Impact of Fermentation on Phenolic Compounds and Antioxidant Capacity of Quinoa



CREA Research Centre for Food and Nutrition, Via Ardeatina 546, I-00178 Roma, Italy
*
Author to whom correspondence should be addressed.
Fermentation 2021, 7(1), 20; https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010020
Submission received: 30 December 2020
/
Revised: 25 January 2021
/
Accepted: 27 January 2021
/
Published: 2 February 2021
Abstract
:Quinoa (Chenopodium quinoa Willd.) is increasingly singled out as a healthy food with an excellent nutritional profile. Besides being suitable for gluten-free diets, it is rich in proteins of excellent quality and is a good source of minerals and vitamins, as well as of natural antioxidants, such as phenolic compounds. The aim of this work is to present how fermentation can affect phenolic compound content and antioxidant capacity of quinoa. It emerged that fermentation can be used to increase phenolic compound content and antioxidant capacity in both quinoa seeds and flours. The use of fermented quinoa flours allowed obtaining bread and pasta richer in phenolic compounds and with a greater antioxidant capacity. Fungi are the main starters used in quinoa seed fermentation, while Lactobacillus strains have been applied to produce sourdoughs. Quinoa has been also fermented to obtain yogurt-like beverages with a higher content in phenolic compounds and a greater antioxidant activity. Strains of Lactobacillus sp. and Bifidobacterium sp. have been used as starters.
1. Introduction
Quinoa (Chenopodium quinoa Willd.) is a pseudocereal crop, indigenous to the Andean region, increasingly singled out as a healthy food with an excellent nutritional profile. It has recently attracted a lot of attention, as it is rich in proteins of excellent quality, with a balanced essential amino acid profile similar to milk and close to the ideal balance recommended by the Food and Agriculture Organization of the United Nations (FAO); it is a good source of minerals (e.g., calcium, iron, and zinc), vitamins, and also natural antioxidants. In addition, it is a gluten-free (GF) crop, and its seeds have been milled and widely used to overcome the technological challenges of GF breadmaking [1].
Quinoa is also a crop with resilience traits. It can withstand extreme environmental conditions, in terms of soils, rainfall, temperature, and altitude. It can thus grow from sea level up to 4500 m above sea level and can adapt to frost, drought and salinity [2].
The inclusion of pseudocereals in the diet of populations other than those from Southern America has been long limited because of traditional dietary habits, disadvantages of longer cooking times, a limited variety of products made from them, and the presence of anti-nutritional factors, such as phytates [3]. However, in recent years, the global demand for quinoa has exploded, because of its claimed “superfood” quality.
A great deal of evidence has shown that consumption of fermented foods can have a preventive effect towards non-communicable diseases, due to the presence of bioactive molecules, which can have antioxidant capacity, cholesterol-lowering effect, or antidiabetic activity [4]. Fermentation of grains and derivatives thereof specifically determines the production of health-promoting components, such as γ-aminobutyric acid, conjugated linoleic acid, folates, and phenolic compounds [4]. The latter are mainly present in bound form (conjugates with sugars, fatty acids, or proteins) in non-fermented grains. Glycosylation and conjugation affect the bioavailability and bioaccessibility of phenolic compounds [5]. Upon fermentation, phenolic bound forms are bio-converted into their free forms, which are more bioavailable and bioaccessible [6].
The aim of this work is to present the effect of fermentation on phenolic compounds and antioxidant capacity of quinoa. To this aim, information on this biotechnology and phenolic compound content in raw quinoa seeds is first presented. Changes in phenolic compound content and profile, as well as on antioxidant capacity of fermented quinoa and quinoa-based fermented foods, are then discussed, based on the starters applied.
2. Fermentation and Phenolic Compounds in Grain-Based Foods
Food fermentation is a food processing technology based on the growth and metabolic activity of microorganisms (i.e., bacteria, yeasts, and mycelial fungi, and their enzymes) for the stabilization and transformation of food matrices [7]. It can occur because microorganisms are indigenously present in the substrate or in the processing environment, or because they are added as a starter culture. In the first case, foods are referred to as wild ferments or spontaneous ferments; in the second case, foods are known as culture-dependent ferments [8]. In both cases, food fermentation occurs provided that a suitable substrate, appropriate microorganism(s), and proper environmental conditions, such as temperature, pH, and moisture content, coexist.
During the fermentative process, several biochemical changes occur in the food matrix, and nutritive and anti-nutritive components are modified in terms of bioactivity and digestibility. In non-fermented grains, phenolic compounds are mainly present in bound form, that is, conjugates with sugars, fatty acids, or proteins.
During fermentation, bound phenolic compounds are bio-converted from their linked or conjugated forms to their free ones, because of the (i) breakdown of the bonds with the grain cell wall components; (ii) activities of enzymes, such as β-glucosidase, decarboxylases, esterases, hydrolases, and reductases; and (iii) metabolic activity of fermenting microorganisms [3]. In their free form, phenolic compounds have a greater bioaccessibility, and the released free aglycones have the potential for increasing antioxidative activity [9]. On the other hand, upon fermentation, a decrease of free phenolic compound content can occur, because they may bind with other molecules present in the food matrix, they might be degraded by microbial enzymes, and they might be hydrolysed by specific microbial strains [3].
Since phenolic compounds act as antioxidants, they also contribute to food antioxidant capacity. Hence, changes in phenolic compound content or profile, upon fermentation, can modulate antioxidant capacity. In addition, many lactic acid bacteria themselves possess enzymatic and non-enzymatic antioxidative mechanisms [9].
Fermentation can thus increase or reduce phenolic compound content and antioxidant capacity of grains [10]. The effect and the degree of influence depend on the species of microorganisms involved in the process [9]. It is pivotal to choose an adequate microorganism able to ferment the food matrix and outcompete contaminating flora [11].
3. Phenolic Compounds in Quinoa
Phenolic compounds (PCs) are a diverse group of phytochemicals ranging from simple phenols to complex polyphenols. These chemical structures share the presence of one or more hydroxyl groups on aromatic ring(s).
PCs are produced by plants in normal and stress conditions; hence, they are ubiquitous in plant-derived foods. In cereals and pseudocereals, they are commonly found in the outer layers of the grains. An appropriate dietary intake of PCs contributes to maintaining health and well-being while lowering the risk for the onset of non-communicable diseases thanks to their antioxidant, anti-inflammatory, and anti-proliferative activity [12].
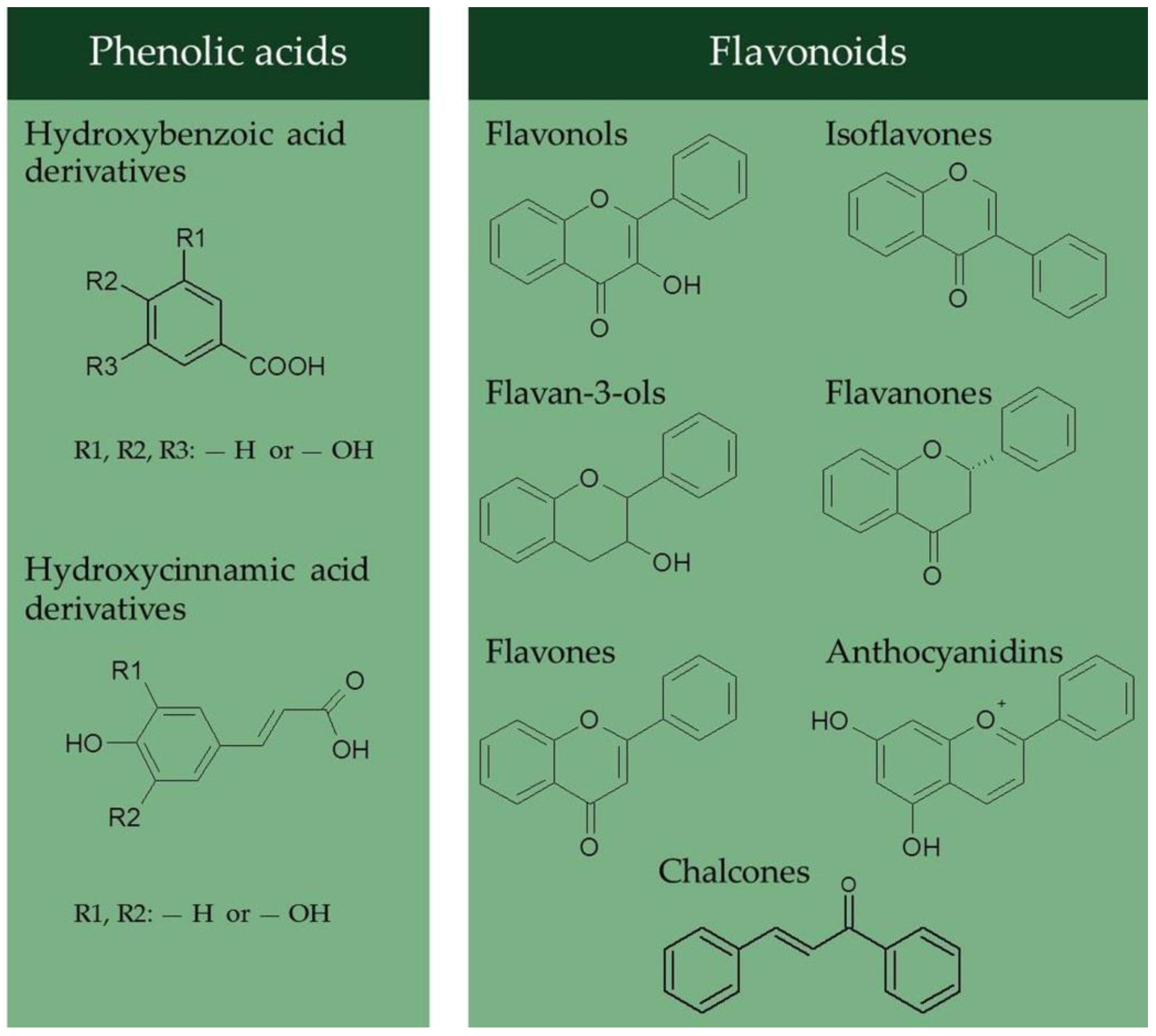
Phenolic acids and flavonoids are the most abundant dietary PCs [12]. The former include hydroxybenzoic and hydroxycinnamic acids derivatives (Figure 1), which occur, respectively, as components of hydrolysable tannins and lignin, and hydroxyacid esters covalently conjugated to the plant cell wall components [13]. Flavonoids show a three-ring structure in the C6–C3–C6 form. They are further classified into flavonols, flavan-3-ols, flavones, isoflavones, flavanones, anthocyanidins, and chalcones, based on the connection of an aromatic ring to the heterocyclic ring (Figure 1).
So far, phenolic compounds have been the most studied non-nutrient phytochemicals in quinoa [14]. They have been mainly quantified by spectrophotometric methods as Total Phenolic Content (TPC). A great variability in TPC has been observed among studies investigating quinoa samples originating from different regions worldwide.
In raw seeds of Titicaca and Puno quinoa, grown in Morocco, TPC was 31.67 and 105.85 mg Gallic Acid Equivalents (GAE) 100 g−1 dry matter (dm), respectively [15]. TPC values ranging from 66 to 202 mg GAE 100 g−1 dm were observed in five quinoa genotypes grown in South Sinai (Egypt) [16]. In the raw seeds of the Chinese quinoa cultivar Jinli-1, TPC was 200.40 mg GAE 100 g−1 dm [17], while in quinoa seeds grown in Korea it was 14.37 mg GAE 100 g−1 dm [18]. Recently, quinoa cultivation has spread also in Europe, and phenolic compounds were determined also in European samples. In raw quinoa seeds grown in Finland, Mattila et al. found that TPC was 181 mg GAE 100 g−1 dm [19], while lower values were reported by Multari et al. [20]. As regards quinoa from Southern America, total free and bound phenolic compounds ranged from 123 to 341 mg GAE 100 g−1 fw (fresh weight) and between 128 and 452 mg GAE 100 g−1 fw, respectively [21]. A lower content of PCs in free form was observed by Vega-Gálvez et al. in samples from Chile [22].
Variability in TPC is due to several factors, such as genetic traits, growing conditions, and post-harvest processing of quinoa seeds. However, differences in analytical procedures for phenolic extraction also contribute to data variability.
Quinoa processing, such as milling and cooking, can modulate the dietary intake of PCs. For instance, upon milling (degree of milling of 27.23%), TPC of the Chinese quinoa cultivar Jinli-1 decreased to 137.36 mg GAE 100 g−1 dm, compared to raw seeds (200.40 mg GAE 100 g−1 dm) [17]. The effect of different types of processing on TPC in quinoa from Brazil has been also recently reported [23]. Compared to the control, toasting significantly decreased TPC, while cooking increased TPC up to 110.65 and 127.54 mg GAE 100 g−1 dm, when it was performed at atmospheric pressure and under pressure, respectively. A higher TPC upon cooking was also observed in cereals [24] and was possibly due to a softening or disintegrating effect on grain tissues that promotes the release of phenolics from the food matrix.
4. Impact of Fermentation on Phenolic Compounds in Quinoa
Fermentation can have multiple effects on phenolic compounds, such as modifications of their content and/or profile, as well as formation of metabolites.
4.1. Fermentation and Phenolic Compound Content
The relationship between fermentation and phenolic compound content has been studied in quinoa seeds and flours.
Microorganisms naturally present in quinoa seeds or previously isolated from them, as well as fungi and Saccharomyces cerevisiae strains (generally applied to the baking and brewing process), were used as fermentation starters.
Strains of Lactobacillus were used to ferment quinoa seeds by Li et al. [25] and Rocchetti et al. [26]. Lactobacillus casei CICC 20995 was used to ferment quinoa seeds from Jiaqi-1, which is an improved variety from the Peruvian altiplano and cultivated in the Shanxi region (China) [25]. In unfermented quinoa seeds, phenolic compounds were found mainly in bound form, which accounted for 85.20%. The level of bound phenolics was 1180 mg GAE 100 g−1 dm, while free phenolic content was 205 mg GAE 100 g−1 dm. In fermented seeds, a 4-fold increase in free phenolics was observed, while the bound phenolic content decreased by approximately 1.5 folds (Table 1). It was supposed that the ability of the enzymatic system of L. casei to catalyze combined polyphenol-protein compounds into a free state was responsible for the decrease in bound phenolic content. Autochthonous lactic acid bacteria (LAB)—Lactobacillus paracasei A1 2.6 and Pediococcus pentosaceus GS B—were used as starters to ferment commercial quinoa seeds native to South America [26]. Quinoa seeds were cooked and acidified before fermentation according to common industrial practices. It was found that fermentation with P. pentosaceus GS B strain was more efficient than with L. paracasei A1 2.6 strain in increasing TPC of quinoa seeds (70.9 vs. 59.6 mg GAE 100 g−1, respectively). However, the most effective process was fermentation of cooked and acidified quinoa seeds by both P. pentosaceus GS B strain and L. paracasei A1 2.6 strain (Table 1). Compared to raw quinoa, TPC increased by 13.8%. The chromatographic analysis of phenolic extracts showed that quinoa was abundant in flavonoids and phenolic acids; however, alkylphenols and lignans were also identified.
Fungi were successfully applied to quinoa seed fermentation, and the effect on PCs was studied. Xu et al. investigated the effect of solid-state fermentation (SSF) by three filamentous fungi (Agaricus bisporus AS2796, Fomitiporia yanbeiensis G1 and Helvella lacunosa X1) on phenolic compound content and antioxidant capacity of quinoa seeds [27]. A. bisporus sp. is also known as the white button mushroom and has anticancer activity, and Fomitiporia (Hymenochaetales, Basidiomycota) has been greatly applied for preventing chronic diseases, while H. lacunosa sp. has been used for enhancing immunity [27]. The application of the above-mentioned strains showed that TPC after SSF was influenced by the starter strains applied and mainly by the fermentation time. When H. lacunosa X1 and F. yanbeiensis G1 were used, after a fermentation time of 14 days, the longer the fermentation time was, the higher TPC was (Table 1). In contrast, the highest TPC value in quinoa fermented by A. bisporus AS2796 was obtained at 35-day fermentation.
Rhizopus microspores var. oligosporus was also used to ferment white quinoa seeds [28]. An increase in TPC from 4.1 to 7.4 mg GAE 100 g−1 dm was observed after a 3-day fermentation. A higher TPC was found after a 5-day fermentation (Table 1). Total Flavonoid Content (TFC) was also determined and a significant increase upon fermentation was observed (Table 1) [28].
The effect of fermentation with three strains of fungi—Rhizopus oligosporus, Aspergillus oryzae and Neurospora intermedia—which are commonly used for the preparation of tempe, koji, and oncom molds, respectively, was explored [29]. The highest value of total phenolics was found when a 6-day fermentation with R. oligosporus was applied. It increased from 0.47 (raw) to 1.96 µg 100 g−1 dm in black quinoa, and up to 0.89 µg 100 g−1 dm in red quinoa (Table 1).
The effect of standard (30 h) and prolonged (40 h) tempe-type fermentation with R. oligosporus ATCC 64063 strain was studied in white, red, and black quinoa [30]. It emerged that both fermentative processes increased TPC compared to raw seeds (Table 1). Moreover, in white and black quinoa, a 40 h fermentation determined a greater increase in TPC than the 30 h treatment.
Baker’s yeast (Saccharomyces cerevisiae NBRC 2375) or brewer’s yeast (S. cerevisiae NBRC 1951) were used to ferment quinoa seeds from Argentina [31]. It was found that TPC in raw quinoa seeds was 39.3 mg GAE 100 g−1 dm and increased by approximately 46% after fermentation with S. cerevisiae NBRC 2375 and NBRC 1951. No statistical differences were found between the two microorganisms (Table 1).
Several Lactobacillus strains were applied also to quinoa flour fermentation. Ayyash et al. studied the effect of solid-state fermentation with two strains of Lactobacillus plantarum—L. plantarum DSM2648 and L. plantarum KX881779—and one strain of Lactobacillus reuteri, KX88177 [32]. TPC in quinoa flour was monitored over 72 h (with 24 h intervals). In non-inoculated quinoa, TPC ranged from approximately 0.11 to 0.18 mg GAE 100 g−1 dm. During 72 h of fermentation, TPC increased significantly (p < 0.05). The highest TPC was found after 72 h fermentation with L. reuteri KX88177 (Table 1).
Quinoa-based ingredients to be used in pasta-making and formulation of bakery products have been obtained from fermentation of quinoa seeds and flours. Lorusso et al. used fermented quinoa flour in pasta-making and investigated the effect on phenolic compound content [33]. Fermentation of raw materials is, in fact, among the possible approaches to obtain functional ingredients for pasta fortification [5]. The autochthonous L. plantarum T6B10 and Lactobacillus rossiae T0A16, isolated from quinoa flour, were used as starters. It was found that the replacement of semolina with raw quinoa flour increased TPC by approximately 37%, and a greater increase (approx. 84%) was observed when fermented quinoa flour was used (Table 1). It was supposed that acidification during fermentation promotes the hydrolysis of complex phenolic compounds and their glycosylated forms into the corresponding phenolic acids which results in a higher content of free total phenolics.
The two LAB strains were also used by Rizzello et al. to obtain a quinoa sourdough for bread-making [34]. TPC in quinoa sourdough was 16.19 mmol kg−1. In the dough obtained by using quinoa sourdough, TPC was 8.78 mmol kg−1, while in the quinoa-based control dough it was 3.65 mmol kg−1 (Table 1).
Fermentation with L. rossiae LB5, L. plantarum 1A7 and Lactobacillus sanfranciscensis DE9 was applied to raw and sprouted quinoa flours [35]. In the obtained sourdough, TPC was 1.8 and 1.2-fold higher than the corresponding flour, respectively (Table 1).
A quinoa sourdough, prepared by fermenting quinoa wholemeal flour with L. plantarum ATCC 8014, was used to make muffins [36]. TFC was 997 mg QE 100 g−1 fw in the wholemeal flour, it decreased in quinoa sourdough and increased up to 1317 mg QE 100 g−1 fw in muffins (Table 1).
Quinoa flour has been fermented to obtain yogurt-like beverages, in order to meet the increasing interest in and demand for non-dairy beverages made with vegetables, fruits, and cereals. Pseudocereals, as well as minor cereals like oat or spelt, and legumes have been, in fact, increasingly investigated as raw ingredients for making functional beverages [39].
Lorusso et al. prepared three yogurt-like beverages by fermenting quinoa with L. rhamnosus SP1, L. plantarum T6B10, and Weissella confusa DSM 20194 [37]. The concentration of total phenols in the fermented beverages was significantly higher than in the non-fermented products. Fermentation with L. rhamnosus SP1 enabled the highest TPC value to be obtained. After 20-day storage, an additional TPC increase was observed (Table 1).
Karovičová et al. prepared fermented quinoa beverages by mixing quinoa flour with water and by fermenting the obtained solution for 6 h with a probiotic culture, including Bifidobacterium sp., Lactobacillus acidophilus, and Streptococcus thermophilus [38]. After fermentation, TPC increased by 27% (Table 1).
4.2. Fermentation and Phenolic Compound Profile
The effect of fermentation on the profile of phenolic compounds has been also investigated. Hydroxybenzoic and hydroxycinnamic acids derivatives, as well as flavonoids, were identified in fermented quinoa and quinoa-based products.
Among hydroxybenzoic acid derivatives, gallic, protocatechuic, and vanillic acids were mainly identified. Gallic acid was detected in quinoa seeds fermented with Rhizopus microspores var. oligosporus (Table 2). It was observed that after 3-day fermentation, gallic acid content increased from 0.01 to 2.37 mg kg−1 but decreased upon longer fermentation (Table 2). Protocathecuic acid was identified in white, red, and black quinoa seeds, and its variation in content was studied upon fermentation with Rhizopus oligosporus ATCC 64063 strain [30]. Protocathecuic acid content was the highest in cooked black quinoa seeds (20.4 µg/g dm) and lowest in white seeds (0.60 µg/g dm) (Table 2). Upon fermentation, protocatechuic acid content ranged from 28.40 to 34.45 µg/g dm in red and black quinoa seeds, while its content was the lowest in white quinoa (1.94–3.69 µg/g dm) (Table 2). Vanillic acid has been determined upon fermentation of quinoa seeds with strains of the Rhizopus sp. and S.cerevisiae. Starzyńska-Janiszewska et al. investigated the content of vanillic acid in bound and free form, in both red and black varieties fermented with a strain of R. oligosporus [29]. As regards the free form, fermentation determined an increase in vanillic acid content in black seeds and a decrease in red seeds, while in the bound form, a 1.13- and 6.23-fold increase occurred upon fermentation of both black and red varieties, respectively (Table 2). The R. oligosporus ATCC 64063 strain was also used to ferment cooked white, red, and black quinoa [30]. The highest increase in vanillic acid content occurred by fermenting for 40 h red quinoa, where it increased from 10.07 to 46.21 µg g−1 dm (Table 2). Vanillic acid content was also monitored in quinoa seeds after a 3- and 5-day fermentation with Rhizopus microspores var. oligosporus [28]. A 1.2–1.4 fold increase occurred (Table 2). A comparable increase was observed by fermenting quinoa seeds with a strain of S. cerevisiae generally used in baking, while a 3.43 fold increase was observed upon fermentation with brewer’s yeast (Table 2) [31]. Alongside vanillic acid, p-hydroxybenzoic acid content also increased when baker’s and brewer’s yeast were applied (Table 2) [31]. On the other hand, a decrease in hydroxybenzoic acid was observed when red and black quinoa seeds were fermented with a strain of R. oligosporus (Table 2) [29].
Regarding hydroxycinnamic acid derivatives, caffeic, chlorogenic, ferulic, and p-coumaric acids were found in quinoa samples. Caffeic acid was identified only among the bound phenolics of red and black quinoa seeds, pre-cooked and fermented with R. oligosporus for 4 days [29]. Its content was 0.034 mg 100 g−1 in fermented seeds of the red variety and 0.160 mg 100 g−1 in fermented black quinoa (Table 2). Chlorogenic acid content increased upon 3-day fermentation with Rhizopus microspores var. oligosporus, while its content did not vary by fermenting quinoa seeds for 5 days (Table 2) [28]. Ferulic acid content decreased upon fermentation with both a strain of R. oligosporus and a strain of S. cerevisiae generally used in brewing (Table 2) [29,31]. On the other hand, when quinoa was fermented with a strain of S. cerevisiae generally used in baking, a 2.35-fold increase in ferulic acid content was observed (Table 2) [31]. Fermentation showed to impact greatly on the content of p-coumaric in quinoa, which increased by 10 and 8.1 fold when the strains of S. cerevisiae generally applied in baking and brewing were used, respectively (Table 2) [31]. On the other hand, when red and black quinoa seeds were fermented with a strain of R. oligosporus sp. [29], p-coumaric decreased (Table 2). In the same sample, a significant decrease in hydroxycinnamic acid was also observed upon fermentation (Table 2) [29].
Regarding flavonoids, catechin was found only in bound form in black and red varieties, and fermentation with R. oligosporus determined a decrease from 0.205 to 0.165 mg 100 g−1 in black seeds and an increase from 0.055 to 0.130 mg 100 g−1 in red seeds (Table 2) [29]. The flavonol kaempferol was found only in free form in red and black quinoa seeds (Table 2) [29]. Upon fermentation with a strain of R. oligosporus sp., its content increased from 0.047 to 0.102 mg 100 g−1 in red quinoa seeds, while in the fermented black seeds it was under the limit of quantitation. After fermentation with the two strains of S. cerevisiae, kaempferol was detected in neither red nor black varieties (Table 2) [31]. Quercetin was found in black and red quinoa in both free and bound form (Table 2). Bound quercetin content increased after fermentation with a strain of R. oligosporus sp., while the free form decreased [29] (Table 2). After fermentation with S. cerevisiae strains, quercetin was also not detected [31]. Free rutin content decreased in red and black quinoa seeds fermented with a strain of R. oligosporus sp. (Table 2) [29], while it increased when it was fermented with the strain R. oligosporus ATCC 64063 for 30 h (Table 2) [30]. Longer fermentation (40 h) negatively affected rutin content (Table 2).
Changes in the phenolic compound profile of quinoa upon fermentation can be due to the activity of microbial enzymes that degrade the grain cell wall structure and thus promote the release of bound phenolic compounds. In addition, biotransformation of phenolic acids by the metabolic pathways of the applied strain can occur. For instance, a 130-fold increase in vanillin content was observed in black quinoa seeds fermented with a strain of R. oligosporus sp. [29]. It is likely that the increase in vanillin was due to microbial biotransformation of ferulic acid to vanillin [40].
The relationship between fermentation and the bioconversion of phenolic compounds in quinoa has been, however, poorly explored and should be further investigated.
5. Impact of Fermentation on Antioxidant Capacity in Quinoa-Based Foods
The antioxidant capacity of a food of plant origin is determined by the concentration and chemical form of components acting as antioxidants. Hence, any processing able to modulate the content of antioxidants in foods impacts their antioxidant capacity.
The antioxidant capacity of quinoa seeds of Jiaqi-1 variety increased after fermentation with L. casei CICC 20995 [25]. It was observed that the half-inhibition concentration (IC50) values of quinoa seeds were 6.65 and 3.43 mg mL−1 before and after fermentation, respectively [25].
Antioxidant capacity of commercial quinoa seeds, native to South America, fermented by L. paracasei A1 2.6 and P. pentosaceus GS B was measured by FRAP and ORAC assays [26]. As far as ORAC assay is concerned, the radical scavenging of fermented quinoa seeds was greater than in raw seeds, and P. pentosaceus enabled the highest value to be obtained (Table 3). When the FRAP assay was applied, a not-detectable reducing power activity was found in all quinoa samples, except for raw quinoa (Table 3).
Antioxidant capacity of quinoa seeds fermented with different fungi was measured by different assays.
Xu et al. measured the antioxidant capacity of quinoa seeds fermented with the three filamentous fungi A. bisporus AS2796, F. yanbeiensis G1, and H. lacunosa X1 by DPPH assay [27]. They found that upon 21- and 35-day fermentation EC50 values were lower than the control, regardless of which fungus was used (Table 3).
The antioxidant capacity of quinoa seeds fermented with R. microspores var. oligosporus was significantly different from the control (raw seeds) only after 5 days of fermentation [28] (Table 3). Antioxidant capacity of colored quinoa fermented with another strain of the Rhyzopus genus, that is, R. oligosporus, and strains of A. oryzae and N. intermedia was measured by ABTS [29]. It emerged that R. oligosporus was the most efficient of the studied strains in enhancing the antioxidant capacity of colored quinoa. In detail, upon a 6-day fermentation, ABTS increased by 276% in black quinoa and by 148% in red quinoa (Table 3).
When quinoa seeds from Argentina were fermented with baker’s yeast (S. cerevisiae NBRC 2375) or brewer’s yeast (S. cerevisiae NBRC 1951), an increase in the antioxidant capacity after fermentation with both strains was observed [31]. Compared to raw quinoa, DPPH values increased by 43 and 33% when baker’s and brewer’s yeast were used, respectively. ABTS values raised by 22 and 27%, and FRAP content increased by 51 and 50% (Table 3).
Antioxidant capacity of quinoa pasta and sourdough started with LAB was measured by DPPH. When two Lactobacillus strains, L. plantarum T6B10 and L. rossiae T0A16, previously isolated from quinoa, were used to produce fermented quinoa pasta [33] and quinoa sourdough bread [34], it was observed that the antioxidant capacity increased. In the study by Lorusso et al. [33], DPPH values increased from 26% in quinoa pasta to 35% in fermented quinoa pasta (Table 3). In quinoa sourdough obtained by fermenting quinoa flour with L. plantarum T6B10 and L. rossiae T0A16, the antioxidant capacity increased up to 71.8% (Table 3) [34].
Montemurro et al. determined the radical scavenging activity in sourdough obtained by raw and sprouted quinoa flour, started with three LAB strains: L. rossiae LB5, L. plantarum 1A7 and L. sanfranciscensis DE9 [35]. Fermentation of raw flour increased DPPH values from 81 to 90%, while a slighter increase was observed when sprouted flour was fermented (from 94 to 97%) (Table 3).
Antioxidant capacity of quinoa flours fermented by SSF with L. plantarum DSM2648, L. reuteri KX88177, and L. plantarum KX881779 had a significant increase (p < 0.05) in DPPH after 24 h of fermentation [32]. The highest value was obtained with L. reuteri KX88177 (Table 3).
Antioxidant capacity of yogurt-like beverages produced by fermenting quinoa with the Lactobacillus strains L. rhamnosus SP1, L. plantarum T6B10, and W. confusa DSM 20194 was significantly different depending on the strain used. The fermented beverage showing the highest antioxidant capacity after fermentation was the one started with L. plantarum T6B10 (Table 3) [37]. Before fermentation, all three beverages showed comparable DPPH values.
6. Conclusions
Fermentation can be used to increase phenolic compound content and antioxidant capacity in both quinoa seeds and flours. Fermented quinoa flours were applied to bread- and pasta-making, and food products richer in phenolic compounds and with greater antioxidant activity were obtained. Fungi were mainly used as starters to ferment quinoa seeds, while Lactobacillus strains were applied to produce sourdoughs. Quinoa has been also fermented to obtain yogurt-like beverages with a higher content in phenolic compounds and a greater antioxidant activity.
In order to maximize the increase in phenolic content, attention should be paid not only to fermentation conditions, but also to quinoa variety used. Studies investigating the effect of fermentation on phenolic compound profile should be encouraged, as well as on the bio-conversion of phenolic compounds.
Author Contributions
Authors equally contributed to the conceptualization, investigation, writing and editing of the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available in the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Melini, F.; Melini, V.; Luziatelli, F.; Ruzzi, M. Current and Forward-Looking Approaches to Technological and Nutritional Improvements of Gluten-Free Bread with Legume Flours: A Critical Review. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1101–1102. [Google Scholar] [CrossRef] [PubMed]
- Bazile, D.; Jacobsen, S.-E.; Verniau, A. The Global Expansion of Quinoa: Trends and Limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adebo, O.A.; Medina-Meza, I.G. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [Green Version]
- Melini, F.; Melini, V.; Luziatelli, F.; Ficca, A.G.; Ruzzi, M. Health-promoting components in fermented foods: An up-to-date systematic review. Nutrients 2019, 11, 1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melini, V.; Melini, F.; Acquistucci, R. Phenolic compounds and bioaccessibility thereof in functional pasta. Antioxidants 2020, 9, 343. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Terefe, N.S. Food Fermentation. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Dimidi, E.; Cox, S.R.; Rossi, M.; Whelan, K. Fermented foods: Definitions and characteristics, impact on the gut microbiota and effects on gastrointestinal health and disease. Nutrients 2019, 11, 1806. [Google Scholar] [CrossRef] [Green Version]
- Hur, S.J.; Lee, S.Y.; Kim, Y.C.; Choi, I.; Kim, G.B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef]
- Gan, R.Y.; Li, H.; Gunaratne, A.; Sui, Z.Q.; Corke, H. Effects of Fermented Edible Seeds and Their Products on Human Health: Bioactive Components and Bioactivities. Compr. Rev. Food Sci. Food Saf. 2017, 16, 489–531. [Google Scholar] [CrossRef]
- Paz, P.C.; Janny, R.J.; Håkansson, Å. Safeguarding of quinoa beverage production by fermentation with Lactobacillus plantarum DSM 9843. Int. J. Food Microbiol. 2020, 324, 108630. [Google Scholar]
- Koch, W. Dietary Polyphenols-Important Non-Nutrients in the Prevention of Chronic Noncommunicable Diseases. Syst. Rev. 2019, 11, 1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahidi, F.; Varatharajan, V.; Oh, W.Y.; Peng, H. Phenolic compounds in agri-food by-products, their bioavailability and health effects. J. Food Bioact. 2019, 5, 57–119. [Google Scholar] [CrossRef] [Green Version]
- Melini, V.; Melini, F. Functional components and anti-nutritional factors in gluten-free grains: A focus on quinoa seeds. Foods 2021. in accepted. [Google Scholar]
- Mhada, M.; Metougui, M.L.; El Hazzam, K.; El Kacimi, K.; Yasri, A. Variations of Saponins, Minerals and Total Phenolic Compounds Due to Processing and Cooking of Quinoa (Chenopodium quinoa Willd.) Seeds. Foods 2020, 9, 660. [Google Scholar] [CrossRef] [PubMed]
- Saad-Allah, K.M.; Youssef, M.S. Phytochemical and genetic characterization of five quinoa (Chenopodium quinoa Willd.) genotypes introduced to Egypt. Physiol. Mol. Biol. Plants 2018, 24, 617–629. [Google Scholar] [CrossRef]
- Han, Y.; Chi, J.; Zhang, M.; Zhang, R.; Fan, S.; Dong, L.; Huang, F.; Liu, L. Changes in saponins, phenolics and antioxidant activity of quinoa (Chenopodium quinoa willd) during milling process. LWT 2019, 114, 108381. [Google Scholar]
- Lim, J.G.; Park, H.; Yoon, K.S. Analysis of saponin composition and comparison of the antioxidant activity of various parts of the quinoa plant (Chenopodium quinoa Willd.). Food Sci. Nutr. 2020, 8, 694–702. [Google Scholar] [CrossRef] [Green Version]
- Mattila, P.H.; Pihlava, J.M.; Hellström, J.; Nurmi, M.; Eurola, M.; Mäkinen, S.; Jalava, T.; Pihlanto, A. Contents of phytochemicals and antinutritional factors in commercial protein-rich plant products. Food Qual. Saf. 2018, 2, 213–219. [Google Scholar] [CrossRef]
- Multari, S.; Marsol-Vall, A.; Keskitalo, M.; Yang, B.; Suomela, J.P. Effects of different drying temperatures on the content of phenolic compounds and carotenoids in quinoa seeds (Chenopodium quinoa) from Finland. J. Food Compos. Anal. 2018, 72, 75–82. [Google Scholar] [CrossRef]
- Abderrahim, F.; Huanatico, E.; Segura, R.; Arribas, S.; Carmen Gonzalez, M.; Condezo-Hoyos, L. Physical features, phenolic compounds, betalains and total antioxidant capacity of coloured quinoa seeds (Chenopodium quinoa Willd.) from Peruvian Altiplano. Food Chem. 2015, 183, 83–90. [Google Scholar] [CrossRef]
- Vega-Gálvez, A.; Zura, L.; Lutz, M.; Jagus, R.; Victoria Agüero, M.; Pastén, A.; Di Scala, K.; Uribe, E. Assessment of dietary fiber, isoflavones and phenolic compounds with antioxidant and antimicrobial properties of quinoa (Chenopodium quinoa Willd.). Chil. J. Agric. Anim. Sci. 2018, 34, 1–11. [Google Scholar] [CrossRef]
- Nickel, J.; Spanier, L.P.; Botelho, F.T.; Gularte, M.A.; Helbig, E. Effect of different types of processing on the total phenolic compound content, antioxidant capacity, and saponin content of Chenopodium quinoa Willd grains. Food Chem. 2016, 209, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Melini, V.; Panfili, G.; Fratianni, A.; Acquistucci, R. Bioactive compounds in rice on Italian market: Pigmented varieties as a source of carotenoids, total phenolic compounds and anthocyanins, before and after cooking. Food Chem. 2019, 277, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, C.; Ji, Y.; Lin, J.; Chen, X.; Qi, B. Improvement of nutritional value, bioactivity and volatile constituents of quinoa seeds by fermentation with Lactobacillus casei. J. Cereal Sci. 2018, 84, 83–89. [Google Scholar] [CrossRef]
- Rocchetti, G.; Miragoli, F.; Zacconi, C.; Lucini, L.; Rebecchi, A. Impact of cooking and fermentation by lactic acid bacteria on phenolic profile of quinoa and buckwheat seeds. Food Res. Int. 2019, 119, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.N.; Guo, S.; Zhang, S.W. Effects of solid-state fermentation on the nutritional components and antioxidant properties from quinoa. Emir. J. Food Agric. 2019, 31, 39–45. [Google Scholar] [CrossRef]
- Hur, J.; Nguyen, T.T.H.; Park, N.; Kim, J.; Kim, D. Characterization of quinoa (Chenopodium quinoa) fermented by Rhizopus oligosporus and its bioactive properties. AMB Express 2018, 8, 143. [Google Scholar] [CrossRef]
- Starzyńska-Janiszewska, A.; Stodolak, B.; Gómez-Caravaca, A.M.; Mickowska, B.; Martin-Garcia, B.; Byczyński, Ł. Mould starter selection for extended solid-state fermentation of quinoa. LWT 2019, 99, 231–237. [Google Scholar] [CrossRef]
- Starzyńska-Janiszewska, A.; Duliński, R.; Stodolak, B.; Mickowska, B.; Wikiera, A. Prolonged tempe-type fermentation in order to improve bioactive potential and nutritional parameters of quinoa seeds. J. Cereal Sci. 2016, 71, 116–121. [Google Scholar] [CrossRef]
- Carciochi, R.A.; Galván-D’Alessandro, L.; Vandendriessche, P.; Chollet, S. Effect of Germination and Fermentation Process on the Antioxidant Compounds of Quinoa Seeds. Plant Foods Hum. Nutr. 2016, 71, 361–367. [Google Scholar] [CrossRef]
- Ayyash, M.; Johnson, S.K.; Liu, S.Q.; Mesmari, N.; Dahmani, S.; Al Dhaheri, A.S.; Kizhakkayil, J. In vitro investigation of bioactivities of solid-state fermented lupin, quinoa and wheat using Lactobacillus spp. Food Chem. 2019, 275, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Lorusso, A.; Verni, M.; Montemurro, M.; Coda, R.; Gobbetti, M.; Rizzello, C.G. Use of fermented quinoa flour for pasta making and evaluation of the technological and nutritional features. LWT Food Sci. Technol. 2017, 78, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Rizzello, C.G.; Lorusso, A.; Montemurro, M.; Gobbetti, M. Use of sourdough made with quinoa (Chenopodium quinoa) flour and autochthonous selected lactic acid bacteria for enhancing the nutritional, textural and sensory features of white bread. Food Microbiol. 2016, 56, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Montemurro, M.; Pontonio, E.; Gobbetti, M.; Rizzello, C.G. Investigation of the nutritional, functional and technological effects of the sourdough fermentation of sprouted flours. Int. J. Food Microbiol. 2019, 302, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Chiş, M.S.; Păucean, A.; Man, S.M.; Vodnar, D.C.; Teleky, B.-E.; Pop, C.R.; Stan, L.; Borsai, O.; Kadar, C.B.; Urcan, A.C.; et al. Quinoa Sourdough Fermented with Lactobacillus plantarum ATCC 8014 Designed for Gluten-Free Muffins—A Powerful Tool to Enhance Bioactive Compounds. Appl. Sci. 2020, 10, 7140. [Google Scholar] [CrossRef]
- Soltani, M.; Hekmat, S.; Ahmadi, L. Microbial and sensory evaluation of probiotic yoghurt supplemented with cereal/pseudo-cereal grains and legumes. Int. J. Dairy Technol. 2018, 71, 141–148. [Google Scholar] [CrossRef]
- Lorusso, A.; Coda, R.; Montemurro, M.; Rizzello, C. Use of Selected Lactic Acid Bacteria and Quinoa Flour for Manufacturing Novel Yogurt-Like Beverages. Foods 2018, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- Karovičová, J.; Kohajdová, Z.; Minarovičová, L.; Lauková, M.; Greifová, M.; Greif, G.; Hojerová, J. Utilisation of quinoa for development of fermented beverages. Potravin. Slovak J. Food Sci. 2020, 14, 465–472. [Google Scholar] [CrossRef]
- Shanker, K.S.; Kishore, K.H.; Kanjilal, S.; Misra, S.; Murty, U.S.N.; Prasad, R.B.N. Biotransformation of ferulic acid to acetovanillone using Rhizopus oryzae. Biocatal. Biotransform. 2007, 25, 109–112. [Google Scholar] [CrossRef]
Figure 1.
Chemical structure of the main dietary phenolic compounds.


{kind=link}
Table 1.
Effect of fermentation on phenolic content in quinoa-based fermented foods.
| Raw Material | Fermented Quinoa-Based Food | Microorganism(s) | Phenolic Content in Raw Quinoa/Quinoa-Based Food | Phenolic Content in Fermented Food | Reference |
|---|---|---|---|---|---|
| Quinoa seeds | Fermented quinoa seeds | Lactobacillus casei CICC 20995 | TPC (free): 205 1 TPC (bound): 1180 1 | TPC (free): 910 1 TPC (bound): 743 1 | Li et al. [25] |
| Quinoa cooked and acidified seeds | Fermented quinoa seeds | Lactobacillus paracasei A1 2.6 strain and/or Pediococcus pentosaceus GS·B strain | TPC (raw): 65.7 1 TPC (cooked): 70.3 1 TPC (cooked+acidified): 60.8 1 | TPC (L. paracasei): 59.6 1 TPC (P. pentosaceus): 70.9 1 TPC (L. paracasei + P. pentosaceus): 74.8 1 | Rocchetti et al. [26] |
| Quinoa seeds | Fermented quinoa seeds | Agaricus bisporus AS2796 | TPC: ≈ 70 1 | TPC (7th day): ≈ 73 1 TPC (14th day): ≈ 81 1 TPC (21th day): ≈ 110 1 TPC (28th day): ≈ 135 1 TPC (35th day): ≈ 138 1 | Xu et al. [27] |
| Quinoa seeds | Fermented quinoa seeds | Fomitiporia yanbeiensis G1 | TPC: ≈ 70 1 | TPC (7th day): ≈ 68 1 TPC (14th day): ≈ 79 1 TPC (21th day): ≈ 110 1 TPC (28th day): ≈ 138 1 TPC (35th day): ≈ 130 1 | Xu et al. [27] |
| Quinoa seeds | Fermented quinoa seeds | Helvella lacunosa X1 | TPC: ≈ 70 1 | TPC (7th day): ≈ 69 1 TPC (14th day): ≈ 79 1 TPC (21th day): ≈ 110 1 TPC (28th day): ≈ 138 1 TPC (35th day): ≈130 1 | Xu et al. [27] |
| Quinoa seeds | Fermented quinoa seeds | Rhizopus microspores var. oligosporus | TPC: 4.1 1 TFC: 1.3 5 | TPC (3 days): 7.4 1 TPC (5 days): 8.0 1 TFC (3 days): 1.6 5 TFC (5 days): 1.9 5 | Hur et al. [28] |
| Quinoa seeds (Black and red varieties from Bolivia) | Fermented quinoa seeds | Aspergillus oryzae | TP (black, raw): 0.47 2 TP (black, pre-cooked): 0.31 2 TP (red, raw): 0.47 2 TP (red, pre-cooked): 0.31 2 | TP (black, 2 days): 0.53 2 TP (black, 4 days): 0.52 2 TP (black, 6 days): 0.56 2 TP (red, 2 days): 0.47 2 TP (red, 4 days): 0.52 2 TP (red, 6 days): 0.60 2 | Starzyńska-Janiszewska et al. [29] |
| Quinoa seeds (Black and red varieties from Bolivia) | Fermented quinoa seeds | Neurospora intermedia | TP (black, raw): 0.47 2 TP (black, pre-cooked): 0.31 2 TP (red, raw): 0.47 2 TP (red, pre-cooked): 0.31 2 | TP (black, 3 days): 0.54 2 TP (black, 5 days): 0.54 2 TP (red, 3 days): 0.51 2 TP (red, 5 days): 0.53 2 | Starzyńska-Janiszewska et al. [29] |
| Quinoa seeds (Black and red varieties from Bolivia) | Fermented quinoa seeds | Rhizopus oligosporus | TP (black, raw): 0.47 2 TP (black, pre-cooked): 0.31 2 TP (red, raw): 0.47 2 TP (red, pre-cooked): 0.31 2 | TP (black, 2 days): 0.77 2 TP (black, 4 days): 1.82 2 TP (black, 6 days): 1.96 2 TP (red, 2 days): 0.50 2 TP (red, 4 days): 0.84 2 TP (red, 6 days): 0.89 2 | Starzyńska-Janiszewska et al. [29] |
| Quinoa seeds (white, red, black varieties) | Fermented quinoa seeds | R. oligosporus ATCC 64063 strain | TPC (white, cooked): 2.76 1 TPC (red, cooked): 2.73 1 TPC (black, cooked): 3.28 1 | TPC (white, fermented 30 h): 8.71 1 TPC (white, fermented 40 h): 9.12 1 TPC (red, fermented 30 h): 7.22 1 TPC (red, fermented 40 h): 7.08 1 TPC (black, fermented 30 h): 7.65 1 TPC (black, fermented 40 h): 8.46 1 | Starzyńska-Janiszewska et al. [30] |
| Quinoa seeds | Fermented quinoa seeds | Saccharomyces cerevisiae NBRC 2375 and NBRC 1951 | TPC: 39.3 1 | TPC (NBRC 2375): ≈ 57 1 TPC (NBRC 1951): ≈ 55 1 | Carciochi et al. [31] |
| Whole grain quinoa flour | Fermented quinoa whole grain flour | Lactobacillus plantarum DSM2648 | TPC (0 h): ≈ 0.13 1 TPC (24 h): ≈ 0.11 1 TPC (48 h): ≈ 0.14 1 TPC (72 h): ≈ 0.18 1 | TPC (0 h): ≈ 0.16 1 TPC (24 h): ≈ 0.36 1 TPC (48 h): ≈ 0.34 1 TPC (72 h): ≈ 0.39 1 | Ayyash et al. [32] |
| Whole grain quinoa flour | Fermented quinoa whole grain flour | L. plantarum KX881779 | TPC (0 h): ≈ 0.13 1 TPC (24 h): ≈ 0.11 1 TPC (48 h): ≈ 0.14 1 TPC (72 h): ≈ 0.18 1 | TPC (0 h): ≈ 0.10 1 TPC (24 h): ≈ 0.20 1 TPC (48 h): ≈ 0.28 1 TPC (72 h): ≈ 0.37 1 | Ayyash et al. [32] |
| Whole grain quinoa flour | Fermented quinoa whole grain flour | Lactobacillus reuteri KX88177 | TPC (0 h): ≈ 0.13 1 TPC (24 h): ≈ 0.11 1 TPC (48 h): ≈ 0.14 1 TPC (72 h): ≈ 0.18 1 | TPC (0 h): ≈ 0.11 1 TPC (24 h): ≈ 0.17 1 TPC (48 h): ≈ 0.36 1 TPC (72 h): ≈ 0.45 1 | Ayyash et al. [32] |
| Quinoa flour | Fermented quinoa pasta | L. plantarum T6B10 Lactobacillus rossiae T0A16 | TPC (semolina pasta): 2.21 3 TPC (quinoa pasta): 3.02 3 | TPC (fermented quinoa pasta): 4.06 3 | Lorusso et al. [33] |
| Quinoa flour | Quinoa sourdough | L. plantarum T6B10 L. rossiae T0A16 | TPC: 3.65 3 | TPC: 8.78 3 | Rizzello et al. [34] |
| Quinoa raw flour | Quinoa sourdough | L. rossiae LB5 L. plantarum 1A7 Lactobacillus sanfranciscensis DE9 | TPC: 3.28 3 | TPC: 5.79 3 | Montemurro et al. [35] |
| Quinoa sprouted flour | Quinoa sourdough | L. rossiae LB5 L. plantarum 1A7 L. sanfranciscensis DE9 | TPC: 5.44 3 | TPC: 6.63 3 | Montemurro et al. [35] |
| Quinoa wholemeal flour | Quinoa sourdough | L. plantarum ATCC 8014 | TFC: 997 4 | TFC (sourdough, 24 h fermentation): 757 4 TFC (muffin): 1317 4 | Chiş et al. [36] |
| Quinoa flour | Yogurt-like beverages | Lactobacillus rhamnosus SP1 L. plantarum T6B10 Weissella confusa DSM 20194 | TPC (B-SP1): 5.3 3 TPC (B-T6B10): 5.2 3 TPC (B-DSM 20194): 4.0 3 | TPC (B-SP1): 5.8 3 TPC (B-T6B10): 8.4 3 TPC (B-DSM 20194): 5.9 3 TPC (B-SP1, 20-day storage): 9.6 3 TPC (B-T6B10, 20-day storage): 9.3 3 TPC (B-DSM 20194, 20-day storage): 7.9 3 | Lorusso et al. [37] |
| Quinoa flour | Fermented quinoa beverages | Commercial probiotic culture including Bifidobacterium sp., Lactobacillus acidophilus, and Streptococcus thermophilus | TPC: 142.37 1 | TPC: 180.33 1 | Karovičová et al. [38] |
1: mg Gallic Acid Equivalents 100 g−1; 2: g 100 g−1 dm; 3: mmol Gallic Acid Equivalents (GAE) kg−1; 4: mg Quercetin Equivalents (QE) 100 g−1 fw; 5: mg Quercetin Equivalents (QE) 100 g−1 dm; TPC: Total Phenolic Content; TP: Total Phenolics; TFC: Total Flavonoid Content.
Table 2.
Effect of fermentation on phenolic compound profile in quinoa-based fermented foods.
| Phenolic Compound | Food Matrix | Content in Raw Quinoa | Content in Fermented Quinoa | Microorganism(s) | Reference |
|---|---|---|---|---|---|
| Hydroxybenzoic acid derivatives | |||||
| Gallic acid | Quinoa seeds | 0.01 mg kg−1 | 2.37 mg kg−1 (3-day fermentation) 0.84 mg kg−1 (5-day fermentation) | Rhizopus microspores var. oligosporus | Hur et al. [28] |
| Hydroxybenzoic acid (bound form) | Red and black quinoa seeds | 1.890 mg 100 g−1 (black) 5.530 mg 100 g−1 (red) | 1.680 mg 100 g−1 (black) 1.775 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| p-Hydroxybenzoic acid | Quinoa seeds | 100% | 831% area relative to raw grain | S. cerevisiae (baker’s yeast) | Carciochi et al. [31] |
| p-Hydroxybenzoic acid | Quinoa seeds | 100% | 868% area relative to raw grain | S. cerevisiae (brewer’s yeast) | Carciochi et al. [31] |
| Protocatechuic acid | White quinoa seeds | 0.60 µg g−1 dm (cooked) | 1.94 µg g−1 dm (30 h) 3.69 µg g−1 dm (40 h) | Rhizopus oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Protocatechuic acid | Red quinoa seeds | 16.09 µg g−1 dm (cooked) | 28.40 µg g−1 dm (30 h) 29.19 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Protocatechuic acid | Black quinoa seeds | 20.4 µg g−1 dm (cooked) | 28.48 µg g−1 dm (30 h) 34.45 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Vanillic acid (free form) | Black and red quinoa seeds | 0.222 mg 100 g−1 (black) 0.762 mg 100 g−1 (red) | 0.482 mg 100 g−1 (black) 0.690 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Vanillic acid (bound form) | Black and red quinoa seeds | 0.990 mg 100 g−1 (black) 0.195 mg 100 g−1 (red) | 1.125 mg 100 g−1 (black) 1.215 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Vanillic acid | White quinoa seeds | 22.02 µg g−1 dm (cooked) | 42.05 µg g−1 dm (30 h) 44.68 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Vanillic acid | Red quinoa seeds | 10.07 µg g−1 dm (cooked) | 43.60 µg g−1 dm (30 h) 46.21 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Vanillic acid | Black quinoa seeds | 12.08 µg g−1 dm (cooked) | 23.10 µg g−1 dm (30 h) 20.86 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Vanillic acid | Quinoa seeds | 1.3 mg kg−1 | 1.55 mg kg−1 (3-day fermentation) 1.83 mg kg−1 (5-day fermentation) | Rhizopus microspores var. oligosporus | Hur et al. [28] |
| Vanillic acid | Quinoa seeds | 100% | 142% area relative to raw grain | S. cerevisiae (baker’s yeast) | Carciochi et al. [31] |
| Vanillic acid | Quinoa seeds | 100% | 343% area relative to raw grain | S. cerevisiae (brewer’s yeast) | Carciochi et al. [31] |
| Hydroxycinnamic acid derivatives | |||||
| Caffeic acid (bound form) | Black and red quinoa seeds | 0.105 mg 100 g−1 (black) 0.030 mg 100 g−1 (red) | 0.160 mg 100 g−1 (black) 0.034 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Chlorogenic acid | Quinoa seeds | 0.002 mg kg−1 | 0.03 mg kg−1 (3-day fermentation) 0.002 mg kg−1 (5-day fermentation) | Rhizopus microspores var. oligosporus | Hur et al. [28] |
| Ferulic acid (free form) | Black and red quinoa seeds | 0.650 mg 100 g−1 (black) 0.888 mg 100 g−1 (red) | 0.230 mg 100 g−1 (black) 0.273 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Ferulic acid (bound form) | Black and red quinoa seeds | 0.16 mg 100 g−1 (black) 0.125 mg 100 g−1 (red) | 0.13 mg 100 g−1 (black) 0.082 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Ferulic acid | Quinoa seeds | 100% | 235% area relative to raw grain | S. cerevisiae (baker’s yeast) | Carciochi et al. [31] |
| Ferulic acid | Quinoa seeds | 100% | 61% area relative to raw grain | S. cerevisiae (brewer’s yeast) | Carciochi et al. [31] |
| Hydroxycinnamic acid (free form) | Black and red quinoa seeds | 12.48 mg 100 g−1 (black) 3.620 mg 100 g−1 (red) | 1.282 mg 100 g−1 (black) 0.670 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| p-Coumaric acid (bound form) | Black and red quinoa seeds | 1.015 mg 100 g−1 (black) 0.025 mg 100 g−1 (red) | 0.720 mg 100 g−1 (black) 0.014 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| p-Coumaric acid | Quinoa seeds | 100% | 1002% area relative to raw grain | S. cerevisiae (baker’s yeast) | Carciochi et al. [31] |
| p-Coumaric acid | Quinoa seeds | 100% | 813% area relative to raw grain | S. cerevisiae (brewer’s yeast) | Carciochi et al. [31] |
| Flavonoids | |||||
| Catechin (bound form) | Black and red quinoa seeds | 0.205 mg 100 g−1 (black) 0.055 mg 100 g−1 (red) | 0.165 mg 100 g−1 (black) 0.130 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Kaempferol (free form) | Black and red quinoa seeds | 0.061 mg 100 g−1 (black) 0.047 mg 100 g−1 (red) | n.q. (black) 0.102 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Kaempferol (bound form) | Black and red quinoa seeds | n.q. (black) n.q. (red) | n.q. (black) n.q. (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Kaempferol | Quinoa seeds | 100% | n.d. | S. cerevisiae (baker’s yeast) | Carciochi et al. [31] |
| Kaempferol | Quinoa seeds | 100% | n.d. | S. cerevisiae (brewer’s yeast) | Carciochi et al. [31] |
| Quercetin (free form) | Black and red quinoa seeds | 0.082 mg 100 g−1 (black) 0.633 mg 100 g−1 (red) | n.q. (black) 0.187 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Quercetin (bound form) | Black and red quinoa seeds | 0.022 mg 100 g−1 (black) 0.043 mg 100 g−1 (red) | 0.061 mg 100 g−1 (black) 0.080 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Quercetin | Quinoa seeds | 100% | n.d. | S. cerevisiae (baker’s yeast) | Carciochi et al. [31] |
| Quercetin | Quinoa seeds | 100% | n.d. | S. cerevisiae (brewer’s yeast) | Carciochi et al. [31] |
| Rutin (free form) | Black and red quinoa seeds | 0.326 mg 100 g−1 (black) 0.580 mg 100 g−1 (red) | 0.054 mg 100 g−1 (black) 0.309 mg 100 g−1 (red) | R. oligosporus | Starzyńska-Janiszewska et al. [29] |
| Rutin | White quinoa seeds | 84.34 µg g−1 dm (cooked) | 107.09 µg g−1 dm (30 h) 104.68 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Rutin | Red quinoa seeds | 57.70 µg g−1 dm (cooked) | 235.11 µg g−1 dm (30 h) 229.98 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
| Rutin | Black quinoa seeds | 39.53 µg g−1 dm (cooked) | 56.37 µg g−1 dm (30 h) 55.52 µg g−1 dm (40 h) | R. oligosporus ATCC 64063 strain | Starzyńska-Janiszewska et al. [30] |
n.d.: not detected; n.q.: not quantitated.
Table 3.
Effect of fermentation on antioxidant capacity in quinoa-based fermented foods.
| Raw Material | Fermented Quinoa-Based Food | Microorganism(s) | Antioxidant Capacity in Raw Quinoa/Quinoa-Based Food | Antioxidant Capacity in Fermented Food | Reference |
|---|---|---|---|---|---|
| Quinoa seeds | Fermented quinoa seeds | Lactobacillus casei CICC 20995 | IC50: 6.65 5 | IC50: 3.43 5 | Li et al. [25] |
| Quinoa cooked and acidified seeds | Quinoa fermented seeds | Lactobacillus paracasei A1 2.6 strain and/or Pediococcus pentosaceus GS·B strain | FRAP (raw): 15.4 4 FRAP (cooked): n.d. FRAP (acidified): n.d. ORAC (raw): 45.84 2 ORAC (cooked): 79.45 2 ORAC (acidified): 66.88 2 | FRAP (L. paracasei): n.d. FRAP (P. pentosaceus): n.d. FRAP (L. paracasei + P. pentosaceus): n.d. ORAC (L. paracasei): 59.68 2 ORAC (P. pentosaceus): 70.11 2 ORAC (L. paracasei + P. pentosaceus): 68.55 2 | Rocchetti et al. [26] |
| Quinoa seeds | Fermented quinoa | Helvella lacunosa X1 | DPPH ≈ 47 1 | DPPH (7th day): ≈ 45 1 DPPH (14th day): ≈ 51 1 DPPH (21th day): ≈ 23 1 DPPH (28th day): ≈ 9 1 DPPH (35th day): ≈ 8 1 | Xu et al. [27] |
| Quinoa seeds | Fermented quinoa | Agaricus bisporus AS2796 | DPPH ≈ 47 1 | DPPH (7th day): ≈ 45 1 DPPH (14th day): ≈ 53 1 DPPH (21th day): ≈ 22 1 DPPH (28th day): ≈ 9 1 DPPH (35th day): ≈ 8 1 | Xu et al. [27] |
| Quinoa seeds | Fermented quinoa | Fomitiporia yanbeiensis G1 | DPPH ≈ 47 1 | DPPH (7th day): ≈ 45 1 DPPH (14th day): ≈ 53 1 DPPH (21th day): ≈ 22 1 DPPH (28th day): ≈ 9 1 DPPH (35th day): ≈ 8 1 | Xu et al. [27] |
| Quinoa seeds | Fermented quinoa seeds | Rhizopus microspores var. oligosporus | DPPH: 3.6 5 | DPPH (3 days): 3.4 5 DPPH (5 days): 2.3 5 | Hur et al. [28] |
| Quinoa seeds (Black and red varieties from Bolivia) | Fermented quinoa seeds | Rhizopus oligosporus | ABTS (black): 49.20 2 ABTS (red): 42.22 2 | ABTS (black, 2 days): 84.38 2 ABTS (black, 4 days): 185.06 2 ABTS (black, 6 days): 227.71 2 ABTS (red, 2 days): 55.68 2 ABTS (red, 4 days): 82.37 2 ABTS (red, 6 days): 105.01 2 | Starzyńska-Janiszewska et al. [29] |
| Quinoa seeds (Black and red varieties from Bolivia) | Fermented quinoa seeds | Aspergillus oryzae | ABTS (black): 49.20 2 ABTS (red): 42.22 2 | ABTS (black, 2 days): 50.13 2 ABTS (black, 4 days): 47.91 2 ABTS (black, 6 days): 48.95 2 ABTS (red, 2 days): 42.40 2 ABTS (red, 4 days): 49.00 2 ABTS (red, 6 days): 53.13 2 | Starzyńska-Janiszewska et al. [29] |
| Quinoa seeds (Black and red varieties from Bolivia) | Fermented quinoa seeds | Neurospora intermedia | ABTS (black): 49.20 2 ABTS (red): 42.22 2 | ABTS (black, 3 days): 53.70 2 ABTS (black, 5 days):56.15 2 ABTS (red, 3 days): 52.23 2 ABTS (red, 5 days): 54.93 2 | Starzyńska-Janiszewska et al. [29] |
| Quinoa seeds | Fermented quinoa seeds | S. cerevisiae NBRC 2375 S. cerevisiae NBRC 1951 | DPPH: 2.60 2 ABTS: 3.50 2 FRAP: 1.20 2 | DPPH (NBRC 2375): ≈ 3.70 2 ABTS (NBRC 2375): ≈ 4.40 2 FRAP (NBRC 2375): ≈ 1.90 2 DPPH (NBRC 1951): ≈ 3.50 2 ABTS (NBRC 1951): ≈ 4.50 2 FRAP (NBRC 1951): ≈ 185 2 | Carciochi et al. [31] |
| Quinoa flour | Fermented quinoa pasta | L. plantarum T6B10 L. rossiae T0A16 | DPPH (semolina pasta): 14 3 DPPH (quinoa pasta): 26 3 | DPPH: 35 3 | Lorusso et al. [33] |
| Quinoa flour | Quinoa sourdough | L. plantarum T6B10 L. rossiae T0A16 | DPPH: 41.7 3 | DPPH: 71.8 3 | Rizzello et al. [34] |
| Quinoa raw flour | Quinoa sourdough | L. rossiae LB5 L. plantarum 1A7 L. sanfranciscensis DE9 | DPPH: 81 3 | DPPH: 90 3 | Montemurro et al. [35] |
| Quinoa sprouted flour | Quinoa sourdough | L. rossiae LB5 L. plantarum 1A7 L. sanfranciscensis DE9 | DPPH: 94 3 | DPPH: 97 3 | Montemurro et al. [35] |
| Whole grain quinoa flour | Fermented quinoa whole grain flour | L. plantarum DSM2648 | DPPH (0 h): ≈ 29 3 DPPH (24 h): ≈ 28 3 DPPH (48 h): ≈ 30 3 DPPH (72 h): ≈ 27 3 | DPPH (0 h): ≈ 27 3 DPPH (24 h): ≈ 68 3 DPPH (48 h): ≈ 66 3 DPPH (72 h): ≈ 64 3 | Ayyash et al. [32] |
| Whole grain quinoa flour | Fermented quinoa whole grain flour | L. reuteri KX88177 | DPPH (0 h): ≈ 29 3 DPPH (24 h): ≈ 28 3 DPPH (48 h): ≈ 30 3 DPPH (72 h): ≈ 27 3 | DPPH (0 h): ≈ 32 3 DPPH (24 h): ≈ 75 3 DPPH (48 h): ≈ 58 3 DPPH (72 h): ≈ 73 3 | Ayyash et al. [32] |
| Whole grain quinoa flour | Fermented quinoa whole grain flour | L. plantarum KX881779 | DPPH (0 h): ≈ 29 3 DPPH (24 h): ≈ 28 3 DPPH (48 h): ≈ 30 3 DPPH (72 h): ≈ 27 3 | DPPH (0 h): ≈ 28 3 DPPH (24 h): ≈ 69 3 DPPH (48 h): ≈ 68 3 DPPH (72 h): ≈ 63 3 | Ayyash et al. [32] |
| Quinoa flour | Yogurt-like beverages | L. rhamnosus SP1 L. plantarum T6B10 W. confusa DSM 20194 | DPPH (B-SP1): 25 3 DPPH (B-T6B10):24 3 DPPH (B-DMS 20194): 29 3 | DPPH (B-SP1, after fermentation): 32 3 DPPH (B-T6B10, after fermentation): 37 3 DPPH (B-DMS 20194, after fermentation): 32 3 DPPH (B-SP1, after storage): 49 3 DPPH (B-T6B10, after storage): 44 3 DPPH (B-DMS 20194, after storage): 38 3 | Lorusso et al. [37] |
| Quinoa flour | Fermented quinoa-based beverages | Commercial probiotic culture including Bifidobacterium sp., Lactobacillus acidophilus, and Streptococcus thermophilus | DPPH: 60.31 3 | DPPH: 69.20 3 | Karovičová et al. [38] |
1: EC50 mg mL−1; 2: µmol Trolox g−1 dm; 3: %; 4: µmol Gallic Acid Equivalent (GAE) 100 g−1 dm; 5: IC50 mg mL−1; n.d.: not detected.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Melini, F.; Melini, V. Impact of Fermentation on Phenolic Compounds and Antioxidant Capacity of Quinoa. Fermentation 2021, 7, 20. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010020
AMA Style
Melini F, Melini V. Impact of Fermentation on Phenolic Compounds and Antioxidant Capacity of Quinoa. Fermentation. 2021; 7(1):20. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010020
Chicago/Turabian StyleMelini, Francesca, and Valentina Melini. 2021. "Impact of Fermentation on Phenolic Compounds and Antioxidant Capacity of Quinoa" Fermentation 7, no. 1: 20. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010020
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.