
Influence of Nitrogen Sources on D-Lactic Acid Biosynthesis by Sporolactobacillus laevolacticus DSM 442 Strain

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Microorganism
2.3. Culture Conditions
2.4. Enzymatic Hydrolysis of Waste, Agricultural and Food Raw Materials
2.5. Influence of Various Nitrogen Sources on D-LA Biosynthesis
2.6. Influence of Pea Seeds Hydrolysate and Unhydrolysed Pea Seeds on D-LA Biosynthesis
2.7. Batch Fermentation
2.8. Fed-Batch Fermentation
2.9. Analysis of Biosynthesis Products
3. Results
3.1. Effects of Various Nitrogen Sources on D-LA Biosynthesis
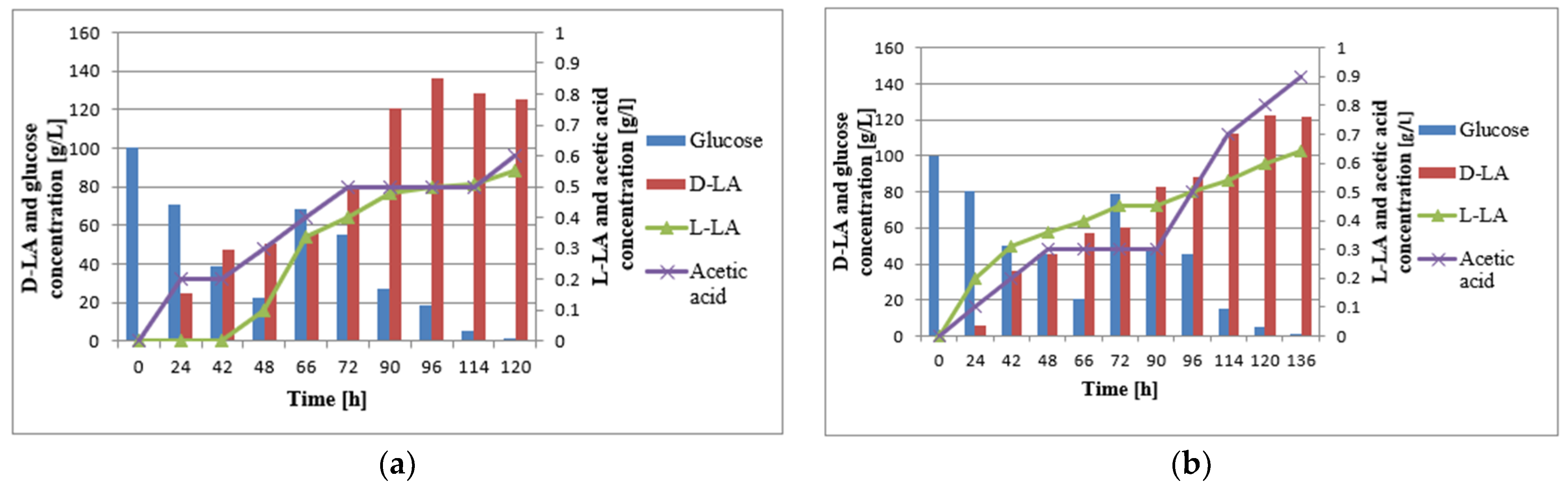
3.2. Batch Fermentation
3.3. Fed-Batch Fermentation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sawai, H.; Na, K.; Sasaki, N.; Mimitsuka, T.; Minegishi, S.; Yamada, S.; Shimizu, M.K.; Yonehara, T. Membrane-integrated fermentation system for improving the optical purity of D-lactic acid produced during continuous fermentation. Biosci. Biotechnol. Biochem. 2011, 75, 2326–2332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demirci, A.; Pometto, A. Enhanced production of D(-)-lactic acid by mutants of Lactobacillus delbrueckii ATCC 964. J. Ind. Microbiol. 1992, 11, 23–28. [Google Scholar] [CrossRef]
- Zheng, H.; Gong, J.; Chen, T.; Chen, X.; Zhao, X. Strain improvement of Sporolactobacillus inulinus ATCC 15538 for acid tolerance and production of D-lactic acid by genome shuffling. Appl. Microbiol. Biotechnol. 2010, 85, 1541–1549. [Google Scholar] [CrossRef]
- Michalczyk, A.; Cieniecka-Rosłonkiewicz, A.; Garbaczewska, S.; Morytz, B.; Białek, A. Badania nad wykorzystaniem biomasy skrobiowej jako źródła węgla w procesie fermentacji D-mleczanowej metodą SHF i SSF. Przem. Chem. 2020, 99, 99–102. [Google Scholar] [CrossRef]
- Smerilli, M.; Neureiter, M.; Wurz, S.; Haas, C.; Frühauf, S.; Fuchs, W. Direct fermentation of potato starch and potato residues to lactic acid by Geobacillus stearothermophilus under non-sterile conditions. J. Chem. Technol. Biotechnol. 2015, 90, 648–657. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, G.; Yu, X.; Chen, H.; Sun, Y.; Chen, G. Pretreatment of corn stover by solid acid for D-lactic acid fermentation. Bioresour. Technol. 2017, 239, 490–495. [Google Scholar] [CrossRef]
- Grosse, C.; Grigsby, W.J.; Noël, M.; Treu, A.; Thévenon, M.F.; Gérardin, P. Optimizing chemical wood modification with oligomeric lactic acid by screening of processing conditions. J. Wood Chem. Technol. 2019, 39, 385–398. [Google Scholar] [CrossRef]
- Yang, S.; Yu, H.; You, Y.; Li, X.; Jiang, J. Effective lactic acid production from waste paper using Streptococcus thermophilus at low enzyme loading assisted by Gleditsia saponin. Carbohyd Polym. 2018, 200, 122–127. [Google Scholar] [CrossRef]
- Vidra, A.; Tóth, A.J.; Németh, Á. Lactic acid production from cane molasses. Liquid Waste Recovery 2017, 2, 13–16. [Google Scholar] [CrossRef] [Green Version]
- Ghasemi, M.; Najafpour, G.; Rahimnejad, M.; Beigi, P.A.; Sedighi, M.; Hashemiyeh, B. Effect of different media on production of lactic acid from whey by Lactobacillus bulgaricus. Afr. J. Biotechnol. 2009, 8, 81–84. [Google Scholar] [CrossRef]
- Ikada, Y.; Jamshidi, K.; Tsuji, H.; Hyon, S.H. Stereocomplex formation between enantiomeric poly(lactides). Macromolecules 1987, 20, 904–906. [Google Scholar] [CrossRef]
- Fukushima, K.; Chang, Y.-H.; Kimura, Y. Enhanced stereocomplex formation of poly(L-lactic acid) and poly(D-lactic acid) in the presence of stereoblock poly(lactic acid). Macromol. Biosci. 2007, 7, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Mehta, R.; Kumar, V.; Bhunia, H.; Upadhyay, S.N. Synthesis of Poly(Lactic Acid): A Review. J. Macromol. Sci. Polym. Rev. 2005, 45, 325–349. [Google Scholar] [CrossRef]
- Reichert, C.L.; Bugnicourt, E.; Coltelli, M.-B.; Cinelli, P.; Lazzeri, A.; Canesi, I.; Braca, F.; Martínez, B.M.; Alonso, R.; Agostinis, L.; et al. Bio-based packaging: Materials, modifications, industrial applications and sustainability. Polymers 2020, 12, 1558. [Google Scholar] [CrossRef] [PubMed]
- Altaf, M.; Naveena, B.J.; Reddy, G. Use of inexpensive nitrogen sources and starch for L-lactic acid production in anaerobic submerged fermentation. Bioresour. Technol. 2007, 98, 498–503. [Google Scholar] [CrossRef]
- Nancib, A.; Nancib, N.; Cherif, D.M.; Boubendir, A.; Fick, M.; Boudrant, J. Joint effect of nitrogen sources and B vitamin supplementation of date juice on lactic acid production by Lactobacillus casei subsp. rhamnosus. Bioresour. Technol. 2005, 96, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Aspmo, S.I.; Horn, S.J.; Eijsink, V.G.H. Use of hydrolysates from Atlantic cod (Gadus morhua L.) viscera as a complex nitrogen source for lactic acid bacteria. FEMS Microbiol. Lett. 2005, 248, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Safari, R.; Motamedzadegan, A.; Ovissipour, M.; Regenstein, J.M.; Gildberg, A.; Rasco, B. Use of hydrolysates from yellow fin tuna (Thunnus albacares) heads as a complex nitrogen source for lactic acid bacteria. Food Bioprocess Technol. 2012, 5, 73–79. [Google Scholar] [CrossRef]
- Hyun, C.K.; Shin, H.K. Utilization of animal blood proteins as nitrogen sources for the cultivation of lactic acid bacteria. Korean J. Appl. Microbiol. Biotechnol. 1997, 25, 218–223. [Google Scholar]
- Paulova, L.; Chmelik, J.; Branska, B.; Patakova, P.; Drahokoupil, M.; Melzoch, K. Comparison of lactic acid production by L. casei in batch, fed-batch and continuous cultivation, testing the use of feather hydrolysate as a complex nitrogen source. Braz. Arch. Biol. Technol. 2020, 63, e20190151. [Google Scholar] [CrossRef]
- Kwon, S.; Lee, P.C.; Lee, E.G.; Chang, Y.K.; Chang, N. Production of lactic acid by Lactobacillus rhamnosus with vitamin supplemented soybean hydrolysate. Enzyme Microb. Technol. 2000, 26, 209–215. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, B.; Li, F.; Xu, K.; Ma, C.; Tao, F.; Li, Q.; Xu, P. Highly efficient production of D-lactate by Sporolactobacillus sp. CASD with simultaneous enzymatic hydrolysis of peanut meal. Appl. Microbiol. Biotechnol. 2011, 89, 1009–1017. [Google Scholar] [CrossRef]
- Hujanen, M.; Linko, Y.Y. Effect of temperature and various nitrogen sources on L(+)-lactic acid production by Lactobacillus casei. Appl. Microbiol. Biotechnol. 1996, 3, 307–313. [Google Scholar] [CrossRef]
- Altaf, M.; Naveena, B.J.; Ready, G. Screening of inexpensive nitrogen sources for production of L(+) lactic acid from starch by amylolytic Lactobacillus amylophilus GV6 in single step fermentation, Food Technol. Biotechnol. 2005, 43, 235–239. [Google Scholar] [CrossRef]
- Li, Y.; Wang, L.; Ju, J.; Yu, B.; Ma, Y. Efficient production of polymer-grade D-lactate by Sporolactobacillus laevolacticus DSM 442 with agricultural waste cottonseed as the sole nitrogen source. Bioresour. Technol. 2013, 142, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.; Gao, Z.; Sun, J.; Wu, B.; He, B. D-Lactic acid production by Sporolactobacillus inulinus YBS1-5 with simultaneous utilization of cottonseed meal and corncob residue. Bioresour. Technol. 2016, 207, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.J.; Jiang, A.L.; Mao, Y.Q.; Bo, W.; He, M.-X.; Hu, W.; Chen, J.-H.; Li, W.-J. Efficient L-lactic acid production from purified sweet sorghum juice coupled with soybean hydrolysate as nitrogen source by Lactobacillus thermophilus A69 strain. J. Chem Technol. Biotechnol. 2019, 94, 1752–1759. [Google Scholar] [CrossRef]
- Wee, Y.J.; Reddy, L.V.A.; Ryu, H.W. Fermentative production of L (+)-lactic acid from starch hydrolyzate and corn steep liquor as inexpensive nutrients by batch culture of Enterococcus faecalis RKY1. J. Chem. Technol. Biotechnol. 2008, 83, 1387–1393. [Google Scholar] [CrossRef]
- Yu, L.; Lei, T.; Ren, X.; Pei, X.; Feng, Y. Response surface optimization of l-(+)-lactic acid production using corn steep liquor as an alternative nitrogen source by Lactobacillus rhamnosus CGMCC 1466. Biochem. Eng. J. 2008, 39, 496–502. [Google Scholar] [CrossRef]
- Liu, B.; Yang, M.; Qi, B.; Chen, X.; Su, Z.; Wan, Y. Optimizing l-(+)-lactic acid production by thermophile Lactobacillus plantarum As.1.3 using alternative nitrogen sources with response surface method. Biochem. Eng. J. 2010, 52, 212–219. [Google Scholar] [CrossRef]
- De la Torre, I.; Ladero, M.; Santos, V.E. Production of d-lactic acid by Lactobacillus delbrueckii ssp. delbrueckii from orange peel waste: Techno-economical assessment of nitrogen sources. Appl. Microbiol. Biotechnol. 2018, 102, 10511–10521. [Google Scholar] [CrossRef]
- Lund, B.; Norddahl, B.; Ahring, B. Production of lactic acid from whey using hydrolysed whey protein as nitrogen source. Biotechnol. Lett. 1992, 14, 851–856. [Google Scholar] [CrossRef]
- Amrane, A.; Prigent, Y. Lactic acid production from lactose in batch culture: Analysis of the data with the help of a mathematical model; relevance for nitrogen source and preculture assessment. Appl. Microbiol. Biotechnol. 1994, 40, 644–649. [Google Scholar] [CrossRef]
- Nancib, N.; Nancib, A.; Boudjelal, A.; Benslimane, C.; Blanchard, F.; Boudrant, J. The effect of supplementation by different nitrogen sources on the production of lactic acid from date juice by Lactobacillus casei subsp. rhamnosus. Bioresour. Technol. 2001, 78, 149–153. [Google Scholar] [CrossRef]
- Clifford, C.E.; Scott, V.B.; Alan, B.K. A Powerful Kjeldahl nitrogen method using peroxymonosulfuric acid. J. Agric. Food Chem. 1985, 33, 1117–1123. [Google Scholar]
- Man, J.C.; Rogosa, M.; Sharpe, M.E. A medium for the cultivation of Lactobacill. J. Appl. Bacteriol. 1960, 23, 130–135. [Google Scholar] [CrossRef]
- Okano, K.; Hama, S.; Kihara, M.; Noda, H.; Tanaka, T.; Kondo, A. Production of optically pure D-lactic acid from brown rice using metabolically engineered Lactobacillus plantarum. Appl Microbiol Biotechnol. 2017, 101, 1869–1875. [Google Scholar] [CrossRef]
- Sreenath, H.K.; Moldes, A.B.; Koegel, R.G.; Straub, R.J. Lactic acid production from agriculture residues. Biotechnol. Lett. 2001, 23, 179–184. [Google Scholar] [CrossRef]
- Moldes, A.B.; Torrado, A.; Converti, A.; Domínguez, J.M. Complete bioconversion of hemicellulosic sugars from agricultural residues into lactic acid by Lactobacillus pentosus. Biotechnol. Appl. Biochem. 2006, 135, 219–227. [Google Scholar] [CrossRef]
- Wee, Y.J.; Kim, J.N.; Ryu, H.W. Biotechnological production of lactic acid and its recent applications. Food Technol. Biotechnol. 2006, 44, 163–172. [Google Scholar]
- Schepers, A.W.; Thibault, J.; Lacroix, C. Lactobacillus helveticus growth and lactic acid production during pH-controlled batch cultures in whey permeate/yeast extract medium. Part I. multiplefactor kinetic analysis. Enzyme Microb. Technol. 2002, 30, 176–186. [Google Scholar] [CrossRef]
- Hsieh, C.M.; Yang, F.C.; Iannott, E.L. The effect of soy protein hydrolyzates on fermentation by Lactobacillus amylovorus. Process Biochem. 1999, 34, 173–179. [Google Scholar] [CrossRef]
- Ishida, N.; Suzuki, T.; Tokuhiro, K.; Nagamori, E.; Onishi, T.; Saitoh, S.; Kitamoto, K.; Takahashi, H. D-Lactic acid production by metabolically engineered Saccharomyces cerevisiae. J. Biosci. Bioeng. 2006, 101, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Benthin, S.; Villadsen, J. Production of optically pure D-lactate by Lactobacillus bulgaricus and purification by crystallization and liquid/liquid extraction. Appl. Microbiol. Biotechnol. 1995, 42, 826–829. [Google Scholar] [CrossRef]
- Okino, S.; Suda, M.; Fujikura, K.; Inui, M.; Yukawa, H. Production of D-lactic acid by Corynebacterium glutamicumunder oxygen deprivation. Biotechnol. Bioprocess Eng. 2008, 78, 449–454. [Google Scholar] [CrossRef]
- Zhang, Y.; Praveen, V.D. Lactic acid biosynthesis from biomass-derived sugars via Lactobacillus delbrueckii fermentation. Biotechnol. Bioprocess Eng. 2013, 36, 1897–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mimitsuka, T.; Na, K.; Morita, K.; Sawai, H.; Minegishi, S.; Henmi, M.; Yamada, K.; Shimizu, S.; Yonehara, T. A membrane-integrated fermentation reactor system: Its effects in reducing the amount of sub-raw materials for D-lactic acid continuous fermentation by Sporolactobacillus laevolacticus. Biosci. Biotechnol. Biochem. 2012, 76, 67–72. [Google Scholar] [CrossRef] [Green Version]
- De Boer, J.P.; De Mattos, M.J.; Neijsseld, O.M. D(-)Lactic acid production by suspended and aggregated continuous cultures of Bacillus laevolacticus. Biosci. Biotechnol. Biochem. 1990, 34, 149–153. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type of Raw Material | Total Nitrogen Content (%) (w/w) | Quantity of Substrate Used in the Production Medium (g/L) |
|---|---|---|
| (NH4)2SO4 | 21 a | 2.4 |
| NH4NO3 | 35 a | 1.4 |
| Urea | 47 a | 1.1 |
| Yeast extract | 10 b | 5.0 |
| Meat extract | 10 b | 5.0 |
| Peptone Bacteriological | 12 b | 4.2 |
| Casein tryptone | 10 b | 5.0 |
| Yeast cake | 8.5 c | 5.9 |
| Corn steep liquor | 1.9 c | 27.0 |
| Baker’s yeast | 2.7 c | 18.5 |
| Wheat middling | 10 c | 5.0 |
| Hazelnuts | 4.1 c | 12.3 |
| Pumpkin seeds | 5.5 c | 9.0 |
| Flax seeds | 3.2 c | 15.6 |
| Sunflower seeds | 2.3 c | 21.7 |
| Alfalfa seeds | 9.1 c | 5.5 |
| Lupine seeds | 6.5 c | 7.8 |
| Pea seeds | 4.1 c | 12.3 |
| Soybean seeds | 8.1 c | 6.2 |
| Source of Nitrogen | Optimal Fermentation Time [h] | Unreacted Glucose Content [g/L] (a) | D-LA Concentration [g/L] (a) | D-LA Yield [%] (b) |
|---|---|---|---|---|
| (NH4)2SO4 | 96 | 63.4, 0.26 | 33.0, 0.56 | 33 |
| NH4NO3 | 96 | 89.3, 0.59 | 8.9, 0.22 | 9 |
| Yeast extract | 96 | 12.8, 1.55 | 83.6, 2.65 | 84 |
| Urea | 120 | 79.4, 0.69 | 17.9, 1.29 | 18 |
| Meat extract | 96 | 62.8, 1.09 | 34.6, 2.09 | 35 |
| Peptone Bacteriological | 96 | 55.3, 1,45 | 36.0, 0.09 | 36 |
| Casein tryptone | 96 | 54.8, 1.22 | 42.9, 0.45 | 43 |
| Yeast cake | 168 | 45.2, 1.25 | 51.3, 0.78 | 51 |
| Corn steep liquor | 168 | 60.2, 1.65 | 36.9, 0.09 | 37 |
| Baker’s yeast | 168 | 50.2, 1.59 | 46.2, 2.51 | 46 |
| Wheat middling | 144 | 52.8, 1.22 | 44.1, 1.65 | 44 |
| Hazelnuts | 120 | 60.7, 1.68 | 34.8, 1.89 | 35 |
| Pumpkin seeds | 120 | 60.5, 2.09 | 35.5, 0.99 | 36 |
| Flax seeds | 120 | 66.2, 1.98 | 30.1, 0.89 | 30 |
| Sunflower seeds | 120 | 59.4, 0.98 | 37.6, 0.78 | 38 |
| Alfalfa seeds | 120 | 36.2, 0.88 | 61.0, 1.89 | 61 |
| Lupine seeds | 120 | 21.20, 0.5 | 74.4, 0.89 | 74 |
| Pea seeds | 120 | 15.3, 0.23 | 81.5, 2.90 | 82 |
| Soybean seeds | 120 | 34.9, 0.89 | 63.6, 3.09 | 64 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michalczyk, A.K.; Garbaczewska, S.; Morytz, B.; Białek, A.; Zakrzewski, J. Influence of Nitrogen Sources on D-Lactic Acid Biosynthesis by Sporolactobacillus laevolacticus DSM 442 Strain. Fermentation 2021, 7, 78. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7020078
Michalczyk AK, Garbaczewska S, Morytz B, Białek A, Zakrzewski J. Influence of Nitrogen Sources on D-Lactic Acid Biosynthesis by Sporolactobacillus laevolacticus DSM 442 Strain. Fermentation. 2021; 7(2):78. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7020078
Chicago/Turabian StyleMichalczyk, Alicja Katarzyna, Sylwia Garbaczewska, Bolesław Morytz, Arkadiusz Białek, and Jerzy Zakrzewski. 2021. "Influence of Nitrogen Sources on D-Lactic Acid Biosynthesis by Sporolactobacillus laevolacticus DSM 442 Strain" Fermentation 7, no. 2: 78. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7020078