated by adding an amino acid-mix to the medium. Like the B-vitamin mix, the amino acid mix is based on composition and concentration of the amino acid profile of 5 g·L−1 yeast extract and 5 g·L−1 meat extract. Without using any complex media compound, and supplementing the media with the amounts of B-vitamins and amino acid that exists in 5 g L −1 yeast and 5 g L−1 meat extract (synthetic MRS medium), a l-lactic acid titer of 10.9 g·L−1 with a yield of 0.55 (w/w) ± 0.04 (w/w) after 70.5 h was achieved, whereas the reference cultivation with 5 g·L−1 yeast extract and 5 g·L−1 meat extract resulted in approximately 99 g·L−1 l-lactic and a yield of 0.91 (w/w) ± 0.02 (w/w) and total glucose consumption. Even the limited MRS medium with 3 g·L−1 yeast extract had an l-lactic acid titer of approximately 40 g·L−1, which was noticeably higher than the synthetic MRS medium.
Therefore, several variations using the limited MRS medium with 3 g·L−1 yeast extract and the supplementation of AA-mix and B-vitamin mix were performed. The supplementation of amino acids contained in 5 g·L−1 yeast extract and 5 g·L−1 meat extract led to a slightly increased titer of 50.8 g·L−1 l-lactic acid after 70.5 h (data not shown). A further, slight increase to 60.9 g·L−1 l-lactic acid after 70.5 h, with a comparable yield, was achieved using 3 g·L−1 yeast extract and the supplementation of the AA-mix and B-vitamin mix with the composition and concentration contained in 5 g·L−1 yeast extract and 5 g·L−1 meat extract (data not shown). However, a total consumption of glucose was not possible during that cultivation time of 70.5 h. To point out potential amino acid limitations, the concentration of free amino acids in the medium was analyzed after cultivation. It was noticed, that the concentration of tryptophan was below the detection limit. The amino acids arginine and cysteine could not be determined by the used analytical method. To estimate the influence of these three amino acids a mixture of 150 mg·L−1 arginine, 50 mg·L−1 cysteine and 80 µg·L−1 tryptophan (Arg/Cys/Trp) was added additionally to the medium, without any supply of a complex media source (synthetic media). The l-lactic acid titer of 30.9 g·L−1 in synthetic medium without yeast extract and Arg/Cys/Trp was still lower than the limited MRS medium with 3 g·L−1 yeast extract, achieving approximately 40 g·L−1 l-lactic acid.
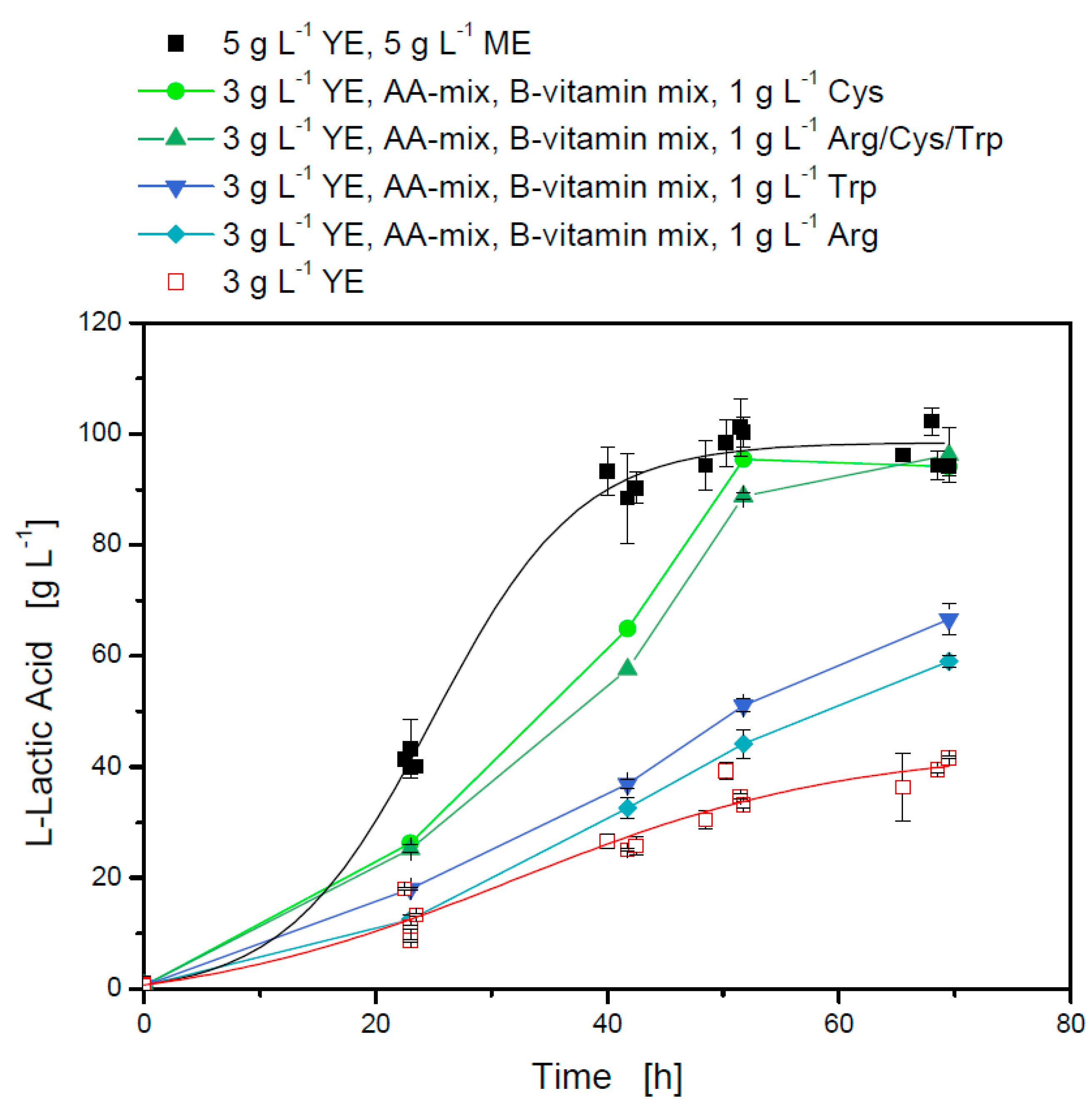
If the limited MRS medium with 3 g·L−1 yeast extract was combined with B-vitamin mix, amino acid-mix and Arg/Cys/Trp, the cultivation was positively influenced (Figure 2). Glucose was totally consumed during the cultivation time and 95.2 g·L−1 l-lactic acid was produced with a yield of 0.89 (w/w) ± 0.02 (w/w). The final titer of this cultivation was comparable to the reference cultivation using 5 g·L−1 yeast extract and 5 g·L−1 meat extract, but the maximum productivity differed. The maximum productivity was on average 3.9 g·L−1·h−1 for the reference cultivation and 2.19 g·L−1·h−1 for the cultivation with limited MRS medium with B-vitamin mix, amino acid-mix, and Arg/Cys/Trp. A further reduction of yeast extract resulted in a decreased l-lactic acid titer without total glucose consumption (data not shown).
To determine the single influence of Arg, Cys, and Trp, 1 g·L−1 of each amino acid was separately added to limited MRS medium with 3 g·L−1 yeast extract, amino acid-mix, and B-vitamin mix (Figure 2). All three amino acids showed a positive effect on the l-lactic acid production and amounted in a final titer of 59.1 g·L−1 for Arg, 66.6 g·L−1 for Trp, and 94.1 g·L−1 for Cys and a yield of 0.90 (w/w) ± 0.04 (w/w), whereas in limited MRS medium, 40 g·L−1 l-lactic acid was produced.
3.3. Agriculture Residue Hydrolysates as Nutrition Source
Instead of complex medium components, different agricultural residue hydrolysates were tested. Rapeseed meal (RM) and distillers’ dried grains with solubles (DDGS) were hydrolyzed either chemically or enzymatically.
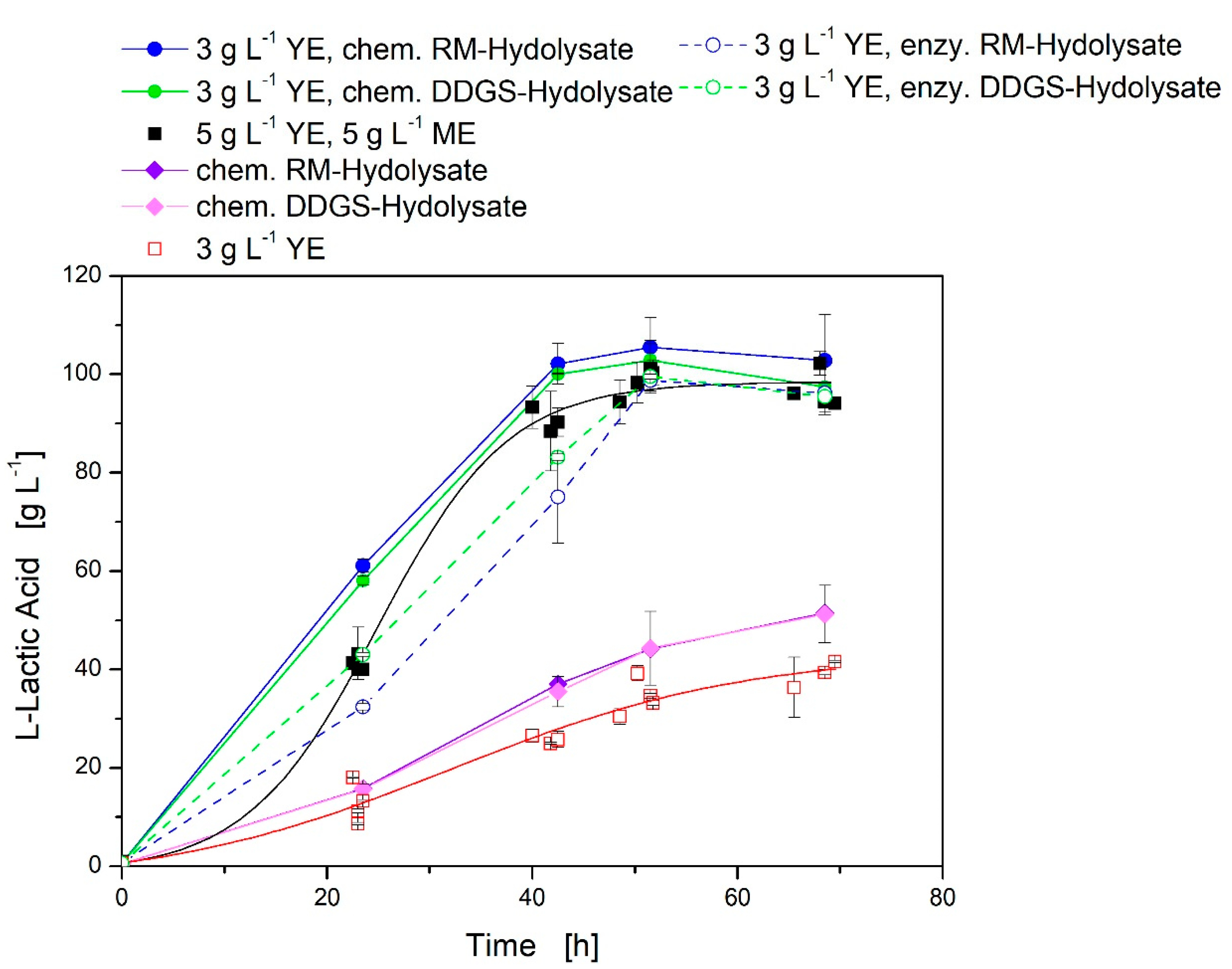
The hydrolysates were tested without the use of yeast extract and with limited MRS medium with 3 g·L−1 yeast extract (Figure 3). For all investigations with hydrolysates, the total nitrogen content was comparable to 5 g·L−1 yeast extract and 5 g·L−1 meat extract. Using hydrolysates without the addition of yeast or meat extract, the l-lactic acid titer averaged 51.4 g·L−1 after 68.5 h, which was half of the titer of the reference cultivation with 5 g·L−1 yeast extract and 5 g·L−1 meat extract.
Adding 3 g·L−1 yeast extract to the MRS medium (limited MRS medium) with either of the chemically hydrolyzed hydrolysates, the titer of 101.7 g·L−1 ± 2.7 g·L−1 was comparable to the reference cultivation. The yield of all cultivations was comparable and amounted to 0.84 (w/w) ± 0.04 (w/w), except cultivation with limited MRS medium (with 3 g·L−1 yeast extract), which yielded in 0.71 (w/w) ± 0.01 (w/w).
Using enzymatic hydrolysis for the digestion of RM and DDGS and under the addition of 3 g·L−1 yeast extract to the MRS medium (limited MRS medium), the l-lactic acid titer was slightly lower compared to the reference but not as good as when using the chemical hydrolysis (Figure 3).
4. Discussion
The composition of the standard MRS medium for cultivation of Lactobacilli [19] was examined with regard to the combination and concentrations of the complex nutrient sources of yeast extract, peptone, and meat extract for l-lactic acid production with L. casei. Combinations of 5 g·L−1 yeast extract, 5 g·L−1 peptone and 5 g·L−1 meat extract, or 5 g·L−1 yeast extract and 5 g·L−1 meat extract resulted in a final titer of >100 g·L−1 l-lactic acid, comparable to the reference cultivation with 5 g·L−1 yeast extract, 10 g·L−1 peptone, and 10 g·L−1 meat extract, which had the highest total calculated nitrogen content of TN = 2.92 g·L−1. The total nitrogen content was not the key factor, as in the studies of Olmos-Dichara et al. [20], Amrane and Prigent [21], and Nancib et al. [22], but the addition of the used nutrient source was the key factor. These nutrient sources differed, beside TN, in vitamins, free amino acids, peptides/proteins, fatty acids, and trace elements [18], which are responsible for bacterial growth and biological activity [23]. Hujanen and Linko [24], Kwon et al. [25], and Klotz et al. [18] identified that lactic acid production is influenced by the type of organic nitrogen source. In this study, the influence of the type of nitrogen source becomes particularly obvious if the maximum productivity of l-lactic acid is compared at a concentration of 10 g·L−1 of each nutrient source. With 10 g·L−1 yeast extract, the maximum productivity was 3.02 g·L−1·h−1; with meat extract, it was 1.89 g·L−1·h−1, and it was significantly reduced with peptone at 0.93 g·L−1·h−1. In the literature, yeast extract is also described as the nutrition source with the highest titer and productivity [21,25,26,27]. However, the highest titer and productivity of l-lactic acid with L. casei ATCC 393 were achieved by a combination of 5 g·L−1 yeast extract and 5 g·L−1 meat extract in the present study. Additionally, the use of costly nitrogen sources was reduced by 60 % by a constant final titer >100 g·L−1 l-lactic acid and an average maximum productivity of 3.9 g·L−1·h−1. For that reason, a modified MRS medium with 5 g·L−1 yeast extract and 5 g·L−1 meat extract was used as a reference and analysis of nutrient requirements.
B-vitamins are crucial for the metabolism of Lactobacilli and have already been investigated with regard to growth, productivity, and final titer of l-lactic acid [28,29,30,31]. The described vitamin requirements in these studies differ for various L. casei strains and are not clear for the used strain ATCC 393. Rogosa et al. [28] and Koser [29] specified the vitamins B2, B3, B5, B6, B7, and B9 as essential, whereas Ledesma et al. [30] described B-vitamins as nonessential for L. casei. Therefore, a synthetic B-vitamin mix was designed based on B-vitamin composition of 5 g·L−1 yeast extract and 5 g·L−1 meat extract and was tested in limited MRS medium with 3 g·L−1 yeast extract in this study. The total B-vitamin mix influenced the l-lactic acid production positively, and the maximum productivity increased from 0.93 g·L−1·h−1 for cultivation with limited MRS medium to 1.15 g·L−1·h−1 with the total B-vitamin mix. Niacin (B3) had the greatest influence on cultivation, which became noticeable by the evaluation of every single vitamin, leading to a maximum productivity of 0.99 g·L−1·h−1, while all other single tested vitamins did not significantly improve the cultivation. Consequently, vitamin B3 is classified as essential for L. casei ATTC 393, and the whole synthetic B-vitamin mix was used for further analysis of amino acid requirements.
Furthermore, based on the amino acid composition of 5 g·L−1 yeast extract and 5 g·L−1 meat extract [18], a synthetic amino acid-mix was investigated. The final titer of 60.9 g·L−1 achieved with limited MRS medium, B-vitamin mix, and amino acid-mix is significantly lower compared to the cultivation with 5 g·L−1 yeast extract and 5 g·L−1 meat extract. This result suggested that essential amino acids were missing in the created amino acid-mix or that a limitation of amino acids occurred during fermentation. A limitation of tryptophan was proven by analyzing the free amino acids in the limited MRS medium with B-vitamin mix and amino acid-mix. Tryptophan, glutamic acid, arginine, cysteine, leucin, tyrosine, tryptophan, and valine are classified as essential amino acids [29,30,32]. Serine, isoleucine, and phenylalanine are described as essential or stimulatory. The used complex nutrient sources of 5 g·L−1 yeast extract and 5 g·L−1 meat extract contain all the listed amino acids, whereas cysteine and arginine could not be measured with the analytic method used. Therefore, a mixture of tryptophan, cysteine, and arginine was added to limited MRS medium with B-vitamin mix and amino acid-mix. With this combination, a comparable final titer of 95.2 g·L−1 l-lactic and yield of 0.89 (w/w) to the MRS medium with 5 g·L−1 yeast extract and 5 g·L−1 meat extract acid was achieved. However, the maximum productivity differed and the productivity of limited MRS medium with supplemented B-vitamin mix; amino acid-mix; and a mixture of Trp, Cys, and Arg was considerably lower than the productivity of the cultivation with 5 g·L−1 yeast extract and 5 g·L−1 meat extract. If each of the three amino acids were tested separately, the amino acid Cys shows major influence on cultivation. Besides total substrate consumption, the final titer of 94.1 g·L−1 gets close to the final titer of the reference cultivation with 5 g·L−1 yeast extract and 5 g·L−1 meat extract. Some species of L. casei cannot synthesize cysteine from methionine or serine, which is an essential amino acid and sulfur source for the growth of the bacteria [33,34]. Especially, a combination of cysteine and methionine stimulate the growth of some L. casei strains [33], which is probably the reason for the glucose consumption and the increased final titer. The results confirmed that these amino acids are essential, and special attention should be given to tryptophan and cysteine. To develop a synthetic medium for l-lactic acid production, the influence of trace elements and nucleosides should additionally be examined, this was was beyond the scope of the present study.
The major aim of this study, however, was the substitution of yeast extract by low-cost protein-rich agricultural hydrolysates for l-lactic acid production with L. casei. A chemical acidic hydrolysis with 3m H2SO4 at 110 °C proved to be a successful hydrolysis method for various protein-rich agricultural residues used in d-lactic acid production without deficits in yield, productivity, or final titer [16]. Hence, this method was transferred to the l-lactic acid production in this study. Also, enzymatic hydrolysates of rapeseed meal and distillers’ dried grains with solubles were tested for the cultivation of L. casei. The total nitrogen amount of hydrolysates used in the cultivations referred to the total nitrogen amount in 5 g·L−1 yeast extract and 5 g·L−1 meat extract. The single use of chemical and enzymatic hydrolysates in MRS medium without complex nutrient sources led to a halved final titer compared to using 5 g·L−1 yeast extract and 5 g·L−1 meat extract. Probably, the supply with essential B-vitamins, which was characterized in this study, is insufficient. Through the hydrolysis conditions, the heat labile B-vitamins of the agriculture residues are destroyed, and through acidic conditions, asparagine, glutamine, serine, and threonine are hydrolyzed whereas tryptophan and tyrosine are destroyed [35]. The use of the limited MRS medium with 3 g·L−1 yeast extract and either type of hydrolysate increased the final titer to 101.7 g·L−1 ± 2.7 g·L−1 l-lactic acid with a constant yield of 0.84 (w/w), which was comparable with the use of 5 g·L−1 yeast extract and 5 g·L−1 meat extract. Probably, 3 g·L−1 yeast extract provided essential B-vitamins and substituted, destroyed, or hydrolyzed amino acids, respectively, for growth and lactic-acid production in sufficient quantities. Also, in the literature, compared to pure yeast extract, combinations of agricultural residues and yeast extract yielded similar results for final titer and productivity of l-lactic acid production using L. casei strains [15,17]. A difference between the enzymatic and chemical hydrolysates is apparent because the maximum productivity decreased for the use of enzymatic hydrolysates by 35% compared to the chemical hydrolysate. The cultivations with the different agriculture residues DDGS and RM demonstrated similar yield, productivity, and final titer. Only the hydrolysis method had a minor impact. The influence of the hydrolysis method on cultivation behavior of Lactobacilli was already described by Brock et al. [16] and is attributable to a higher free amino acids content and lower peptide amount in chemical produced hydrolysates with 3m H2SO4.
All in all, the content of cost-intensive, complex media components was successfully reduced from 25 g·L−1 in total to between 10 g·L−1 to 3 g·L−1 yeast extract and was supplemented by agricultural residue hydrolysates with equal yield, productivity, and final titer of l-lactic acid. Yeast extract and other complex media components costs about 9000–10,000 € t−1 [1] and cover 25–38% of the total costs of biotechnological lactic acid production [11,12]. The costs of the tested protein-rich agricultural residues depend on the cost of corn or rapeseed meal and average between 120 and 220 € t−1 [36,37,38]. Raw material costs are also caused by the used sulfuric acid (128 € t−1) and neutralization agent Ca(OH)2 (100 € t−1) [39,40] as well as investment costs and annual cost of a plant. A hydrolysate based on the used agricultural residues with the same total nitrogen content as yeast extract costs approximately 1150 € t−1 using acidic hydrolysis in a hydrolysis reactor with a volume of 80 m3 and an annual operating time of 8300 h. If 70% of the yeast extract is replaced by a hydrolysate of DDGS or RM, the costs of l-lactic acid production with Lactobacillus casei could prospectively be reduced by 23%.
5. Conclusions
This study demonstrated that not the total nitrogen content of the cultivation media but rather the composition of the complex nutrition sources regarding B-vitamins and amino acids is significant for l-lactic acid production of L. casei. Moreover, 70% of expensive complex media components was supplemented by low-cost hydrolysates of agricultural residues, i.e., rapeseed meal (RM) and distillers’ dried grains with solubles (DDGS), with unchanged yield, productivity, and final titer of l-lactic acid. The reduced cost of approximately 23% of the biotechnological l-lactate production could make PLA production more economical, and the use of fossil-based thermoplastics could be significantly reduced. It has been demonstrated that the chemical hydrolysis with 3m H2SO4 is also suited for l-lactic acid as well as d-lactic acid production with different Lactobacilli species. If the new knowledge of nutritional requirements of L. casei is combined with the promising protein-rich hydrolysates, it is to be expected, that the content of yeast extract could be further reduced or totally supplemented by hydrolysates of agricultural residues.
Author Contributions
S.K. designed and performed the cultivation experiments. S.B. designed and performed the hydrolysis and analysis of the hydrolysates. S.K. wrote the manuscript. A.K. and U.P. supervised the experiments and revised the manuscript. A.K. performed the needed changes during the revision process. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the German Federal Ministry of Food and Agriculture, following a decision of the German Bundestag, via the Agency of Renewable Resources (grant No. 22016816).
Conflicts of Interest
The authors declare no conflict of interest. The funder had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Maddipati, P.; Atiyeh, H.K.; Bellmer, D.D.; Huhnke, R.L. Ethanol production from syngas by Clostridium strain P11 using corn steep liquor as a nutrient replacement to yeast extract. Bioresour. Technol. 2011, 102, 6494–6501. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, M.A.; Tashiro, Y.; Sonomoto, K. Recent advances in lactic acid production by microbial fermentation processes. Biotechnol. Adv. 2013, 31, 877–902. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tashiro, Y.; Sonomoto, K. Fermentative production of lactic acid from renewable materials: Recent achievements, prospects, and limits. J. Biosci. Bioeng. 2015, 119, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Chopin, A. Organization and regulation of genes for amino acid biosynthesis in lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 21–37. [Google Scholar] [CrossRef]
- Deguchi, Y.; Morishita, T. Nutritional requirements in multiple auxotrophic lactic acid bacteria: Genetic lesions affecting amino acid biosynthetic pathways in Lactococcus lactis, Enterococcus faecium, and Pediococcus acidilactici. Biosci. Biotechnol. Biochem. 1992, 56, 913–918. [Google Scholar] [CrossRef]
- Klotz, S.; Kaufmann, N.; Kuenz, A.; Prüße, U. Biotechnological production of enantiomerically pure d-lactic acid. Appl. Microbiol. Biotechnol. 2016, 100, 9423–9437. [Google Scholar] [CrossRef]
- Liu, S.-Q.; Holland, R.; Crow, V. The potential of dairy lactic acid bacteria to metabolise amino acids via non-transaminating reactions and endogenous transamination. Int. J. Food Microbiol. 2003, 86, 257–269. [Google Scholar] [CrossRef]
- de Oliveira, R.A.; Komesu, A.; Rossell, C.E.V.; Maciel Filho, R. Challenges and opportunities in lactic acid bioprocess design—From economic to production aspects. Biochem. Eng. J. 2018, 133, 219–239. [Google Scholar] [CrossRef]
- Garlotta, D. A literature review of poly (lactic acid). J. Polym. Environ. 2001, 9, 63–84. [Google Scholar] [CrossRef]
- Hofvendahl, K.; Hahn–Hägerdal, B. Factors affecting the fermentative lactic acid production from renewable resources1. Enzym. Microb. Technol. 2000, 26, 87–107. [Google Scholar] [CrossRef]
- González, M.I.; Álvarez, S.; Riera, F.; Álvarez, R. Economic evaluation of an integrated process for lactic acid production from ultrafiltered whey. J. Food Eng. 2007, 80, 553–561. [Google Scholar] [CrossRef]
- Tejayadi, S.; Cheryan, M. Lactic acid from cheese whey permeate. Productivity and economics of a continuous membrane bioreactor. Appl. Microbiol. Biotechnol. 1995, 43, 242–248. [Google Scholar] [CrossRef]
- Sikder, J.; Roy, M.; Dey, P.; Pal, P. Techno-economic analysis of a membrane-integrated bioreactor system for production of lactic acid from sugarcane juice. Biochem. Eng. J. 2012, 63, 81–87. [Google Scholar] [CrossRef]
- Sommer, R. Yeast extracts: Production, properties and components. Food Aust. 1998, 50, 181–183. [Google Scholar]
- Pauli, T.; Fitzpatrick, J.J. Malt combing nuts as a nutrient supplement to whey permeate for producing lactic by fermentation with Lactobacillus casei. Process Biochem. 2002, 38, 1–6. [Google Scholar] [CrossRef]
- Brock, S.; Kuenz, A.; Prüße, U. Impact of hydrolysis methods on the utilization of agricultural residues as nutrient source for D-lactic acid production by Sporolactobacillus inulinus. Fermentation 2019, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Hujanen, M.; Linko, S.; Linko, Y.-Y.; Leisola, M. Optimisation of media and cultivation conditions for L (+)(S)-lactic acid production by Lactobacillus casei NRRL B-441. Appl. Microbiol. Biotechnol. 2001, 56, 126–130. [Google Scholar] [CrossRef]
- Klotz, S.; Kuenz, A.; Prüße, U. Nutritional requirements and the impact of yeast extract on the d-lactic acid production by Sporolactobacillus inulinus. Green Chem. 2017, 19, 4633–4641. [Google Scholar] [CrossRef]
- De Man, J.C.; Rogosa, M.; Sharpe, M.E. A Medium for the Cultivation of Lactobacilli. J. Appl. Bacteriol. 1960, 23, 130–135. [Google Scholar] [CrossRef]
- Olmos-Dichara, A.; Ampe, F.; Uribelarrea, J.L.; Pareilleux, A.; Goma, G. Growth and lactic acid production by Lactobacillus casei ssp. rhamnosus in batch and membrane bioreactor: Influence of yeast extract and Tryptone enrichment. Biotechnol. Lett. 1997, 19, 709–714. [Google Scholar] [CrossRef]
- Amrane, A.; Prigent, Y. Influence of yeast extract concentrationon batch cultures of Lactobacillus helveticus: Growth and production coupling. World J. Microbiol. Biotechnol. 1998, 14, 529–534. [Google Scholar] [CrossRef]
- Nancib, N.; Nancib, A.; Boudjelal, A.; Benslimane, C.; Blanchard, F.; Boudrant, J. The effect of supplementation by different nitrogen sources on the production of lactic acid from date juice by Lactobacillus casei subsp. rhamnosus. Bioresour. Technol. 2001, 78, 149–153. [Google Scholar] [CrossRef]
- Ding, S.; Tan, T. L-lactic acid production by Lactobacillus casei fermentation using different fed-batch feeding strategies. Process Biochem. 2006, 41, 1451–1454. [Google Scholar] [CrossRef]
- Hujanen, M.; Linko, Y.Y. Effect of temperature and various nitrogen sources on L(+)-lactic acid production by Lactobacillus casei. Appl. Microbiol. Biotechnol. 1996, 45, 307–313. [Google Scholar] [CrossRef]
- Kwon, S.; Lee, P.C.; Lee, E.G.; Keun Chang, Y.; Chang, N. Production of lactic acid by Lactobacillus rhamnosus with vitamin-supplemented soybean hydrolysate. Enzym. Microb. Technol. 2000, 26, 209–215. [Google Scholar] [CrossRef]
- Aeschlimann, A.; von Stockar, U. The effect of yeast extract supplementation on the production of lactic acid from whey permeate by Lactobacillus helueticus. Appl. Microbiol. Biotechnol. 1990, 32, 398–402. [Google Scholar] [CrossRef]
- Aspmo, S.I.; Horn, S.J.; Eijsink, V.G.H. Use of hydrolysates from Atlantic cod (Gadus morhua L.) viscera as a complex nitrogen source for lactic acid bacteria. FEMS Microbiol. Lett. 2005, 248, 65–68. [Google Scholar] [CrossRef]
- Rogosa, M.; Franklin, J.G.; Perry, K.D. Correlation of the vitamin requirements with cultural and biochemical characters of Lactobacillus spp. J. Gen. Microbiol. 1961, 25, 473–482. [Google Scholar] [CrossRef] [Green Version]
- Koser, S.A. Vitamin requirements of bacteria and yeasts. Arch. Intern. Med. 1968, 122, 192. [Google Scholar] [CrossRef]
- Ledesma, O.V.; De Ruiz Holgado, A.P.; Oliver, G.; De Giori, G.S.; Raibaud, P.; Galpin, J.V. A synthetic medium for comparative nutritional studies of lactobacilli. J. Appl. Bacteriol. 1977, 42, 123–133. [Google Scholar] [CrossRef]
- Yoo, I.-K.; Chang, H.N.; Lee, E.G.; Chang, Y.K.; Moon, S.-H. Effect of B vitamin supplementation on lactic acid production by Lactobacillus casei. J. Ferment Bioeng. 1997, 84, 172–175. [Google Scholar] [CrossRef]
- Dunn, M.S.; Shankman, S.; Camien, M.N.; Block, H. The amino acid requirements of twenty-three lactic acid bacteria. J. Biol. Chem. 1947, 168, 1–22. [Google Scholar] [PubMed]
- Bogicevic, B.; Berthoud, H.; Portmann, R.; Bavan, T.; Meile, L.; Irmler, S. Cysteine biosynthesis in Lactobacillus casei: Identification and characterization of a serine acetyltransferase. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef] [Green Version]
- Irmler, S.; Schafer, H.; Beisert, B.; Rauhut, D.; Berthoud, H. Identification and characterization of a strain-dependent cystathionine beta/gamma-lyase in Lactobacillus casei potentially involved in cysteine biosynthesis. FEMS Microbiol. Lett. 2009, 295, 67–76. [Google Scholar] [CrossRef]
- Fountoulakis, M.; Lahm, H.-W. Hydrolysis and amino acid composition analysis of proteins. J. Chromatogr. A 1998, 826, 109–134. [Google Scholar] [CrossRef]
- Stein, H.H. Distillers dried grains with solubles (DDGS) in diets fed to swine. Swine Focus 2007, 1, 1–8. [Google Scholar]
- Proplanta. Hamburg Großhandelspreise vom 18.08.2020-Getreide, Mühlenprodukte, Futtermittel und Ölsaaten. Available online: https://www.proplanta.de/markt-und-preis/hamburger-getreideboerse/hamburg-grosshandelspreise-vom-18-08-2020-getreide-muehlenprodukte-futtermittel-und-oelsaaten_notierungen1597770118.html (accessed on 23 August 2020).
- DTN. Progressive Farmer, DDG Weekly Update-DTN Weekly Average DDG Price Unchanged 2/24/2020. Available online: https://www.dtnpf.com/agriculture/web/ag/livestock/article/2020/02/24/dtn-weekly-average-ddg-price (accessed on 23 August 2020).
- DKL Engineering, I. Sulphuric acid on the webᵀᴹ-Market and cost information. Available online: www.sulphuric-acid.com/sulphuric-acid-on-the-web/Market-Info.htm (accessed on 23 August 2020).
- Schnippe, F. Löschkalk als preisgünstige Alternative. Top Agrar 2004, 6, 6–9. [Google Scholar]
Figure 1.
Influence of different B-vitamins on l-lactate production with L. casei in limited MRS medium with 3 g·L−1 yeast extract in 24-well plates at 37 °C and 400 rpm. Black, closed squares modified MRS medium with 5 g·L−1 yeast extract (YE) and 5 g·L−1 meat extract (ME), red, opened squares limited MRS medium with 3 g·L−1 yeast extract.
Figure 1.
Influence of different B-vitamins on l-lactate production with L. casei in limited MRS medium with 3 g·L−1 yeast extract in 24-well plates at 37 °C and 400 rpm. Black, closed squares modified MRS medium with 5 g·L−1 yeast extract (YE) and 5 g·L−1 meat extract (ME), red, opened squares limited MRS medium with 3 g·L−1 yeast extract.

Figure 2.
Influence of different medium composition on l-lactate production with L. casei in 24-well plates at 37 °C and 400 rpm: black, closed squares modified MRS medium with 5 g·L−1 yeast extract (YE) and 5 g·L−1 meat extract (ME); red, opened squares limited MRS medium with 3 g·L−1 yeast extract.
Figure 2.
Influence of different medium composition on l-lactate production with L. casei in 24-well plates at 37 °C and 400 rpm: black, closed squares modified MRS medium with 5 g·L−1 yeast extract (YE) and 5 g·L−1 meat extract (ME); red, opened squares limited MRS medium with 3 g·L−1 yeast extract.

Figure 3.
Use of different agricultural residue hydrolysates instead of complex medium components in modified MRS medium with 5 g·L−1 yeast extract (YE) and 5 g·L−1 meat extract (ME) and limited MRS medium with 3 g·L−1 yeast extract for l-lactate production with L. casei in 24-well plates at 37 °C and 400 rpm: the total nitrogen amount was kept constant comparable to 5 g·L−1 yeast extract and 5 g·L−1 meat extract. Black, closed squares modified MRS medium with 5 g·L−1 yeast extract and 5 g·L−1 meat extract; red, opened squares limited MRS medium with 3 g·L−1 yeast extract.
Figure 3.
Use of different agricultural residue hydrolysates instead of complex medium components in modified MRS medium with 5 g·L−1 yeast extract (YE) and 5 g·L−1 meat extract (ME) and limited MRS medium with 3 g·L−1 yeast extract for l-lactate production with L. casei in 24-well plates at 37 °C and 400 rpm: the total nitrogen amount was kept constant comparable to 5 g·L−1 yeast extract and 5 g·L−1 meat extract. Black, closed squares modified MRS medium with 5 g·L−1 yeast extract and 5 g·L−1 meat extract; red, opened squares limited MRS medium with 3 g·L−1 yeast extract.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Reduction of the complex components yeast extract (YE), peptone (P), and meat extract (ME) in MRS medium and effect on final titer after 70.5 h and maximum productivity of l-lactic acid: 37 °C, 400 rpm, and 24-well plates. TN was calculated based on results of Klotz et al. [18].
Table 1.
Reduction of the complex components yeast extract (YE), peptone (P), and meat extract (ME) in MRS medium and effect on final titer after 70.5 h and maximum productivity of l-lactic acid: 37 °C, 400 rpm, and 24-well plates. TN was calculated based on results of Klotz et al. [18].
| YE (g·L−1) | P (g·L−1) | ME (g·L−1) | TN (g·L−1) | Glucose (g·L−1) | l-Lactic Acid (g·L−1) | Max. Productivity (g·L−1 h−1) |
|---|---|---|---|---|---|---|
| 5 | 10 | 10 | 2.97 | 0 | 106.3 ± 5.5 | 4.63 ± 0.06 |
| 5 | 0 | 5 | 1.11 | 0 | 113.3 ± 3.7 | 4.76 ± 0.08 |
| 5 | 5 | 5 | 1.72 | 0 | 105.1 ± 0.5 | 4.39 ± 0.22 |
| 10 | 0 | 0 | 0.95 | 0 | 113.3 ± 1.1 | 3.02 ± 0.08 |
| 5 | 5 | 0 | 1.09 | 0 | 108.1 ± 2.9 | 2.67 ± 0.26 |
| 0 | 0 | 10 | 1.25 | 0 | 102.1 ± 1.1 | 1.89 ± 0.01 |
| 5 | 0 | 0 | 0.48 | 33.5 ± 0.9 | 87.2 ± 0.5 | 1.69 ± 0.04 |
| 0 | 5 | 5 | 1.24 | 38.7 ± 0.1 | 64.36 ± 0.5 | 1.62 ± 0.02 |
| 0 | 0 | 5 | 0.63 | 54.8 ± 6.1 | 59.0 ± 4.8 | 1.62 ± 0.38 |
| 3 | 0 | 0 | 0.29 | 65.4 ± 0.6 | 57.9 ± 0.1 | 1.02 ± 0.01 |
| 0 | 10 | 0 | 1.24 | 75.7 ± 2.8 | 33.8 ± 1.7 | 0.93 ±0.01 |
| 0 | 5 | 0 | 0.62 | 78.8 ± 4.3 | 18.7 ± 0.8 | 0.54 ± 0.01 |
| 0 | 0 | 0 | 0 | 114 ± 0.1 | 6.1 ± 0.1 | 0.21 ± 0.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Krull, S.; Brock, S.; Prüße, U.; Kuenz, A. Hydrolyzed Agricultural Residues—Low-Cost Nutrient Sources for l-Lactic Acid Production. Fermentation 2020, 6, 97. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040097
AMA Style
Krull S, Brock S, Prüße U, Kuenz A. Hydrolyzed Agricultural Residues—Low-Cost Nutrient Sources for l-Lactic Acid Production. Fermentation. 2020; 6(4):97. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040097
Chicago/Turabian StyleKrull, Susan, Silvia Brock, Ulf Prüße, and Anja Kuenz. 2020. "Hydrolyzed Agricultural Residues—Low-Cost Nutrient Sources for l-Lactic Acid Production" Fermentation 6, no. 4: 97. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation6040097
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.