
Acetaldehyde Stimulation of the Growth of Zymomonas mobilis Subjected to Ethanol and Other Environmental Stresses: Effect of Other Metabolic Electron Acceptors and Evidence for a Mechanism


Abstract
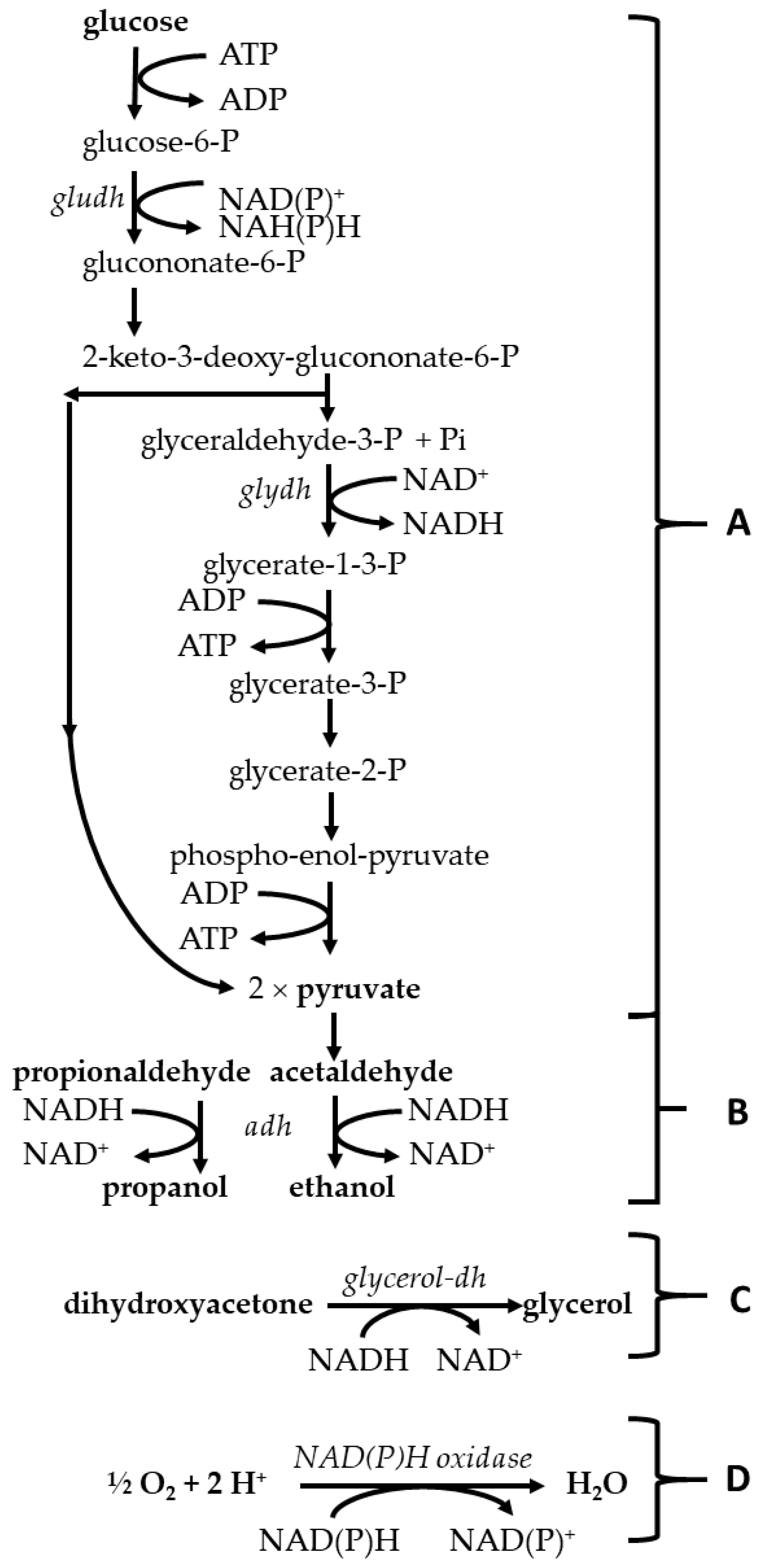
:1. Introduction
2. Materials and Methods
2.1. Organism, Media, Inoculum Preparation and Experimental Procedures.
2.2. Glucose Utilization in Nongrowing Cultures
2.3. Analyses
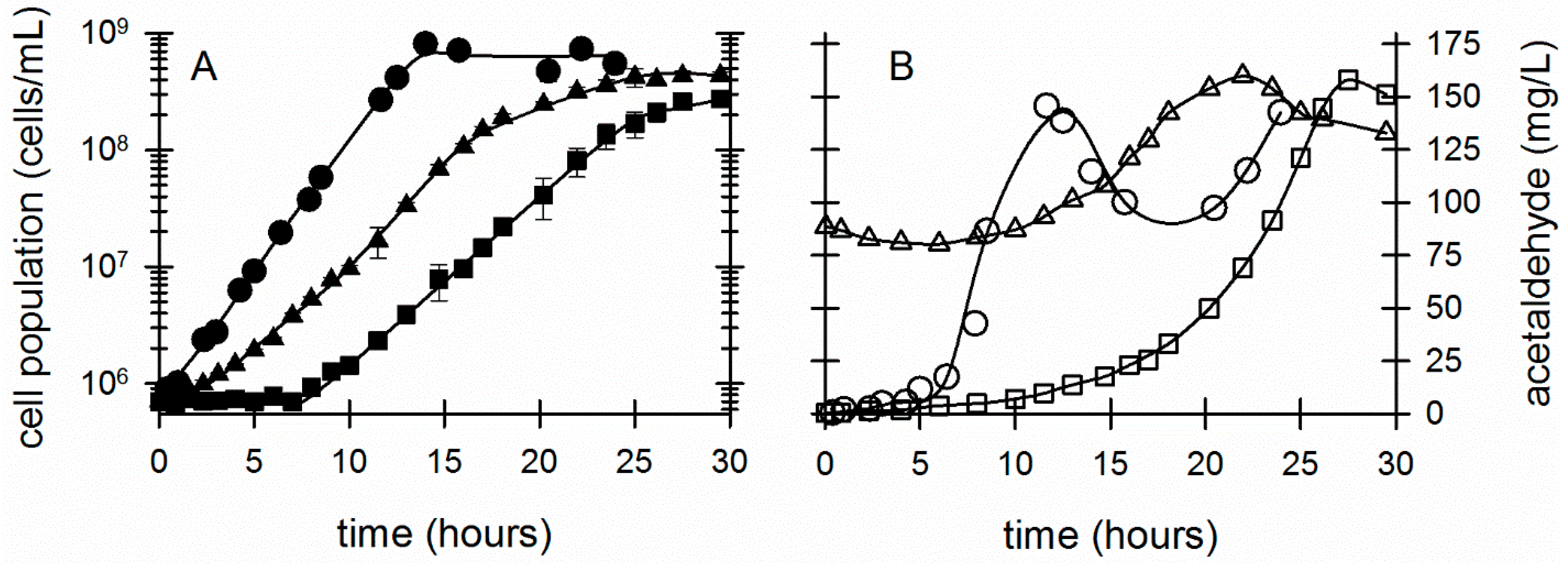
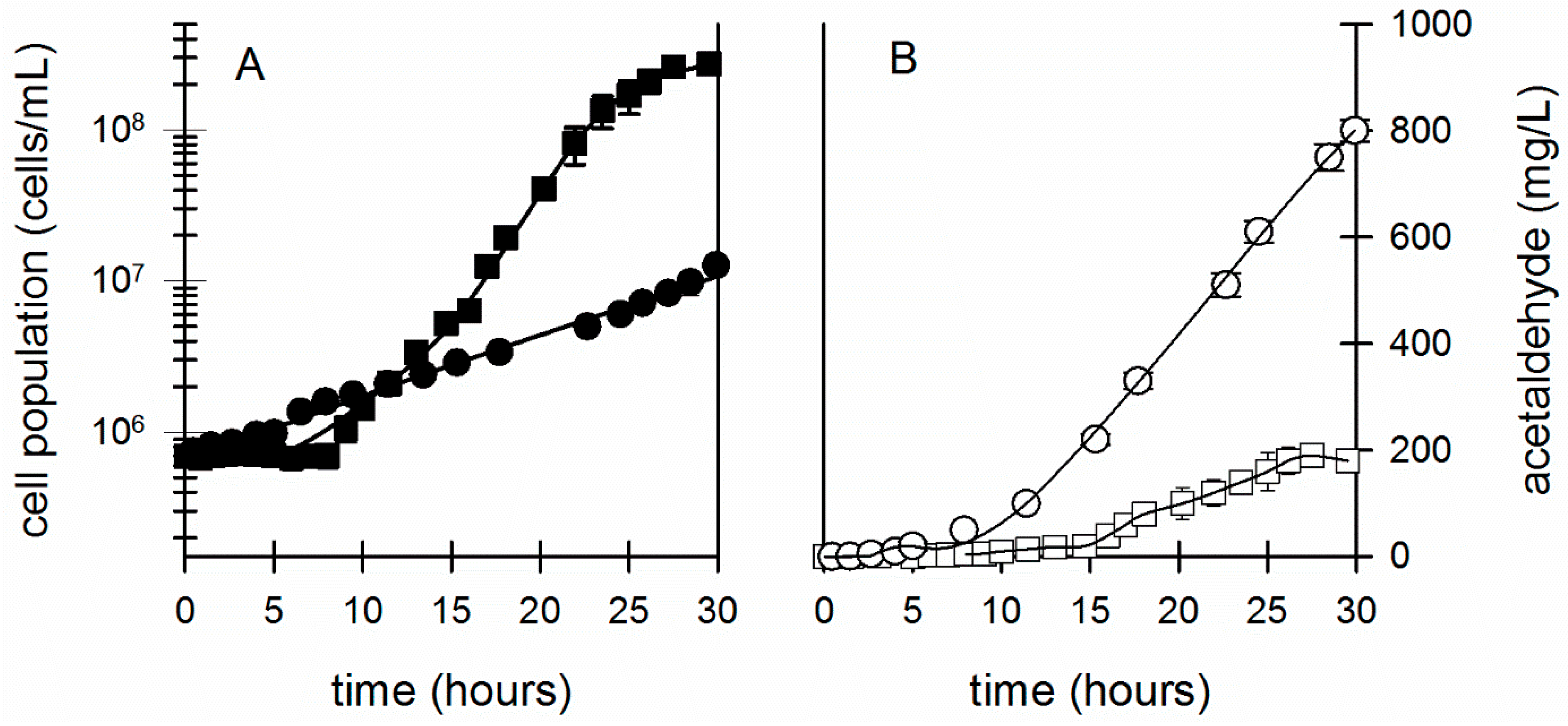
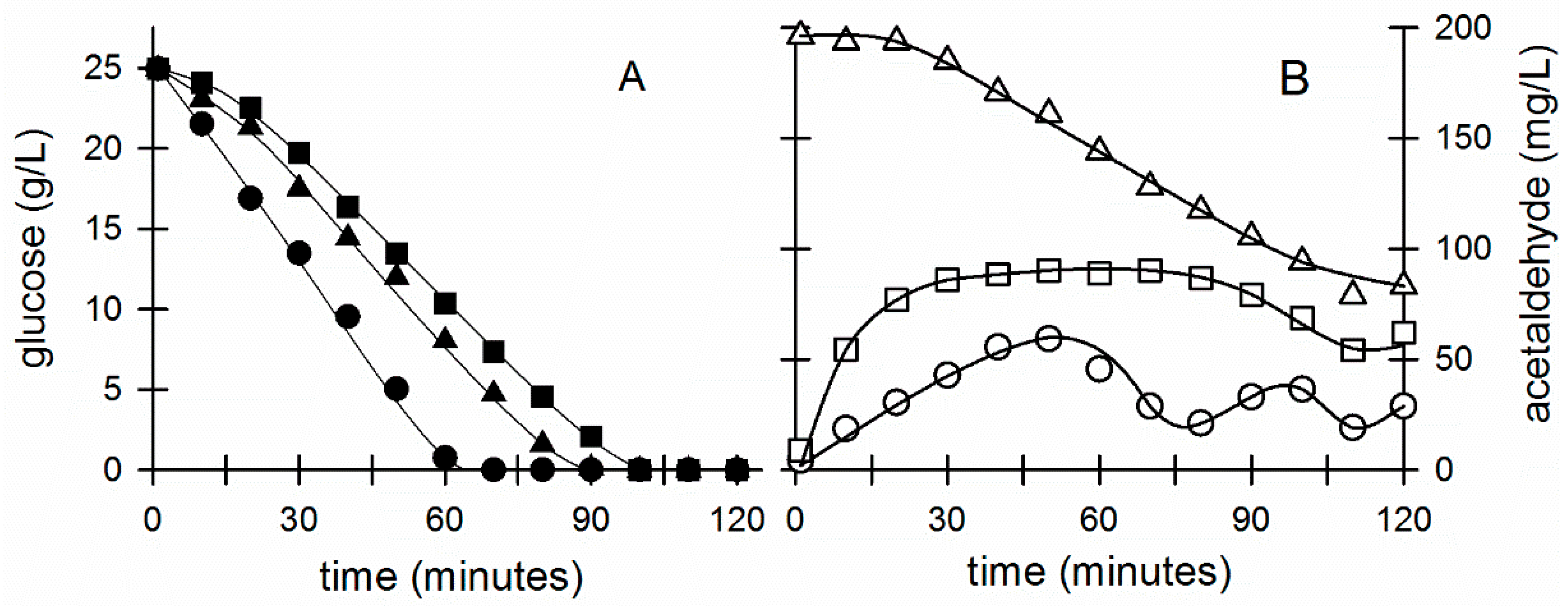
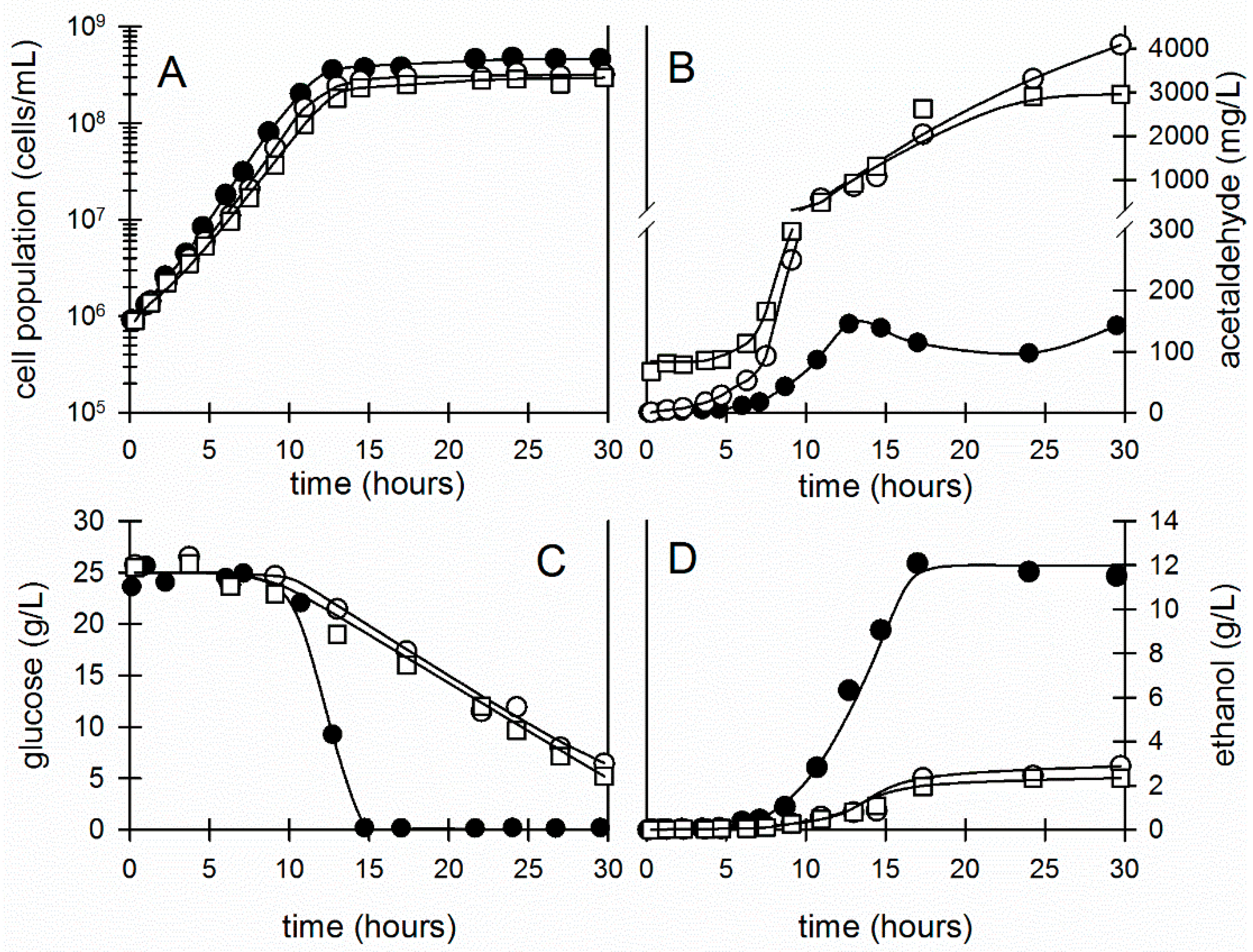
3. Results
3.1. Evidence That the Acetaldehyde Effect in Z. mobilis Is Redox-Based and Leads to an Enhanced Glycolytic Flux
3.2. Effect of Acetaldehyde on Z. mobilis Exposed to Other Environmental Stresses
4. Discussion
5. Conclusions and Recommendations for Further Work
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Althuri, A.; Gujjala, L.K.S.; Banerjee, R. Partially consolidated bioprocessing of mixed lignocellulosic feedstocks for ethanol production. Bioresour. Technol. 2017, 245, 530–539. [Google Scholar] [CrossRef]
- Panesar, P.S.; Marwaha, S.S.; Kennedy, J.F. Zymomonas mobilis: An alternative ethanol producer. J. Chem. Technol. Biotechnol. 2006, 81, 623–635. [Google Scholar] [CrossRef]
- Weir, P.M. The ecology of Zymomonas: A review. Folia Microbiol. 2016, 61, 385–392. [Google Scholar] [CrossRef]
- Yanase, H.; Miyawaki, H.; Sakurai, M.; Kawakami, A.; Matsumoto, M.; Haga, K.; Kojima, M.; Okamoto, K. Ethanol production from wood hydrolysate using genetically engineered Zymomonas mobilis. Appl. Microbiol. Biotechnol. 2012, 94, 1667–1678. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Yang, Y.; Liu, C.-G.; Yang, S.; Bai, F.-W. Engineering Zymomonas mobilis for Robust Cellulosic Ethanol Production. Trends Biotechnol. 2019, 37, 960–972. [Google Scholar] [CrossRef] [PubMed]
- Swings, J.; de Ley, J. The biology of Zymomonas. Bacteriol. Rev. 1977, 41, 1–46. [Google Scholar] [CrossRef]
- Franden, M.A.; Pilath, H.M.; Mohagheghi, A.; Pienkos, P.T.; Zhang, M. Inhibition of growth of Zymomonas mobilis by model compounds found in lignocellulosic hydrolysates. Biotechnol. Biofuels 2013, 6, 99. [Google Scholar] [CrossRef] [Green Version]
- Vriesekoop, F.; Rasmusson, M.; Pamment, N. Respective effects of sodium and chloride ions on filament formation and growth and ethanol production in Zymomonas mobilis fermentations. Lett. Appl. Microbiol. 2002, 35, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, A.R.; Hansson, H.; Pamment, N.B. Acetaldehyde stimulation of growth of Saccharomyces cerevisiae in the presence of inhibitors found in lignocellulose-to-ethanol fermentations. J. Ind. Microbiol. Biotechnol. 2000, 25, 104–108. [Google Scholar] [CrossRef]
- Barber, A.R.; Vriesekoop, F.; Pamment, N.B. Effects of acetaldehyde on Saccharomyces cerevisiae exposed to a range of chemical and environmental stresses. Enzym. Microb. Technol. 2002, 30, 240–250. [Google Scholar] [CrossRef]
- Barber, A.R.; Henningsson, M.; Pamment, N.B. Acceleration of high gravity yeast fermentations by acetaldehyde addition. Biotechnol. Lett. 2002, 24, 891–895. [Google Scholar] [CrossRef]
- Stanley, G.A.; Hobley, T.J.; Pamment, N.B. Effect of acetaldehyde on Saccharomyces cerevisiae and Zymomonas mobilis subjected to environmental shocks. Biotechnol. Bioeng. 1997, 53, 71–78. [Google Scholar] [CrossRef]
- Vriesekoop, F.; Pamment, N. Acetaldehyde addition and pre-adaptation to the stressor together virtually eliminate the ethanol-induced lag phase in Saccharomyces cerevisiae. Lett. Appl. Microbiol. 2005, 41, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Vriesekoop, F.; Barber, A.R.; Pamment, N.B. Acetaldehyde mediates growth stimulation of ethanol-stressed Saccharomyces cerevisiae: Evidence of a redox-driven mechanism. Biotechnol. Lett. 2007, 29, 1099–1103. [Google Scholar] [CrossRef]
- Hucker, B.; Vriesekoop, F. Acetaldehyde stimulates ethanol-stressed Saccharomyces cerevisiae, grown on various carbon sources. Folia Microbiol. 2008, 53, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Vriesekoop, F.; Haass, C.; Pamment, N.B. The role of acetaldehyde and glycerol in the adaptation to ethanol stress of Saccharomyces cerevisiae and other yeasts. FEMS Yeast Res. 2009, 9, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, H. A colorimetric estimation of reducing sugars in potatoes with 3,5-dinitrosalicylic acid. Potato Res. 1973, 16, 176–179. [Google Scholar] [CrossRef]
- Hobley, T.J.; Pamment, N.B. Liquid injection gas chromatography and the ALDH assay overestimate free acetaldehyde in complex fermentation medium. Biotechnol. Tech. 1997, 11, 39–42. [Google Scholar] [CrossRef]
- Moon, J.-H.; Lee, H.-J.; Park, S.-Y.; Song, J.-M.; Park, M.-Y.; Park, H.-M.; Sun, J.; Park, J.-H.; Kim, B.Y.; Kim, J.-S. Structures of Iron-Dependent Alcohol Dehydrogenase 2 from Zymomonas mobilis ZM4 with and without NAD+ Cofactor. J. Mol. Biol. 2011, 407, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Horbach, S.; Strohhacker, J.; Welle, R.; de Graaf, A.; Sahm, H. Enzymes involved in the formation of glycerol-3-phosphate and the by-products dihydroxyacetone and glycerol in Zymomonas mobilis. FEMS Microbiol. Lett. 1994, 120, 37–44. [Google Scholar] [CrossRef]
- Bolhuis, H.; Martín-Cuadrado, A.B.; Rosselli, R.; Pašić, L.; Rodriguez-Valera, F. Transcriptome analysis of Haloquadratum walsbyi: Vanity is but the surface. BMC Genom. 2017, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jeckelmann, J.-M.; Erni, B. Transporters of glucose and other carbohydrates in bacteria. Pflügers Archiv—Eur. J. Physiol. 2020, 472, 1129–1153. [Google Scholar] [CrossRef]
- Bringer, S.; Finn, R.K.; Sahm, H. Effect of oxygen on the metabolism of Zymomonas mobilis. Arch. Microbiol. 1984, 139, 376–381. [Google Scholar] [CrossRef]
- Martien, J.I.; Hebert, A.S.; Stevenson, D.M.; Regner, M.R.; Khana, D.B.; Coon, J.J.; Amador-Noguez, D. Systems-Level Analysis of Oxygen Exposure in Zymomonas mobilis: Implications for Isoprenoid Production. mSystems 2019, 4, e00284-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toh, H.; Doelle, H. Changes in the growth and enzyme level of Zymomonas mobilis under oxygen-limited conditions at low glucose concentration. Arch. Microbiol. 1997, 168, 46–52. [Google Scholar] [CrossRef]
- Krúče, R.; Auzin, L. An elevation of the molar growth yield of Zymomonas mobilis during aerobic exponential growth. Arch. Microbiol. 1997, 167, 167–171. [Google Scholar] [CrossRef]
- Snoep, J.L.; Arfman, N.; Yomano, L.P.; Westerhoff, H.V.; Conway, T.; Ingram, L.O. Control of the glycolytic flux in Zymomonas mobilis by glucose-6-phosphate dehydrogenase activity. Biotechnol. Bioeng. 1996, 51, 190–197. [Google Scholar] [CrossRef]
- Osman, Y.A.; Conway, T.; Bonetti, S.J.; Ingram, L.O. Glycolytic flux in Zymomonas mobilis: Enzyme and metabolite levels during batch fermentation. J. Bacteriol. 1987, 169, 3726–3736. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, H.; Rollin, C.; Guyonvarch, A.; Guerquin-Kern, J.-L.; Cocaign-Bousquet, M.; Lindley, N.D. Carbon-flux distribution in the central metabolic pathways of Corynebacterium glutamicum during growth on fructose. JBIC J. Biol. Inorg. Chem. 1998, 254, 96–102. [Google Scholar] [CrossRef]
- Garrigues, C.; Loubiere, P.; Lindley, N.D.; Cocaign-Bousquet, M. Control of the shift from homolactic acid to mixed-acid fermentation in Lactococcus lactis: Predominant role of the NADH/NAD+ ratio. J. Bacteriol. 1997, 179, 5282–5287. [Google Scholar] [CrossRef] [Green Version]
- Girbal, L.; Soucaille, P. Regulation of Clostridium acetobutylicum metabolism as revealed by mixed-substrate steady-state continuous cultures: Role of NADH/NAD ratio and ATP pool. J. Bacteriol. 1994, 176, 6433–6438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldrich, H.C.; McDowell, L.; Barbosa, M.F.; Yomano, L.P.; Scopes, R.K.; Ingram, O.L. Immunocytochemical localization of glycolytic and fermentative enzymes in Zymomonas mobilis. J. Bacteriol. 1992, 174, 4504–4508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalnenieks, U. Physiology of Zymomonas mobilis: Some Unanswered Questions. Adv. Microb. Physiol. 2006, 51, 73–117. [Google Scholar] [CrossRef]
- Hohmann, S. Osmotic Stress Signaling and Osmoadaptation in Yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roustan, J.L.; Sablayrolles, J.M. Modification of acetaldehyde concentration during alcoholic fermentation and effect on fermentation kinetics. J. Biosci. Bioeng. 2002, 93, 375–376. [Google Scholar] [CrossRef]
- Struch, T.; Neuss, B.; Bringer-Meyer, S.; Sahm, H. Osmotic adjustment of Zymomonas mobilis to concentrated glucose solutions. Appl. Microbiol. Biotechnol. 1991, 34, 518–523. [Google Scholar] [CrossRef]
- Tamarit, J.; Cabiscol, E.; Aguilar, J.; Ros, J. Differential inactivation of alcohol dehydrogenase isoenzymes in Zymomonas mobilis by oxygen. J. Bacteriol. 1997, 179, 1102–1104. [Google Scholar] [CrossRef] [Green Version]
- Pamment, N.B.; Hall, R.J.; Barford, J.P. Mathematical modeling of lag phases in microbial growth. Biotechnol. Bioeng. 1978, 20, 349–381. [Google Scholar] [CrossRef] [PubMed]
- Rolfe, M.D.; Rice, C.J.; Lucchini, S.; Pin, C.; Thompson, A.; Cameron, A.D.S.; Alston, M.; Stringer, M.F.; Betts, R.P.; Baranyi, J.; et al. Lag Phase Is a Distinct Growth Phase That Prepares Bacteria for Exponential Growth and Involves Transient Metal Accumulation. J. Bacteriol. 2011, 194, 686–701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.-P.; Xie, J.; Cheng, Y.; Zhang, Z.; Zhao, Y.; Qian, Y.-F. Response of Shewanella putrefaciens to low temperature regulated by membrane fluidity and fatty acid metabolism. LWT 2020, 117, 108638. [Google Scholar] [CrossRef]
- Bekers, M.; Vigants, A.; Laukevics, J.; Toma, M.; Rapoports, A.; Zikmanis, P. The effect of osmo-induced stress on product formation by Zymomonas mobilis on sucrose. Int. J. Food Microbiol. 2000, 55, 147–150. [Google Scholar] [CrossRef]
- Wang, W.-T.; Dai, L.-C.; Wu, B.; Qi, B.-F.; Huang, T.-F.; Hu, G.-Q.; He, M.-X. Biochar-mediated enhanced ethanol fermentation (BMEEF) in Zymomonas mobilis under furfural and acetic acid stress. Biotechnol. Biofuels 2020, 13, 10–28. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Electron Acceptors (ea) | Lag Time (h) | Δ Lag Time (h) | lag Time (as % Stress) | µ (h−1) ea 2 | µea/µstress 3 | Final Population, Xea (Cells mL−1) | Xea/Xstress 4 |
|---|---|---|---|---|---|---|---|
| Acetaldehyde (2 mM) | 3 | −6.0 | 67 | 0.36 | 1.05 | 3.4 × 108 | 1.36 |
| Propionaldehyde (2 mM) | 3 | −6.0 | 67 | 0.36 | 1.05 | 3.5 × 108 | 1.38 |
| Pyruvate (2 mM) | 7 | −2.0 | 22 | 0.35 | 1.02 | 2.7 × 108 | 1.08 |
| Dihydroxyacetone (2 mM) | 8 | −1.0 | 11 | 0.34 | 1.00 | 2.6 × 108 | 1.04 |
| Oxygen (aerobic culture) | 0 | −9.0 | 100 | 0.09 | 0.26 | 3.2 × 107 | 0.13 |
| Stress | Without Acetaldehyde Addition | With Acetaldehyde Addition | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lag Time (h) | Specific Growth Rate, μ (h−1) | µstress/µ0 1 | Final Population, X (Cells mL−1) | Xstress/X0 2 | Added Acetaldehyde (mg L−1) | Lag Time (h) | Δ Lag Time (h) | Δ Lag Time (as % Control) | µacH 3 | µacH/µstress 4 | Final Population, XacH (Cells mL−1) | XacH/Xstress 5 | |
| Nonstressed control | 0 | 0.52 | NA 7 | 4.2 × 108 | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| Ethanol (50 g/L) | 9 | 0.34 | 0.66 | 2.5 × 108 | 0.6 | 90 | 2.5 | 6.5 | 72 | 0.38 | 1.13 | 1.2 × 108 | 1.68 |
| Temperature increase 20–40 °C | 3.5 | 0.35 | 0.67 | 2.1 × 107 | 0.05 | 90 | 3 | 0.5 | 12 | 0.35 | 1 | 8.0 × 107 | 3.8 |
| Osmotic | Irregular 6 | ND 8 | 3.9 × 107 | 0.1 | 9 | Stimulated | Irregular | 4.2 × 107 | 1.08 | ||||
| (250 g/L sorbitol) | 90 | Unchanged | Irregular | 4.1 × 107 | 1.05 | ||||||||
| Sodium chloride | Irregular | ND | 1.4 × 107 | 0.03 | 9 | Irregular | Irregular | 1.4 × 107 | 1 | ||||
| 175 mM | 90 | Irregular | Irregular | 1.4 × 107 | 1 | ||||||||
| Ammonium chloride | Irregular | ND | 2.1 × 107 | 0.04 | 9 | Stimulated | Irregular | 2.1 × 107 | 1 | ||||
| 175 mM | 90 | Stimulated | Irregular | 2.1 × 107 | 1 | ||||||||
| Sodium sulfate | 1.5 | 0.22 | 0.42 | 1.9 × 107 | 0.04 | 9 | 0.5 | 1 | 67 | 0.24 | 1.1 | 2.1 × 107 | 1.1 |
| 175 mM | 90 | 0.5 | 1 | 67 | 0.26 | 1.21 | 2.1 × 107 | 1.1 | |||||
| Acetate | 0 | 0.24 | 0.47 | 5.2 × 107 | 0.12 | 9 | 0 | 0 | 0 | 0.29 | 1.19 | 8.9 × 107 | 1.71 |
| 175 mM | 90 | 2 | 2 | NA | 0.29 | 1.19 | 8.0 × 107 | 1.54 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vriesekoop, F.; Pamment, N.B. Acetaldehyde Stimulation of the Growth of Zymomonas mobilis Subjected to Ethanol and Other Environmental Stresses: Effect of Other Metabolic Electron Acceptors and Evidence for a Mechanism. Fermentation 2021, 7, 80. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7020080
Vriesekoop F, Pamment NB. Acetaldehyde Stimulation of the Growth of Zymomonas mobilis Subjected to Ethanol and Other Environmental Stresses: Effect of Other Metabolic Electron Acceptors and Evidence for a Mechanism. Fermentation. 2021; 7(2):80. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7020080
Chicago/Turabian StyleVriesekoop, Frank, and Neville B. Pamment. 2021. "Acetaldehyde Stimulation of the Growth of Zymomonas mobilis Subjected to Ethanol and Other Environmental Stresses: Effect of Other Metabolic Electron Acceptors and Evidence for a Mechanism" Fermentation 7, no. 2: 80. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7020080