
Valorization of Bread Waste to a Fiber- and Protein-Rich Fungal Biomass


 , ,
, ,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Substrate
2.2. Microorganism
2.3. Batch and Fed-Batch Cultivation in Shake Flasks
2.4. Fed-Batch Cultivation in 4.5 L Bioreactors
2.5. Batch and Fed-Batch in a 26 L Bubble Column Bioreactor
2.6. Characterizations
2.7. Statistical Analysis
3. Results and Discussion
3.1. Substrate Analysis
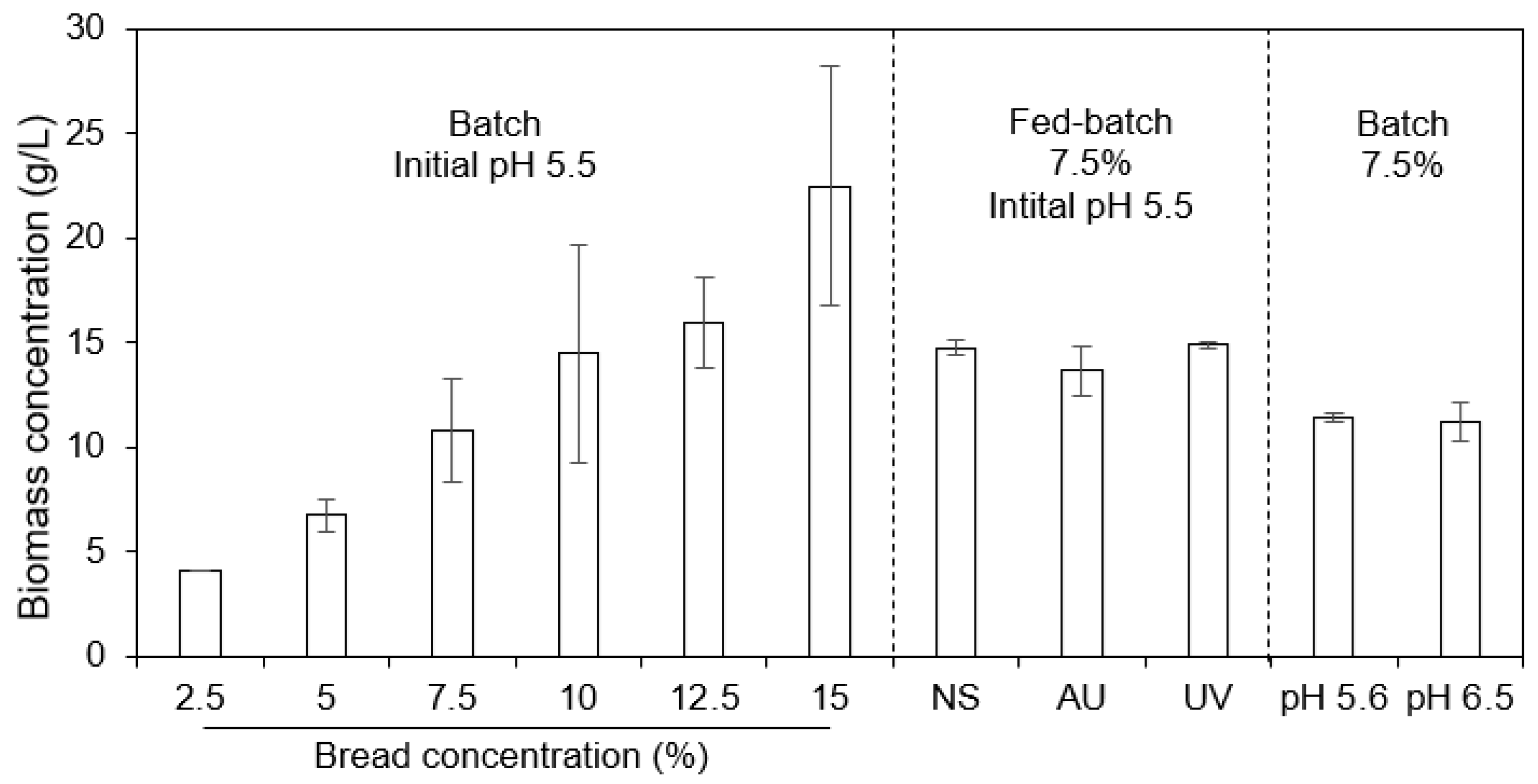
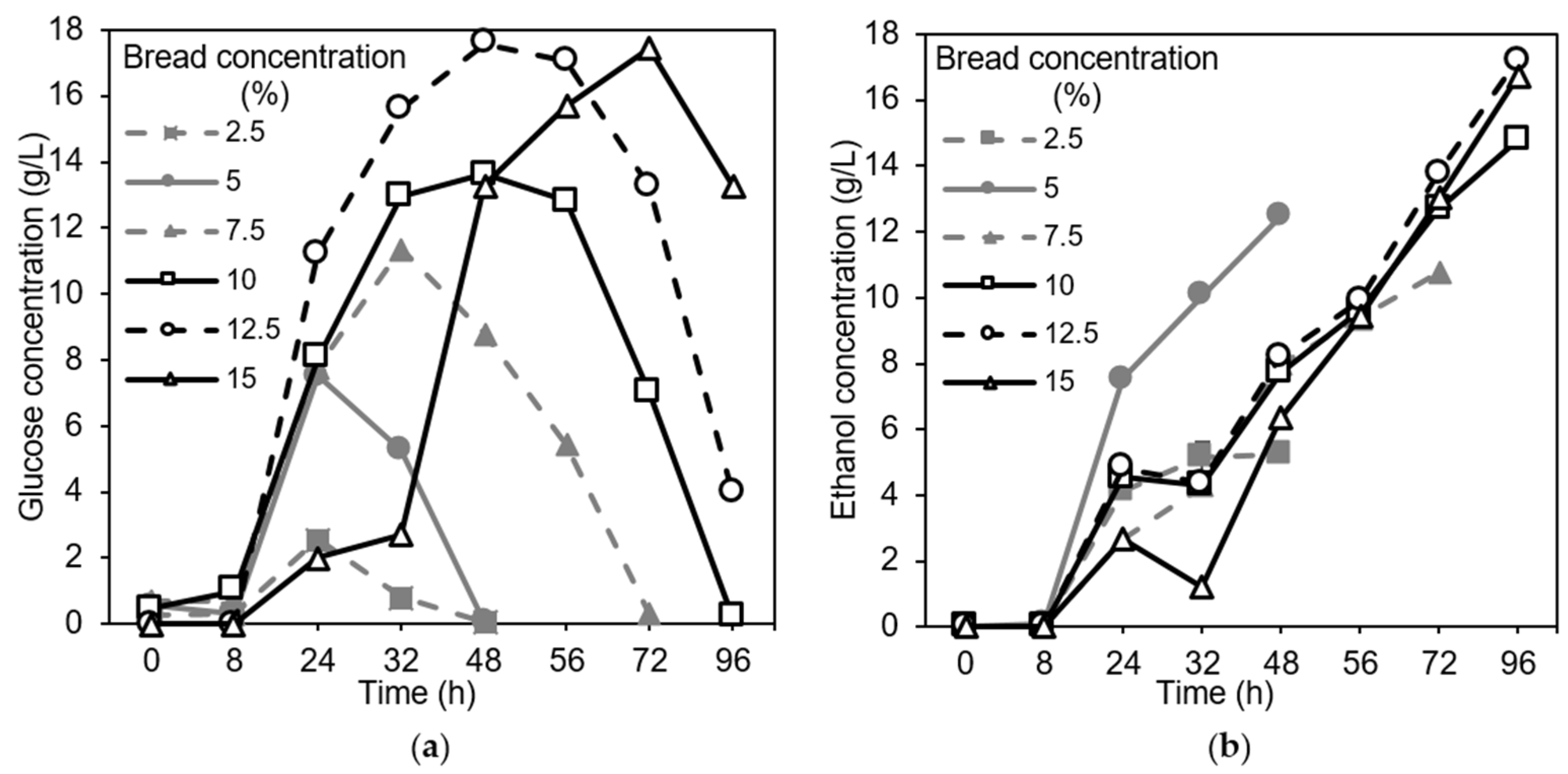
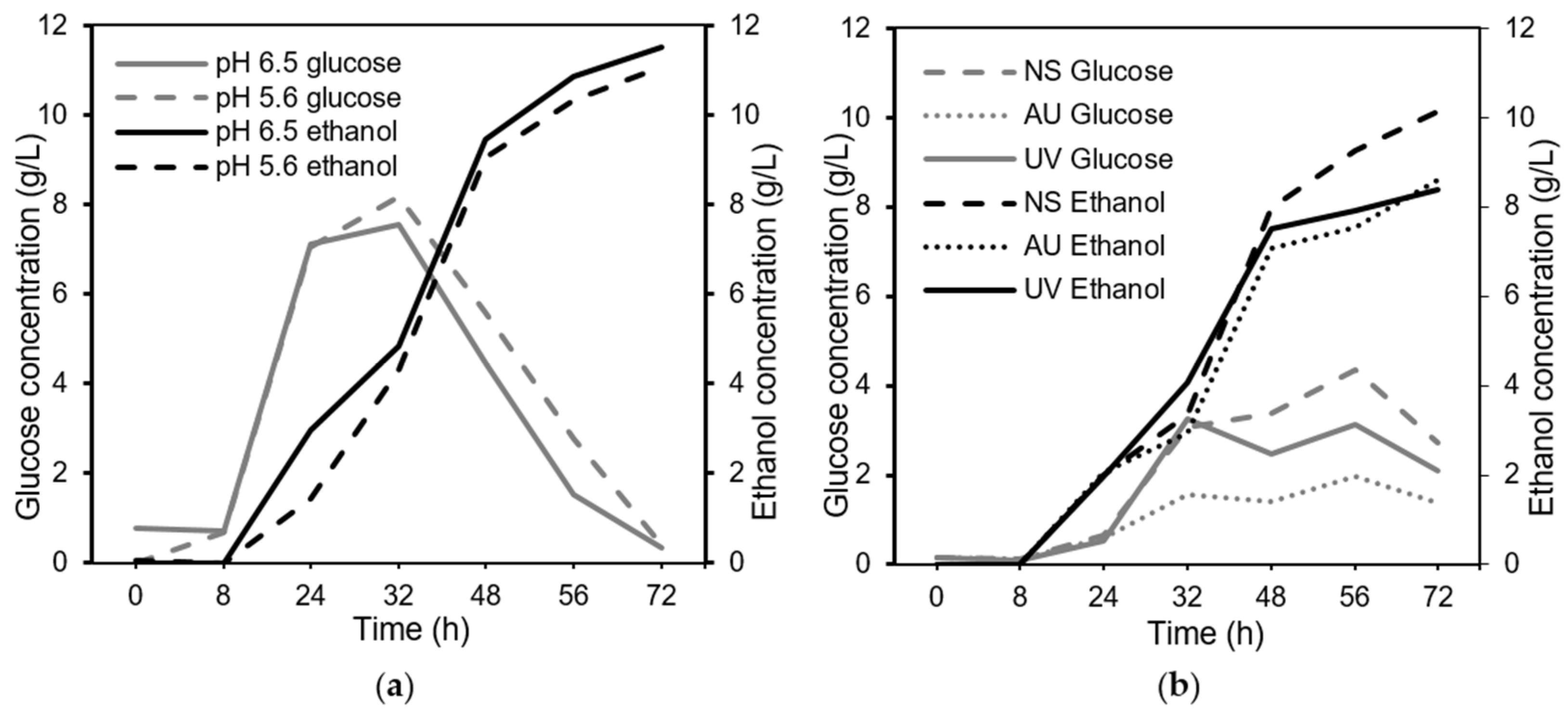
3.2. Effects of pH, Substrate Concentration, and Bread Sterilization on Fungal Growth in Shake Flasks
3.3. Cultivation in 4.5 L Bioreactors: Effects of Reactor Design and Aeration Rate
3.4. Fungal Biomass Production in a 26 L Bioreactor
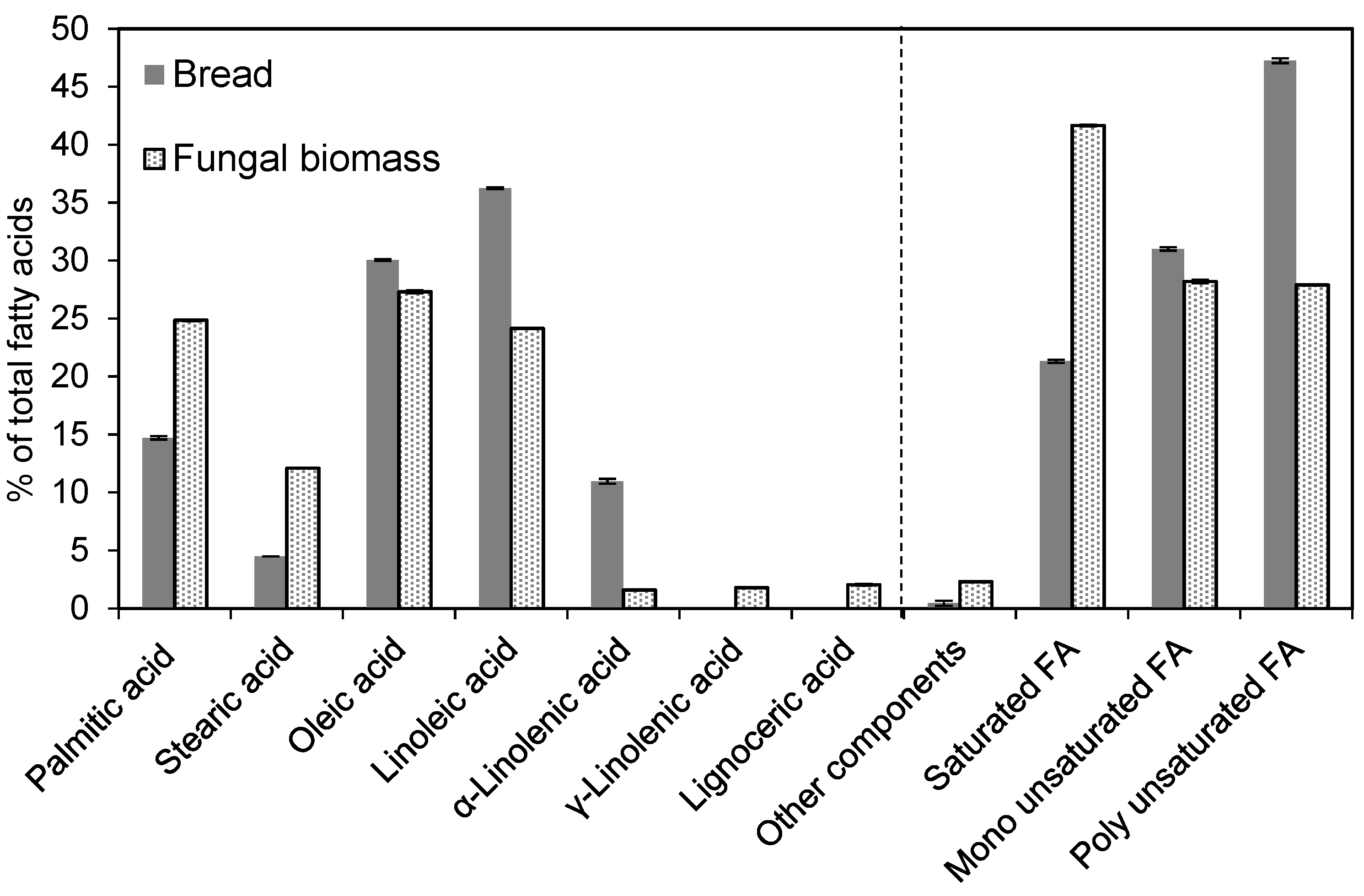
3.5. Nutritional Value of the Produced Biomass: Evaluating the Amino Acid and Fatty Acid Profiles
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- European Commission. Food Waste. Available online: https://ec.europa.eu/food/safety/food_waste_en (accessed on 28 February 2021).
- Brancoli, P.; Rousta, K.; Bolton, K. Life cycle assessment of supermarket food waste. Resour. Conserv. Recycl. 2017, 118, 39–46. [Google Scholar] [CrossRef]
- Souza Filho, P. Fungi-based biorefinery model for food industry waste: A progress towards circular economy. In Skrifter Från Högskolan i Borås; Högskolan i Borås: Borås, Sweden, 2018. [Google Scholar]
- Wiebe, M.G. Myco-protein from Fusarium venenatum: A well-established product for human consumption. Appl. Microbiol. Biotechnol. 2002, 58, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Wiebe, M.G. QuornTM Myco-protein—Overview of a successful fungal product. Mycologist 1999, 18, 17–20. [Google Scholar] [CrossRef]
- Hosseini, S.M.; Khosravi-Darani, K.; Mohammadifar, M.A.; Nikoopour, H. Production of Mycoprotein by Fusarium venenatum Growth on Modified Vogel Medium. Asian J. Chem. 2009, 21, 4017–4022. [Google Scholar]
- Denny, A.; Aisbitt, B.; Lunn, J. Mycoprotein and health. Nutr. Bull. 2008, 33, 298–310. [Google Scholar] [CrossRef]
- Dunlop, M.V.; Kilroe, S.P.; Bowtell, J.L.; Finnigan, T.J.A.; Salmon, D.L.; Wall, B.T. Mycoprotein represents a bioavailable and insulinotropic non-animal-derived dietary protein source: A dose–response study. Br. J. Nutr. 2017, 118, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Finnigan, T.J.A. 13—Mycoprotein: Origins, production and properties. In Handbook of Food Proteins; Williams, P.A., Ed.; Woodhead Publishing: Cambridge, UK, 2011; pp. 335–352. [Google Scholar]
- Filho, P.F.S.; Nair, R.B.; Andersson, D.; Lennartsson, P.R.; Taherzadeh, M.J. Vegan-mycoprotein concentrate from pea-processing industry byproduct using edible filamentous fungi. Fungal Biol. Biotechnol. 2018, 5, 1–10. [Google Scholar] [CrossRef]
- Troiano, D.; Orsat, V.; Dumont, M. Status of filamentous fungi in integrated biorefineries. Renew. Sustain. Energy Rev. 2020, 117, 109472. [Google Scholar] [CrossRef]
- Benabda, O.; Kasmi, M.; Kachouri, F.; Hamdi, M. Valorization of the powdered bread waste hydrolysate as growth medium for baker yeast. Food Bioprod. Process. 2018, 109, 1–8. [Google Scholar] [CrossRef]
- Kawa-Rygielska, J.; Pietrzak, W.; Czubaszek, A. Characterization of fermentation of waste wheat-rye bread mashes with the addition of complex enzymatic preparations. Biomass Bioenergy 2012, 44, 17–22. [Google Scholar] [CrossRef]
- Srichuwong, S.; Fujiwara, M.; Wang, X.; Seyama, T.; Shiroma, R.; Arakane, M.; Mukojima, N.; Tokuyasu, K. Simultaneous saccharification and fermentation (SSF) of very high gravity (VHG) potato mash for the production of ethanol. Biomass Bioenergy 2009, 33, 890–898. [Google Scholar] [CrossRef]
- Kłosowski, G.; Mikulski, D.; Czupryński, B.; Kotarska, K. Characterisation of fermentation of high-gravity maize mashes with the application of pullulanase, proteolytic enzymes and enzymes degrading non-starch polysaccharides. J. Biosci. Bioeng. 2010, 109, 466–471. [Google Scholar] [CrossRef]
- Kumar, J.V.; Mathew, R.; Shahbazi, A. Bioconversion of solid food wastes to ethanol. Analyst 1998, 123, 497–502. [Google Scholar] [CrossRef]
- Lennartsson, P.; Taherzadeh, M.; Edebo, L. Rhizopus. In Encyclopedia of Food Microbiology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 284–290. [Google Scholar]
- Ferreira, J.A.; Lennartsson, P.R.; Edebo, L.; Taherzadeh, M.J. Zygomycetes-based biorefinery: Present status and future prospects. Bioresour. Technol. 2013, 135, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahimi, F.; Khanahmadi, M.; Roodpeyma, S.; Taherzadeh, M. Ethanol production from bread residues. Biomass Bioenergy 2008, 32, 333–337. [Google Scholar] [CrossRef]
- Melikoglu, M.; Webb, C. 4—Use of Waste Bread to Produce Fermentation Products. In Food Industry Wastes: Assessment and Recuperation of Commodities; Kosseva, M.R., Webb, C., Eds.; Elsevier Science & Technology: San Diego, CA, USA, 2013; pp. 63–76. [Google Scholar]
- Melikoglu, M.; Lin, C.S.K.; Webb, C. Stepwise optimisation of enzyme production in solid state fermentation of waste bread pieces. Food Bioprod. Process. 2013, 91, 638–646. [Google Scholar] [CrossRef]
- Gmoser, R.; Sintca, C.; Taherzadeh, M.J.; Lennartsson, P.R. Combining submerged and solid state fermentation to convert waste bread into protein and pigment using the edible filamentous fungus N. intermedia. Waste Manag. 2019, 97, 63–70. [Google Scholar] [CrossRef]
- Svensson, S.E.; Ferreira, J.A.; Hakkarainen, M.; Adolfsson, K.H.; Zamani, A. Fungal textiles: Wet spinning of fungal microfibers to produce monofilament yarns. Sustain. Mater. Technol. 2021, 28, e00256. [Google Scholar] [CrossRef]
- Wikandari, R.; Millati, R.; Lennartsson, P.R.; Harmayani, E.; Taherzadeh, M.J. Isolation and Characterization of Zygomycetes Fungi from Tempe for Ethanol Production and Biomass Applications. Appl. Biochem. Biotechnol. 2012, 167, 1501–1512. [Google Scholar] [CrossRef]
- Mariotti, F.; Tomé, D.; Mirand, P.P. Converting Nitrogen into Protein—Beyond 6.25 and Jones’ Factors. Crit. Rev. Food Sci. Nutr. 2008, 48, 177–184. [Google Scholar] [CrossRef]
- Swedish Institute for Standards. Animal Feeding Stuffs—Determination of Amino Acids Content (ISO 13903:2005); Swedish Standards Institute: Stockholm, Sweden, 2005. [Google Scholar]
- Hames, B.; Scarlata, C.; Sluiter, A. Determination of Protein Content in Biomass; National Renewable Energy Laboratory: Golden, CO, USA, 2008; pp. 1–5. [Google Scholar]
- Filho, P.F.S.; Zamani, A.; Taherzadeh, M.J. Edible Protein Production by Filamentous Fungi using Starch Plant Wastewater. Waste Biomass Valorization 2018, 10, 2487–2496. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Production of ethanol and biomass from thin stillage byNeurospora intermedia: A pilot study for process diversification. Eng. Life Sci. 2015, 15, 751–759. [Google Scholar] [CrossRef] [Green Version]
- Papagianni, M. Fungal morphology and metabolite production in submerged mycelial processes. Biotechnol. Adv. 2004, 22, 189–259. [Google Scholar] [CrossRef]
- Ferreira, J. Spent sulphite liquor for cultivation of an edible Rhizopus sp. Bioresources 2012, 7, 173–188. [Google Scholar]
- Oliveira, M.D.S.; Feddern, V.; Kupski, L.; Cipolatti, E.; Badiale-Furlong, E.; de Souza-Soares, L.A. Changes in lipid, fatty acids and phospholipids composition of whole rice bran after solid-state fungal fermentation. Bioresour. Technol. 2011, 102, 8335–8338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibarruri, J.; Hernández, I. Valorization of cheese whey and orange molasses for fungal biomass production by submerged fermentation with Rhizopus sp. Bioprocess Biosyst. Eng. 2019, 42, 1285–1300. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Operation Mode | Configuration | Substrate Concentration (%) | Aeration (vvm *) | Time (h) |
|---|---|---|---|---|
| Shake flasks (200 mL working volume) | ||||
| Batch | 2.5, 5.0, 7.5, 10.0, 12.5, 15.0 | n/a | 48, 72, 96 | |
| Fed-batch | 7.5 a | 72 | ||
| Bioreactor (3.5 L working volume) | ||||
| Fed-batch | Airlift | 7.5 a | 1 | 72 |
| Bubble column | 7.5 a | 1 | 72 | |
| Bubble column pH control | 7.5 b | 1 | 72 | |
| Bubble column pH control | 4.5 c | 1.0, 1.42, 0.61 | 48 | |
| Bioreactor (20 L working volume) | ||||
| Batch | Bubble column | 7.5 | 1.0 | 48 |
| Batch | Bubble column pH control | 4.5 | 1.42 | 48 |
| Fed-batch | Bubble column pH control | 4.5 c | 1.42 | 48 |
| Component | g/100 g Dry Bread |
|---|---|
| Starch | 52.5 ± 3.5 |
| Ash | 2.4 ± 0.01 |
| Total Kjeldahl nitrogen | 2.3 ± 0.02 |
| Protein a | 12.8 ± 0.1 |
| Protein b | 13.4 ± 0.1 |
| Crude fat | 4.9 ± 0.1 |
| Reactor Design | pH Control | Aeration Rate (vvm) | Cultivation Time (h) | Biomass Concentration (g/L) | Ethanol Concentration (g/L) |
|---|---|---|---|---|---|
| Airlift | no | 1.0 | 72 a | 12.4 ± 2.8 | 8.1 ± 2.5 |
| Bubble column | no | 1.0 | 72 a | 12.3 ± 2.5 | 8.6 ± 0.6 |
| yes | 1.0 | 72 a | 10.9 ± 0.1 | 8.8 ± 3.5 | |
| yes | 0.61 | 48 b | 7.9 ± 0.1 | 8.8 ± 5.3 | |
| yes | 1.0 | 48 b | 7.2 ± 0.4 | 4.0 ± 1.4 | |
| yes | 1.42 | 48 b | 8.3 ± 0.5 | 1.5 ± 1.9 |
| Bread Concentration (%) | Mode | Aeration (vvm) | Biomass Concentration (g/L) |
|---|---|---|---|
| 7.5 | Batch, without pH control | 1.0 | 11.73 ± 1.6 |
| 4.5 | Batch, with pH control | 1.42 | 7.9 |
| 4.5 | Fed-batch, with pH control | 1.42 | 8.0 |
| Cultivation Scale | Bread Concentration (%) | Aeration Rate (vvm) | Protein (%) |
|---|---|---|---|
| Shake flasks | 2.5 | 27.4 a ± 1.8 | |
| 5 | 29.6 b ± 0.3 | ||
| 7.5 | n/a | 32.3 b ± 1.0 | |
| 10 | 32.0 b ± 0.6 | ||
| 12.5 | 31.3 b ± 1.0 | ||
| 15 | 30.0 b ± 1.4 | ||
| 4.5 L bioreactor | 4.5 | 0.61 | 35.8 b ± 1.8 |
| 4.5 | 1.0 | 33.0 b ± 1.4 | |
| 4.5 | 1.42 | 31.2 b ± 2.1 | |
| 26 L bioreactor | 4.5–7.5 | 1.0–1.42 | 30.7 b ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Svensson, S.E.; Bucuricova, L.; Ferreira, J.A.; Souza Filho, P.F.; Taherzadeh, M.J.; Zamani, A. Valorization of Bread Waste to a Fiber- and Protein-Rich Fungal Biomass. Fermentation 2021, 7, 91. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7020091
Svensson SE, Bucuricova L, Ferreira JA, Souza Filho PF, Taherzadeh MJ, Zamani A. Valorization of Bread Waste to a Fiber- and Protein-Rich Fungal Biomass. Fermentation. 2021; 7(2):91. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7020091
Chicago/Turabian StyleSvensson, Sofie E., Ludmila Bucuricova, Jorge A. Ferreira, Pedro F. Souza Filho, Mohammad J. Taherzadeh, and Akram Zamani. 2021. "Valorization of Bread Waste to a Fiber- and Protein-Rich Fungal Biomass" Fermentation 7, no. 2: 91. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7020091