Dynamic Co-Cultivation Process of Corynebacterium glutamicum Strains for the Fermentative Production of Riboflavin

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids and Growth Conditions
2.2. Molecular Genetic Techniques and Strains Construction
2.3. Analytical Procedures
2.4. Bioreactor Conditions
3. Results
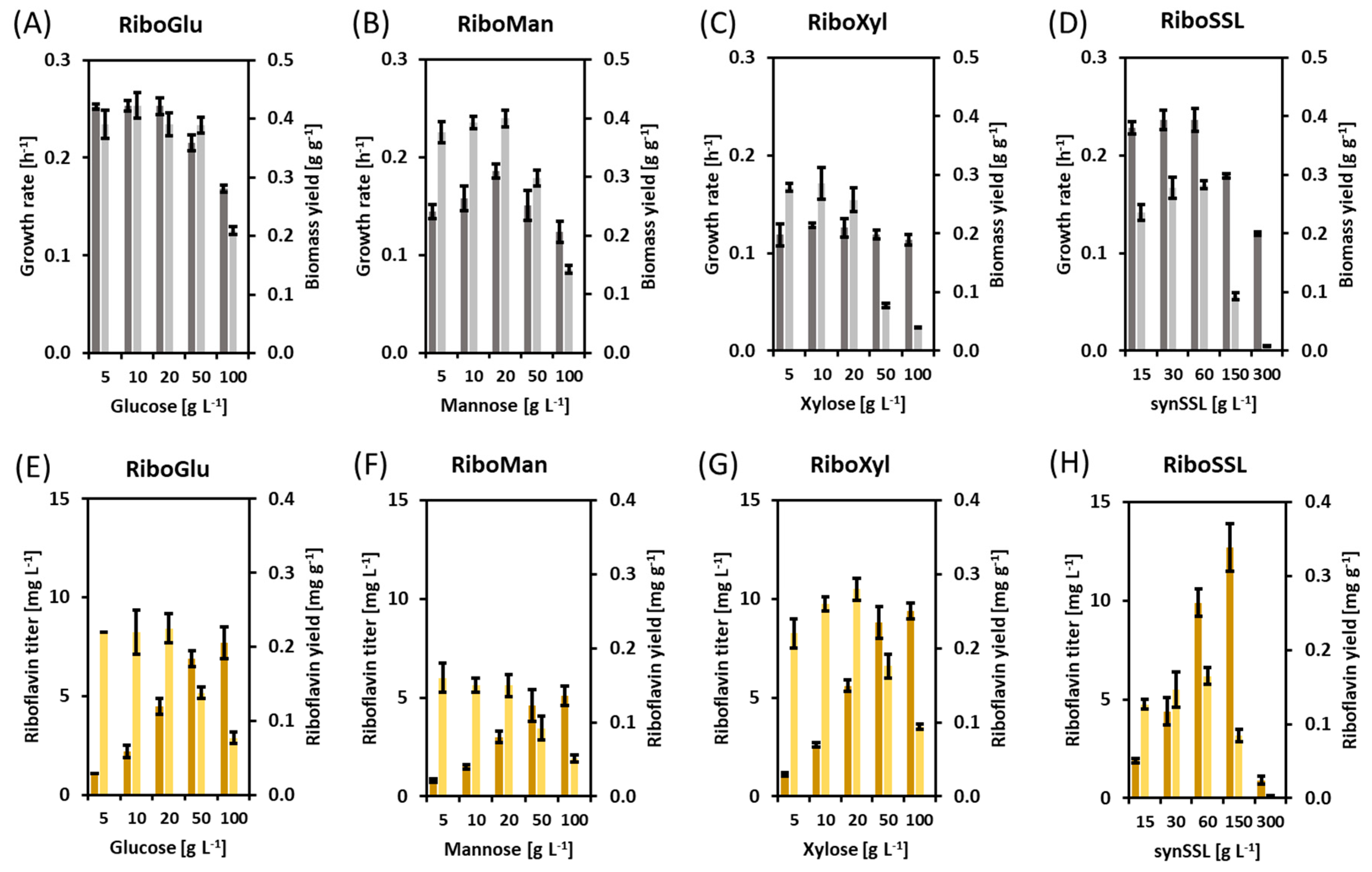
3.1. Riboflavin Production in Flask Fermentations with Different Carbon Sources and Concentrations
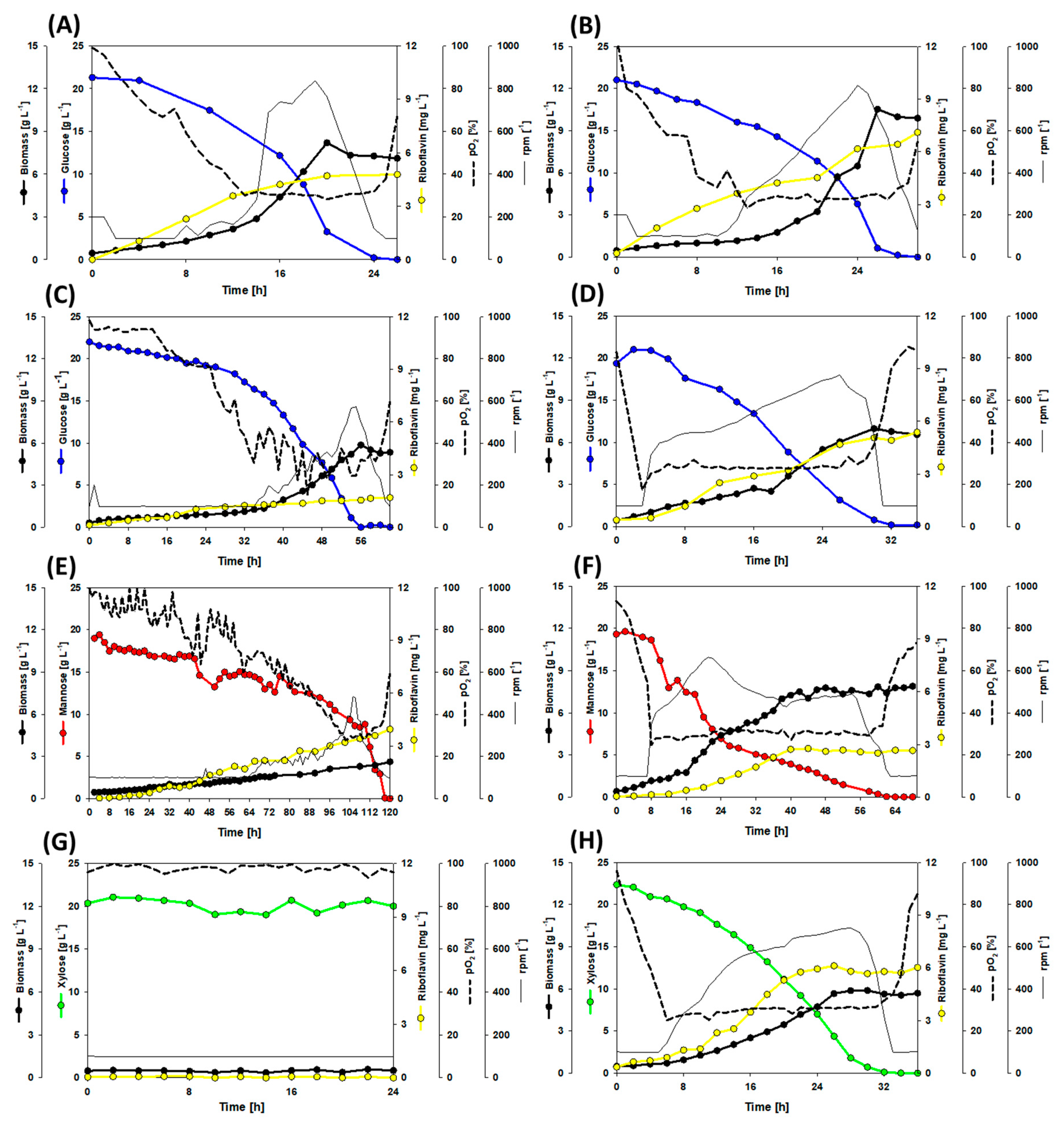
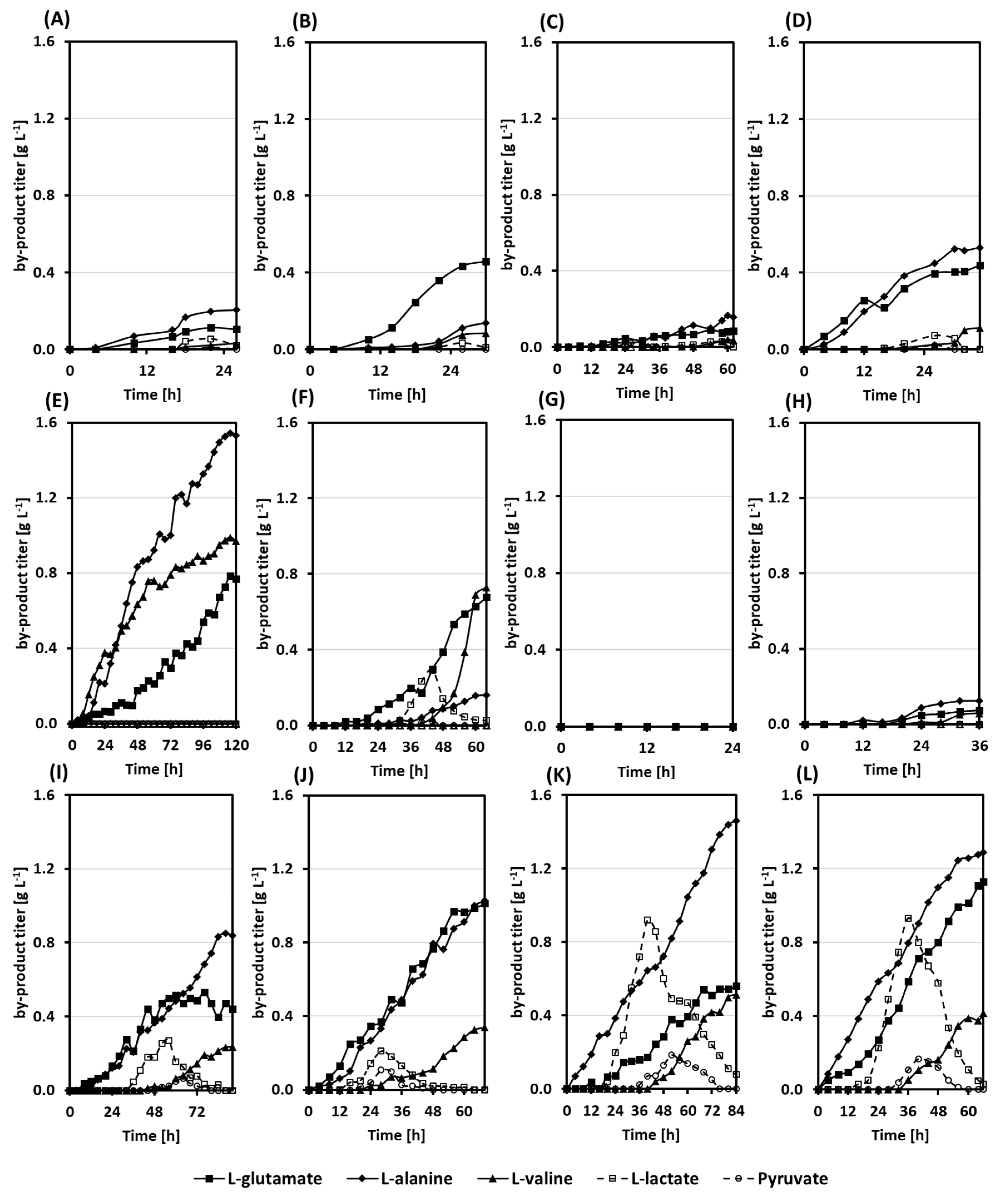
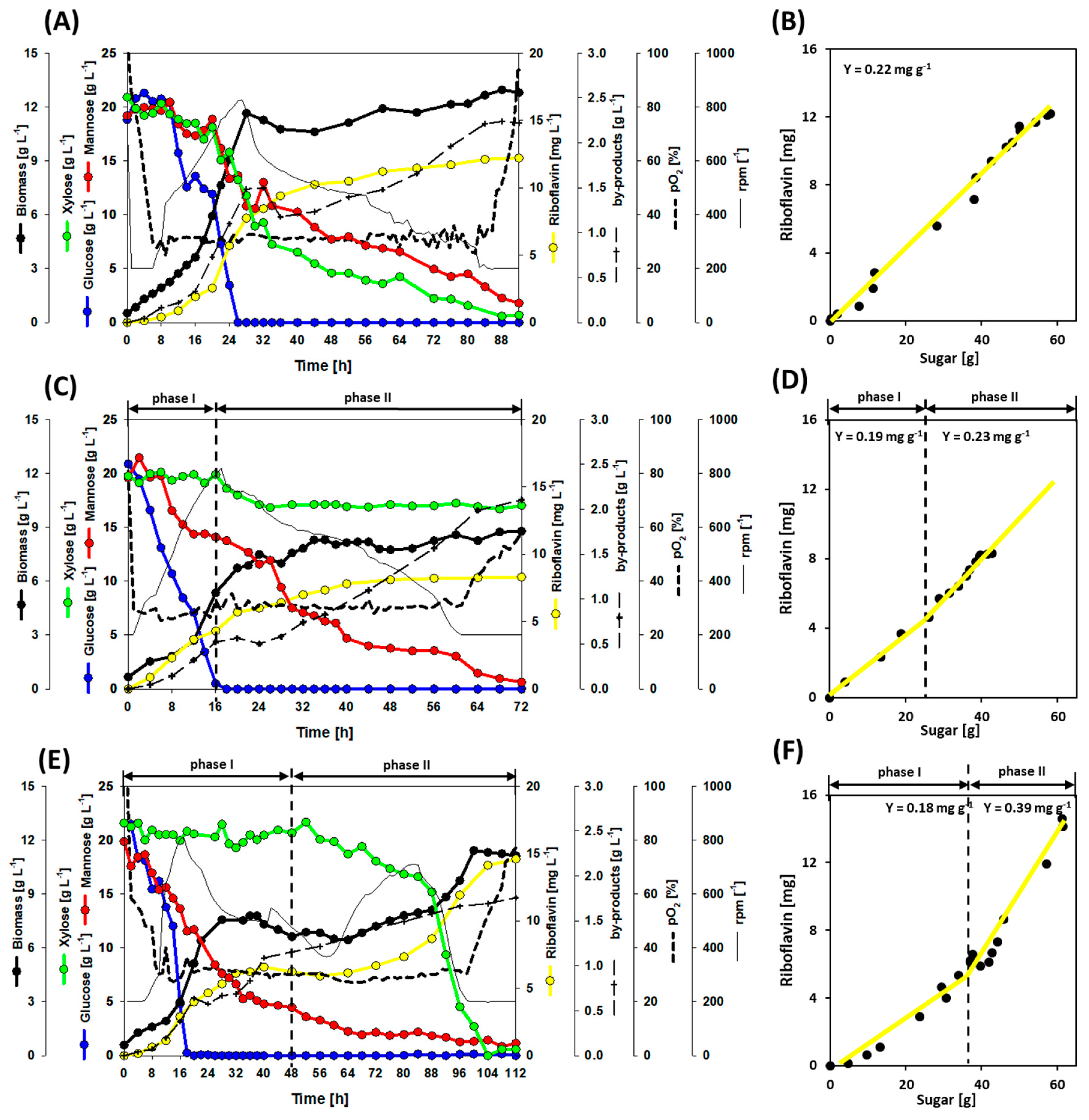
3.2. Growth and Production Behavior of the Riboflavin Producing Strains in Lab-Scale Bioreactor Fermentations
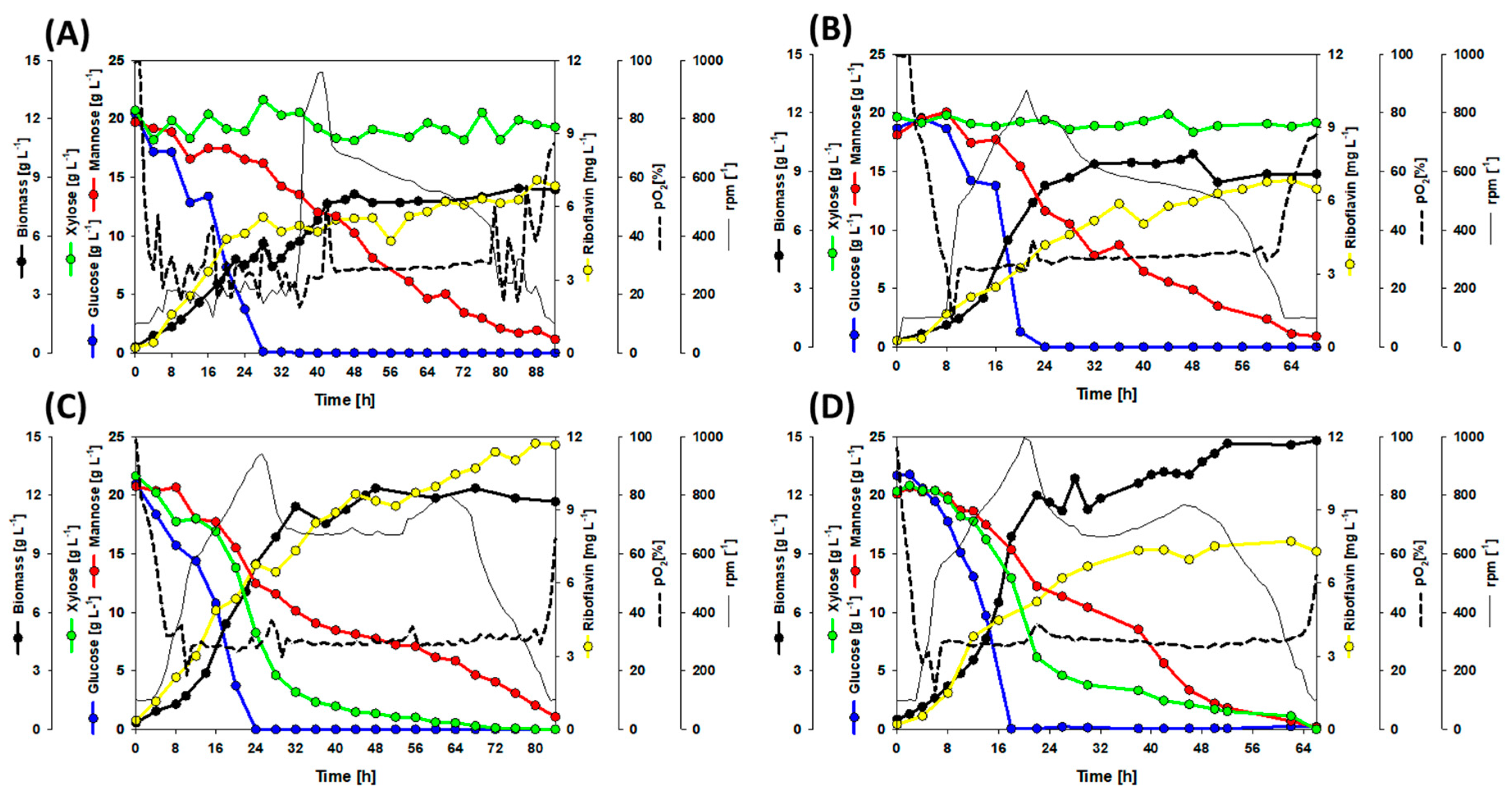
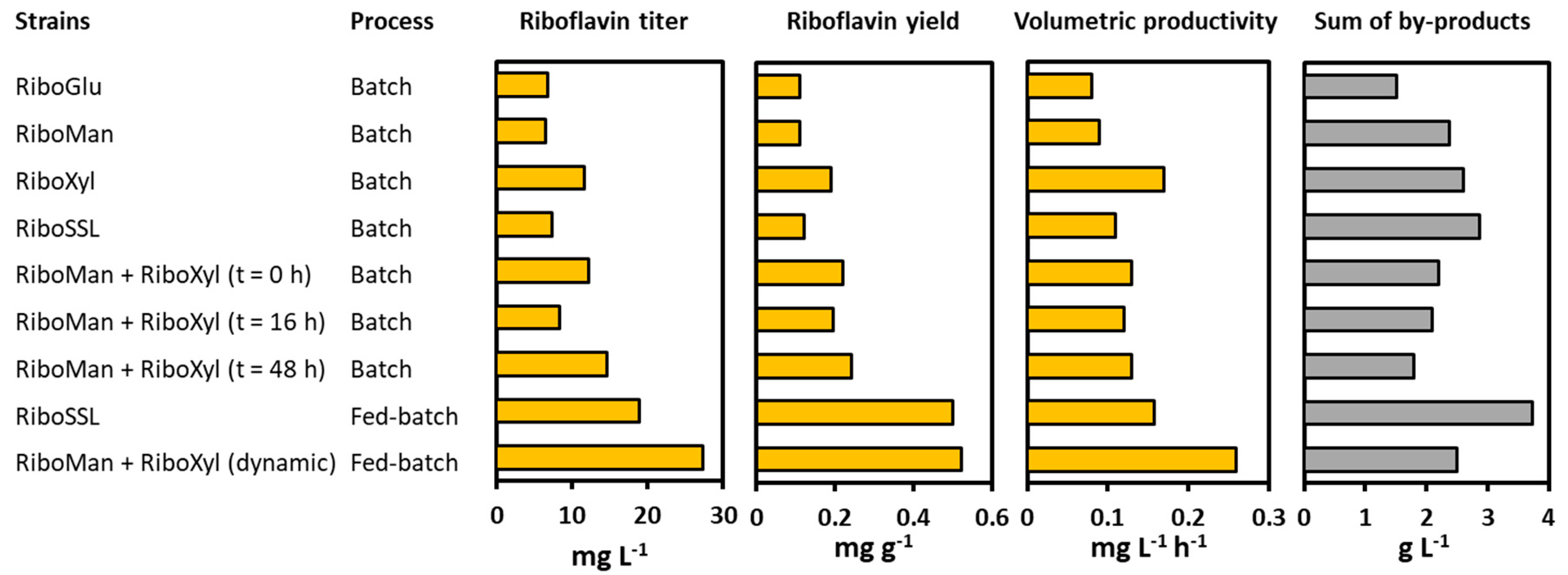
3.3. A Co-Cultivation Strategy of C. glutamicum Strains Improved Fermentation Performance
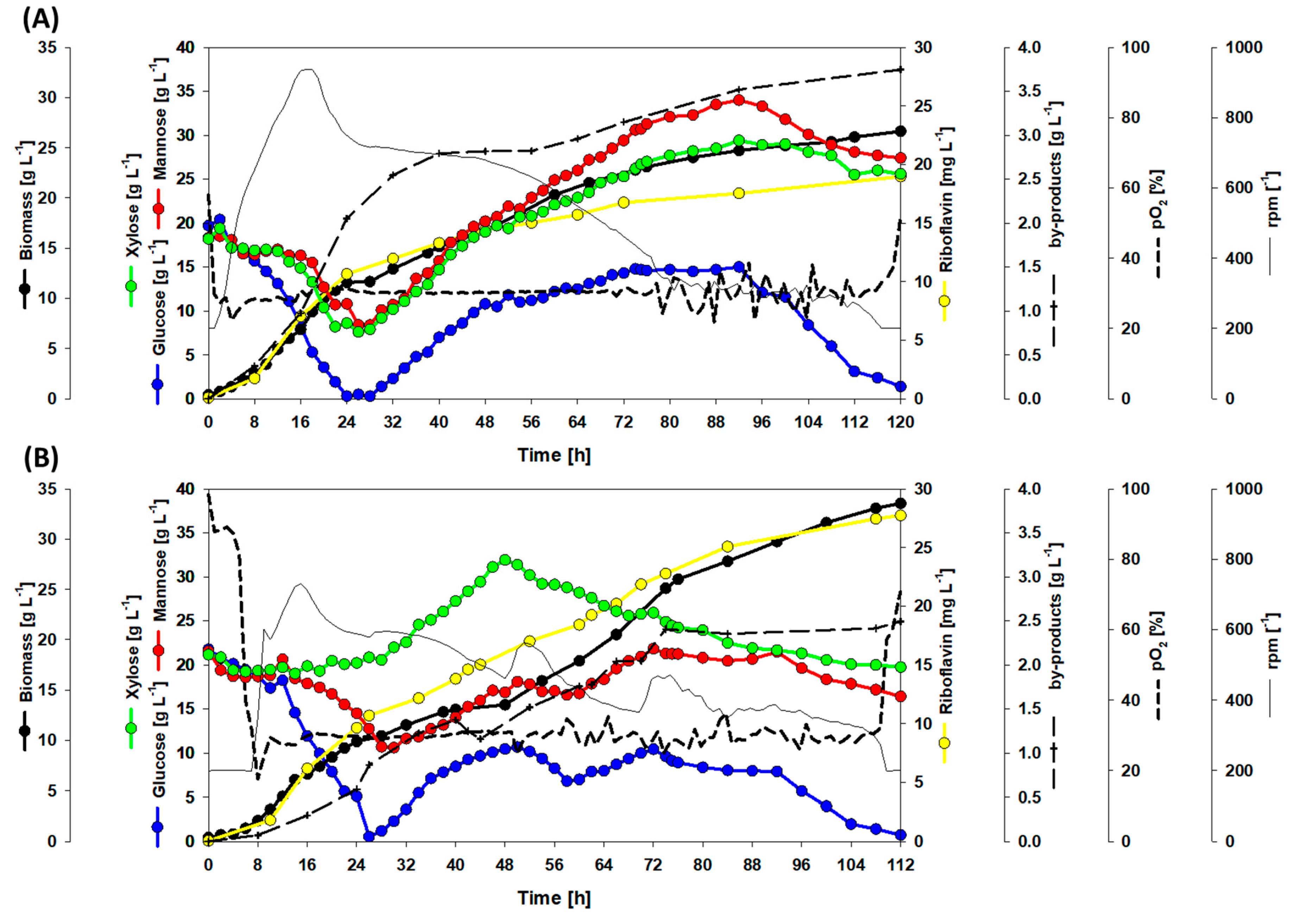
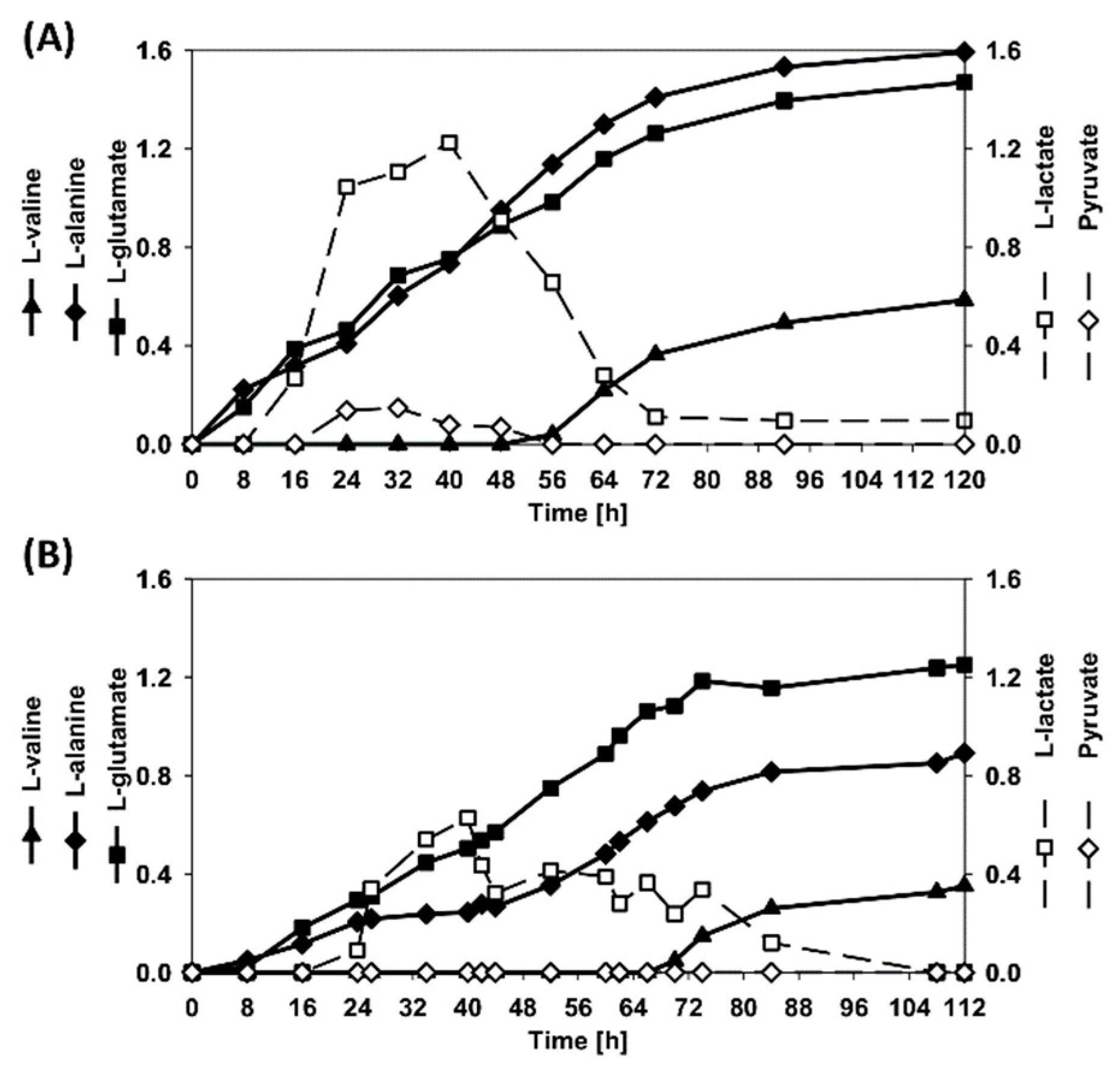
3.4. Dynamic Co-Inoculation Fed-Batch Process Enhanced Productivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fahmy, Y.; Fahmy, T.Y.A.; Mobarak, F.; El-Sakhawy, M.; Fadl, M. Agricultural Residues (Wastes) for Manufacture of Paper, Board, and Miscellaneous Products: Background Overview and Future Prospects; Social Science Research Network: Rochester, NY, USA, 2017. [Google Scholar]
- Bajpai, P. Brief description of the pulp and papermaking process. In Biotechnology for Pulp and Paper Processing; Bajpai, P., Ed.; Springer: Singapore, 2018; pp. 9–26. ISBN 978-981-10-7853-8. [Google Scholar]
- Jiménez, L.; Pérez, I.; García, J.C.; Rodríguez, A.; Ferrer, J.L. Influence of ethanol pulping of wheat straw on the resulting paper sheets. Process Biochem. 2002, 37, 665–672. [Google Scholar] [CrossRef]
- Rodríguez, A.; Espinosa, E.; Domínguez-Robles, J.; Sánchez, R.; Bascón, I.; Rosal, A. Different solvents for organosolv pulping. In Pulp and Paper Processing; Books on Demand: Norderstedt, Germany, 2018. [Google Scholar] [CrossRef] [Green Version]
- Aro, T.; Fatehi, P. Production and application of lignosulfonates and sulfonated lignin. ChemSusChem 2017, 10, 1861–1877. [Google Scholar] [CrossRef] [PubMed]
- Petersen, A.M.; Haigh, K.; Görgens, J.F. Techno-economics of integrating bioethanol production from spent sulfite liquor for reduction of greenhouse gas emissions from sulfite pulping mills. Biotechnol. Biofuels 2014, 7, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueda, C.; Calvo, P.A.; Moncalián, G.; Ruiz, G.; Coz, A. Biorefinery options to valorize the spent liquor from sulfite pulping. J. Chem. Technol. Biotechnol. 2015, 90, 2218–2226. [Google Scholar] [CrossRef]
- Rødsrud, G.; Lersch, M.; Sjöde, A. History and future of world’s most advanced biorefinery in operation. Biomass Bioenergy 2012, 46, 46–59. [Google Scholar] [CrossRef]
- Humpert, D.; Ebrahimi, M.; Czermak, P. Membrane technology for the recovery of lignin: A review. Membranes 2016, 6, 42. [Google Scholar] [CrossRef]
- Lawford, H.G.; Rousseau, J.D. Production of ethanol from pulp mill hardwood and softwood spent sulfite liquors by genetically engineered E. coli. Appl. Biochem. Biotechnol. 1993, 39–40, 667–685. [Google Scholar] [CrossRef]
- Novy, V.; Krahulec, S.; Longus, K.; Klimacek, M.; Nidetzky, B. Co-fermentation of hexose and pentose sugars in a spent sulfite liquor matrix with genetically modified Saccharomyces cerevisiae. Bioresour. Technol. 2013, 130, 439–448. [Google Scholar] [CrossRef]
- Alexandri, M.; Venus, J. Feedstock flexibility in sustainable chemistry: Bridging sectors still not sufficiently familiar with each other—Sshowcases of ongoing and emerging initiatives. Curr. Opin. Green Sustain. Chem. 2017, 8, 24–29. [Google Scholar] [CrossRef]
- Bates, C.J. Riboflavin. In Encyclopedia of Human Nutrition, 3rd ed.; Caballero, B., Ed.; Academic Press: Waltham, MA, USA, 2013; pp. 158–165. ISBN 978-0-12-384885-7. [Google Scholar]
- Pinto, J.T.; Zempleni, J. Riboflavin. Adv. Nutr. 2016, 7, 973–975. [Google Scholar] [CrossRef] [Green Version]
- Revuelta, J.L.; Ledesma-Amaro, R.; Lozano-Martinez, P.; Díaz-Fernández, D.; Buey, R.M.; Jiménez, A. Bioproduction of riboflavin: A bright yellow history. J. Ind. Microbiol. Biotechnol. 2017, 44, 659–665. [Google Scholar] [CrossRef]
- Ledesma-Amaro, R.; Serrano-Amatriain, C.; Jiménez, A.; Revuelta, J.L. Metabolic engineering of riboflavin production in Ashbya gossypii through pathway optimization. Microb. Cell Factories 2015, 14, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Hu, W.; Wang, Z.; Chen, T. Production of riboflavin and related cofactors by biotechnological processes. Microb. Cell Factories 2020, 19, 31. [Google Scholar] [CrossRef] [PubMed]
- Wendisch, V.F. Microbial production of amino acid-related compounds. Adv. Biochem. Eng. Biotechnol. 2017, 159, 255–269. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.; Rohles, C.M.; Wittmann, C. Metabolically engineered Corynebacterium glutamicum for bio-based production of chemicals, fuels, materials, and healthcare products. Metab. Eng. 2018, 50, 122–141. [Google Scholar] [CrossRef]
- Wendisch, V.F. Metabolic engineering advances and prospects for amino acid production. Metab. Eng. 2020, 58, 17–34. [Google Scholar] [CrossRef]
- Kogure, T.; Inui, M. Recent advances in metabolic engineering of Corynebacterium glutamicum for bioproduction of value-added aromatic chemicals and natural products. Appl. Microbiol. Biotechnol. 2018, 102, 8685–8705. [Google Scholar] [CrossRef]
- Pérez-García, F.; Wendisch, V.F. Transport and metabolic engineering of the cell factory Corynebacterium glutamicum. FEMS Microbiol. Lett. 2018, 365, fny166. [Google Scholar] [CrossRef] [Green Version]
- Meiswinkel, T.M.; Gopinath, V.; Lindner, S.N.; Nampoothiri, K.M.; Wendisch, V.F. Accelerated pentose utilization by Corynebacterium glutamicum for accelerated production of lysine, glutamate, ornithine and putrescine. Microb. Biotechnol. 2013, 6, 131–140. [Google Scholar] [CrossRef]
- Wendisch, V.F.; Brito, L.F.; Gil Lopez, M.; Hennig, G.; Pfeifenschneider, J.; Sgobba, E.; Veldmann, K.H. The flexible feedstock concept in industrial biotechnology: Metabolic engineering of Escherichia coli, Corynebacterium glutamicum, Pseudomonas, Bacillus and yeast strains for access to alternative carbon sources. J. Biotechnol. 2016, 234, 139–157. [Google Scholar] [CrossRef]
- Sgobba, E.; Wendisch, V.F. Synthetic microbial consortia for small molecule production. Curr. Opin. Biotechnol. 2019, 62, 72–79. [Google Scholar] [CrossRef]
- Burmeister, A.; Hilgers, F.; Langner, A.; Westerwalbesloh, C.; Kerkhoff, Y.; Tenhaef, N.; Drepper, T.; Kohlheyer, D.; von Lieres, E.; Noack, S.; et al. A microfluidic co-cultivation platform to investigate microbial interactions at defined microenvironments. Lab Chip 2018, 19, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, H.; Wendisch, V.F. Exploring the role of sigma factor gene expression on production by Corynebacterium glutamicum: Sigma factor H and FMN as example. Front. Microbiol. 2015, 6, 740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin, Y.-W.; Park, J.-B.; Park, Y.-C.; Kim, K.H.; Seo, J.-H. Metabolic engineering of Corynebacterium glutamicum to produce GDP-L-fucose from glucose and mannose. Bioprocess Biosyst. Eng. 2013, 36, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Abe, S.; Takayama, K.-I.; Kinoshita, S. Taxonomical studies on glutamic acid-producing bacteria. J. Gen. Appl. Microbiol. 1967, 13, 279–301. [Google Scholar] [CrossRef]
- Henke, N.A.; Heider, S.A.E.; Peters-Wendisch, P.; Wendisch, V.F. Production of the marine carotenoid astaxanthin by metabolically engineered Corynebacterium glutamicum. Mar. Drugs 2016, 14, 124. [Google Scholar] [CrossRef]
- Stansen, C.; Uy, D.; Delaunay, S.; Eggeling, L.; Goergen, J.-L.; Wendisch, V.F. Characterization of a Corynebacterium glutamicum lactate utilization operon induced during temperature-triggered glutamate production. Appl. Environ. Microbiol. 2005, 71, 5920–5928. [Google Scholar] [CrossRef] [Green Version]
- Eggeling, L.; Bott, M.; Bott, M. Handbook of Corynebacterium Glutamicum; CRC Press: Boca Raton, FL, USA, 2005; ISBN 978-0-429-12905-6. [Google Scholar]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012; ISBN 978-1-936113-41-5. [Google Scholar]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Sasaki, M.; Teramoto, H.; Inui, M.; Yukawa, H. Identification of mannose uptake and catabolism genes in Corynebacterium glutamicum and genetic engineering for simultaneous utilization of mannose and glucose. Appl. Microbiol. Biotechnol. 2011, 89, 1905–1916. [Google Scholar] [CrossRef]
- Kiefer, P.; Heinzle, E.; Zelder, O.; Wittmann, C. Comparative metabolic flux analysis of lysine-producing Corynebacterium glutamicum cultured on glucose or fructose. Appl. Environ. Microbiol. 2004, 70, 229–239. [Google Scholar] [CrossRef] [Green Version]
- Paczia, N.; Nilgen, A.; Lehmann, T.; Gätgens, J.; Wiechert, W.; Noack, S. Extensive exometabolome analysis reveals extended overflow metabolism in various microorganisms. Microb. Cell Factories 2012, 11, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-García, F.; Peters-Wendisch, P.; Wendisch, V.F. Engineering Corynebacterium glutamicum for fast production of L-lysine and L-pipecolic acid. Appl. Microbiol. Biotechnol. 2016, 100, 8075–8090. [Google Scholar] [CrossRef] [PubMed]
- Wendisch, V.F.; Lindner, S.N.; Meiswinkel, T.M. Use of glycerol in biotechnological applications. Biodiesel—Quality, Emissions and By-Products; InTechOpen: London, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Khanna, S.; Goyal, A.; Moholkar, V.S. Microbial conversion of glycerol: Present status and future prospects. Crit. Rev. Biotechnol. 2012, 32, 235–262. [Google Scholar] [CrossRef] [PubMed]
- Holmberg, C.; Beijer, L.; Rutberg, B.; Rutberg, L. Glycerol catabolism in Bacillus subtilis: Nucleotide sequence of the genes encoding glycerol kinase (glpK) and glycerol-3-phosphate dehydrogenase (glpD). J. Gen. Microbiol. 1990, 136, 2367–2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Ren, S.; Ye, X.P. Lactic acid production from glycerol using CaO as solid base catalyst. Fuel Process. Technol. 2014, 120, 40–47. [Google Scholar] [CrossRef]
- Matano, C.; Uhde, A.; Youn, J.-W.; Maeda, T.; Clermont, L.; Marin, K.; Krämer, R.; Wendisch, V.F.; Seibold, G.M. Engineering of Corynebacterium glutamicum for growth and L-lysine and lycopene production from N-acetyl-glucosamine. Appl. Microbiol. Biotechnol. 2014, 98, 5633–5643. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-K.; Shen, C.-R.; Liu, C.-L. N-acetylglucosamine: Production and applications. Mar. Drugs 2010, 8, 2493–2516. [Google Scholar] [CrossRef] [Green Version]
- Bertram, R.; Rigali, S.; Wood, N.; Lulko, A.T.; Kuipers, O.P.; Titgemeyer, F. Regulon of the N-acetylglucosamine utilization regulator NagR in Bacillus subtilis. J. Bacteriol. 2011, 193, 3525–3536. [Google Scholar] [CrossRef] [Green Version]
- Marx, A.; Hans, S.; Möckel, B.; Bathe, B.; de Graaf, A.A.; McCormack, A.C.; Stapleton, C.; Burke, K.; O’Donohue, M.; Dunican, L.K. Metabolic phenotype of phosphoglucose isomerase mutants of Corynebacterium glutamicum. J. Biotechnol. 2003, 104, 185–197. [Google Scholar] [CrossRef]
- Brüsseler, C.; Späth, A.; Sokolowsky, S.; Marienhagen, J. Alone at last!—Heterologous expression of a single gene is sufficient for establishing the five-step Weimberg pathway in Corynebacterium glutamicum. Metab. Eng. Commun. 2019, 9, e00090. [Google Scholar] [CrossRef]
- Kamada, N.; Yasuhara, A.; Ikeda, M. Significance of the non-oxidative route of the pentose phosphate pathway for supplying carbon to the purine-nucleotide pathway in Corynebacterium ammoniagenes. J. Ind. Microbiol. Biotechnol. 2003, 30, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Hennig, G.; Haupka, C.; Brito, L.F.; Rückert, C.; Cahoreau, E.; Heux, S.; Wendisch, V.F. Methanol-essential growth of Corynebacterium glutamicum: Adaptive laboratory evolution overcomes limitation due to methanethiol assimilation pathway. Int. J. Mol. Sci. 2020, 21, 3617. [Google Scholar] [CrossRef]
- Kundu, B.; Sarkar, D.; Ray, N.; Talukdar, A. Understanding the riboflavin biosynthesis pathway for the development of antimicrobial agents. Med. Res. Rev. 2019, 39, 1338–1371. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, F.; Brito, L.F.; Wendisch, V.F. Function of L-pipecolic acid as compatible solute in Corynebacterium glutamicum as basis for its production under hyperosmolar conditions. Front. Microbiol. 2019, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.-Z.; Ruan, H.-Z.; Chen, X.-L.; Zhang, F.; Zhang, W. Equilibrium of the intracellular redox state for improving cell growth and L-lysine yield of Corynebacterium glutamicum by optimal cofactor swapping. Microb. Cell Factories 2019, 18, 65. [Google Scholar] [CrossRef] [Green Version]
- Valgepea, K.; Adamberg, K.; Nahku, R.; Lahtvee, P.-J.; Arike, L.; Vilu, R. Systems biology approach reveals that overflow metabolism of acetate in Escherichia coli is triggered by carbon catabolite repression of acetyl-CoA synthetase. BMC Syst. Biol. 2010, 4, 166. [Google Scholar] [CrossRef] [Green Version]
- Pronk, J.T.; Yde Steensma, H.; van Dijken, J.P. Pyruvate metabolism in Saccharomyces cerevisiae. Yeast 1996, 12, 1607–1633. [Google Scholar] [CrossRef]
- Cocaign-Bousquet, M.; Lindley, N.D. Pyruvate overflow and carbon flux within the central metabolic pathways of Corynebacterium glutamicum during growth on lactate. Enzyme Microb. Technol. 1995, 17, 260–267. [Google Scholar] [CrossRef]
- Wieschalka, S.; Blombach, B.; Eikmanns, B.J. Engineering Corynebacterium glutamicum for the production of pyruvate. Appl. Microbiol. Biotechnol. 2012, 94, 449–459. [Google Scholar] [CrossRef]
- Khuat, H.B.T.; Kaboré, A.K.; Olmos, E.; Fick, M.; Boudrant, J.; Goergen, J.-L.; Delaunay, S.; Guedon, E. Lactate production as representative of the fermentation potential of Corynebacterium glutamicum 2262 in a one-step process. Biosci. Biotechnol. Biochem. 2014, 78, 343–349. [Google Scholar] [CrossRef] [Green Version]
- Veldmann, K.H.; Dachwitz, S.; Risse, J.M.; Lee, J.-H.; Sewald, N.; Wendisch, V.F. Bromination of L-tryptophan in a fermentative process with Corynebacterium glutamicum. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Georgi, T.; Rittmann, D.; Wendisch, V.F. Lysine and glutamate production by Corynebacterium glutamicum on glucose, fructose and sucrose: Roles of malic enzyme and fructose-1,6-bisphosphatase. Metab. Eng. 2005, 7, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Murai, K.; Sasaki, D.; Kobayashi, S.; Yamaguchi, A.; Uchikura, H.; Shirai, T.; Sasaki, K.; Kondo, A.; Tsuge, Y. Optimal ratio of carbon flux between glycolysis and the pentose phosphate pathway for amino acid accumulation in Corynebacterium glutamicum. ACS Synth. Biol. 2020, 9, 1615–1622. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, F.; Risse, M.J.; Friehs, K.; Wendisch, V.F. Fermentative production of L-pipecolic acid from glucose and alternative carbon sources. Biotechnol. J. 2017, 12. [Google Scholar] [CrossRef]
- Zhang, X.; Lai, L.; Xu, G.; Zhang, X.; Shi, J.; Koffas, M.A.G.; Xu, Z. Rewiring the central metabolic pathway for high-yield l-serine production in Corynebacterium glutamicum by using glucose. Biotechnol. J. 2019, 14, 1800497. [Google Scholar] [CrossRef]
- Zhang, J.; Qian, F.; Dong, F.; Wang, Q.; Yang, J.; Jiang, Y.; Yang, S. De novo engineering of Corynebacterium glutamicum for l-proline production. ACS Synth. Biol. 2020, 9, 1897–1906. [Google Scholar] [CrossRef]
- Mustafi, N.; Grünberger, A.; Mahr, R.; Helfrich, S.; Nöh, K.; Blombach, B.; Kohlheyer, D.; Frunzke, J. Application of a genetically encoded biosensor for live cell imaging of L-valine production in pyruvate dehydrogenase complex-deficient Corynebacterium glutamicum strains. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Mahr, R.; Gätgens, C.; Gätgens, J.; Polen, T.; Kalinowski, J.; Frunzke, J. Biosensor-driven adaptive laboratory evolution of l-valine production in Corynebacterium glutamicum. Metab. Eng. 2015, 32, 184–194. [Google Scholar] [CrossRef]
- Pérez-García, F.; Ziert, C.; Risse, J.M.; Wendisch, V.F. Improved fermentative production of the compatible solute ectoine by Corynebacterium glutamicum from glucose and alternative carbon sources. J. Biotechnol. 2017, 258, 59–68. [Google Scholar] [CrossRef]
- Sanchez, S.; Rodríguez-Sanoja, R.; Ramos, A.; Demain, A.L. Our microbes not only produce antibiotics, they also overproduce amino acids. J. Antibiot. 2017. [Google Scholar] [CrossRef]
- Sgobba, E.; Stumpf, A.K.; Vortmann, M.; Jagmann, N.; Krehenbrink, M.; Dirks-Hofmeister, M.E.; Moerschbacher, B.; Philipp, B.; Wendisch, V.F. Synthetic Escherichia coli-Corynebacterium glutamicum consortia for l-lysine production from starch and sucrose. Bioresour. Technol. 2018, 260, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Chen, T.; Chen, X.; Jingyu, W.; Zhao, X. Enhanced riboflavin production by expressing heterologous riboflavin operon from Bacillus cereus ATCC14579 in Bacillus subtilis. Chin. J. Chem. Eng. 2010, 18, 129–136. [Google Scholar] [CrossRef]
- Lin, Z.; Xu, Z.; Li, Y.; Wang, Z.; Chen, T.; Zhao, X. Metabolic engineering of Escherichia coli for the production of riboflavin. Microb. Cell Factories 2014, 13, 104. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Chen, T.; Ma, X.; Shen, Z.; Zhao, X. Enhancement of riboflavin production with Bacillus subtilis by expression and site-directed mutagenesis of zwf and gnd gene from Corynebacterium glutamicum. Bioresour. Technol. 2011, 102, 3934–3940. [Google Scholar] [CrossRef]
- Burgess, C.; O’Connell-Motherway, M.; Sybesma, W.; Hugenholtz, J.; van Sinderen, D. Riboflavin production in Lactococcus lactis: Potential for in situ production of vitamin-enriched foods. Appl. Environ. Microbiol. 2004, 70, 5769–5777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henke, N.A.; Wendisch, V.F. Improved astaxanthin production with Corynebacterium glutamicum by application of a membrane fusion protein. Mar. Drugs 2019, 17, 621. [Google Scholar] [CrossRef] [Green Version]
- Tsuge, Y.; Hori, Y.; Kudou, M.; Ishii, J.; Hasunuma, T.; Kondo, A. Detoxification of furfural in Corynebacterium glutamicum under aerobic and anaerobic conditions. Appl. Microbiol. Biotechnol. 2014, 98, 8675–8683. [Google Scholar] [CrossRef]
- Walter, T.; Veldmann, K.H.; Götker, S.; Busche, T.; Rückert, C.; Kashkooli, A.B.; Paulus, J.; Cankar, K.; Wendisch, V.F. Physiological response of Corynebacterium glutamicum to indole. Microorganisms 2020, 8, 1945. [Google Scholar] [CrossRef]
- Tsuge, Y.; Kudou, M.; Kawaguchi, H.; Ishii, J.; Hasunuma, T.; Kondo, A. FudC, a protein primarily responsible for furfural detoxification in Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2016, 100, 2685–2692. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains and Plasmids | Description | Source |
|---|---|---|
| Strains | ||
| Corynebacterium glutamicum | wild-type strain ATCC 13032, auxotrophic for biotin | [29] |
| RiboGlu | Corynebacterium glutamicum carrying the plasmids pEKEx3-sigH and pSH1 | This work |
| RiboMan | Corynebacterium glutamicum carrying the plasmids pEKEx3-sigH and pSH1-manA | This work |
| RiboXyl | Corynebacterium glutamicum carrying the plasmids pEKEx3-sigH and pSH1-xylAB | This work |
| RiboSSL | Corynebacterium glutamicum carrying the plasmids pEKEx3-sigH and pSH1-manA-xylAB | This work |
| Plasmids | ||
| pSH1 | KanR, C. glutamicum/E. coli shuttle plasmid (Ptuf, pHM1519 OriVCg) | [30] |
| pEKEx3 | SpecR, C. glutamicum/E. coli shuttle plasmid (Ptac, lacI, pBL1 OriVCg) | [31] |
| pSH1-manA | KanR, pSH1 overexpressing manA from Corynebacterium glutamicum | This work |
| pSH1-xylAB | KanR, pSH1 overexpressing xylA from Xanthomonas campestris and xylB from Corynebacterium glutamicum | This work |
| pSH1-manA-xylAB | KanR, pSH1 overexpressing manA from Corynebacterium glutamicum, xylA from Xanthomonas campestris and xylB from Corynebacterium glutamicum | This work |
| pEKEx3-sigH | SpecR, pEKEx3 overexpressing sigH from Corynebacterium glutamicum | [27] |
| Primers | Sequence (5′ to 3′) |
|---|---|
| xylAB-fw | CCTGCAGGTCGACTCTAGAGGAAAGGAGGCCCTTCAGATGAGCAACACCGTTTTCATC |
| xylAB-rv | GAGCTCGGTACCCGGGGATCTAGTACCAACCCTGCGTTGC |
| manA-fw | CCTGCAGGTCGACTCTAGAGGAAAGGAGGCCCTTCAGATGGAGCTATTGGAAGGCTCAC |
| manA-rv | GAGCTCGGTACCCGGGGATCCTAAACCCTAGCGAGGAATAC |
| xyl-manA-fw | GGTACTAGATCCCCGGGTACGAAAGGAGGCCCTTCAGATGGAGCTATTGGAAGGCTCAC |
| xyl-manA-rv | GGCCAGTGAATTCGAGCTCGGTACCTAAACCCTAGCGAGGAATAC |
| pSH1-conf-fw | ACCGGCTCCAGATTTATCAG |
| pSH1-conf-rv | ATCTTCTCTCATCCGCCA |
| Carbon Source | [Carbon Source] | Growth Rate | Biomass | Biomass Yield | Riboflavin Titer | Riboflavin Yield |
|---|---|---|---|---|---|---|
| g L−1 | h−1 | g L−1 | g g−1 | mg L−1 | mg g−1 | |
| Glucose | 5 | 0.25 ± 0.00 | 2.0 ± 0.1 | 0.39 ± 0.02 | 1.1 ± 0.0 | 0.22 ± 0.01 |
| Glucose | 10 | 0.25 ± 0.01 | 4.2 ± 0.2 | 0.42 ± 0.02 | 2.2 ± 0.3 | 0.22 ± 0.03 |
| Glucose | 20 | 0.25 ± 0.01 | 7.8 ± 0.4 | 0.39 ± 0.02 | 4.5 ± 0.4 | 0.23 ± 0.02 |
| Glucose | 50 | 0.22 ± 0.01 | 19.4 ± 0.7 | 0.39 ± 0.01 | 6.9 ± 0.4 | 0.14 ± 0.01 |
| Glucose | 100 | 0.17 ± 0.00 | 20.9 ± 0.6 | 0.21 ± 0.01 | 7.7 ± 0.8 | 0.08 ± 0.01 |
| Mannose | 5 | 0.14 ± 0.01 | 1.9 ± 0.1 | 0.38 ± 0.02 | 0.8 ± 0.1 | 0.16 ± 0.01 |
| Mannose | 10 | 0.16 ± 0.01 | 3.9 ± 0.1 | 0.39 ± 0.01 | 1.5 ± 0.1 | 0.15 ± 0.01 |
| Mannose | 20 | 0.19 ± 0.01 | 8.0 ± 0.3 | 0.40 ± 0.01 | 3.0 ± 0.3 | 0.15 ± 0.02 |
| Mannose | 50 | 0.15 ± 0.02 | 14.9 ± 0.7 | 0.30 ± 0.01 | 4.6 ± 0.8 | 0.09 ± 0.02 |
| Mannose | 100 | 0.12 ± 0.01 | 14.2 ± 0.7 | 0.14 ± 0.01 | 5.1 ± 0.5 | 0.05 ± 0.00 |
| Xylose | 5 | 0.12 ± 0.01 | 1.4 ± 0.0 | 0.28 ± 0.01 | 1.1 ± 0.1 | 0.22 ± 0.02 |
| Xylose | 10 | 0.13 ± 0.00 | 2.9 ± 0.3 | 0.29 ± 0.03 | 2.6 ± 0.1 | 0.26 ± 0.01 |
| Xylose | 20 | 0.13 ± 0.01 | 5.2 ± 0.4 | 0.26 ± 0.02 | 5.6 ± 0.3 | 0.28 ± 0.01 |
| Xylose | 50 | 0.12 ± 0.00 | 3.9 ± 0.2 | 0.08 ± 0.00 | 8.8 ± 0.8 | 0.18 ± 0.02 |
| Xylose | 100 | 0.11 ± 0.01 | 4.0 ± 0.1 | 0.04 ± 0.00 | 9.4 ± 0.4 | 0.09 ± 0.00 |
| SynSSL | 15 | 0.23 ± 0.01 | 3.5 ± 0.2 | 0.24 ± 0.01 | 1.9 ± 0.1 | 0.12 ± 0.01 |
| SynSSL | 30 | 0.24 ± 0.01 | 8.3 ± 0.6 | 0.28 ± 0.02 | 4.4 ± 0.7 | 0.15 ± 0.02 |
| SynSSL | 60 | 0.24 ± 0.01 | 17.0 ± 0.4 | 0.28 ± 0.01 | 9.9 ± 0.7 | 0.17 ± 0.01 |
| SynSSL | 150 | 0.18 ± 0.00 | 14.0 ± 0.8 | 0.09 ± 0.01 | 12.7 ± 1.2 | 0.08 ± 0.01 |
| SynSSL | 300 | 0.06 ± 0.00 | 2.6 ± 0.2 | 0.01 ± 0.00 | 0.9 ± 0.2 | 0.00 ± 0.00 |
| Carbon Source | Strain | Biomass | Biomass Yield | Riboflavin Titer | Riboflavin Yield | Volumetric Productivity |
|---|---|---|---|---|---|---|
| g L−1 | g g−1 | mg L−1 | mg g−1 | mg L−1 h−1 | ||
| Glucose | RiboGlu | 6.1 | 0.31 | 4.8 | 0.24 | 0.18 |
| Glucose | RiboMan | 9.9 | 0.49 | 7.1 | 0.36 | 0.24 |
| Glucose | RiboXyl | 5.3 | 0.27 | 1.7 | 0.08 | 0.03 |
| Glucose | RiboSSL | 6.5 | 0.33 | 5.3 | 0.26 | 0.12 |
| Mannose | RiboGlu | 2.6 | 0.13 | 3.9 | 0.20 | 0.03 |
| Mannose | RiboMan | 3.9 | 0.20 | 2.6 | 0.13 | 0.04 |
| Xylose | RiboXyl | 6.7 | 0.33 | 6.0 | 0.30 | 0.17 |
| SynSSL | RiboGlu | 8.4 | 0.14 | 6.8 | 0.11 | 0.08 |
| SynSSL | RiboMan | 8.9 | 0.15 | 6.5 | 0.11 | 0.09 |
| SynSSL | RiboXyl | 11.7 | 0.20 | 11.7 | 0.19 | 0.17 |
| SynSSL | RiboSSL | 12.8 | 0.21 | 7.3 | 0.12 | 0.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-García, F.; Burgardt, A.; Kallman, D.R.; Wendisch, V.F.; Bar, N. Dynamic Co-Cultivation Process of Corynebacterium glutamicum Strains for the Fermentative Production of Riboflavin. Fermentation 2021, 7, 11. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010011
Pérez-García F, Burgardt A, Kallman DR, Wendisch VF, Bar N. Dynamic Co-Cultivation Process of Corynebacterium glutamicum Strains for the Fermentative Production of Riboflavin. Fermentation. 2021; 7(1):11. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010011
Chicago/Turabian StylePérez-García, Fernando, Arthur Burgardt, Dina R. Kallman, Volker F. Wendisch, and Nadav Bar. 2021. "Dynamic Co-Cultivation Process of Corynebacterium glutamicum Strains for the Fermentative Production of Riboflavin" Fermentation 7, no. 1: 11. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010011