
Use of Kluyveromyces marxianus to Increase Free Monoterpenes and Aliphatic Esters in White Wines

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Technological Screening of Strain Km L2009
2.2.1. Killer Activity Assay
2.2.2. Sulfite Tolerance Assay
2.2.3. Alcohol Tolerance Assay
2.3. Laboratory-Scale Fermentations
2.4. Wine Production and Monitoring
2.4.1. Vinification
2.4.2. Microbiological Analyses
2.4.3. Molecular Analyses
2.4.4. Chemical Analyses
2.4.5. Volatile Compound Analyses
2.4.6. Sensory Analyses
2.4.7. Statistical and Explorative Multivariate Analyses
3. Results and Discussion
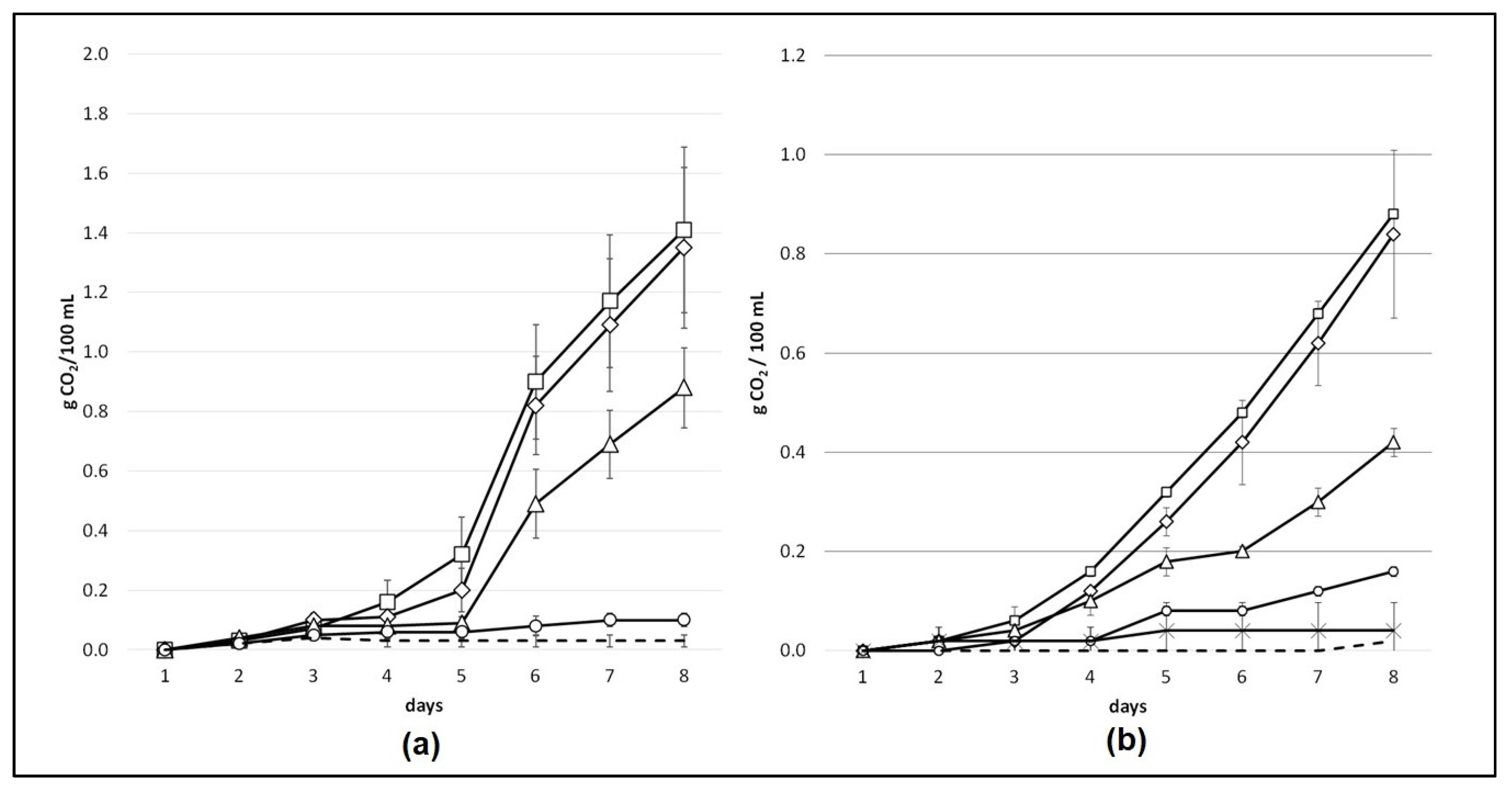
3.1. Technological Screening of Strain Km L2009
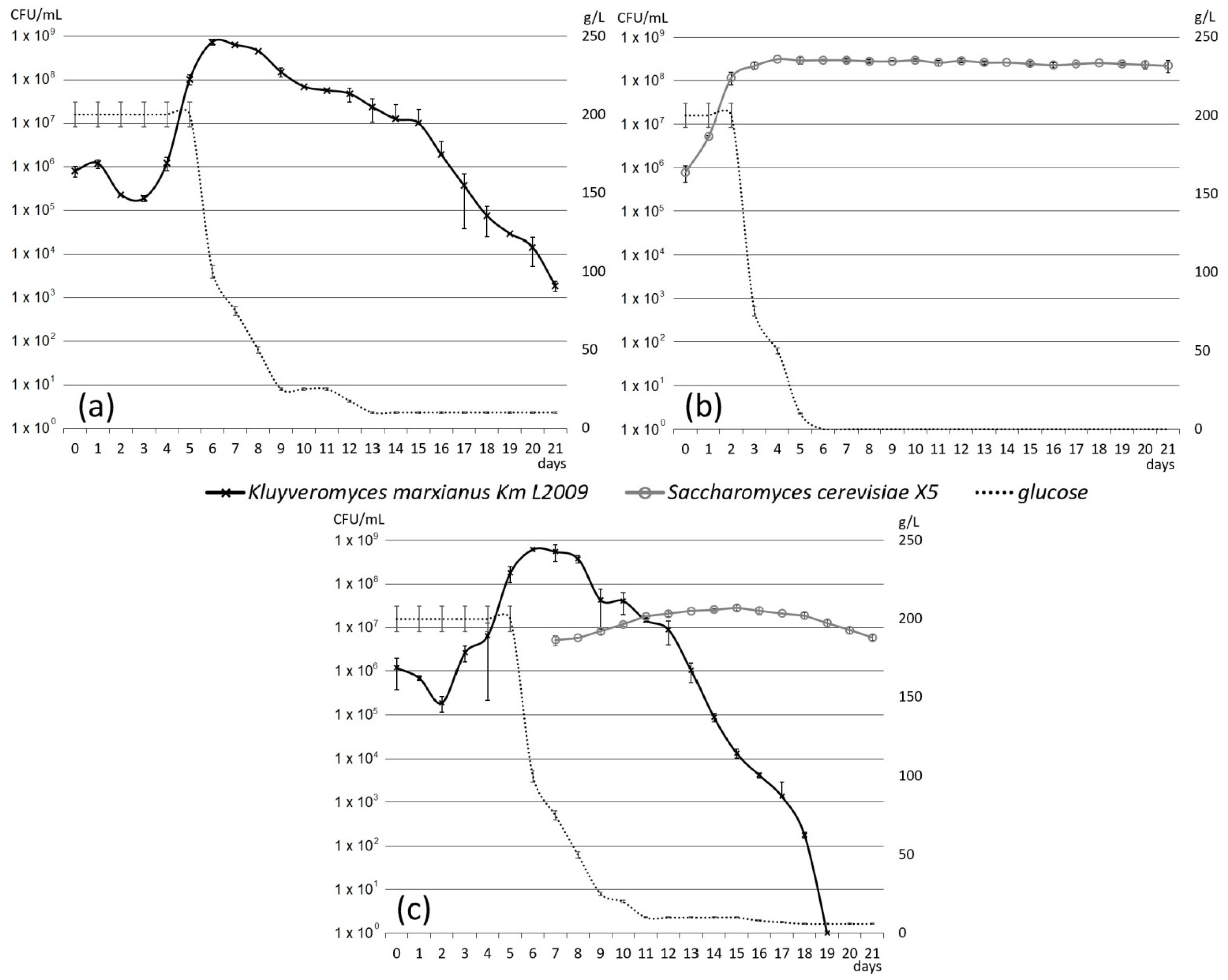
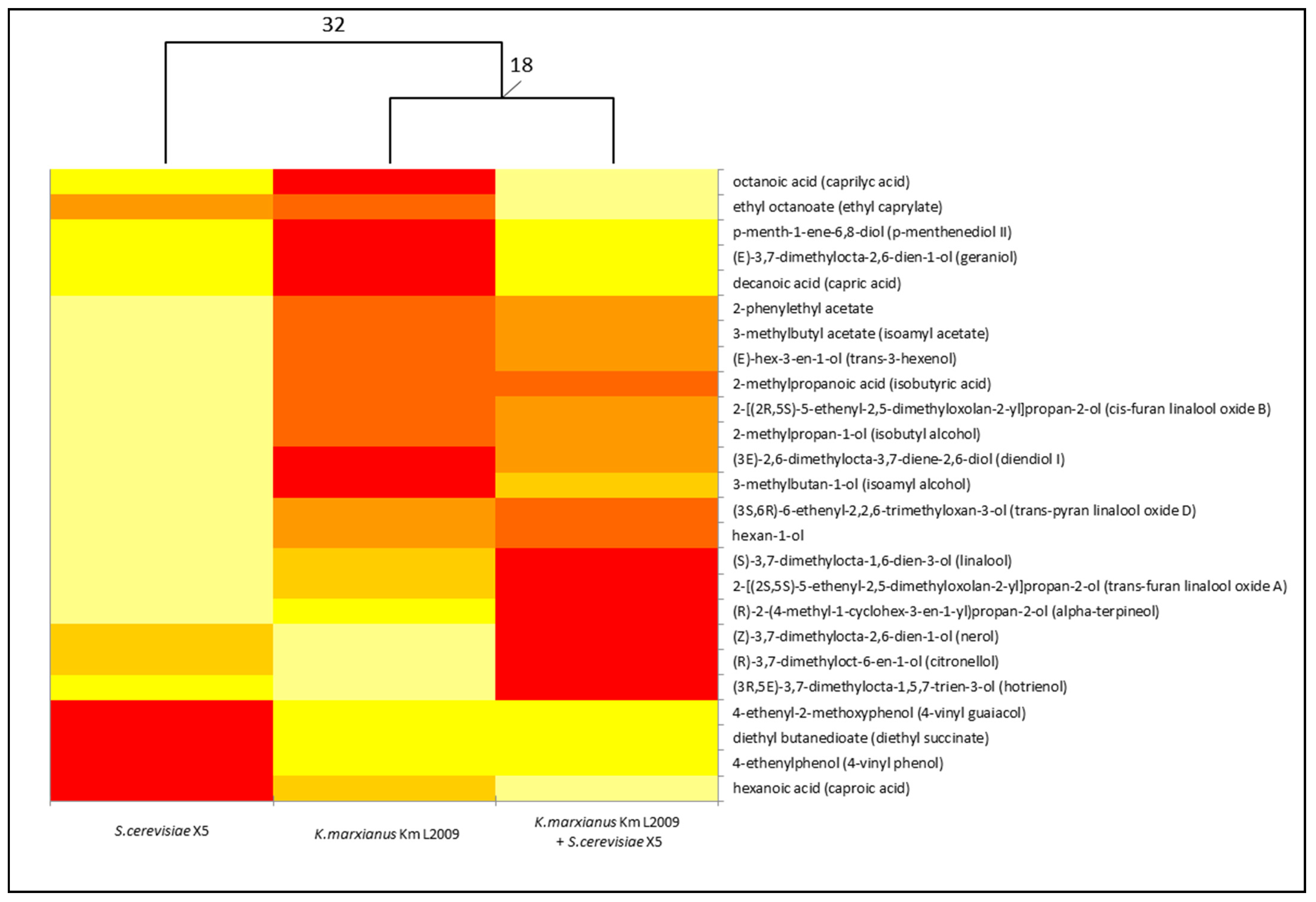
3.2. Laboratory-Scale Fermentations
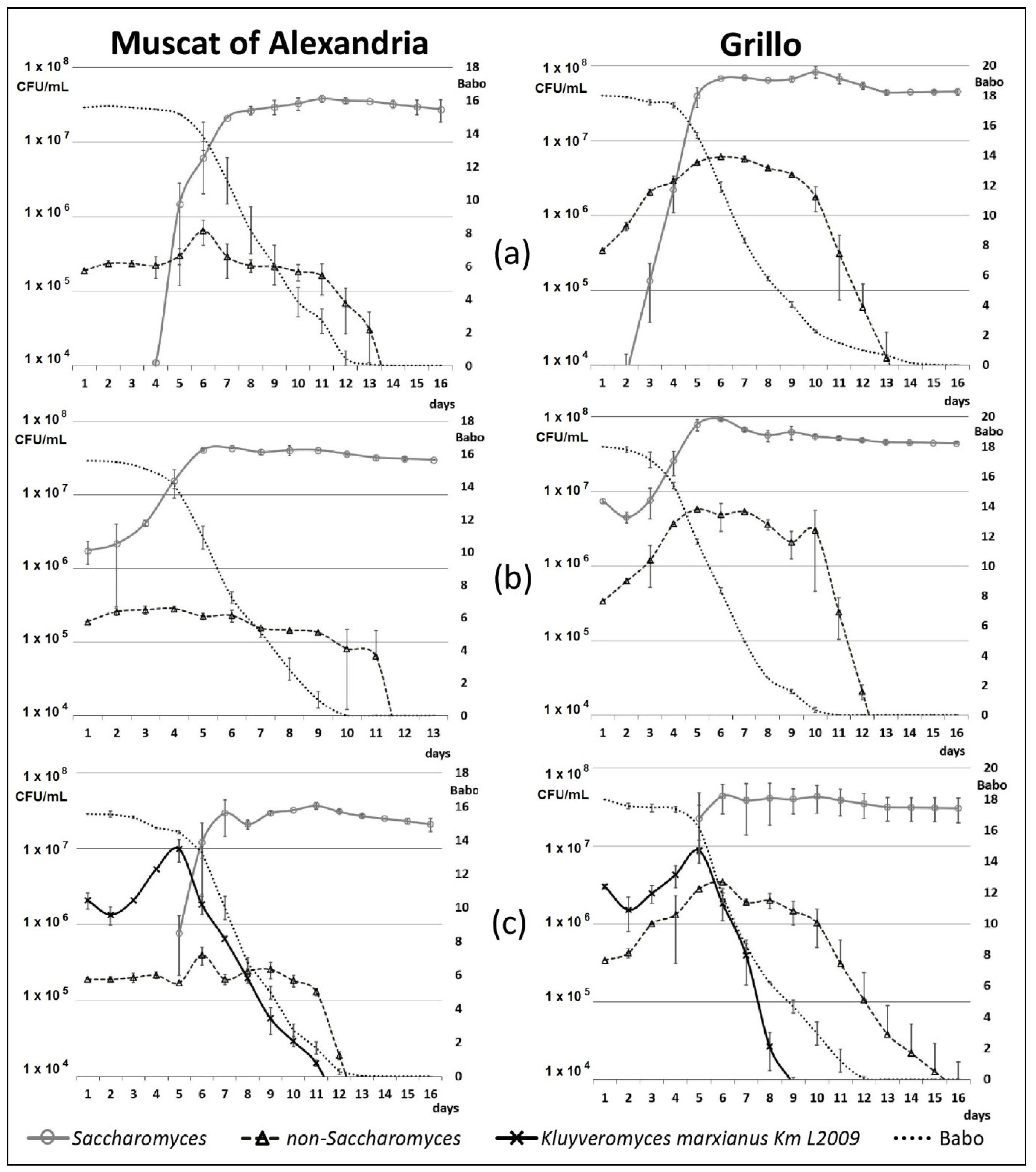
3.3. Wine Production and Monitoring
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aranda, A. Enological Repercussions of Non-Saccharomyces Species. Fermentation 2019, 5, 68. [Google Scholar] [CrossRef] [Green Version]
- Morata, A. Enological Repercussions of Non-Saccharomyces Species in Wine Biotechnology. Fermentation 2019, 5, 72. [Google Scholar] [CrossRef] [Green Version]
- Morata, A. Enological Repercussions of Non-Saccharomyces Species 2.0. Fermentation 2020, 6, 110. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, C.; Borneman, A.R. Yeasts found in vineyards and wineries. Yeast 2016, 34, 111–128. [Google Scholar] [CrossRef]
- Mateo, J.J.; Maicas, S. Application of non-Saccharomyces yeasts to wine-making process. Fermentation 2016, 2, 14. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F. Non-Saccharomyces wine yeasts have a promising role in biotechnological approaches to winemaking. Ann. Microbiol. 2011, 61, 25–32. [Google Scholar] [CrossRef]
- Romani, C.; Lencioni, L.; Biondi Bartolini, A.; Ciani, M.; Mannazzu, I.; Domizio, P. Pilot Scale Fermentations of Sangiovese: An Overview on the Impact of Saccharomyces and Non-Saccharomyces Wine Yeasts. Fermentation 2020, 6, 63. [Google Scholar] [CrossRef]
- Giaramida, P.; Ponticello, G.; Di Maio, S.; Squadrito, M.; Genna, G.; Barone, E.; Scacco, A.; Corona, O.; Amore, G.; Di Stefano, R.; et al. Candida zemplinina for production of wines with less alcohol and more glycerol. S. Afr. J. Enol. Vitic. 2013, 34, 204–211. [Google Scholar] [CrossRef] [Green Version]
- Lodder, J.; Kreger-van Rij, N.J.W. The Yeasts: A Taxonomic Study; North-Holland Publishing, Co.: Amsterdam, The Netherlands, 1952; pp. 155–158. [Google Scholar]
- Lachance, M.A. Kluyveromyces van der Walt (1971). In The Yeasts, A Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 471–482. [Google Scholar]
- Fonseca, G.G.; Heinzle, E.; Wittmann, C.; Gombert, A.K. The yeast Kluyveromyces marxianus and its biotechnological potential. Appl. Microbiol. Biotechnol. 2008, 79, 339–354. [Google Scholar] [CrossRef] [PubMed]
- Karim, A.; Gerliani, N.; Aïder, M. Kluyveromyces marxianus: An emerging yeast cell factory for applications in food and biotechnology. Int. J. Food Microbiol. 2020, 333, 108818. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, J.P.; Etschmann, M.M.W.; Schrader, J.; de Billerbeck, G.M. Cell factory applications of the yeast Kluyveromyces marxianus for the biotechnological production of natural flavour and fragrance molecules. Yeast 2015, 32, 3–16. [Google Scholar] [CrossRef]
- Reyes-Sánchez, F.; Páez-Lerma, J.B.; Rojas-Contreras, J.A.; López-Miranda, J.; Soto-Cruz, N.Ó.; Reinhart-Kirchmayr, M.R. Study of the Enzymatic Capacity of Kluyveromyces marxianus for the Synthesis of Esters. J. Mol. Microbiol. Biotechnol. 2019, 29, 1–9. [Google Scholar] [CrossRef]
- Papon, N.; Courdavault, V.; Clastre, M.; Bennett, R.J. Emerging and emerged pathogenic Candida species: Beyond the Candida albicans paradigm. PLoS Pathog. 2013, 9, e1003550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Food Safety Authority. Scientific opinion on the update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA (2017–2019). EFSA J. 2020, 18, e05966. [Google Scholar] [CrossRef] [Green Version]
- Piemolini-Barreto, L.T.; Zacaria, J.; Delamare, A.P.; Antonio, R.V.; Echeverrigaray, S. Variation in phenolic compounds, anthocyanins, and color in red wine treated with enzymatic extract of Kluyveromyces marxianus. World J. Microbiol. Biotechnol. 2014, 30, 1541–1547. [Google Scholar] [CrossRef]
- Piemolini-Barreto, L.T.; Antonio, R.V.; Echeverrigaray, S. Comparison of a pectinoltytic extract of Kluyveromyces marxianus and a commercial enzyme preparation in the production of Ives (Vitis labrusca) grape juice. World J. Microbiol. Biotechnol. 2015, 31, 755–762. [Google Scholar] [CrossRef]
- Sieiro, C.; Villa, T.G.; da Silva, A.F.; García-Fraga, B.; Vilanova, M. Albariño wine aroma enhancement through the use of a recombinant polygalacturonase from Kluyveromyces marxianus. Food Chem. 2014, 145, 179–185. [Google Scholar] [CrossRef]
- Kourkoutas, Y.; McErlean, C.; Kanellaki, M.; Hack, C.J.; Marchant, R.; Banat, I.M.; Koutinas, M.M. High-temperature wine making using the thermotolerant yeast strain Kluyveromyces marxianus IMB3. Appl. Biochem. Biotechnol. 2004, 112, 25–35. [Google Scholar] [CrossRef]
- Vigentini, I.; Maghradze, D.; Petrozziello, M.; Bonello, F.; Mezzapelle, V.; Valdetara, F.; Failla, O.; Foschino, R. Indigenous Georgian wine associated-yeasts and grape cultivars to edit the wine quality in a precision oenology perspective. Front. Microbiol. 2016, 7, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rollero, S.; Zietsman, A.J.J.; Buffetto, F.; Schückel, J.; Ortiz-Julien, A.; Divol, B. Kluyveromyces marxianus Secretes a Pectinase in Shiraz Grape Must That Impacts Technological Properties and Aroma Profile of Wine. J. Agric. Food Chem. 2018, 66, 11739–11747. [Google Scholar] [CrossRef] [PubMed]
- Polizzotto, G.; Barone, E.; Ponticello, G.; Fasciana, T.; Barbera, D.; Corona, O.; Amore, G.; Giammanco, A.; Oliva, D. Isolation, identification and oenological characterization of non-Saccharomyces yeasts in a Mediterranean island. Lett. Appl. Microbiol. 2016, 63, 131–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maicas, S.; Mateo, J.J. Hydrolysis of terpenyl glycosides in grape juice and other fruit juices: A review. Appl. Microbiol. Biotechnol. 2005, 67, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Regodón, J.A.; Peréz, F.; Valdés, M.E.; De Miguel, C.; Ramìrez, M. A simple and effective procedure for selection of wine yeast strains. Food Microbiol. 1997, 14, 247–254. [Google Scholar] [CrossRef]
- Caridi, A.; Cufari, A.; Ramondino, D. Isolation and clonal pre-selection of enological Saccharomyces. J. Gen. Appl. Microbiol. 2002, 48, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Gump, B.H.; Zoecklein, B.W.; Fugelsang, K.C.; Whiton, R.S. Comparison of analytical methods for prediction of prefermentation nutritional status of grape juice. Am. J. Enol. Vitic. 2002, 53, 325–329. [Google Scholar]
- Ribérau-Gayon, P.; Glories, Y.; Maujean, A.; Dubourdieu, D. Handbook of Enology. Volume 2. The Chemistry of Wine: Stabilization and Treatments, 2nd ed.; John Wiley & Sons Ltd.: Chichester, UK, 2006; pp. 206–209, 301–331. [Google Scholar]
- Hall, J.F. Detection of wild yeasts in the brewery. J. Inst. Brew. 1971, 77, 513–516. [Google Scholar] [CrossRef]
- Pallmann, C.L.; Brown, J.A.; Olineka, T.L.; Cocolin, L.; Mills, D.A.; Bisson, L.F. Use of WL Medium to profile native flora fermentations. Am. J. Enol. Vitic. 2001, 52, 198–203. Available online: https://www.ajevonline.org/content/52/3/198 (accessed on 15 May 2021).
- Romancino, D.; Di Maio, S.; Muriella, R.; Oliva, D. Analysis of non Saccharomyces yeast populations isolated from grape must from Sicily (Italy). J. Appl. Microbiol. 2008, 105, 2248–2254. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Liu, A.; Xue, B.; Liu, Y. Yeast species associated with spontaneous wine fermentation of Cabernet Sauvignon from Ningxia, China. World J. Microbiol. Biotechnol. 2011, 27, 2475–2482. [Google Scholar] [CrossRef]
- Fowell, R.R. The identification of wild yeast colonies on Lysine Agar. J. Appl. Bacteriol. 1965, 28, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Di Maio, S.; Polizzotto, G.; Planeta, D.; Oliva, D. A method to discriminate between the Candida stellata and Saccharomyces cerevisiae in mixed fermentation on WLD and Lysine Agar Media. S. Afr. J. Enol. Vitic. 2011, 32, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Belloch, C.; Barrio, E.; Uruburu, F.; Garcia, M.D.; Querol, A. Characterisation of four species of the genus Kluyveromyces by mitochondrial DNA restriction analysis. System. Appl. Microbiol. 1997, 20, 397–408. [Google Scholar] [CrossRef]
- Querol, A.; Barrio, E.; Huerta, T.; Ramón, D. Molecular monitoring of wine fermentations conducted by active dry yeast strains. Appl. Envir. Microbiol. 1992, 58, 2948–2953. [Google Scholar] [CrossRef] [Green Version]
- Di Maio, S.; Polizzotto, G.; Di Gangi, E.; Foresta, G.; Genna, G.; Verzera, A.; Scacco, A.; Amore, G.; Oliva, D. Biodiversity of indigenous Saccharomyces populations from old wineries of south-eastern Sicily (Italy): Preservation and economic potential. PLoS ONE 2012, 7, e30428. [Google Scholar]
- The International Organisation of Vine and Wine: Compendium of International Methods of Analysis of Wines and Musts. Available online: http://www.oiv.int/en/technical-standards-and-documents/methods-of-analysis/compendium-of-international-methods-of-analysis-of-wines-and-musts-2-vol (accessed on 3 March 2021).
- Di Stefano, R.; Cravero, M.C.; Gentilini, N. Metodi per lo studio dei polifenoli dei vini. L’Enotecnico 1989, 25, 83–89. [Google Scholar]
- Corona, O. Wine-making with protection of must against oxidation in a warm semi-arid terroir. S. Afr. J. Enol. Vitic. 2010, 31, 58–63. [Google Scholar] [CrossRef]
- Ilc, T.; Werck-Reichhart, D.; Navrot, N. Meta-Analysis of the Core Aroma Components of Grape and Wine Aroma. Front. Plant Sci. 2016, 7, 1472. [Google Scholar] [CrossRef] [Green Version]
- International Organization for Standardization, Geneva, Switzerland. UNI ISO 10399 (2010). Sensory Analysis—Methodology—Duo-Trio Test. Available online: https://www.iso.org/standard/74219.html (accessed on 3 March 2021).
- International Organization for Standardization, Geneva, Switzerland. UNI ISO 5495 (2005). Sensory Analysis—Methodology—Paired Comparison Test. Available online: https://www.iso.org/standard/31621.html (accessed on 5 May 2021).
- International Organization for Standardization, Geneva, Switzerland. UNI ISO 8589 (2007). Sensory Analysis—General Guidance for the Design of Test Rooms. Available online: https://www.iso.org/standard/36385.html (accessed on 3 March 2021).
- Roessler, E.B.; Pangborn, R.M.; Sidel, J.L.; Stone, H. Expanded statistical tables for estimation significance in paired-preference, paired-difference, duo-trio and triangle tests. J. Food Sci. 1978, 43, 940–943. [Google Scholar] [CrossRef]
- Martorana, A.; Alfonzo, A.; Gaglio, R.; Settanni, L.; Corona, O.; La Croce, F.; Vagnoli, P.; Caruso, T.; Moschetti, G.; Francesca, N. Evaluation of different conditions to enhance the performances of Lactobacillus pentosus OM13 during industrial production of Spanish-style table olives. Food Microbiol. 2017, 61, 150–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangorrín, M.P.; Zajonskovsky, I.E.; Lopes, C.A.; Rodríguez, M.E.; Giraudo de van Broock, M.R.; Caballero, A.C. Killer behaviour in wild wine yeasts associated with Merlot and Malbec type musts spontaneously fermented from northwestern Patagonia (Argentina). J. Basic Microbiol. 2001, 41, 105–113. [Google Scholar] [CrossRef]
- Borneman, A.R.; Forgan, A.H.; Kolouchova, R.; Fraser, J.A.; Schmidt, S.A. Whole Genome Comparison Reveals High Levels of Inbreeding and Strain Redundancy Across the Spectrum of Commercial Wine Strains of Saccharomyces cerevisiae. G3 Genes Genomes Genet. 2016, 6, 957–971. [Google Scholar] [CrossRef] [Green Version]
- Pozo-Bayón, M.Á.; Monagas, M.; Bartolomé, B.; Moreno-Arribas, M.V. Wine features related to safety and consumer health: An integrated perspective. Crit. Rev. Food Sci. Nutr. 2012, 52, 31–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hjelmeland, A.K.; Zweigenbaum, J.; Ebeler, S.E. Profiling monoterpenol glycoconjugation in Vitis vinifera L. cv. Muscat of Alexandria using a novel putative compound database approach, high resolution mass spectrometry and collision induced dissociation fragmentation analysis. Anal. Chim. Acta 2015, 887, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Hodson, G.; Wilkes, E.; Azevedo, S.; Battaglene, T. Methanol in wine. BIO Web Conf. 2017, 9, 02028. [Google Scholar] [CrossRef]
- Marais, J. Terpenes in the aroma of grapes and wines: A review. S. Afr. J. Enol. Vitic. 1983, 4, 49–58. [Google Scholar] [CrossRef]
- Bartowsky, E.J.; Pretorius, I.S. Microbial formation and modification of flavor and off-flavor compounds in wine. In Biology of Microorganisms on Grapes, in Must and in Wine; König, H., Unden, G., Fröhlich, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 209–231. [Google Scholar] [CrossRef]
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl. Microbiol. Biotechnol. 2016, 100, 9861–9874. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Boekhout, T.; Gojkovic, Z.; Katz, M. Evaluation of non-Saccharomyces yeasts in the fermentation of wine, beer and cider for the development of new beverages. J. Inst. Brew. 2018, 124, 389–402. [Google Scholar] [CrossRef]
- Gamero, A.; Dijkstra, A.; Smit, B.; de Jong, C. Aromatic Potential of Diverse Non-Conventional Yeast Species for Winemaking and Brewing. Fermentation 2020, 6, 50. [Google Scholar] [CrossRef]
- Belitz, H.D.; Grosch, W. Food Chemistry, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 857–859. [Google Scholar] [CrossRef]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its importance to wine aroma—A review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Mateo, J.J.; Jiménez, M. Monoterpenes in grape juice and wines. J. Chromatogr. A 2000, 881, 557–567. [Google Scholar] [CrossRef]
- Ferrari, G.; Lablanquie, O.; Cantagrel, R.; Ledauphin, J.; Payot, T.; Fournier, N.; Guichard, E. Determination of key odorant compounds in freshly distilled cognac using GC-O, GC-MS, and sensory evaluation. J. Agric. Food Chem. 2004, 52, 5670–5676. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Míguez, M.J.; Cacho, J.F.; Ferreira, V.; Vicario, I.M.; Heredia, F.J. Volatile components of Zalema white wines. Food Chem. 2007, 100, 1464–1473. [Google Scholar] [CrossRef]
- Dragone, G.; Mussatto, S.I.; Oliveira, J.M.; Teixeira, J.A. Characterization of volatile compounds in an alcoholic beverage produced by whey fermentation. Food Chem. 2009, 112, 929–935. [Google Scholar] [CrossRef] [Green Version]
- Duarte, W.F.; Dias, D.R.; Oliveira, J.M.; Teixeira, J.A.; de Almeida e Silva, J.B.; Schwan, R.F. Characterization of different fruit wines made from cacao, cupuassu, gabiroba, jaboticaba and umbu. LWT Food Sci. Technol. 2010, 43, 1564–1572. [Google Scholar] [CrossRef]
- Vilanova, M.; Genisheva, Z.; Masa, A.; Oliveira, J.M. Correlation between volatile composition and sensory properties in Spanish Albariño wines. Microchem. J. 2010, 95, 240–246. [Google Scholar] [CrossRef]
- Sànchez-Palomo, E.; García-Carpintero, E.G.; Gómez Gallego, M.Á.; Gonzàlez Viñas, M.Á. The aroma of rojal red wines from La Mancha region—Determination of key odorants. In Gas Chromatography in Plant Science, Wine Technology, Toxicology and Some Specific Applications; Salih, B., Çelikbıçak, Ö., Eds.; Intech: Rijeka, Croatia, 2012; pp. 147–170. [Google Scholar] [CrossRef]
- Fariña, L.; Villar, V.; Ares, G.; Carrau, F.; Dellacassa, E.; Boido, E. Volatile composition and aroma profile of Uruguayan Tannat wines. Food Res. Int. 2015, 69, 244–255. [Google Scholar] [CrossRef]
- Scafidi, P.; Pisciotta, A.; Patti, D.; Tamborra, P.; Di Lorenzo, R.; Barbagallo, M.G. Effect of artificial shading on the tannin accumulation and aromatic composition of the Grillo cultivar (Vitis vinifera L.). BMC Plant Biol. 2013, 13, 175. [Google Scholar] [CrossRef] [Green Version]
- Englezos, V.; Rantsiou, K.; Torchio, F.; Rolle, L.; Gerbi, V.; Cocolin, L. Exploitation of the non-Saccharomyces yeast Starmerella bacillaris (synonym Candida zemplinina) in wine fermentation: Physiological and molecular characterizations. Int. J. Food Microbiol. 2015, 199, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Benito, S. The impacts of Lachancea thermotolerans yeast strains on winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 6775–6790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scacco, A.; Oliva, D.; Di Maio, S.; Polizzotto, G.; Genna, G.; Tripodi, G.; Lanza, C.M.; Verzera, A. Indigenous Saccharomyces cerevisiae strains and their influence on the quality of Cataratto, Inzolia and Grillo white wines. Food Res. Int. 2012, 46, 1–9. [Google Scholar] [CrossRef]
- Di Maio, S.; Genna, G.; Gandolfo, V.; Amore, G.; Ciaccio, M.; Oliva, D. Presence of Candida zemplinina in Sicilian Musts and Selection of a Strain for Wine Mixed Fermentations. S. Afr. J. Enol. Vitic. 2012, 33, 80–87. [Google Scholar] [CrossRef]
- Nardi, T.; Corich, V.; Giacomini, A.; Blondin, B. A sulphite-inducible form of the sulphite efflux gene SSU1 in a Saccharomyces cerevisiae wine yeast. Microbiology 2010, 156, 1686–1696. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
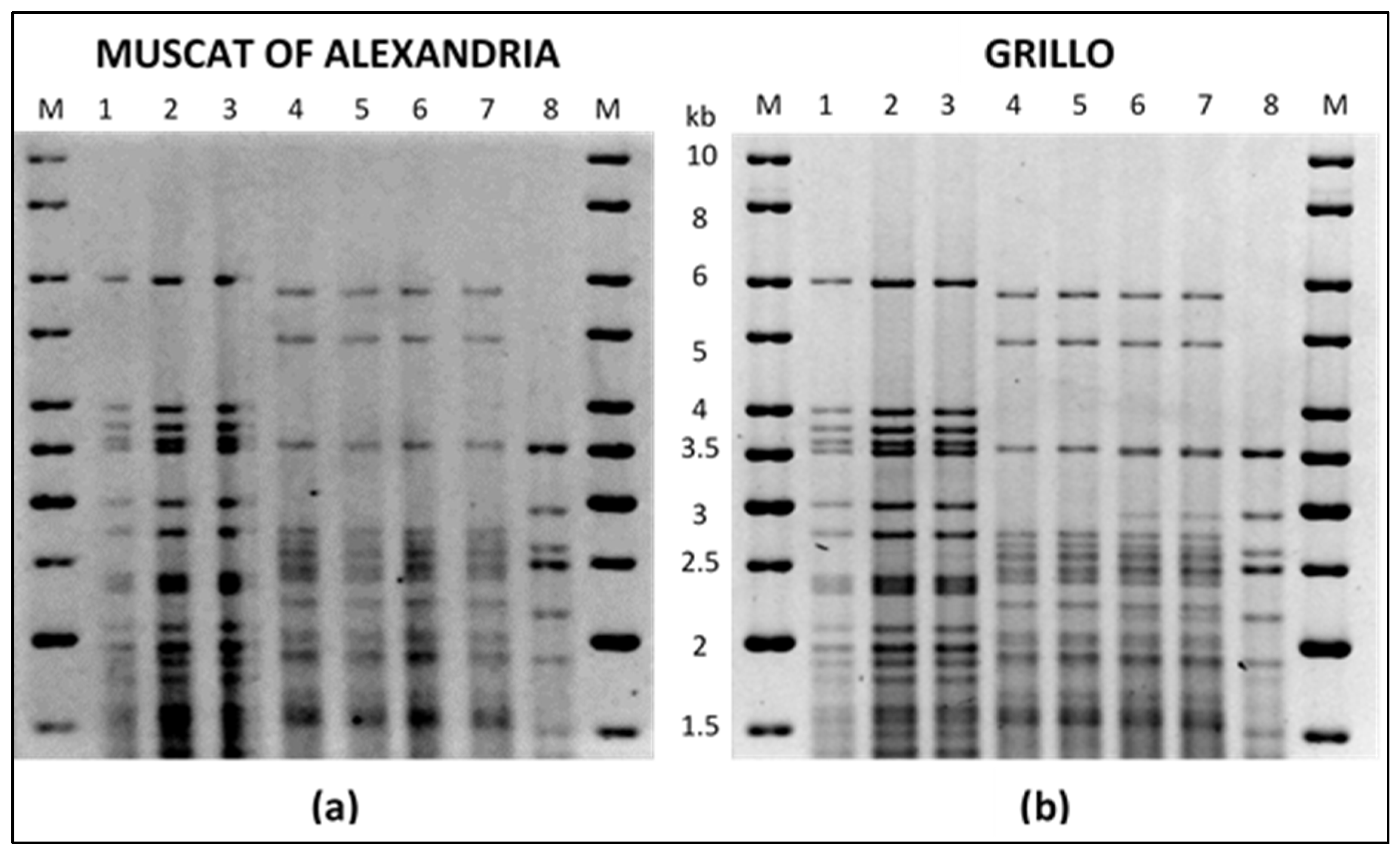{kind=link}
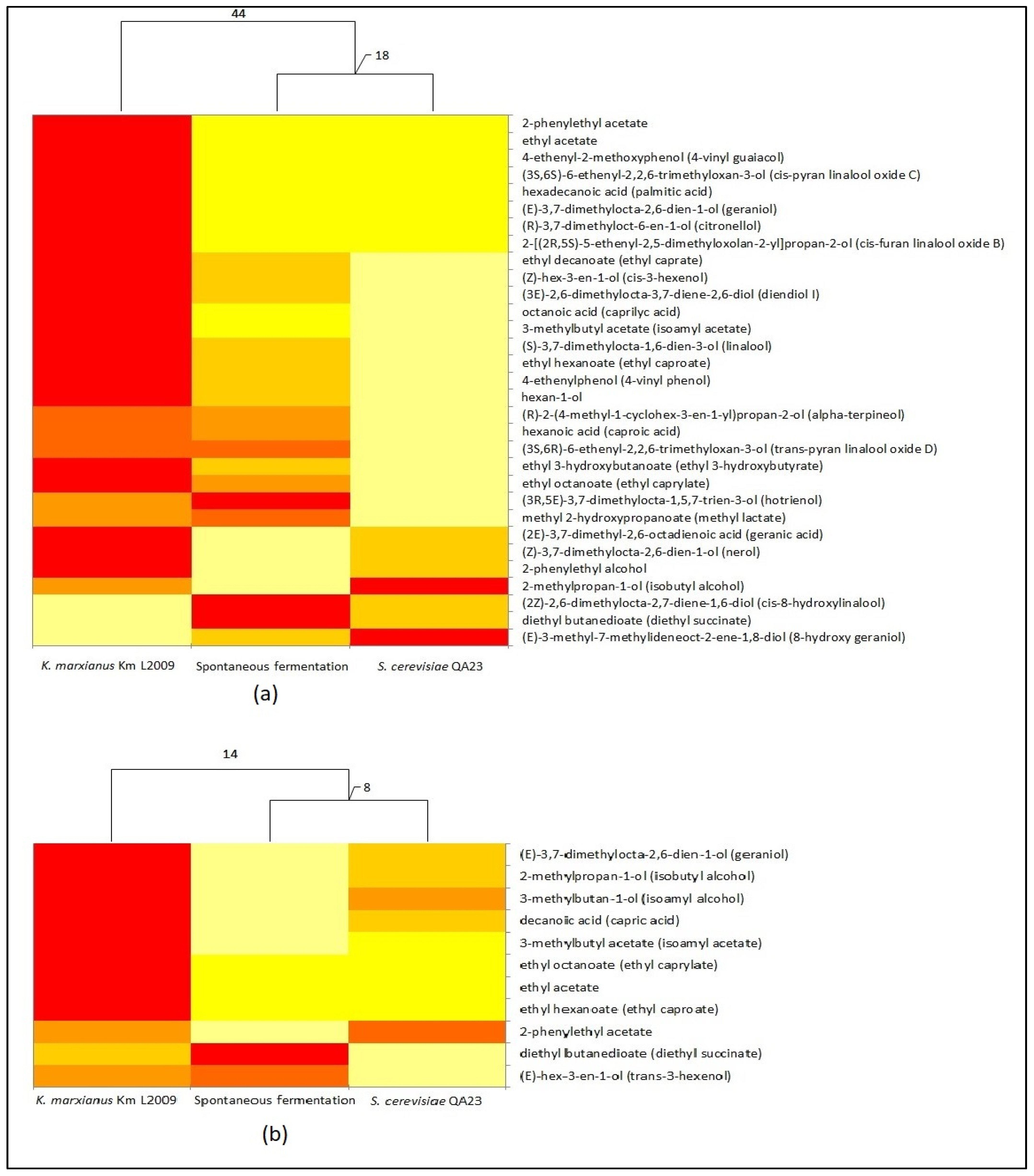{kind=link}
| Yeast Strain | Killer Activity |
|---|---|
| Saccharomyces cerevisiae Actiflore® F33 (Laffort Oenologie, Bordeaux, France) | + |
| Saccharomyces cerevisiae CK S102 (Fermentis, Marcq-en-Barœul, France) | + |
| Saccharomyces cerevisiae Cross EvolutionTM (Lallemand, Montreal, QC, Canada) | + |
| Saccharomyces cerevisiae Ezferm 44 (Esseco, San Martino di Trecate, Italy) | + |
| Saccharomyces cerevisiae Fermol Arome Plus (AEB, Brescia, Italy) | + |
| Saccharomyces cerevisiae Fermol Davis 522 (AEB, Brescia, Italy) | + |
| Saccharomyces cerevisiae Fermol Super 16 (AEB, Brescia, Italy) | − |
| Saccharomyces cerevisiae FR-WP (Ferrari, Verona, Italy) | − |
| Saccharomyces cerevisiae Lalvin BA11 (Lallemand, Montreal, QC, Canada) | + |
| Saccharomyces cerevisiae Lalvin EC1118 (Lallemand, Montreal, QC, Canada) | − |
| Saccharomyces cerevisiae Lalvin ICV K1 (Lallemand, Montreal, QC, Canada) | + |
| Saccharomyces cerevisiae Lalvin QA23 (Lallemand, Montreal, QC, Canada) | − |
| Saccharomyces cerevisiae PDM (Maurivin, Toowoomba, Australia) | − |
| Saccharomyces cerevisiae SIHA7 (Eaton, Nettersheim, Germany) | + |
| Saccharomyces cerevisiae UCLM S325 (Fermentis, Marcq-en-Barœul, France) | − |
| Saccharomyces cerevisiae Uvaferm DV10 (Lallemand, Montreal, QC, Canada) | −/+ |
| Saccharomyces cerevisiae Zymaflore® VL1 (Laffort Oenologie, Bordeaux, France) | + |
| Saccharomyces cerevisiae Zymaflore® VL3 (Laffort Oenologie, Bordeaux, France) | −/+ |
| Saccharomyces cerevisiae Zymaflore® ST (Laffort Oenologie, Bordeaux, France) | + |
| Saccharomyces cerevisiae Zymaflore® X5 (Laffort Oenologie, Bordeaux, France) | − |
| Saccharomyces cerevisiae Zymaflore® X16 (Laffort Oenologie, Bordeaux, France) | + |
| K. marxianus Km L2009 | S. cerevisiae X5 | K. marxianus Km L2009 + S. cerevisiae X5 | p-Values | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Alcohol (% v/v) | 9.70 a | ± | 0.16 | 11.68 c | ± | 0.03 | 10.16 b | ± | 0.01 | 0.001 |
| Total Dry Extract (g/L) | 67.2 c | ± | 2.5 | 0.8 a | ± | 0.0 | 48.1 b | ± | 0.2 | 0.001 |
| pH | 3.51 b | ± | 0.02 | 3.51 b | ± | 0.02 | 3.15 a | ± | 0.01 | 0.001 |
| Glycerol (g/L) | 6.13 b | ± | 0.21 | 4.40 a | ± | 0.00 | 5.55 b | ± | 0.35 | 0.001 |
| Tartaric Acid (g/L) | 1.62 | ± | 0.02 | 1.62 | ± | 0.02 | 1.59 | ± | 0.02 | |
| Malic Acid (g/L) | 1.14 a | ± | 0.02 | 1.22 b | ± | 0.04 | 1.14 a | ± | 0.02 | 0.05 |
| Lactic Acid (g/L) | 0.02 a | ± | 0.01 | 0.03 b | ± | 0.01 | 0.02 ab | ± | 0.01 | 0.05 |
| Glucose + Fructose (g/L) | 43.89 c | ± | 0.73 | 0.07 a | ± | 0.01 | 25.24 b | ± | 0.54 | 0.001 |
| Acetic Acid (g/L) | 1.22 a | ± | 0.01 | 2.36 c | ± | 0.00 | 2.15 b | ± | 0.04 | 0.001 |
| Citric Acid (g/L) | 0.74 b | ± | 0.04 | 0.37 a | ± | 0.01 | 2.20 c | ± | 0.08 | 0.001 |
| Total Phenolics (mg/L) | 112 | ± | 9 | 111 | ± | 3 | 103 | ± | 3 | |
| Total Catechins (mg/L) | 4.8 b | ± | 0.6 | 11.1 c | ± | 0.2 | 1.6 a | ± | 0.2 | 0.001 |
| Methanol (mg/L) | 13 | ± | 6 | 20 | ± | 3 | 16 | ± | 1 | |
| IUPAC Name (Synonym) | K. marxianus Km L2009 | S. cerevisiae X5 | K. marxianus Km L2009 + S. cerevisiae X5 | Odour Threshold Range | Sensory Description | |||
|---|---|---|---|---|---|---|---|---|
| Aliphatic alcohols | ||||||||
| 2-methylpropan-1-ol (isobutyl alcohol) | 1535.0 ± 61.0 | b | 405.6 ± 80.9 | a | 1247.3 ± 287.6 | b | 40,000–500,000 | Oily, bitter, green |
| 3-methylbutan-1-ol (isoamyl alcohol) | 92,148.1 ± 2278.0 | c | 72,384.9 ± 4352.2 | a | 81,779.0 ± 4090.2 | b | 30,000–300,000 | Burnt, alcohol |
| hexan-1-ol | 281.5 ± 21.4 | ab | 234.7 ± 22.4 | a | 288.2 ± 34.4 | b | 4000–8000 | Flower, green, cut grass |
| (E)-hex-3-en-1-ol (trans-3-hexenol) | 27.1 ± 5. 7 | b | 18.1 ± 2.6 | a | 26.1 ± 3.8 | ab | 1000 | Green |
| (Z)-hex-3-en-1-ol (cis-3-hexenol) | 54.8 ± 9.2 | 43.0 ± 6.0 | 52.2 ± 4.3 | 400 | Green, cut grass | |||
| Total aliphatic alcohols | 94,046.5 ± 2298.9 | c | 73,086.3 ± 4389.0 | a | 83,392.8 ± 3858.4 | b | ||
| Aliphatic esters | ||||||||
| 3-methylbutyl acetate (isoamyl acetate) | 507.1 ± 76.3 | b | 167.5 ± 12.2 | a | 463.2 ± 168.6 | b | 30–260 | Banana, apple, pear |
| ethyl hexanoate (ethyl caproate) | 104.5 ± 46.6 | 108.7 ± 25.1 | 61.2 ± 11.5 | 14–80 | Green apple, fruity | |||
| ethyl acetate | 43.2 ± 5.6 | 42.7 ± 14.3 | 39.3 ± 3.8 | 7500–17,620 | Fruity, solvent | |||
| methyl 2-hydroxypropanoate (methyl lactate) | 59.2 ± 8.0 | 46.9 ± 11.0 | 56.2 ± 14.2 | |||||
| ethyl octanoate (ethyl caprylate) | 16.8 ± 2.1 | b | 14.6 ± 2.8 | b | 7.0 ± 2.0 | a | 5–580 | Sweet, fruity |
| diethyl butanedioate (diethyl succinate) | 0.0 ± 0.0 | a | 88.1 ± 34.1 | b | 0.0 ± 0.0 | a | 75,000–200,000 | vinous |
| Total aliphatic esters | 730.8 ± 51.0 | b | 468.5 ± 50.0 | a | 626.9 ± 187.1 | ab | ||
| Aliphatic acids | ||||||||
| 2-methylpropanoic acid (isobutyric acid) | 262.1 ± 34.8 | b | 9.8 ± 4.3 | a | 240.8 ± 29.8 | b | 2300–8100 | Rancid, butter, cheese |
| hexanoic acid (caproic acid) | 181.0 ± 18.8 | ab | 236.0 ± 74.8 | b | 115.0 ± 41.3 | a | 420–8000 | Sweat |
| octanoic acid (caprilyc acid) | 174.1 ± 17.6 | c | 98.5 ± 16.2 | b | 57.2 ± 9.4 | a | 500–13,000 | Sweat, cheese |
| decanoic acid (capric acid) | 82.6 ± 11.7 | b | 52.4 ± 9.5 | a | 49.6 ± 13.3 | a | 1000–10,000 | Rancid fat |
| hexadecanoic acid (palmitic acid) | 267.0 ± 58.4 | 206.2 ± 38.3 | 196.8 ± 57.4 | |||||
| Total aliphatic acids | 966.8 ± 79.3 | b | 602.9 ± 124.0 | a | 659.4 ± 137.9 | a | ||
| Monoterpenes | ||||||||
| 2-[(2S,5S)-5-ethenyl-2,5-dimethyloxolan-2-yl]propan-2-ol (trans-furan linalool oxide A) | 11.9 ± 2.2 | ab | 8.9 ± 1.2 | a | 16.0 ± 5.7 | b | 6000–7000 | Leafy, sweet, floral, creamy, earthy |
| 2-[(2R,5S)-5-ethenyl-2,5-dimethyloxolan-2-yl]propan-2-ol (cis-furan linalool oxide B) | 22.8 ± 1.6 | b | 10.5 ± 7.5 | a | 19.5 ± 3.5 | ab | 6000 | Leafy, sweet, floral, creamy, earthy |
| (S)-3,7-dimethylocta-1,6-dien-3-ol (linalool) | 292.7 ± 33.7 | ab | 221.1 ± 29.2 | a | 361.9 ± 84.1 | b | 15–100 | Floral |
| (3R,5E)-3,7-dimethylocta-1,5,7-trien-3-ol (hotrienol) | 20.5 ± 2.8 | a | 26.5 ± 4.5 | ab | 40.5 ± 13.9 | b | 110 | Linden |
| (R)-2-(4-methyl-1-cyclohex-3-en-1-yl)propan-2-ol (alpha-terpineol) | 57.0 ± 6.3 | a | 43.9 ± 6.5 | a | 90.2 ± 23.6 | b | 250–400 | Pine, lily of the valley |
| (3S,6S)-6-ethenyl-2,2,6-trimethyloxan-3-ol (cis-pyran linalool oxide C) | 83.6 ± 11.1 | 76.7 ± 6.9 | 89.6 ± 25.2 | 3000–3600 | Leafy, sweet, floral, creamy, earthy | |||
| (3S,6R)-6-ethenyl-2,2,6-trimethyloxan-3-ol (trans-pyran linalool oxide D) | 48.0 ± 7.5 | b | 31.9 ± 3.0 | a | 51.5 ± 6.4 | b | 3000–5400 | Leafy, sweet, floral, creamy, earthy |
| (R)-3,7-dimethyloct-6-en-1-ol (citronellol) | 36.4 ± 6.8 | a | 43.3 ± 6.8 | ab | 52.4 ± 8.9 | b | 18–100 | Green lemon |
| (Z)-3,7-dimethylocta-2,6-dien-1-ol (nerol) | 16.0 ± 3.1 | 16.4 ± 14.0 | 16.8 ± 3.9 | 400 | Rose, lime | |||
| (E)-3,7-dimethylocta-2,6-dien-1-ol (geraniol) | 40.0 ± 6.2 | b | 8.9 ± 10.4 | a | 12.8 ± 9.9 | a | 30–130 | Roses, geranium |
| (3E)-2,6-dimethylocta-3,7-diene-2,6-diol (diendiol I) | 864.9 ± 50.7 | b | 549.3 ± 95.0 | a | 774.1 ± 194.1 | ab | Odorless | |
| 3,7-dimethylocta-1-en-3,7-diol (endiol) | 37.3 ± 7.0 | 24.0 ± 6.3 | 32.4 ± 11.5 | |||||
| 2,6-dimethylocta-1,7-diene-3,6-diol (diendiol II) | 111.5 ± 14.7 | 73.8 ± 8.8 | 95.0 ± 29.2 | |||||
| p-menth-1-ene-6,8-diol (p-menthenediol II) | 24.3 ± 6.2 | b | 11.3 ± 2.1 | a | 13.6 ± 1.9 | a | ||
| (E)-3-methyl-7-methylideneoct-2-ene-1,8-diol (8-hydroxy geraniol) | 11.6 ± 4.1 | 12.0 ± 2.5 | 16.6 ± 6.1 | |||||
| (2Z)-2,6-dimethylocta-2,7-diene-1,6-diol (cis-8-hydroxylinalool) | 11.5 ± 2.7 | 12.3 ± 4.4 | 16.7 ± 8.2 | |||||
| (2E)-3,7-dimethyl-2,6-octadienoic acid (geranic acid) | 51.7 ± 8.7 | 35.5 ± 15.4 | 53.4 ± 14.7 | |||||
| Total monoterpenes | 1741.7 ± 131.9 | b | 1206.3 ± 80.9 | a | 1753.0 ± 404.3 | b | ||
| Volatile phenols | ||||||||
| 4-ethenyl-2-methoxyphenol (4-vinyl guaiacol) | 55.1 ± 10.0 | a | 175.2 ± 27.8 | b | 64.0 ± 27.3 | a | 440–1100 | Smoky, vanilla, clovelike |
| 4-ethenylphenol (4-vinyl phenol) | 38.0 ± 11.4 | a | 125.9 ± 21.2 | b | 26.7 ± 6.0 | a | 180–770 | Pharmaceutical, elastoplast, gouache |
| 1-(4-hydroxy-3-methoxyphenyl)ethanone (acetovanillone) | 24.4 ± 11.5 | 17.7 ± 3.2 | 14.2 ± 4.4 | 1000 | Sweet spices | |||
| Total volatile phenols | 117.5 ± 28.8 | a | 318.8 ± 48.6 | b | 104.9 ± 32.6 | a | ||
| Benzenoids | ||||||||
| 2-phenylethyl acetate | 5282.6 ± 541.5 | b | 57.5 ± 12.2 | a | 4629.3 ± 1150.6 | b | 250–650 | Rose, honey, apple, sweetish, flowery |
| 2-phenylethanol | 8789.4 ± 1461.2 | 9161.4 ± 209.0 | 8517.7 ± 1064.6 | 7500–14,000 | Floral, roses | |||
| Total benzenoids | 14,072.0 ± 926.6 | b | 9218.9 ± 209.2 | a | 13,147.0 ± 2138.4 | b | ||
| IUPAC Name (Synonym) | Spontaneous Fermentation | strain S. cerevisiae QA23 | strain K. marxianus Km L2009 | Odour Threshold Range | Sensory Description | |||
|---|---|---|---|---|---|---|---|---|
| Aliphatic alcohols | ||||||||
| 2-methylpropan-1-ol (isobutyl alcohol) | 6785.8 ± 365.3 | a | 12,447.2 ± 1350.2 | c | 10,653.6 ± 665.1 | b | 40,000–500,000 | Oily, bitter, green |
| 3-methylbutan-1-ol (isoamyl alcohol) | 124,063.0 ± 7570.8 | 132,503.3 ± 8271.5 | 136,284.5 ± 6431.6 | 30,000–300,000 | Burnt, alcohol | |||
| hexan-1-ol | 672.8 ± 103.1 | ab | 564.7 ± 49.4 | a | 791.1 ± 128.2 | b | 4000–8000 | Flower, green, cut grass |
| (E)-hex-3-en-1-ol (trans-3-hexenol) | 18.4 ± 9.5 | 19.8 ± 6.6 | 28.5 ± 3.4 | 1000 | Green | |||
| (Z)-hex-3-en-1-ol (cis-3-hexenol) | 133.0 ± 8.0 | b | 104.3 ± 11.8 | a | 180.2 ± 9.3 | c | 400 | Green, cut grass |
| Total aliphatic alcohols | 131,673.0 ± 7631.0 | a | 145,639.3 ± 8942.9 | ab | 147,937.9 ± 5962.5 | b | ||
| Aliphatic esters | ||||||||
| 3-methylbutyl acetate (isoamyl acetate) | 5169.4 ± 893.9 | a | 3696.8 ± 306.5 | a | 7970.0 ± 937.5 | b | 30–260 | Banana, apple, pear |
| ethyl hexanoate (ethyl caproate) | 1171.3 ± 296.3 | ab | 955.2 ± 151.2 | a | 1464.4 ± 163.8 | b | 14–80 | Green apple, fruity |
| ethyl acetate | 367.9 ± 39.0 | a | 324.6 ± 41.9 | a | 722.7 ± 86.7 | b | 7500–17,620 | Fruity, solvent |
| methyl 2-hydroxypropanoate (methyl lactate) | 82.3 ± 20.5 | b | 50.5 ± 6.6 | a | 74.4 ± 10.8 | ab | ||
| ethyl octanoate (ethyl caprylate) | 1520.5 ± 163.4 | b | 831.2 ± 96.3 | a | 1812.8 ± 97.1 | c | 5–580 | Sweet, fruity |
| ethyl 3-hydroxybutanoate (ethyl 3-hydroxybutyrate) | 47.1 ± 15.8 | ab | 28.7 ± 11.3 | a | 58.3 ± 16.3 | b | 20,000 | fruity |
| ethyl decanoate (ethyl caprate) | 445.0 ± 162.4 | a | 281.9 ± 10.0 | a | 712.7 ± 92.3 | b | 200–500 | Sweet, fruity |
| diethyl butanedioate (diethyl succinate) | 32.3 ± 12.7 | b | 20.7 ± 5.0 | ab | 11.6 ± 3.4 | a | 75,000–200,000 | Vinous |
| Total esters and acetates | 8835.8 ± 1441.2 | b | 6189.6 ± 543.5 | a | 12,826.9 ± 1033.7 | c | ||
| Aliphatic acids | ||||||||
| hexanoic acid (caproic acid) | 3485.7 ± 254.4 | ab | 2222.1 ± 218.7 | a | 3696.0 ± 1,060.9 | b | 420–8000 | Sweat |
| octanoic acid (caprilyc acid) | 7739.6 ± 733.0 | b | 6289.2 ± 710.9 | a | 10,161.6 ± 589.1 | c | 500–13,000 | Sweat, cheese |
| decanoic acid (capric acid) | 1943.2 ± 936.3 | 2328.4 ± 245.3 | 3,057.1 ± 232.2 | 1000–10,000 | Rancid fat | |||
| hexadecanoic acid (palmitic acid) | 266.7 ± 17.9 | a | 218.2 ± 70.2 | a | 432.8 ± 60.9 | b | ||
| Total aliphatic acids | 13,435.2 ± 1481.5 | a | 11,057.9 ± 903.9 | a | 17,347.5 ± 1433.9 | b | ||
| Monoterpenes | ||||||||
| 2-[(2R,5S)-5-ethenyl-2,5-dimethyloxolan-2-yl]propan-2-ol (cis-furan linalool oxide B) | 20.8 ± 20.2 | a | 21.9 ± 3.7 | a | 57.8 ± 4.2 | b | 6000 | Leafy, sweet, floral, creamy, earthy |
| (S)-3,7-dimethylocta-1,6-dien-3-ol (linalool) | 332.8 ± 89.8 | ab | 260.7 ± 45.0 | a | 437.3 ± 47.3 | b | 15–100 | Floral |
| (3R,5E)-3,7-dimethylocta-1,5,7-trien-3-ol (hotrienol) | 27.7 ± 2.6 | b | 14.7 ± 2.9 | a | 23.0 ± 5.9 | b | 110 | Linden |
| (R)-2-(4-methyl-1-cyclohex-3-en-1-yl)propan-2-ol (alpha-terpineol) | 77.9 ± 28.1 | b | 28.9 ± 12.6 | a | 84.0 ± 4.7 | b | 250–400 | Pine, lily of the valley |
| (3S,6S)-6-ethenyl-2,2,6-trimethyloxan-3-ol (cis-pyran linalool oxide C) | 82.8 ± 8.8 | a | 70.8 ± 10.4 | a | 117.6 ± 3.2 | b | 3000–3600 | Leafy, sweet, floral, creamy, earthy |
| (3S,6R)-6-ethenyl-2,2,6-trimethyloxan-3-ol (trans-pyran linalool oxide D) | 104.7 ± 7.7 | b | 66.3 ± 11.1 | a | 101.6 ± 13.9 | b | 3000–5400 | Leafy, sweet, floral, creamy, earthy |
| (R)-3,7-dimethyloct-6-en-1-ol (citronellol) | 10.2 ± 1.5 | a | 11.2 ± 1.3 | a | 20.8 ± 6.5 | b | 18–100 | Green lemon |
| (Z)-3,7-dimethylocta-2,6-dien-1-ol (nerol) | 14.3 ± 5.9 | a | 20.2 ± 4.1 | ab | 26.5 ± 1.8 | b | 400 | Rose, lime |
| (E)-3,7-dimethylocta-2,6-dien-1-ol (geraniol) | 13.7 ± 1.6 | a | 15.7 ± 4.8 | a | 31.4 ± 4.7 | b | 30–130 | Roses, geranium |
| (3E)-2,6-dimethylocta-3,7-diene-2,6-diol (diendiol I) | 609.5 ± 27.1 | a | 496.8 ± 79.6 | a | 792.0 ± 126.6 | b | Odorless | |
| 3,7-dimethylocta-1-en-3,7-diol (endiol) | 35.4 ± 16.9 | 26.7 ± 5.4 | 29.7 ± 4.4 | |||||
| 2,6-dimethylocta-1,7-diene-3,6-diol (diendiol II) | 107.7 ± 37.2 | 75.1 ± 8.0 | 116.9 ± 16.9 | |||||
| p-menth-1-ene-6,8-diol (p-menthenediol II) | 42.6 ± 21.9 | 46.2 ± 27.6 | 54.2 ± 15.3 | |||||
| (E)-3-methyl-7-methylideneoct-2-ene-1,8-diol (8-hydroxy geraniol) | 23.1 ± 6.7 | ab | 30.8 ± 6.5 | b | 17.0 ± 3.3 | a | ||
| (2Z)-2,6-dimethylocta-2,7-diene-1,6-diol (cis-8-hydroxylinalool) | 28.6 ± 5.0 | c | 19.6 ± 1.7 | b | 13.2 ± 1.8 | a | ||
| (2E)-3,7-dimethyl-2,6-octadienoic acid (geranic acid) | 303.7 ± 45.1 | a | 419.9 ± 54.7 | a | 553.9 ± 85.5 | b | ||
| Total monoterpenes | 1835.5 ± 124.2 | a | 1625.5 ± 130.1 | a | 2476.9 ± 218.4 | b | ||
| Volatile phenols | ||||||||
| 4-ethenyl-2-methoxyphenol (4-vinyl guaiacol) | 54.5 ± 21.8 | a | 34.0 ± 12.4 | a | 328.9 ± 42.9 | b | 440–1100 | Smoky, vanilla, clovelike |
| 4-ethenylphenol (4-vinyl phenol) | 500.4 ± 124.8 | b | 195.4 ± 63.2 | a | 814.8 ± 23.2 | c | 180–770 | Pharmaceutical, elastoplast, gouache |
| Total volatile phenols | 554.9 ± 105.0 | b | 229.4 ± 75.4 | a | 1143.7 ± 49.7 | c | ||
| Benzenoids | ||||||||
| 2-phenylethyl acetate | 378.9 ± 85.4 | a | 347.9 ± 43.4 | a | 569.1 ± 10.9 | b | 250–650 | Rose, honey, apple, sweetish, flowery |
| 2-phenylethyl alcohol | 6139.9 ± 707.8 | a | 8695.7 ± 845.8 | b | 10,308.8 ± 1195.2 | b | 7500–14,000 | Floral, roses |
| Total benzenoids | 6518.8 ± 788.2 | a | 9043.6 ± 887.0 | b | 10,877.9 ± 1189.0 | b | ||
| IUPAC Name (Synonym) | Spontaneous Fermentation | strain S. cerevisiae QA23 | strain K. marxianus Km L2009 | Odour Threshold Range | Sensory Description | |||
|---|---|---|---|---|---|---|---|---|
| Aliphatic alcohols | ||||||||
| 2-methylpropan-1-ol (isobutyl alcohol) | 10,631.2 ± 2108.7 | a | 14,878.1 ± 4148.9 | ab | 18,931.2 ± 18,179 | b | 40,000–500,000 | Oily, bitter, green |
| 3-methylbutan-1-ol (isoamyl alcohol) | 136,320.3 ± 17,597.8 | a | 185,100.3 ± 21,295.9 | b | 213,619.9 ± 23,524.7 | b | 30,000–300,000 | Burnt, alcohol |
| hexan-1-ol | 787.5 ± 44.0 | 800.7 ± 87.4 | 797.8 ± 134.7 | 4000–8000 | Flower, green, cut grass | |||
| (E)-hex-3-en-1-ol (trans-3-hexenol) | 14.9 ± 1.9 | b | 10.8 ± 1.4 | a | 14.1 ± 2.3 | ab | 1000 | Green |
| (Z)-hex-3-en-1-ol (cis-3-hexenol) | 23.9 ± 4.2 | 18.9 ± 2.7 | 22.3 ± 2.4 | 400 | Green, cut grass | |||
| Total aliphatic alcohols | 147,777.8 ± 19,378.1 | a | 200,808.8 ± 17,829.9 | b | 233,385.3 ± 22,276.0 | b | ||
| Aliphatic esters | ||||||||
| 3-methylbutyl acetate (isoamyl acetate) | 1360.1 ± 232.4 | a | 1964.1 ± 264.9 | a | 2983.6 ± 459.3 | b | 30–260 | Banana, apple, pear |
| ethyl hexanoate (ethyl caproate) | 343.4 ± 73.6 | a | 414.2 ± 59.1 | a | 753.4 ± 134.7 | b | 14–80 | Green apple, fruity |
| ethyl acetate | 266.7 ± 51.0 | a | 266.2 ± 26.6 | a | 474.8 ± 137.3 | b | 7500–17,620 | Fruity, solvent |
| methyl 2-hydroxypropanoate (methyl lactate) | 31.4 ± 13.2 | 37.3 ± 5.5 | 39.0 ± 12.1 | |||||
| ethyl octanoate (ethyl caprylate) | 486.3 ± 40.0 | a | 472.7 ± 155.3 | a | 838.6 ± 155.1 | b | 5–580 | Sweet, fruity |
| ethyl 3-hydroxybutanoate (ethyl 3-hydroxybutyrate) | 20.3 ± 2.4 | 23.0 ± 8.6 | 20.3 ± 1.0 | 20,000 | Fruity | |||
| ethyl decanoate (ethyl caprate) | 140.7 ± 46.6 | 129.7 ± 33.1 | 167.2 ± 41.5 | 200–500 | Sweet, fruity | |||
| diethyl butanedioate (diethyl succinate) | 18.2 ± 6.0 | b | 9.3 ± 2.5 | a | 13.9 ± 3.9 | ab | 75,000–200,000 | Vinous |
| Total aliphatic esters | 2667.1 ± 290.7 | a | 3316.5 ± 473.9 | a | 5290.8 ± 911.9 | b | ||
| Aliphatic acids | ||||||||
| hexanoic acid (caproic acid) | 1116.1 ± 161.7 | 1275.3 ± 263.0 | 1846.2 ± 595.7 | 420–8000 | Sweat | |||
| octanoic acid (caprilyc acid) | 2949.1 ± 374.2 | 3275.8 ± 339.2 | 4448.8 ± 1370.3 | 500–13,000 | Sweat, cheese | |||
| decanoic acid (capric acid) | 658.3 ± 497.8 | a | 1128.5 ± 199.5 | ab | 1798.6 ± 716.1 | b | 1000–10,000 | Rancid fat |
| hexadecanoic acid (palmitic acid) | 217.9 ± 58.0 | 318.5 ± 129.5 | 402.7 ± 79.7 | |||||
| Total aliphatic acids | 4941.4 ± 967.2 | a | 5998.1 ± 899.7 | ab | 8496.3 ± 2720.1 | b | ||
| Monoterpenes | ||||||||
| (Z)-3,7-dimethylocta-2,6-dien-1-ol (nerol) | 2.9 ± 3.5 | 10.0 ± 9.0 | 7.5 ± 6.4 | 400 | Rose, lime | |||
| (E)-3,7-dimethylocta-2,6-dien-1-ol (geraniol) | 2.6 ± 3.1 | a | 6.9 ± 2.2 | ab | 11.2 ± 3.6 | b | 30–130 | Rose, geranium |
| 3,7-dimethylocta-1-en-3,7-diol (endiol) | 17.4 ± 3.5 | 17.4 ± 20.3 | 3.2 ± 3.7 | |||||
| 2,6-dimethylocta-1,7-diene-3,6-diol (diendiol II) | 9.3 ± 10.8 | 3.6 ± 4.3 | 0.0 ± 0.0 | |||||
| (E)-3-methyl-7-methylideneoct-2-ene-1,8-diol (8-hydroxy geraniol) | 4.4 ± 5.2 | 4.2 ± 4.8 | 5.0 ± 5.9 | |||||
| (2Z)-2,6-dimethylocta-2,7-diene-1,6-diol (cis-8-hydroxylinalool) | 5.6 ± 6.5 | 5.6 ± 6.7 | 5.1 ± 6.2 | |||||
| (2E)-3,7-dimethyl-2,6-octadienoic acid (geranic acid) | 34.8 ± 40.6 | 50.6 ± 5.6 | 73.9 ± 22.8 | |||||
| Total monoterpenes | 77.0 ± 51.0 | 98.3 ± 42.6 | 105.9 ± 4.4 | |||||
| Volatile phenols | ||||||||
| 4-ethenyl-2-methoxyphenol (4-vinyl guaiacol) | 210.5 ± 54.3 | 197.9 ± 47.3 | 298.1 ± 60.5 | 440–1100 | Smoky, vanilla, clovelike | |||
| 4-ethenylphenol (4-vinyl phenol) | 78.2 ± 28.2 | 86.8 ± 15.9 | 137.7 ± 62.6 | 180–770 | Pharmaceutical, elastoplast, gouache | |||
| Total volatile phenols | 288.7 ± 79.4 | 284.7 ± 61.7 | 435.8 ± 121.6 | |||||
| Benzenoids | ||||||||
| 2-phenylethyl acetate | 167.4 ± 34.0 | a | 301.9 ± 56.6 | b | 276.6 ± 67.1 | b | 250–650 | Rose, honey, apple, sweetish; flowery |
| Total benzenoids | 167.4 ± 34.0 | a | 301.9 ± 56.6 | b | 276.6 ± 67.1 | b | ||
| Pairs of Wines | Number of Judges | Number of Correct Answers | Number of Correct AnswersRequired for p < α | Significance | |
|---|---|---|---|---|---|
| Muscat of Alexandria wines | Spontaneous vs. Spontaneous | 25 | 13 | 18 for p < 0.05 | n.s. |
| S. cerevisiae vs. S. cerevisiae | 25 | 13 | 18 for p < 0.05 | n.s. | |
| Km L2009 vs. Km L2009 | 25 | 13 | 18 for p < 0.05 | n.s. | |
| Spontaneous vs. S. cerevisiae | 25 | 18 | 18 for p < 0.05 | p < 0.05 | |
| Spontaneous vs. Km L2009 | 23 | 12 | 16 for p < 0.05 | n.s. | |
| S. cerevisiae vs. Km L2009 | 23 | 16 | 16 for p < 0.05 | p < 0.05 | |
| Grillo wines | Spontaneous vs. Spontaneous | 25 | 11 | 18 for p < 0.05 | n.s. |
| S. cerevisiae vs. S. cerevisiae | 25 | 13 | 18 for p < 0.05 | n.s. | |
| Km L2009 vs. Km L2009 | 25 | 14 | 18 for p < 0.05 | n.s. | |
| Spontaneous vs. S. cerevisiae | 23 | 16 | 16 for p < 0.05 | p < 0.05 | |
| Spontaneous vs. Km L2009 | 23 | 18 | 18 for p < 0.01 | p < 0.01 | |
| S. cerevisiae vs. Km L2009 | 23 | 16 | 16 for p < 0.05 | p < 0.05 |
| Pairs of Wines | Number of Judges | Number of Correct Answers | Number of Correct Answers Required for p < α | Significance | |
|---|---|---|---|---|---|
| Muscat of Alexandria wines | Spontaneous vs. Spontaneous | 25 | 11 vs. 14 | 18 for p < 0.05 | n.s. |
| S. cerevisiae vs. S. cerevisiae | 25 | 12 vs. 13 | 18 for p < 0.05 | n.s. | |
| Km L2009 vs. Km L2009 | 25 | 13 vs. 12 | 18 for p < 0.05 | n.s. | |
| Spontaneous vs. S. cerevisiae | 25 | 12 vs. 13 | 18 for p < 0.05 | n.s. | |
| Spontaneous vs. Km L2009 | 23 | 11 vs. 12 | 17 for p < 0.05 | n.s. | |
| S. cerevisiae vs. Km L2009 | 23 | 12 vs. 11 | 17 for p < 0.05 | n.s. | |
| Grillo wines | Spontaneous vs. Spontaneous | 25 | 11 vs. 14 | 18 for p < 0.05 | n.s. |
| S. cerevisiae vs. S. cerevisiae | 25 | 13 vs. 12 | 18 for p < 0.05 | n.s. | |
| Km L2009 vs. Km L2009 | 25 | 12 vs. 13 | 18 for p < 0.05 | n.s. | |
| Spontaneous vs. S. cerevisiae | 23 | 11 vs. 12 | 17 for p < 0.05 | n.s. | |
| Spontaneous vs. Km L2009 | 23 | 11 vs. 12 | 17 for p < 0.05 | n.s. | |
| S. cerevisiae vs. Km L2009 | 23 | 6 vs. 17 | 17 for p < 0.05 | p < 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barone, E.; Ponticello, G.; Giaramida, P.; Squadrito, M.; Fasciana, T.; Gandolfo, V.; Ardizzone, F.; Monteleone, M.; Corona, O.; Francesca, N.; et al. Use of Kluyveromyces marxianus to Increase Free Monoterpenes and Aliphatic Esters in White Wines. Fermentation 2021, 7, 79. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7020079
Barone E, Ponticello G, Giaramida P, Squadrito M, Fasciana T, Gandolfo V, Ardizzone F, Monteleone M, Corona O, Francesca N, et al. Use of Kluyveromyces marxianus to Increase Free Monoterpenes and Aliphatic Esters in White Wines. Fermentation. 2021; 7(2):79. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7020079
Chicago/Turabian StyleBarone, Eleonora, Giovanna Ponticello, Pieramaria Giaramida, Margherita Squadrito, Teresa Fasciana, Valentina Gandolfo, Francesco Ardizzone, Manuela Monteleone, Onofrio Corona, Nicola Francesca, and et al. 2021. "Use of Kluyveromyces marxianus to Increase Free Monoterpenes and Aliphatic Esters in White Wines" Fermentation 7, no. 2: 79. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7020079