1. Introduction
Coffee is the most consumed drink in the world; for instance, during 2018–2019, around 10 million tons of coffee grains were consumed [
1,
2]. However, the pest known as coffee berry borer beetle (
Hypothenemus hampei) has the largest negative economic impact to coffee farms, as it causes worldwide annual loses above USD 500 million, and severe infestation may result in 50% crop loss [
3]. Bored coffee beans (BCBs) lack the quality required for roasting and alternative uses aside from the extraction of oil must be sought [
4]. Coffee residues such as the outer shell, pulp, and spent coffee grounds have been used for microbial production of bioethanol, biodiesel, biogas, citric acid, gibberellic acid, and xylanase, amongst other products [
5]. These examples demonstrate the potential of BCB as a source of carbohydrates for the microbial production of industrial metabolites with high added value such as hyaluronic acid.
Hyaluronic acid (HA) is a linear glycosaminoglycan with alternating β-1,3 and β-1,4 glycosidic linkages [
6], whose viscoelastic properties, high moisture retention, and biocompatibility allow for a wide range of biomedical applications that include the treatment of osteoarthritis by viscosupplementation, ophthalmic surgery, cosmetic surgeries, controlled release of drugs, tissue engineering, moisturizing creams for the skin, wound healing, etc. The cost of HA ranges from 2000 to 60,000 USD/kg, depending on the application, purity, and molecular weight [
7,
8]. HA can be obtained from animal tissues such as rooster combs and bovine vitreous humor; however, the amount of tissues is scant, the risk of viral infections is high, and the purification steps are costly. This situation has spurred the interest in the microbial production of HA, from which yields are high, the production costs are lower, and the purification processes are more efficient [
9]. Some strains/species of Streptococci show a high production of HA, such as
Streptococcus zooepidemicus,
S. pyogenes,
S. equisimilis, and
S. thermophiles, and other genetically modified microorganisms such as
Bacillus subtilis,
Lactococcus lactis,
Escherichia coli, and
Lactobacillus acidophilus [
6,
10]. Particularly,
Streptococcus zooepidemicus has regulatory acceptance by the FDA in the USA and by the MHRA in the U.K., and is already used industrially [
6,
8]. However, the costs associated with the production of HA (mainly the sources of carbon and/or nitrogen for cultivation media) have lagged behind the commercial competitiveness of the microbial path [
11]. A plausible alternative to reduce the cost of bacterial cultivation is the use of agroindustrial residues, such as those from the coffee industry.
The goal of this study was to evaluate the production of HA by S. zooepidemicus from bored coffee beans. To the best of our knowledge, this is the first report on the production of HA from coffee residues.
2. Materials and Methods
2.1. Chemical Composition of Bored Coffee Beans
Bored coffee beans (BCBs,
Coffea arabica) were kindly donated by a coffee farmer from the village of Pluma Hidalgo in the state of Oaxaca, México. The beans were ground in a mill Pulvex® (Maren, CD México, México) (Pulvex
®) and sieved in a test sieve RX-29 (W.S. Tayler, Mentor, OH, USA); the particles retained in sieves 40 (0.42 mm) and 60 (0.25 mm) were used to determine the chemical composition and to carry out both the acid (AcH) and enzymatic hydrolysis (EnH). Ground beans were analyzed regarding moisture content [
12], ash content [
13], water extractives [
14], solvent extractives [
15], lignin [
16], holocellulose [
17], and cellulose [
18].
Streptococcus equi subsp.
zooepidemicus ATCC 35246 was purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA).
2.2. Hydrolysis of Bored Coffee Beans
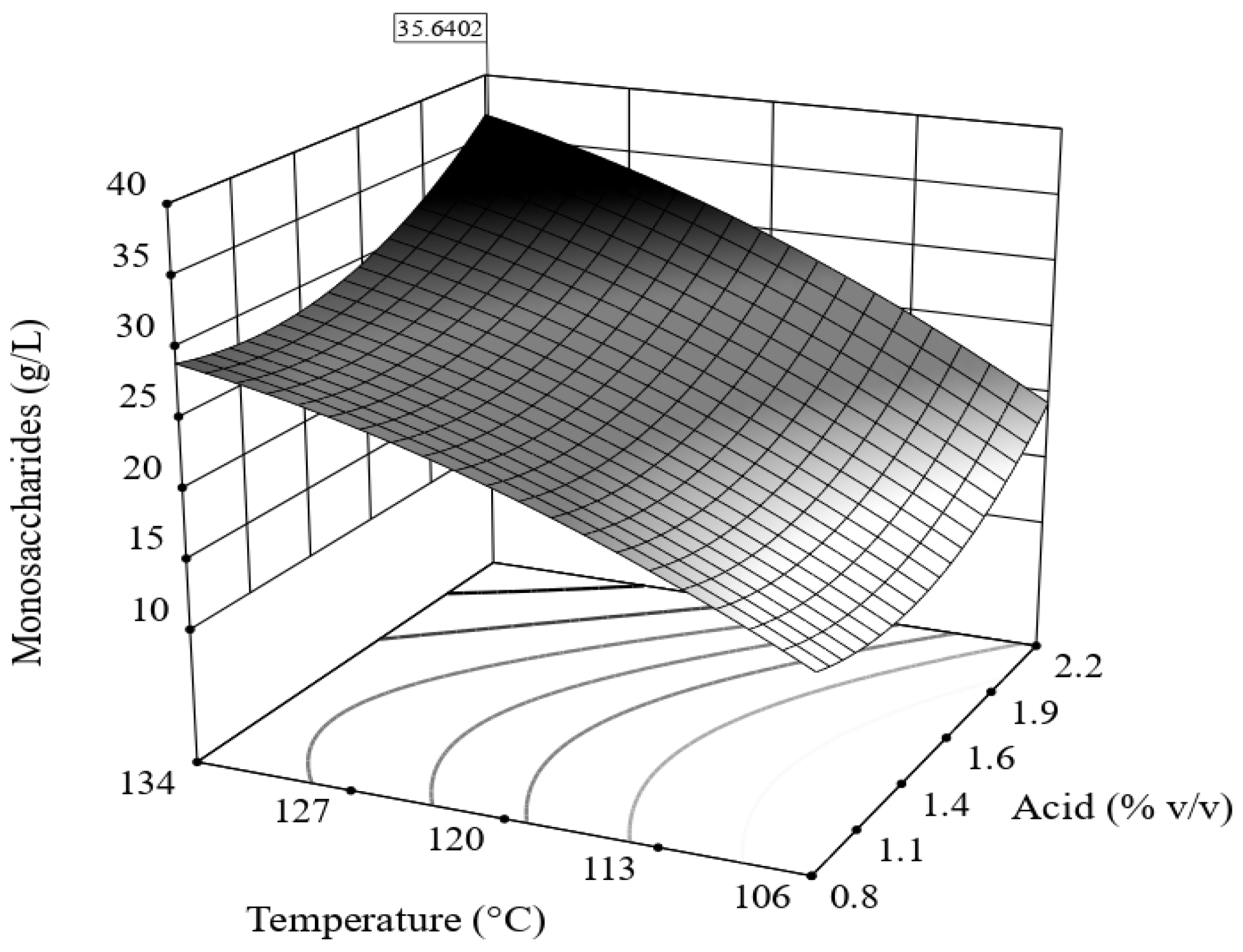
BCBs were hydrolyzed either by acid or enzymatically (10% load of solids i.e., 1 g of solids/10 mL solution) at conditions established with a central composite design (2 factors, 5 central points (replicates), and 4 axial points).
The acid hydrolyses (AcH) were carried out by varying the concentration of HCl (1–2% v/v) and the temperature (110–135 °C) for 45 min. After acid hydrolysis, samples were neutralized and then centrifuged at 10,000 rpm for 10 min. The supernatants were then passed first through a Sep-Pack C-18 (Waters®, Milford, MA, USA) cartridge and then through a 0.45 µm Millipore membrane. Glucose, xylose, arabinose, and mannose were quantified in an HPLC system Alliance e2695 (Waters®, Milford, MA, USA) provided with an RI detector (Waters® 2414), and an Aminex HPX-87C (300 mm × 7.8 mm, Bio-Rad, Hercules, CA, USA) column. HPLC-grade water was used as the eluent at 0.6 mL/min at 85 °C.
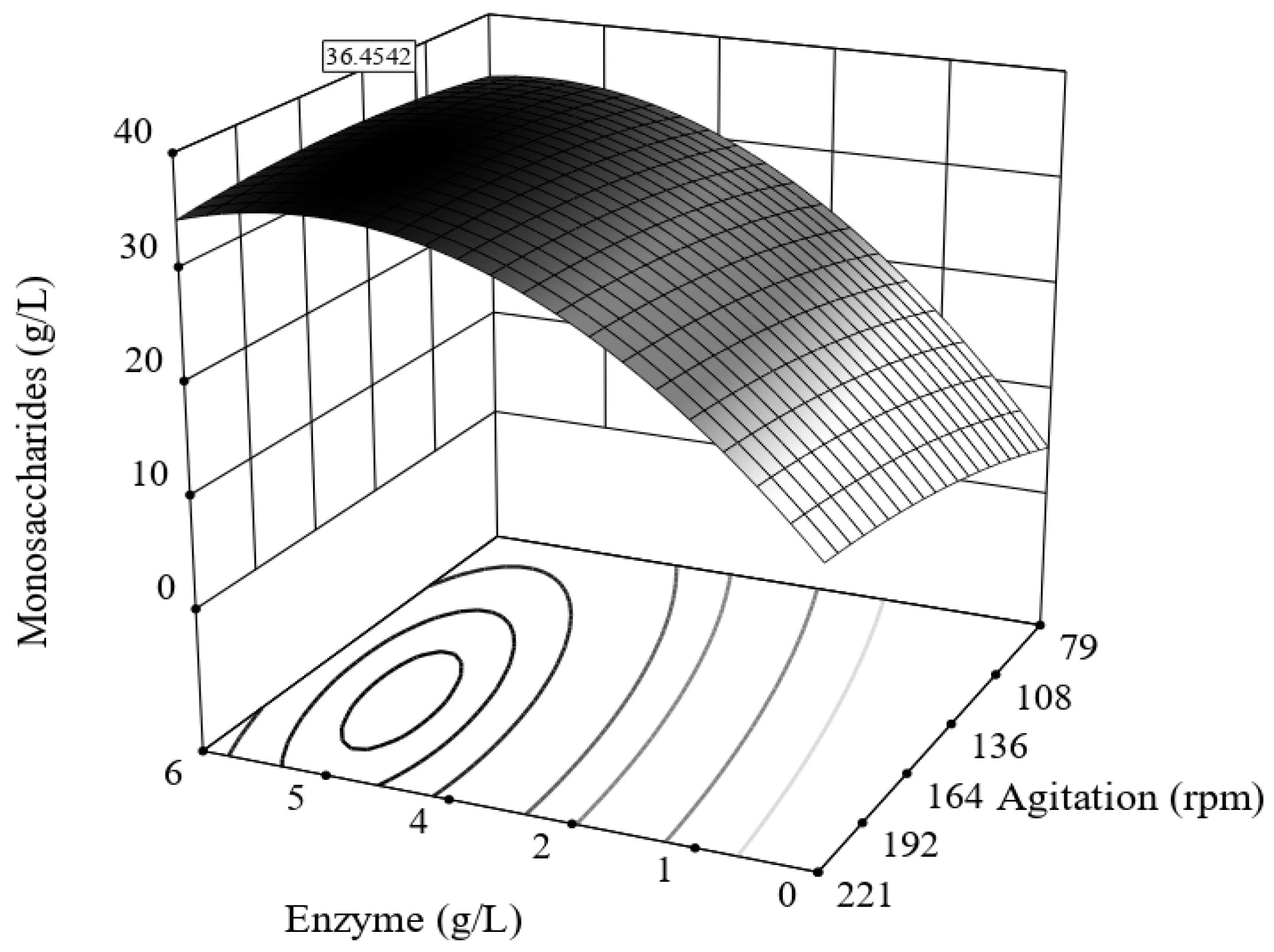
Enzymatic hydrolysis (EnH) was carried out at 50 °C and pH 5 (citrate buffer) for 20 h on an orbital shaker KS 4000i (IKA® Works, Inc., Wilmington, NC, USA) (KS 4000i, IKA®). Celluzyme (a mixture of cellulolytic enzymes from Trichoderma longibrachiatum, 90,000 CMCase/g activity) was employed. The concentration of enzyme (1–5 g/L buffer) and stirring (100–200 rpm) were the factors evaluated.
Once the enzymatic hydrolyses were conducted, the enzyme was thermally denatured; the samples centrifuged at 10,000 rpm for 10 min; the supernatant neutralized; and the concentrations of glucose, mannose, xylose, and arabinose were determined by HPLC, as described above.
The yields (δ) of monosaccharides were calculated as shown in Equation (1).
The coefficients 0.9 and 0.88 result from the addition of water to each monosaccharide (hexose and pentose, respectively) after hydrolysis.
The relationship between the selected factors and the response variable was obtained from the central composite design that can be described by a quadratic polynomial model Equation (2):
where β
0 is the constant term of the fitted model; β
1 and β
2, β
11 and β
22 are regression coefficients (linear and quadratic, respectively); β
12 is the interaction coefficient; V
1 and V
2 are the independent coded variables; and ε is the experimental error. Data analysis, surface responses, polynomial models, and ANOVA were all conducted with Design-Expert 11 software. The effects were considered statistically significant at
pvalue < 0.05.
To investigate the effect of organic extractives on the hydrolysis and fermentations, BCBs were acetone-extracted and then hydrolyzed either by acid or enzymatically under the best conditions dictated by the statistical model.
2.3. Post-Treatment of Acid Hydrolyzates
Acid hydrolyzates (AcH and AcHD) were ion-exchanged (250 g resin/Lhydrolyzates) with Amberlite IRA 96, (purchased from Sigma Aldrich, St. Louis, MO, USA) (Aldrich) in order to remove compounds such as hydroxymethyl furfural (HMF), furfural, and formic acid, which inhibit bacterial growth. Before and after ion-exchange treatment, the concentration of monosaccharides was quantified as above, whereas HMF, furfural, and formic acid were determined by HPLC provided with a diode array (Waters® 2998) and an Aminex HPX-87H column (300 mm × 7.8 mm, Bio-Rad), and run at 50 °C, using 5 mM H2SO4 at 0.6 mL/min as the eluent.
2.4. Microorganism and Culture Conditions
For preservation, Streptococcus equi subsp. zooepidemicus was periodically sub-cultured in brain-heart infusion (BHI) agar at 37 °C for 24 h.
Hyaluronic acid was biosynthesized by preparing a S. zooepidemicus inoculum as follows: a single colony was transferred to 200 mL BHI broth and then incubated at 37 °C at 200 rpm on an orbital shaker for 12 h. Later, this culture was inoculated (10% v/v) in the reactor production medium.
Batch cultures were formed in a 3 L bioreactor (Applikon®, JG Delft, Netherlands) (Applikon®) with a working volume of 2 L; the culture medium consisted of (g/L): monosaccharides in hydrolyzates, either acid or enzymatic (30); yeast extract (10); NaCl (2); K2HPO4 (2.5); and MgSO4×7H2O (1.5). A control cultivation was also performed using synthetic media with the same composition as above but with 30 g/L of glucose as the carbon source instead of hydrolyzates. Culture media were autoclaved for 15 min. Cultures were carried out at 37 °C, 300 rpm, pH was set constant at 7 (controlled with NaOH 5M during cultivation), with 1 vvm air flow, and 5% dissolved oxygen. All the runs were duplicated.
The concentration of biomass, HA, monosaccharides, and lactic acid were determined in samples as follows: The biomass content was determined by dry weight; first hyaluronic acid was separated from cells by treating culture samples with equal volumes of sodium dodecyl sulfate (0.1
w/
v) for 10 min [
19] and the mixture centrifuged at 10,000 rpm for 10 min. Then, the precipitate was washed twice with distilled water and centrifuged again at 10,000 rpm for 10 min, and then dried and weighed. The supernatant was used to quantify products (hyaluronic acid, monosaccharides, and organic acids) by HPLC. Hyaluronic acid was determined by following the methodology described by [
19] using an IR detector (Waters
® 2414), an Ultrahydrogel 2000 column (7.8 × 300 mm; Waters) at 70 °C, and NaNO
3 100 mM as the eluent at 0.8 mL/min. Monosaccharides were quantified as described in
Section 2.2, whereas LA was determined under the same conditions as furfural and formic acid described before.
2.5. Kinetic Parameters
Maxima specific growth rate (µ
max,1/h), HA, LA production (q
HA-max, q
LA-max, g/g h), and substrate consumption (q
G-max, q
M-max, g/g h) were calculated in the exponential phase for each time interval by taking the mean cell concentration (
, g/L), as shown in Equations (3)–(5). Yields (Y
LA/S, Y
HA/S, y Y
X/S, g/g) were calculated from the slope of the straight line obtained by plotting increasing amounts of products (LA − LA
0), (HA − HA
0), or biomass (X − X
0) as function of the substrate consumption (S − S
0) according to Equation (6). Productivities (g/L h) for HA (r
HA) and LA (r
LA) were calculated according to Equation (7) by dividing the respective variable (HA or LA) over fermentation time. Samples were obtained in 1 and 2 h periods. Equations (3)–(7):
where G, M, and X, are glucose, mannose, and biomass concentrations, respectively; Δ is the increased at each interval in the respective variable.
4. Discussion
The chemical composition of bored coffee beans is similar to that of green coffee as reported by others [
20,
21,
22]. According to our results, given the high carbohydrate polymers content (cellulose, galactomannans, and arabinogalactans) and the low lignin content, bored coffee bean (BCB) is an excellent candidate to be employed for the production of HA by microbial cultures. Notably, agricultural residues with higher lignin contents often require a pretreatment (physical, chemical, or biological) in order to remove lignin; examples of such materials include rice straw (12–24%), wheat straw (17–19%), corn straw (8%), and corn stubble (7–21%) [
23]. Moreover, water extractives, which contain carbohydrates, represent 25 wt % in bored coffee beans. Different studies have reported the presence of caffeine, phenolic compounds (chlorogenic acids), diterpenes (cafestol and khaweol), sucrose, glucose, and fructose in aqueous extracts of green coffee [
24,
25]. The differences found in the chemical composition between green coffee beans and bored coffee beans can be attributed to the differences in the environmental conditions and soil composition [
1].
Streptococcus zooepidemicus uses monosaccharides as the substrate for the production of HA; therefore, the polysaccharides from bored coffee beans need to be hydrolyzed. The majority of studies on the revalorization of industrial coffee residues have focused mainly on the pulp, outer skin, coffee parchment, and spent coffee grounds [
5]; however, there are no reports on the hydrolysis of bored coffee beans. In this study, acid and enzymatic hydrolyses of BCB and acetone-extracted BCB were carried out.
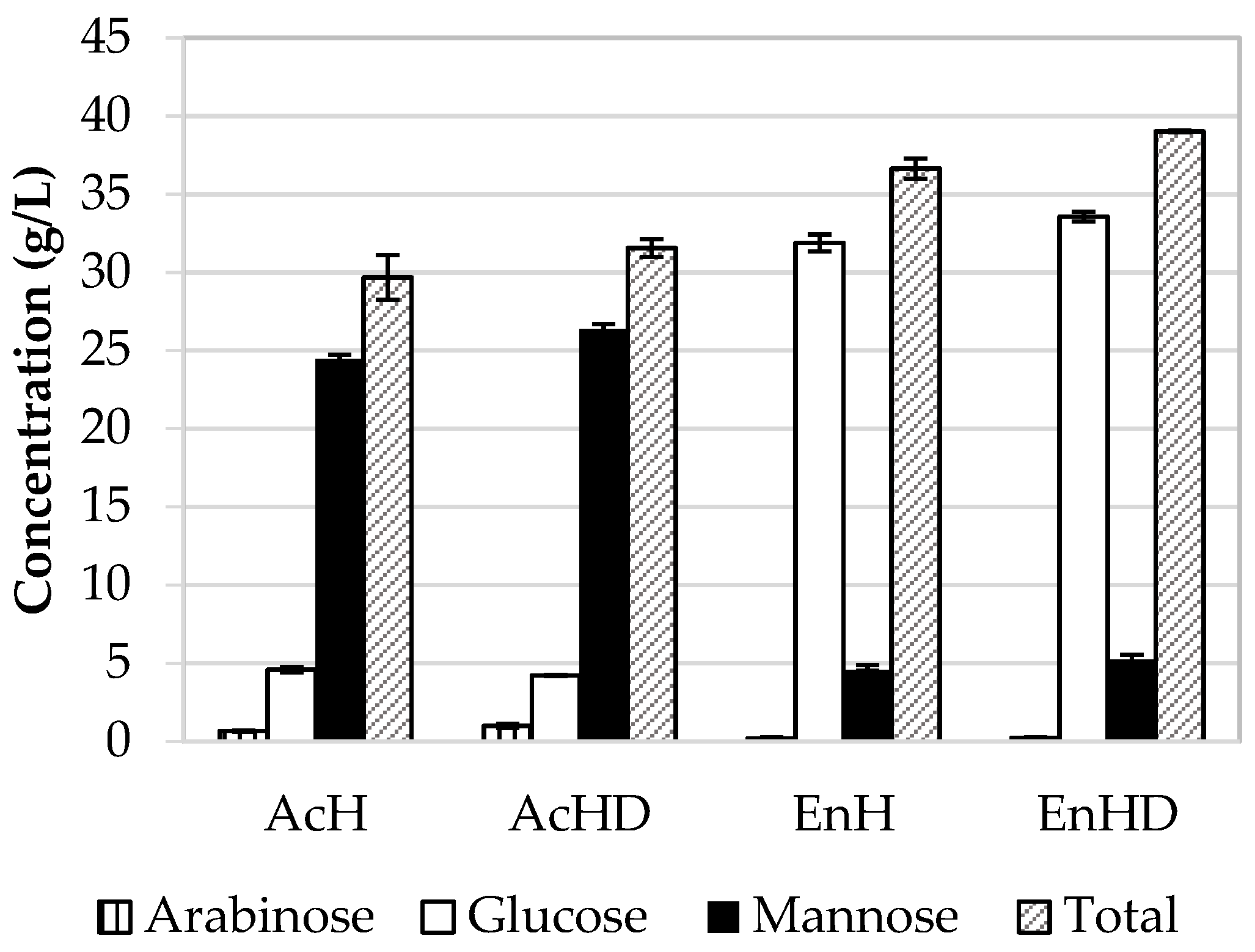
From acid hydrolysis (AcH), similar amounts of products were obtained as in non-extracted bored coffee beans (32.5 g/L,
Table 2) and acetone-extracted BCBs (31.6 ± 0.57 g/L). Juarez et al. [
26] found 33.4 g/L reductive sugar in the hydrolysis of spent coffee grounds; Kumar et al. [
27] obtained 30.29 g/L reductive sugars from sugar cane bagasse using HCl (2.5%
v/
v), and Pedroso et al. [
28] obtained 17.8 g/L sugars from the hydrolysis of rice husks using HCl (1.5%
v/
v).
The statistical model for acid hydrolysis (Equation (8)) suggests that a higher concentration of monosaccharides should have been obtained. This finding indicates that some monosaccharides were degraded, as HMF (0.103 g/L) and formic acid (3 g/L) were found. During acid hydrolysis, monosaccharides may degrade to furfurals and formic acid. Kupiainen et al. [
29] showed that increasing H
2SO
4 concentration from 1–5% in the hydrolysis of spent coffee grounds increased the concentrations of HMF and furfural (0.056 and 0.012 g/L, respectively). Taherzadeh et al. [
30] reported that the addition of 2 g/L of HMF decreased the specific growth rate of
S. cerevisiae 40% compared to a control with no HMF. Therefore, it is important to determine the hydrolysis conditions that maximize the recovery of monosaccharides and reduce the generation of furfurals and acetic and formic acids.
Notably, acid hydrolyzates require an ion-exchange treatment for the elimination of HMF, furfural, and formic acid to avoid the inhibition of S. zooepidemicus growth before HA production. Sugar retention in ion-exchange resin might be attributed to the interaction of OH groups with tertiary amines in the resin. Considering the loss of sugars, the concentration of monosaccharides was adjusted to the concentration of sugars before the ion-exchange resin treatment.
The differences between 3 and 5 g/L enzyme loads are minimal because the surface response is already at the maximum and it is more convenient from an economical point of view to keep the enzyme load at the medium point (3 g/L).
A higher quantity of monosaccharides was obtained from the enzymatic hydrolysis of acetone-extracted BCB (EnHD, 39 ± 0.60 g/L) in comparison to non-extracted bored coffee beans (EnH, 36.7 ± 0.65 g/L monosaccharides); the higher yield in acetone-extracted BCB might be associated with an increased exposure of polysaccharides in BCBs, as seen by Kwon et al. [
31], who reported an increased yield of glucose (from 15 to 18 g/L) after extracting spent coffee grounds. Niglio et al. [
32] obtained 35 g/L of sugars from the enzymatic hydrolysis of alkali-treated coffee silverskin. Seong et al. [
33] carried out enzymatic hydrolysis of HCl-treated spent coffee grounds and obtained 40.6 g/L of sugars. Notably, bored coffee beans and acetone-extracted bored coffee beans require no alkali or acid pretreatments and the obtained yields are consistent with the yields reported.
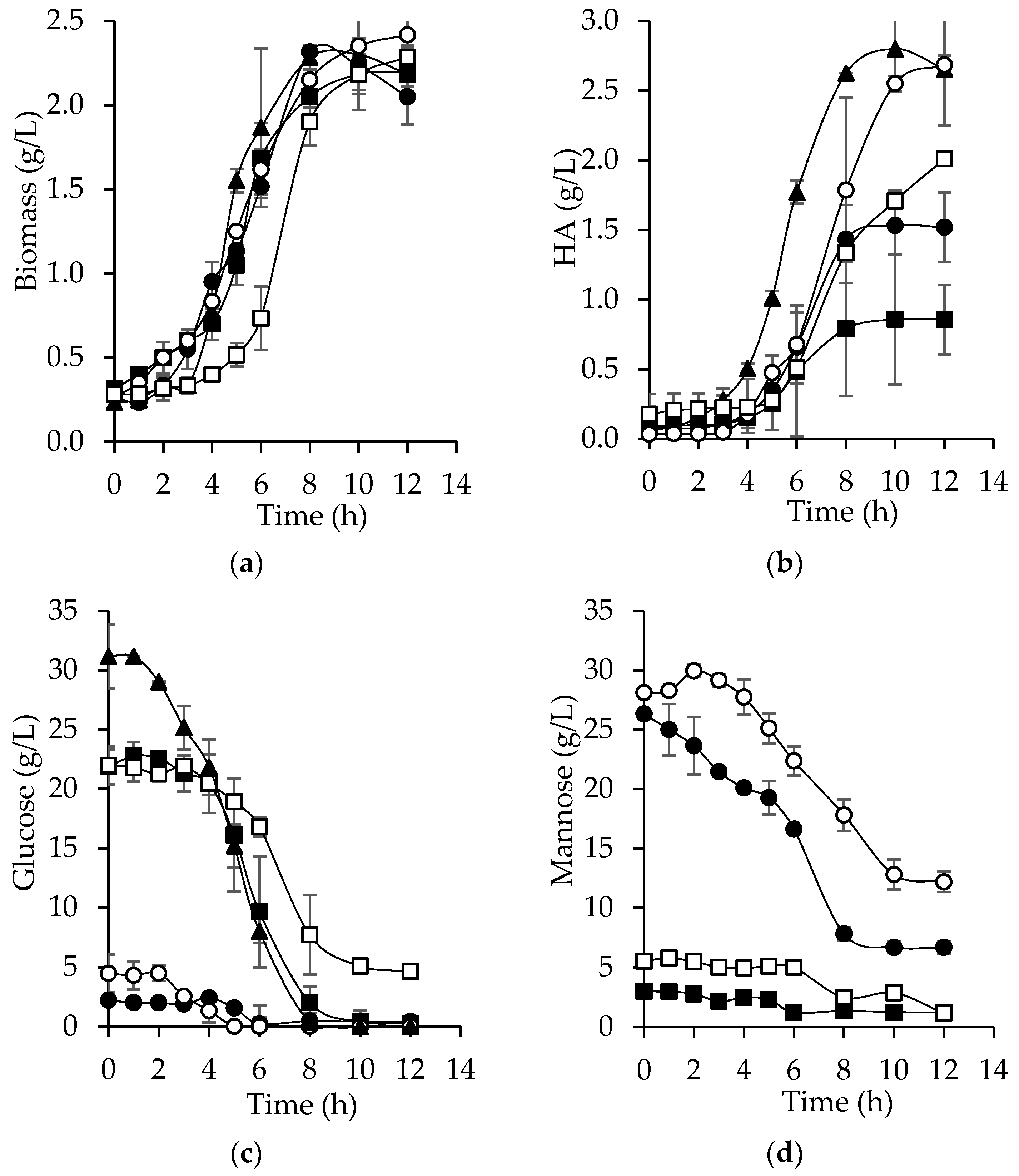
Both the acid and enzymatic hydrolyzates were used for the microbial production of HA. Bacterial growth was similar in all culture media (
Figure 4a). The stationary phase was reached at 8 h, except for EnHD. Interestingly, biomass yields (Y
X/S) were higher with both hydrolyzates compared to the synthetic medium, possibly due to the existence of unidentified substrates. It was expected that the highest concentration of HA would be reached with the enzymatic hydrolyzates because the main substrate in these is glucose. Furthermore, in the enzymatic hydrolysis, compounds such as furfurals and formic acid, which inhibit the bacterial growth, were absent. However, the highest concentration of HA (2.7 g/L) obtained with bored coffee beans hydrolyzates was achieved with AcHD at 12 h (
Figure 4b), similar to the amount obtained with the synthetic medium at 8 h (2.8 g/L). This behavior can be explained because there were probably other unidentified substrates in the AcHD that
S. zooepidemicus was able to convert into HA. Additionally, a higher concentration of HA was obtained using acetone-extracted bored coffee beans (AcHD and EnHD), achieving 1.8- and 2.3-fold increase in the HA amount, respectively, compared to un-extracted samples, probably because un-extracted bored coffee beans might contain compounds with antimicrobial activity such as quinic, malic, and chlorogenic acids, and caffeine [
34]. The main monosaccharide in acid hydrolyzates is mannose; in the enzymatic hydrolyzates, the main sugar is glucose. Glucose originates from the degradation of cellulose, while the lower amount of mannose might be due to the low activity of endogenous mannanase in cellulase preparations from
Trichoderma [
35], as cellulase from fungi exhibits relatively low levels of galactomannan degradation in softwoods [
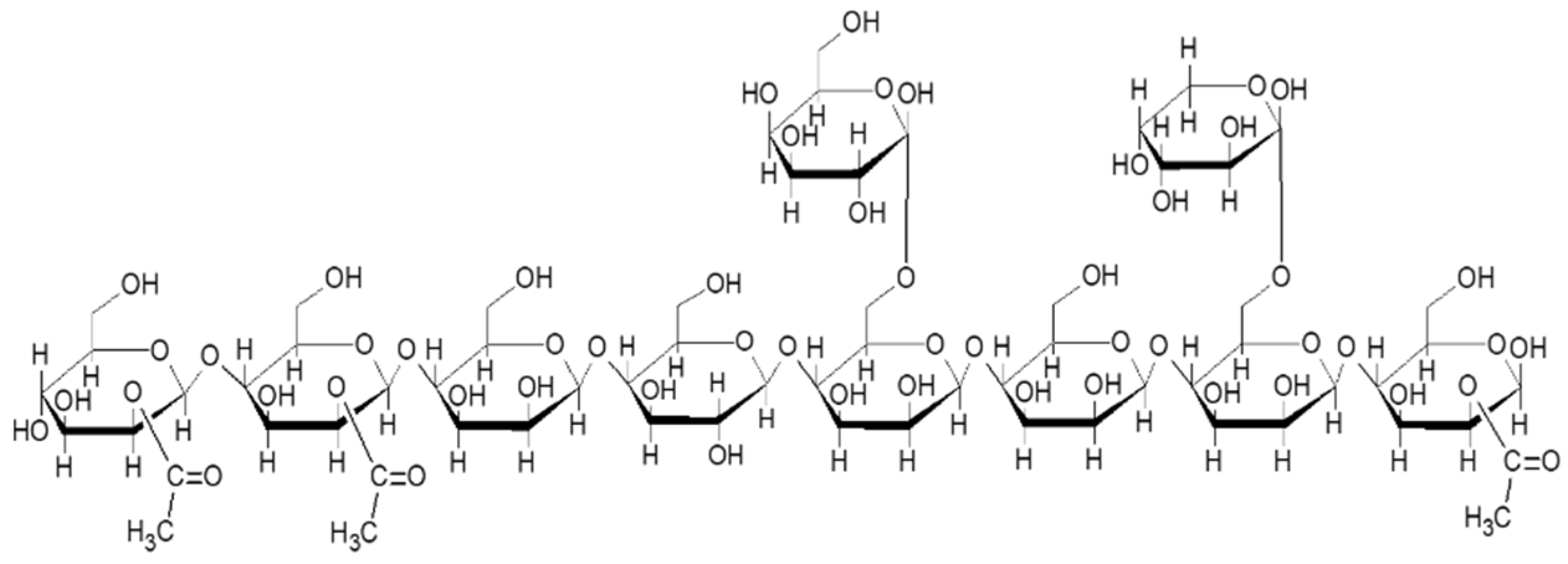
36]. Mannose originates from galactomannans, which, in green coffee, consist of a linear molecule composed of mannose residues linked by β-1,4 bonds with lateral galactose substituents with α-1,6 linkages, as seen in
Figure 5. Other randomly distributed elements were also found such as glucose linked in the main backbone with β-1, 4 bonds, arabinose as a lateral group, and acetyl groups (up to 11%) in mannose [
37].
Glucose consumption in the enzymatic hydrolyzates was higher and faster, as shown in
Figure 4c and
Table 5, compared to the consumption of mannose with acid hydrolyzates (
Figure 4d,
Table 5). Glucose is consumed more rapidly because mannose requires two enzymatic reactions to become fructose 6-P. First, it is converted into mannose 6-P by hexokinase, which has a lower affinity for mannose compared to glucose. Then, in order to be able to enter to glycolysis, it is converted to fructose 6-P by phosphomannose isomerase [
38].
Interestingly, at 10 h, the consumption of mannose stopped (
Figure 4d), which can be attributed to acetyl decorations that
S. zooepidemicus is unable to assimilate. Chen et al. [
39] demonstrated that by deacetylating xylan from corn baffles, the consumption of xylose improved and ethanol yields increased 10% during fermentation. Regarding arabinose, at the end of cultivation, no consumption was observed.
We obtained 2.0 g/L HA using EnHD hydrolyzates at 12 h cultivation. Productivity (r
HA,
Table 5) and the maximum specific rate of HA production (q
HA-max) behaved similarly to the concentration, although the highest substrate conversion to HA was obtained with AcHD (Y
HA/S = 0.13 g/g). The yields of HA obtained with the hydrolyzates ranged between 0.04 and 0.13 g/g, which are comparable to those reported for this species (
Table 6). For instance, Rohit et al. [
6] reported 0.09 g/g (Y
HA/S) from palm sugar (Palmyra palm sugar), whereas Pan et al. [
7] reported a 0.06 g/g yield from sugarcane molasses. Moreover, the maximum specific rate of HA production (q
HA-max) was obtained with acetone-extracted acid hydrolyzates (0.27 g/g h), which is 2.4-fold higher than the value reported by Mohan et al. [
40] with 30 g/L glucose (0.11 g/g h).
The production of HA obtained in this work is compared to those obtained in other studies in
Table 6. It can be seen that the amount of HA obtained in this work was slightly higher without considering that the other reports employed a higher initial concentration of sugars, which demonstrates the potential of bored coffee beans for the production of HA.
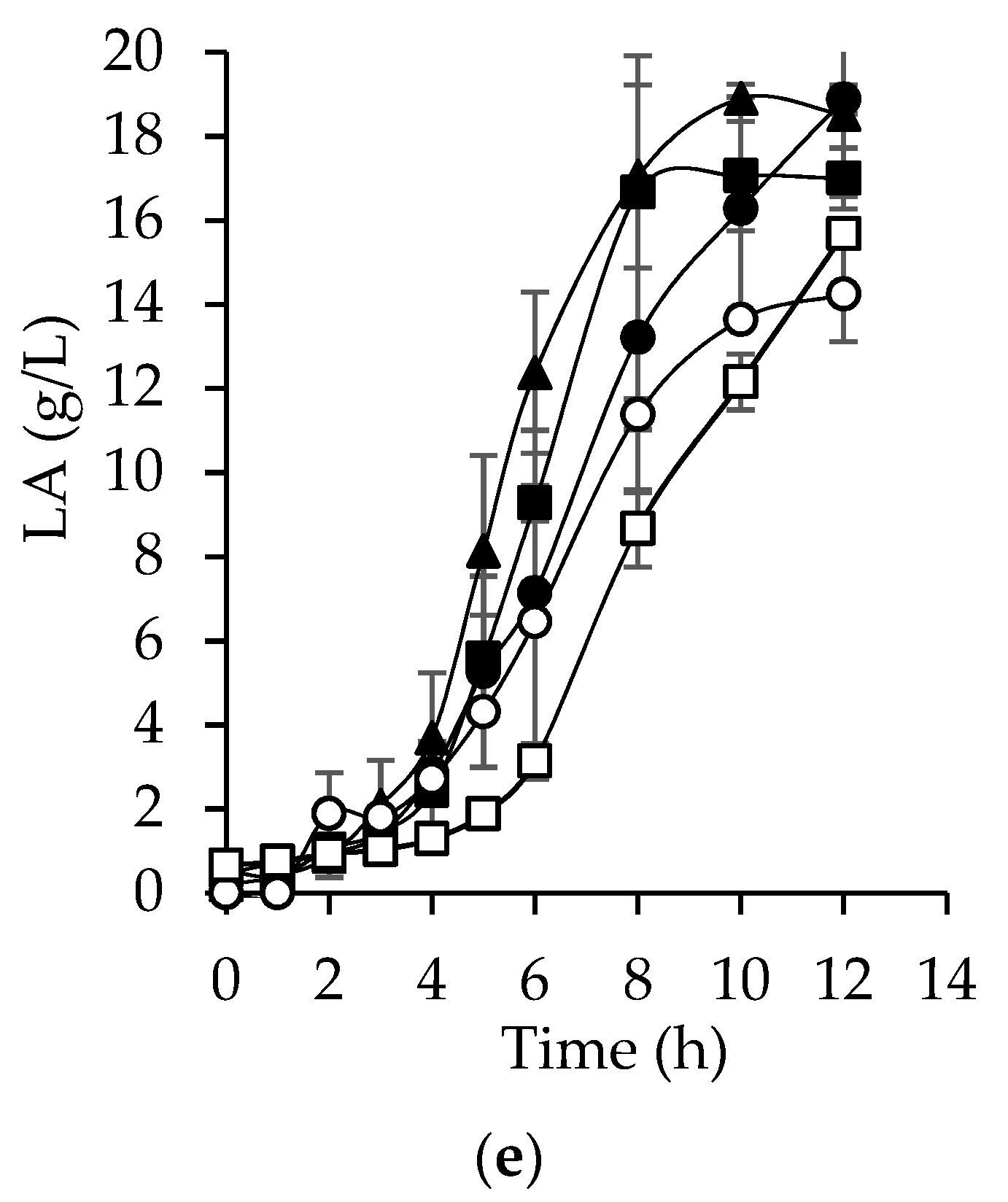
Lactic acid (LA) is a primary metabolite produced simultaneously with HA by
S. zooepidemicus. Higher production of LA (
Figure 4e), productivity (r
LA,
Table 5), and specific rate (q
LA) were obtained with EnHD (18.8 g/L), slightly higher than those obtained with the synthetic medium (18.6 g/L). However, the concentration of LA with all hydrolyzates was at least 14 g/L and yields (Y
LA/S) ranged between 0.60 and 0.79 g/g, as seen in
Table 5.
S. zooepidemicus is a homolactic bacteria that uses the majority of the substrate for LA synthesis [
42], which means that bacteria converted from 60% to 80% of the substrate into LA. The LA productivities obtained in this study (1.3 to 1.9 g/L h) agree with values obtained from other microorganisms such as
Rhizopus oryzae NRRL 395 and
Lactococcus lactis ATCC 19435, whose productivities are 1.62 and 1 g/L h, respectively [
43].
Importantly, LA is the main product from the metabolism of S. zooepidemicus. LA is a versatile metabolite with applications in different industries such as pharmaceuticals, cosmetics, chemicals, and food. The possibility of collecting both HA and LA simultaneously is attractive since their separation is easy, thus increasing the added value of BCBs.




 and
and 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}