
Growth of Non-Saccharomyces Native Strains under Different Fermentative Stress Conditions


Departamento de Investigación Agroambiental, Instituto Madrileño de Investigación y Desarrollo Rural, Agrario y Alimentario (IMIDRA), El Encín, A-2, km 38.2, 28805 Alcalá de Henares, Madrid, Spain
*
Author to whom correspondence should be addressed.
Fermentation 2021, 7(3), 124; https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030124
Submission received: 30 June 2021
/
Revised: 16 July 2021
/
Accepted: 17 July 2021
/
Published: 22 July 2021
(This article belongs to the Special Issue Enological Repercussions of Non-Saccharomyces Species 3.0)
Abstract
:The selection of yeast strains adapted to fermentation stresses in their winegrowing area is a key factor to produce quality wines. Twelve non-Saccharomyces native strains from Denomination of Origin (D.O.) “Vinos de Madrid” (Spain), a warm climate winegrowing region, were tested under osmotic pressure, ethanol, and acidic pH stresses. In addition, mixed combinations between non-Saccharomyces and a native Saccharomyces cerevisiae strain were practised. Phenotypic microarray technology has been employed to study the metabolic output of yeasts under the different stress situations. The yeast strains, Lachancea fermentati, Lachancea thermotolerans, and Schizosaccharomyces pombe showed the best adaptation to three stress conditions examined. The use of mixed cultures improved the tolerance to osmotic pressure by Torulaspora delbrueckii, S. pombe, and Zygosaccharomyces bailii strains and to high ethanol content by Candida stellata, S. pombe, and Z. bailii strains regarding the control. In general, the good adaptation of the native non-Saccharomyces strains to fermentative stress conditions makes them great candidates for wine elaboration in warm climate areas.
1. Introduction
Viticulture and wine elaboration are agricultural sectors greatly influenced by climate change [1]. The general influence of temperature increases has the immediate oenological consequence of an increased sugar content and, consequently, ethanol content. In addition, wines present a reduced acidity and increased pH, which implies a risk of undesired microbial proliferation and changes in the wine color, taste, and aroma [2,3,4]. While the external environment is continuously changing, the yeasts are exposed simultaneously to numerous stress conditions (oxidative, osmotic, and ethanol stress, among others). Yeast cells possess systems to response to stress conditions including the rapid synthesis of protective molecules and the activation of signal transduction pathways that induce secondary events as the reactivation of enzyme activities and the transcription of genes encodes factors with protective functions [5]. Therefore, a biotechnological approach to mitigate product depreciation due to climate change could be the selection of yeast strains that are able to respond to these stress situations without important viability loss [6].
Typically, the inoculation of a S. cerevisiae strain to carry out the vinification is a common practise in winemaking. However, many other species of yeasts belonging to non-Saccharomyces genera present oenological aptitude of interest. This group not only contributes to alcoholic fermentation but also are helpful to solve specific oenological problems [7,8], to modulate wine aroma [7,9] and to control the activity of undesired microbes [10,11]. Some non-Saccharomyces species are now commercially available as T. delbrueckii, L. thermotolerans, M. pulcherrima, S. pombe, and Pichia kluyveri [12]. The yeast species T. delbrueckii is probably considered the most suitable non-Saccharomyces yeast for wine elaboration [13]. It presents a good fermentative capacity and has positive aspects as low acetic acid and ethanol production, high glycerol and desirable aroma compounds release [13,14,15]. The fission yeast S. pombe has been recognized to improve some of the sensory parameters in wine, especially those linked to acidity control and wine color stability [16,17]. Two other genera abundantly found to be associated with grapes are Hanseniaspora and Candida, where C. stellata exhibit a positive impact in winemaking due to its fructophilic character, high glycerol, and extracellular enzyme production [18]. On the other hand, Hanseniaspora species are generally related to the increase of almost all acetate esters and some species stabilize the color of wine [19]. Most of non-Saccharomyces yeasts produce small changes in wine composition, while others prevent the addition of chemical compounds [20]. Thus, the main advantage in the use of L. thermotolerans is its inherent production of lactic acid, which avoid the addition of tartaric acid; in the same way, M. pulcherrima possess an antimicrobial effect by the secretion of the natural compound, pulcherrimin [21]. However, as long as non-Saccharomyces yeasts can confer beneficial characteristics to the resulting wines, most of them have a lesser fermentative capacity than S. cerevisiae. Consequently, controlled mixed fermentations that use more than one selected yeast strain play an increasingly important role in wine elaboration [22,23].
In the present work, phenotypic microarray technology was applied to test the growth capacity of microorganisms under different stressors using small volumes of culture. Twelve native non-Saccharomyces strains and one S. cerevisiae were studied alone or in mixed culture for better understanding of the behavior of these yeasts under different fermentative stress conditions given in a warm climate area (central Spain). Then, two non-Saccharomyces (S. pombe CLI 1085 and C. stellata CLI 920) and S. cerevisiae CLI 519 strains were assayed at higher volume under high osmotic pressure to analyze their fermentative kinetic and physicochemical changes associated with this stress situation.
2. Materials and Methods
2.1. Yeast Strains
Twelve non-Saccharomyces yeast strains (Table 1) isolated from spontaneous fermentations in Malvar grape variety have been included in this study. The biotechnological potential for elaboration of high-quality wines from D.O. “Vinos de Madrid” have helped to their selection [24,25]. The S. cerevisiae CLI 519 from IMIDRA collection was considered as control and selected on its good adaptative capacity to stress conditions inherent to wine fermentation in warm climates such as the Madrid winegrowing region (central Spain) [26].
2.2. Phenotypic Microarray Analysis
Phenotypic microarray (PM) technology enables growth of microorganism or consortia in 96‒well plate, each with a different substrate, stressor, or nutrient by providing a phenotypic characterization of the cultures [27,28]. This technology is commercially available through Biolog, Inc. (Hayward, CA, USA). The system allows for culturing in small volumes where cells growing up under aerobic or anaerobic conditions and their metabolic output is colorimetrically measured on a microplate reader or Omnilog unit (Biolog, Hayward, CA, USA). By comparing growth on plates, phenotypic variation between yeast strains can be detected [28,29]. PM technology uses a reporter system, which utilizes a redox-sensitive tetrazolium dye that can be reduced to a soluble purple formazan product, correlates with an increase in the metabolic rate of a cell, which is oxidizing a carbon source. Therefore, the presence of a chemical compound that it can oxidize by cell have the effect of increasing the cell metabolism and reducing the tetrazolium dye leads to a change from colorless into purple; this change is irreversible and cumulative. Instead, if the cell cannot oxidize a chemical, its metabolic rate does not rise, and no change of color takes place [30]. The cumulative amount of formazan is measured spectrophotometrically at 590 nm and is directly proportional to the number of metabolically active cells [31]. For PM analysis, each 96-well contained growth medium with a final concentration of 0.67% (w/v) yeast nitrogen base (YNB) and 6% (w/v) glucose in a total assay volume of 120 µL, supplemented with 2.6 µL of yeast nutrient mixture [26] and 0.2 µL dye D (Biolog). The final volume of growth medium was 30 µL using sterile distilled water, and inhibitory reagents were added as appropriate to maintain this volume. Thus, a stock solution of 80% (w/v) sorbitol adjusted to generate 20% and 30% (w/v) concentrations in a final volume of 120 µL. For ethanol, 5% (v/v), 8% (v/v) and 13% (v/v) were used to induce ethanol stress. Assays with pH used media YNB modified with phosphoric acid and then sterilized by filtration. Cell suspensions from inoculums were adjusted to a transmittance of 62 % (around 5 × 106 cells/mL) and 90 µL of a mixture of these cells and IFY buffer (Biolog) was inoculated to each well in the 96-well plate.
The Omnilog reader photographs the plates every 15 min, reflecting the metabolic output from dye conversion in each well. Finally, the percentage of redox signal intensity was determined by dividing the redox signal intensity under each stress condition and their non-stressed condition after 25 h incubation.
2.3. Small-Scale Fermentations
Sterile flasks of 120 mL were filled with YNB + glucose (87.5 g/L) medium (as control) and YNB + glucose (87.5 g/L) + 30% sorbitol medium (as stress condition) and were fermented at 30 °C. The fermenters were inoculated with 106 cells/mL of S. cerevisiae CLI 519 (p-Sc), S. pombe CLI 1085 (p-Sp) and C. stellata CLI 920 (p-Cs) in pure cultures. Mixed fermentation trials were simultaneously inoculated with the same concentration (106 cells/mL) of non-Saccharomyces cultures and S. cerevisiae strain. These pure and mixed cultures were tested as culture standard conditions (p-Sc, p-Sp, p-Cs, m-Sc/Sp, m-Sc/Cs) as for high osmotic pressure studies (p-Sc30%, p-Sp30%, p-Cs30%, m-Sc/Sp30%, m-Sc/Cs30%). The fermentation process was monitored daily by weight loss [32]. When the weight was constant, the fermentation was considered finished. Finally, samples were centrifuged and analyzed by HPLC.
2.4. Analytical Determinations
Chemical parameters as ethanol, glucose, acetic acid, glycerol and trehalose related to yeasts’ fermentative ability and their response to stressful conditions were analyzed in samples by HPLC. The Dionex Ultimate 3000 HPLC system (Thermo Scientific, Waltham, MA, USA) was equipped with a RefractoMax 520 refractive index (RI) detector and a Rezex ROA-Organic Acid H+ column (300 × 7.8 mm) at room temperature. Five mmol/L sulfuric acid was used as mobile phase, at a flow rate of 1 mL/min and a sample volume of 10 µL.
2.5. Data Statistical Analysis
Analysis of variance was carried out by ANOVA Tukey’s test to examine significant differences in fermentative parameters produced by mixed cultures of C. stellata CLI 920 and S. pombe CLI 1085 strains regarding their respective controls (pure cultures of S. cerevisiae with or without stress factor). The data were analyzed with SPSS Statistics 25 Software (SPSS Inc., Chicago, IL, USA).
3. Results
3.1. Fermentative Stress Tolerance of Non-Saccharomyces Yeast Strains in Aerobic (AE) and Anaerobic (AN) Conditions
The 12 native non-Saccharomyces strains from IMIDRA Collection were exposed to stress conditions present during the fermentation process: osmotic pressure, alcohol content, and pH. The resistance results are expressed in redox signal intensity units, which is the percentage of Biolog growth in a strain in comparison with this value for the same strain in non-stressed conditions.
The metabolic output of non-Saccharomyces strains has been analyzed in AE and AN condition in standard YNB culture medium for 96 h. Their metabolic activities are represented in Figure S1 (Supplementary Materials). All strains grew in both conditions at different percentages although a higher response is usually observed under aerobic situations. The species P. toletana, P. carsonii, S. pombe, and T. delbrueckii showed a parallel growth in both culture situations. Instead, the larger differences between AE and AN status were observed in the development of M. pulcherrima, W. anomalus and Z. bailii yeast strains.
3.1.1. Tolerance of Wine Yeast to Osmotic Stress
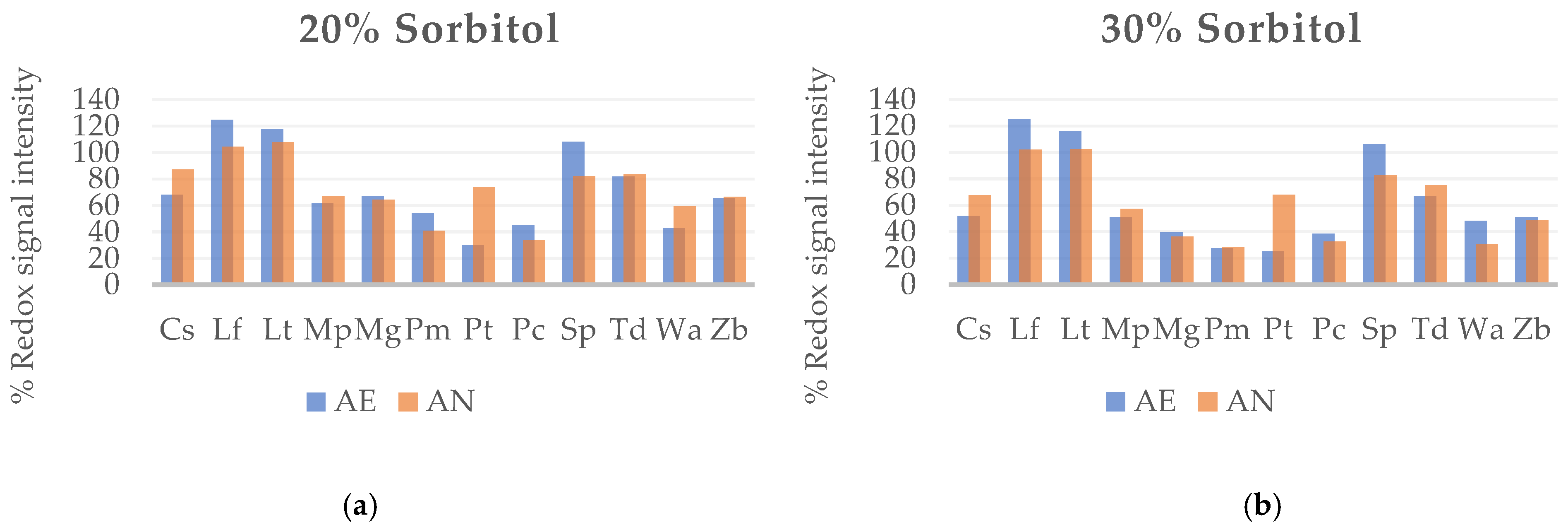
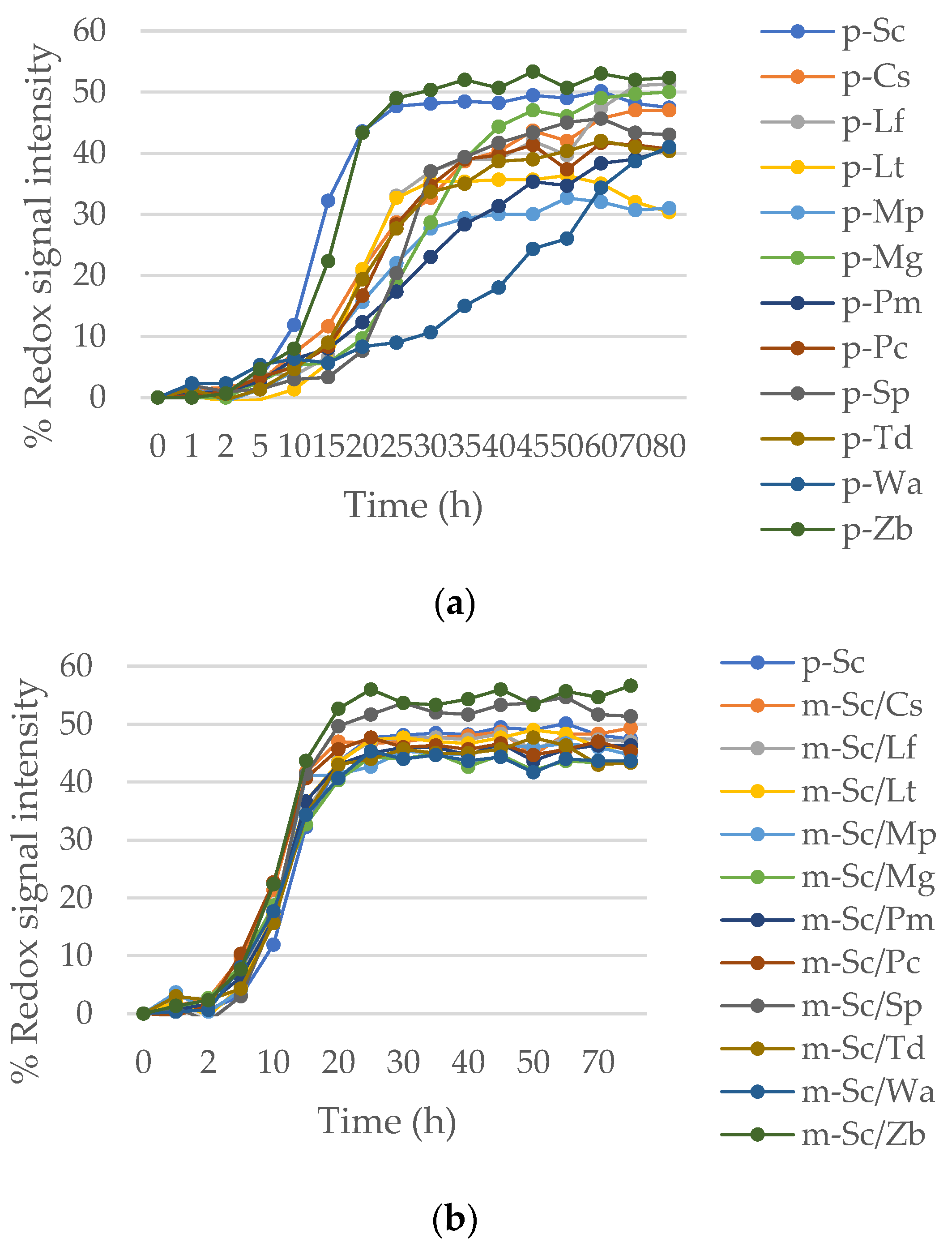
Sorbitol, a non-fermentable carbon source by yeasts, was employed to induce osmotic stress. In the presence of 20% sorbitol (Figure 1a), the strains L. fermentati (formerly Zygosaccharomyces fermentati), L. thermotolerans, and S. pombe exhibited the highest metabolic output between 100% and 125% under aerobic conditions. In anaerobiosis, the growth ratio of non-Saccharomyces is similar to in the presence of oxygen in general, though remain high by L. fermentati and L. thermotolerans. Strains C. stellata and P. toletana showed an opposed tendency over others and their growth is not extremely high, behave better on AN condition.
The osmophilic strains, L. fermentati, L. thermotolerans and S. pombe presented the highest resistance in AE and AN again at 30% sorbitol. The yeast strains that showed the worst adaptation to fermentative stress in this trial have been M. guilliermondii, P. membranaefaciens, P. carsonii, and W. anomalus with percentages between 27% and 48% (Figure 1b). As the same case observed before, C. stellata and P. toletana are better adapted to osmotic stress in AN condition. The strains M. pulcherrima and Z. bailii exhibited intermediate values around 50% with very similar behavior in AE and AN environment.
3.1.2. Tolerance of Wine Yeast to Ethanol
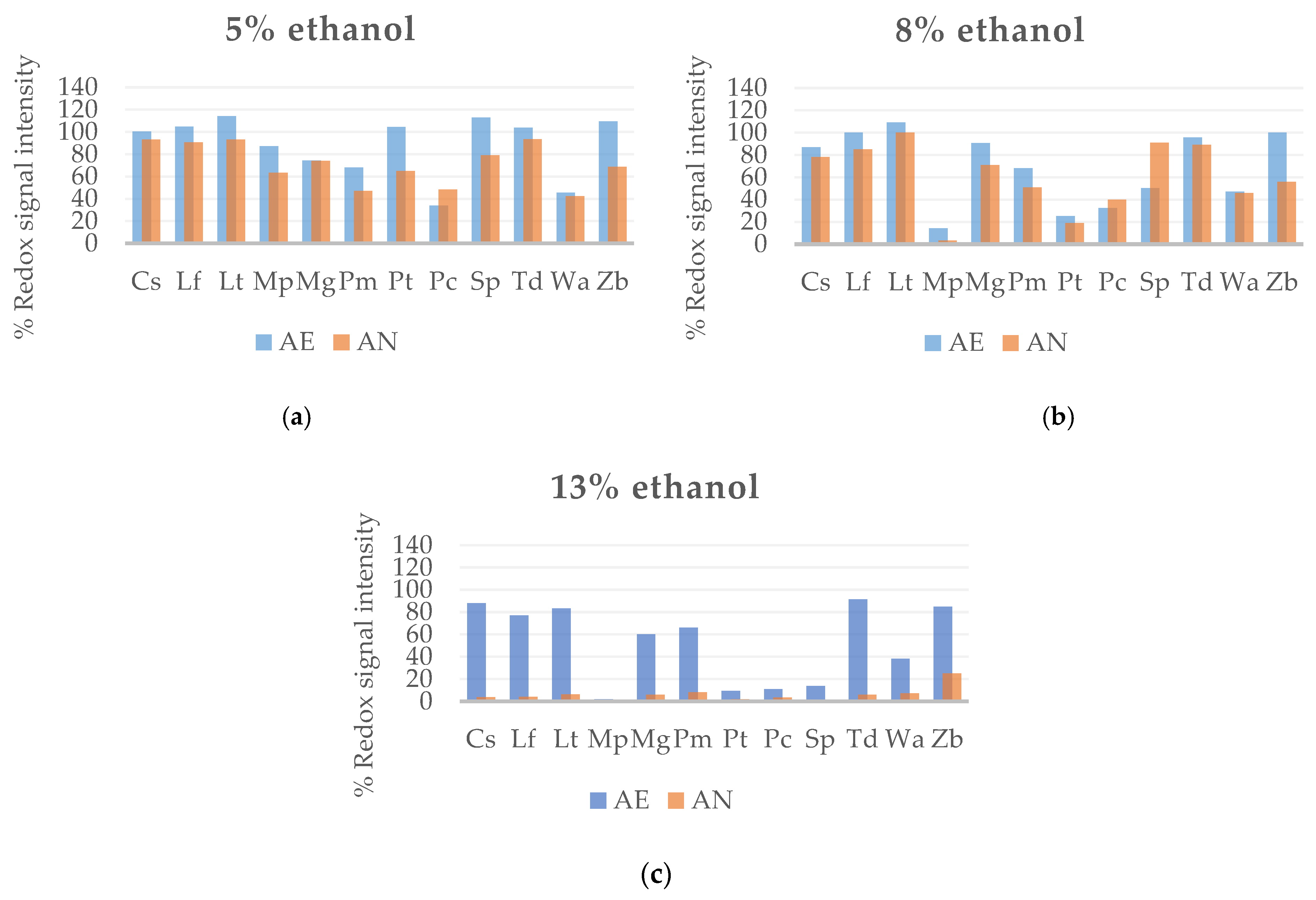
The resistance of yeasts to alcohol degree was tested at concentrations between 5% to 13% of ethanol to determine tolerant and sensitive yeast strains (Figure 2). The growth response of non-Saccharomyces strains was analyzed in 5% ethanol medium, and it can be observed that all of them grew well at this ethanol content, especially in the presence of oxygen (Figure 2a). Several yeast species showed growth ratios above 100%, as in case of C. stellata, L. fermentati, L. thermotolerans, P. toletana, S. pombe, T. delbrueckii, and Z. bailii. In contrast, P. membranaefaciens, P. carsonii, and W. anomalus presented the highest sensibility to 5% of ethanol (Figure 2a). When ethanol content was increased at 8% (Figure 2b), the development capacity of strains was reduced, being more affected the strains, M. pulcherrima and P. toletana. In this trial, P. carsonii and S. pombe grew better under anaerobic conditions. The growth capacity of strains was considerably reduced in the presence of 13% of ethanol, remaining very low or even non-existent in AN condition. Only Z. bailii reached values of 25% in Biolog units (Figure 2c).
3.1.3. Tolerance of Wine Yeast to Acidic pH
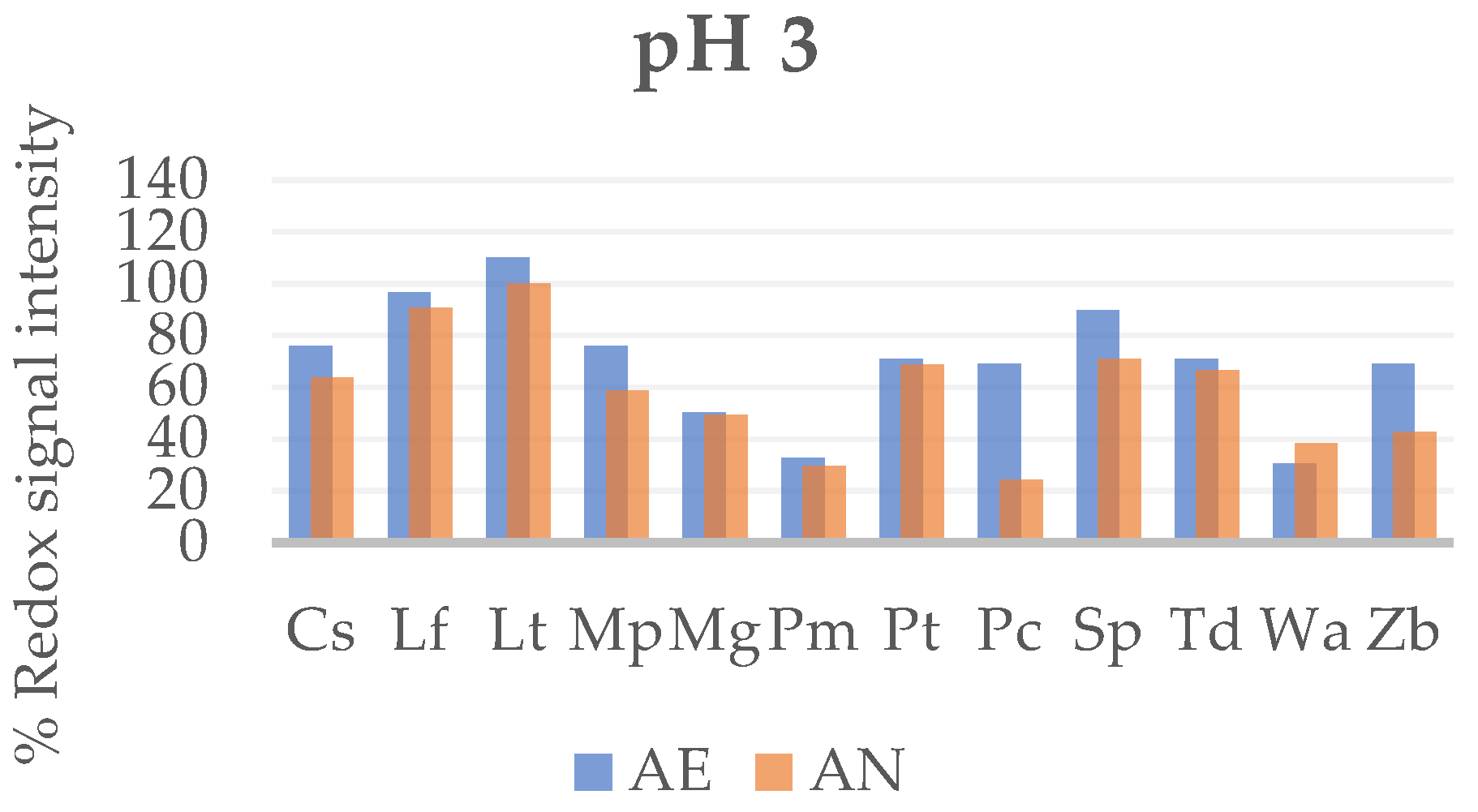
In general, there are not major differences for the strains between AE and AN condition as a whole (Figure 3). The strains with a good metabolic output were L. fermentati (96–90%), L. thermotolerans (110–100%) and S. pombe (90–71%); by contrast, P. membranaefaciens, P. carsonii in AN, and W. anomalus were more sensitive to stress caused by low pH (Figure 3), with values below 40%.
3.2. Phenotypic Response to Stress Conditions (30% Sorbitol and 13% Ethanol) in Mixed Cultures under Anaerobic Fermentation
The phenotypic response is studied for two stress situations, osmotic pressure at 30% of sorbitol and ethanol concentration at 13%. Mixed cultures are fermented in AN and 1:1 proportion of S. cerevisiae together with each of the twelve non-Saccharomyces included in this work.
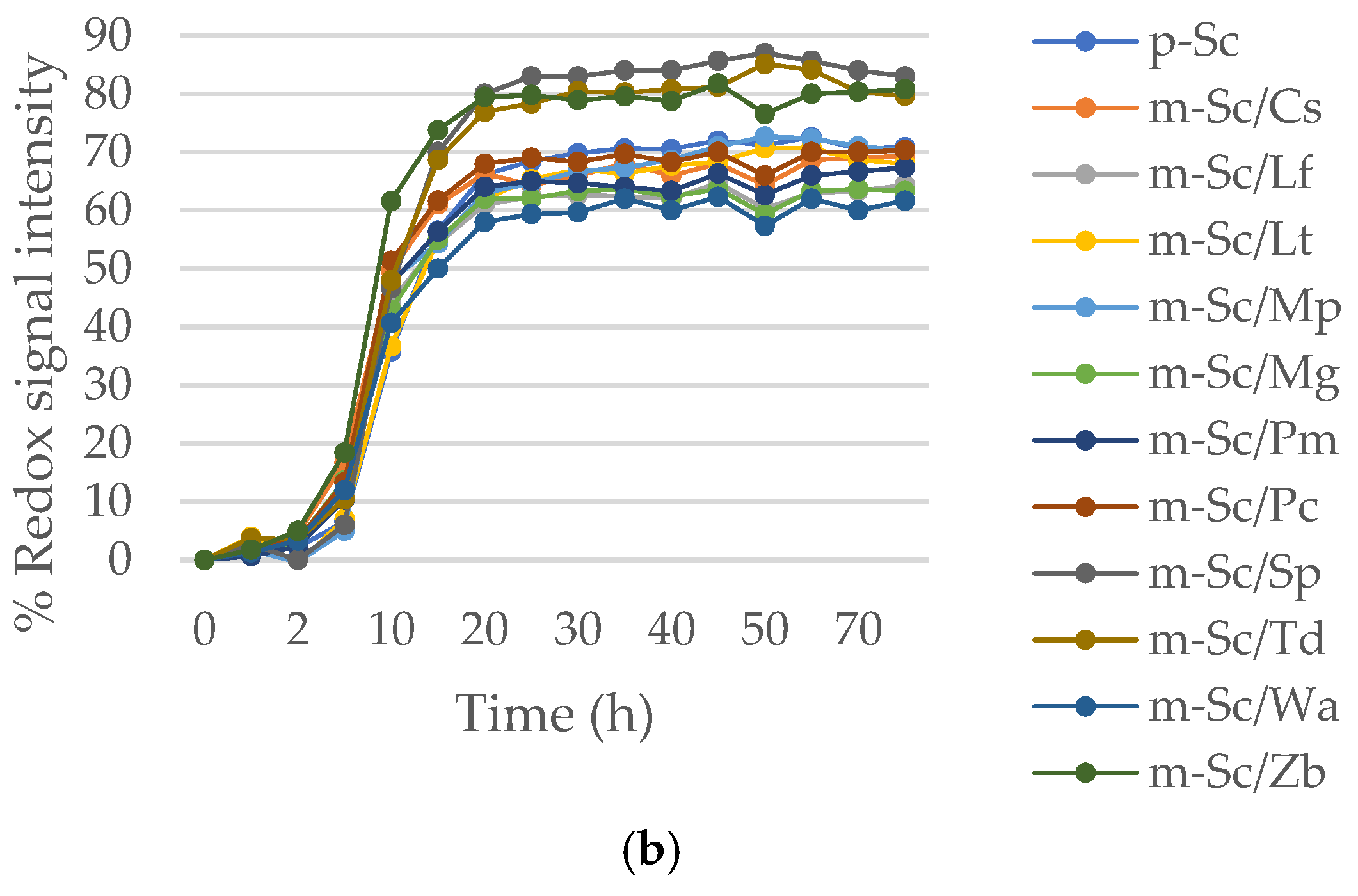
In pure cultures exposed to high osmotic pressure (Figure 4a), control culture of S. cerevisiae has undergone a large increase in its metabolic rate after 5 h of fermentation, completing the entire process only overtaken by pure cultures of L. fermentati (p-Lf) and L. thermotolerans (p-Lt). These two non-Saccharomyces strains had already shown their osmophilic character before (Figure 1a,b). The combinations between S. cerevisiae and S. pombe (m-Sc/Sp), T. delbrueckii (m-Sc/Td) and Z. bailii (m-Sc/Zb) (Figure 4b) presented a metabolic output upper than control in the presence of 30% sorbitol.
Similar behavior has been observed when the stress factor is the 13% of ethanol (Figure 5). The S. cerevisiae strain showed better adaptation to high levels of ethanol in the medium, only overtaken by Z. bailii (Figure 5a). However, there is a rise in the growth response when S. cerevisiae is associated with the non-Saccharomyces, Z. bailii (m-Sc/Zb) and S. pombe (m-Sc/Sp) (Figure 5b). The metabolic output was improved in the association of S. cerevisiae with the rest of non-Saccharomyces since their growth was higher than pure culture of non-Saccharomyces alone (Figure 5a). The non-conventional species that showed good resistance to ethanol were Z. bailii (p-Zb), L. fermentati (p-Lf) and M. guilliermondii (p-Mg) in pure culture (Figure 5a).
3.3. Small Scale Fermentations with Candida stellata CLI 920 and Schizosaccharomyces Pombe CLI 1085 in Mixed Culture with Saccharomyces cerevisiae CLI 519 under Osmotic Stress
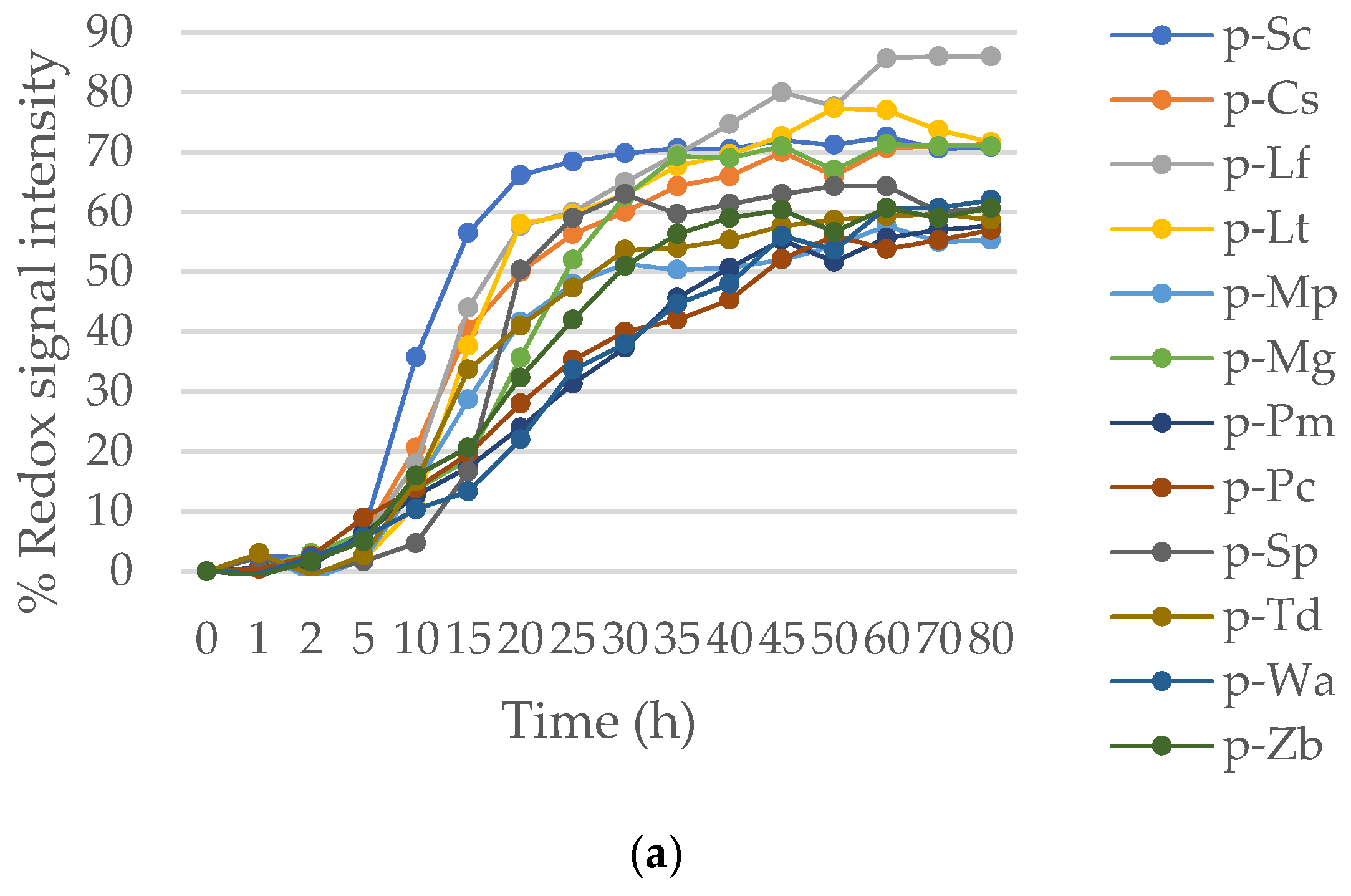
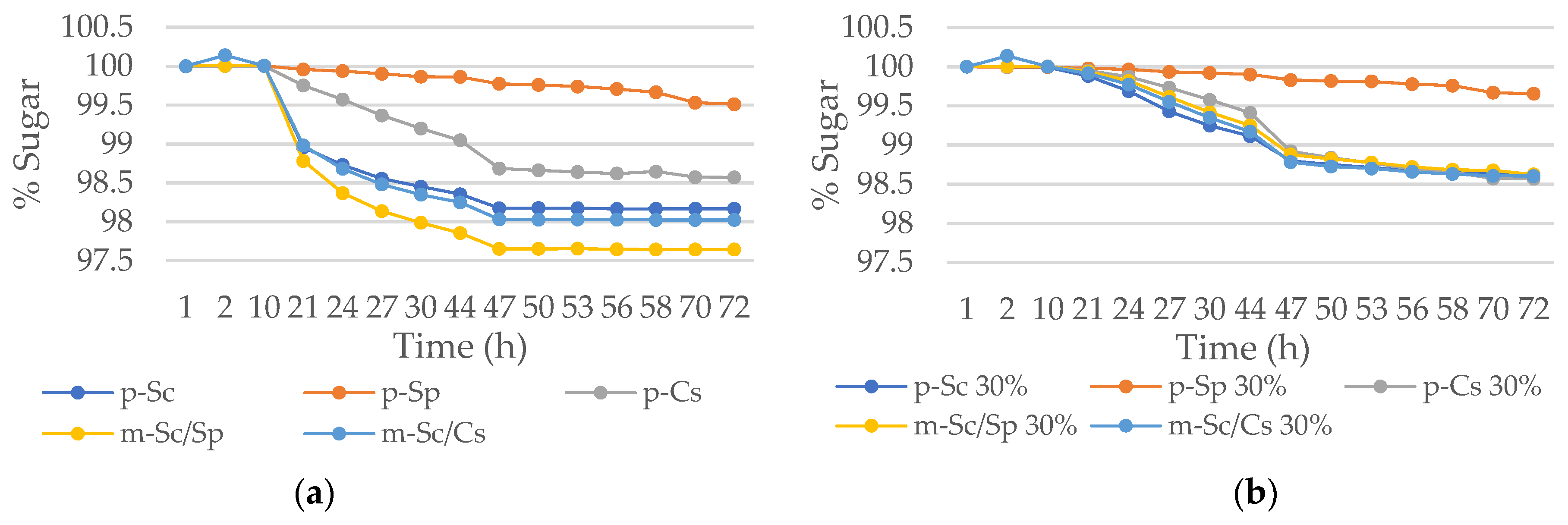
Microvinifications were carried out in a synthetic medium with the non-Saccharomyces species, C. stellata and S. pombe inoculated at 50% together with S. cerevisiae in a control medium and in a high osmotic pressure medium generated with 30% of sorbitol. The fermentative kinetic in standard medium (Figure 6a) indicates that the fermentation process finished from 47 h, except for pure culture of S. pombe (p-Sp). The control p-Sc and both mixed cultures presented a kinetic pattern very similar. By contrast, the non-Saccharomyces pure cultures (p-Cs and p-Sp) presented a slow fermentation, especially for p-Sp was more long-delayed and incomplete.
Figure 6b corresponds to cited combinations but under high osmotic pressure media. The worst adapted strain was S. pombe p-Sp, being all others very similar. S. cerevisiae p-Sc has shown a good adaptation to osmotic pressure again, but it is worth highlighting the similar fermentative behavior of p-Cs to the control p-Sc.
The chemical compounds analyzed at final of fermentation are shown in Table 2. Yeasts consume glucose content in p-Sc and mixed cultures when the culture medium has not fermentative stress element. However, pure cultures of non-Saccharomyces finished the fermentation with elevated amount of glucose under the same situation, highlighting p-Sp (45.8 g/L of glucose); both showed significant differences (p > 0.05) respect to their control (p-Sc) (Table 2). Only p-Sp was unable to consume the glucose portion under stressed condition (30% sorbitol), affecting to its ethanol production and pH values at final of fermentation. The highest ethanol content was produced by C. stellata in pure culture under osmotic stress. This data is in contrast with its behavior under standard conditions where p-Cs finished with 13.53 g/L of glucose.
Glycerol and trehalose contents are consistent with the conditions of study, a higher production is observed under high osmotic pressure. Mixed cultures secreted high amount of glycerol with similar values to the control of S. cerevisiae. The non-Saccharomyces, S. pombe was the strain that produced a lesser quantity of glycerol (Table 2).
There are some differences concerning to acetic acid contents (Table 2). Mixed cultures presented high concentration similar to the control of S. cerevisiae; these values are more elevated under osmotic stress situations. By contrast, significant differences are observed in both non-Saccharomyces pure cultures regarding to the control, showing very low values (0.10 g/L in p-Sp30% and 0.00 g/L in p-Cs30%).
4. Discussion
According to the hypothesis proposed by Ungar [33], climate change lacks daily relevance that may encourage people to obtain knowledge about the topic. For winemakers, weather phenomena have become very important since it is related to the annual course of the vineyard operations and to the quality of final products, even for their place in the market [1]. A key factor to reach the biotechnological success with wine yeasts is to know their tolerance to environmental stress conditions. There is a correlation between fermentative capacity and high tolerance to stress [34] and stress tolerance is a good criterion for selecting oenologically interesting yeasts [35]. The actual work is a first screening in synthetic media of native non-Saccharomyces yeasts resistance to stresses currently caused by global warming in wine elaboration, in order to select the best yeast strains as strategy to mitigate its effects on wine quality. Thus, twelve autochthonous non-Saccharomyces strains and one S. cerevisiae strain from grapes and musts of cellars belonging to D.O. “Vinos de Madrid” has been employed alone or combined in fermentations since these strains are adapted to conditions associated with their specific winegrowing area [24,36].
Several yeast members that constitute wine fermentation microbiota as Saccharomyces, Hanseniaspora, Pichia, Candida, Starmerella, Torulaspora, and Lachancea are facultative anaerobes microorganisms that can grow and survive under these conditions [37]. Nevertheless, most of non-Saccharomyces yeasts have more oxygen demands and low fermentative power in comparison with S. cerevisiae [38,39,40,41,42]. In the present work, the non-Saccharomyces strains grew with a similar profile independently of the presence of oxygen without any stress factor, but M. pulcherrima, W. anomalus, and Z. bailii showed larger oxygen dependence since their metabolic output was noticeably higher in aerobiosis. The yeast species M. pulcherrima has demonstrated in previous studies [39,42,43,44] that its capacity to contribute actively to the outcome of the fermentation is directly related to oxygen availability, being considered an obligate aerobe microorganism. In fact, this non-Saccharomyces species has displayed a fully respiratory glucose metabolism [41,44]. In the same way, Contreras et al. [45] (also observed that Z. bailii AWRI 1578 presented the capacity to metabolize sugars without the collateral production of ethanol with a determined aeration regime, producing wines with lower alcohol content in sequential culture with S. cerevisiae. In addition, a fully aerobic or weakly fermentative metabolism has been described in W. anomalus and it is known for film formation covering bulk wines in unfilled vessels [46,47].
One of the first stress conditions supported in the fermentation process by yeasts is produced by an elevated concentration of osmotically active substances of the grape must, mainly glucose and fructose. Yeast cells respond to such hyperosmotic shock through the modification of their cell wall and cytoskeleton and the synthesis and accumulation of compatible compounds, which help to reestablish an osmotic equilibrium [6]. In the present work, 20% and 30% of sorbitol generate the hyperosmotic scenario by yeasts. In general, the growth percentage in the non-Saccharomyces strains is similar with or without oxygen under osmotic stress created by the different contents of sorbitol. In the work of Krantz et al. [48], they excluded oxygen from the culture and noted that the stress response did not differ qualitatively from that under aerobic conditions and, particularly, encompasses the oxidative stress response genes. They observed an almost perfect coincidence between genes showing up- or downregulated expression after osmotic stress under aerobic and anaerobic conditions. In our study, the most resistant non-Saccharomyces strains to osmotic pressure have been L. fermentati CLI 1220, L. thermotolerans CLI 1219, S. pombe CLI 1085 in AE and AN. These results agree with those obtained previously [26] where the non-Saccharomyces, L. thermotolerans and S. pombe strains presented high tolerance under 30% sorbitol. Accordingly, a broad study about the genus Lachancea [49] presented three yeast species mainly associated with grape must and wine fermentation processes: L. fermentati, L. thermotolerans and L. lanzarotensis. These species demonstrated an optimal growth on the 50% D-glucose supplemented medium, suggesting to their increased osmotic tolerance. To mitigate such stress, L. thermotolerans strains showed the ability to assimilate glycerol while L. fermentati and L. lanzarotensis strains displayed variability in glycerol assimilation [49]. For its part, the application of S. pombe has been suggested in warm viticulture area characterized by grape juices with high sugar content, pH values close to 4 and low malic acid concentration [50,51]. In addition, the cell wall of S. pombe presents a big thickness and a particular presence of galacto-mannoproteins in the outer layer that give them structural strength enough to resist high osmotic pressures [17,52].
The increasing ethanol concentration during the alcoholic fermentation represents another stress factor by yeasts. The study of suppression of the cellular growth in the presence of ethanol is a simple and useful method by classification of a high number of strains by its ability to tolerate alcohol [53]. During the present work, the presence of 5% ethanol is not a limiting factor by the metabolic output of most non-Saccharomyces strains, though P. carsonii and W. anomalus presented lower growth values. The fermentative capacity of these two non-Saccharomyces strains (P. carsonii CLI 1221 and W. anomalus CLI 1218) was tested in previous works [25,54]. Both strains showed a fermentative kinetics studied as weight loss in CO2 grams released, considerably less than a S. cerevisiae native strain [54]; probably, this low fermentative power can be related with their evident low ethanol tolerance. When ethanol content rises to 8%, M. pulcherrima and P. toletana viability was considerably reduced. These results are in accordance with those obtained in a previous work, where these strains presented one of the lowest tolerances to ethanol content [26]. In general, the fermentative capacity of M. pulcherrima species is considered low, with many strains reaching around 4% v/v in ethanol [55,56], although some studies have denoted the production of ethanol up 6–10% v/v [43,57]. Under 13% ethanol stress condition, the non-Saccharomyces cell survival are enhanced in the presence of oxygen, although the resistance of Z. bailii CLI 622 also highlights in an anaerobic environment. The genus Zygosaccharomyces is included in the group of spoilage yeasts for the food and drinks industries, with Z. bailii and Z. rouxii being more frequent. This group presents key physiological characteristics as ability to vigorously ferment hexose sugars such as glucose and fructose, resistance to weak-acid preservatives, extreme osmotolerance and high resistance to both ethanol and acetic acid that contribute to their spoilage capacity [58,59,60,61,62]. Sousa et al. [58] described a physiological strategy carried out by Z. bailii where the presence of ethanol in the medium appears to reinforce acetic acid control and plays a protective role for this yeast, inhibiting the acid uptake ad maintaining its intracellular concentration below toxic levels.
Due to climatic change, pH values are increased, affecting the global wine quality [63]. Furthermore, higher pH values hold the risk of higher microbial contamination mainly in the early stages of fermentation course. To overcome this problem, the correction of the initial pH in musts is a common practise but it may involve an additional stress factor for the survival and growth of some yeast species [64]. In our assay, we tested the yeast performance at pH 3.0 and, in general, most strains were metabolically active at this acidic pH value. It is worth noting that L. fermentati, L. thermotolerans and S. pombe were more tolerant yeast strains again. It is well known that the Lachancea and Schizosaccharomyces species have the capacity to modulate different organic acids during fermentation, varying the pH of the wine. Thus, a microbiological acidification with L. thermotolerans is a classical use of this yeast species in warm climate viticultural areas [65]. This character is due to the metabolic activity of L. thermotolerans, which is a good L-lactic producer [66]. A L. thermotolerans native strain from D.O. “Vinos de Madrid” (Spain) led to a fall to pH 2.43 because of 8.32 g/L of lactic acid production in Malvar white wines elaboration [56]. Kapsopoulou et al. [67] reported similar pH value (2.9) due to the lactic acid production (9.6 g/L) using L. thermotolerans strain in pure culture. This biological acidification can improve the overall wine quality in warm areas [68]. In the same way, L. fermentati strains has also been described as lactic acid producer [49] and proposed to sour beer or low alcohol beer elaborations [69,70,71]. In addition, the production of lactic acid from glucose and cellobiose has been demonstrated in S. pombe [72]. Alternatively, some authors have proposed the combined use of S. pombe and L. thermotolerans to solve problems in warm regions [73,74], since S. pombe consumes totally malic acid achieving the microbiological stabilization objective and L. thermotolerans produces lactic acid increasing the acidity of wines from low sourness musts.
In the last years, non-Saccharomyces species are considered capable of helping to improve specific characteristics of wine quality [75,76,77], depending on certain yeasts species and strains used. Generally, one disadvantage of non-Saccharomyces species is their low fermentative activity and their low resistance to additives as sulfur dioxide [8]. Thus, the use of some non-Saccharomyces together with a high fermentative S. cerevisiae strain is proposed as a strategy to metabolize all sugars into ethanol [78]. In the present work, non-Saccharomyces strains were fermented in mixed combination with S. cerevisiae under two fermentative stress conditions (30% sorbitol and 13% ethanol). Over each inoculation, yeast strains are exposed to a number of stress conditions that require a period of adaptation. The time needed for the adaptation is known as the lag-phase, characterized by the absence of cellular growth. This lag-phase, typical for all newly inoculated cultures, is so another signal of a stress-induced adaptation phase [6,79]. In our work, it is possible to see that the pure cultures under osmotic or ethanol stresses presented a lag-phase longer (around 5 h) than the mixed cultures (around 2 h). The presence of S. cerevisiae in the medium seems to shorten this adaptation period.
The growth percentage of two non-Saccharomyces strains (L. fermentati CLI 1220 and L. thermotolerans CLI 1219) exceeds the S. cerevisiae values in pure culture at 30% sorbitol. A study carried out by De Kock [23] stated that a strain of L. thermotolerans produced not only glycerol but also more polyols than S. cerevisiae, which could provide more protection against osmotic stress situation. Instead, the metabolic output of Lachancea species decreased in mixed culture with S. cerevisiae. Several studies attribute this decline in L. thermotolerans to the impact of parameters as temperature [80], lack of oxygenation [42], cell-to-cell contact [81] and the secretion of toxic compounds by S. cerevisiae [82,83]. However, L. fermentati in co-culture fermentations has been poorly studied yet [49]. Three combinations presented higher growth percentage than the control when osmotic pressure stress is applied to mixed cultures: mixed cultures of T. delbrueckii, S. pombe, and Z. bailii with S. cerevisiae. The yeast species, T. delbrueckii is known as a highly resistant microorganism to stress conditions and is often isolated from fruit juices, bakery [84] and botrytized musts [85,86], but there are not so many studies about its resistance mechanisms in high-sugar ambient. Our results are in accordance with Bely et al. [14] where they observed the highest biomass presence in mixed T. delbrueckii/S. cerevisiae culture on high-sugar fermentation. As stated before, the high resistance to ethanol in Z. bailii and S. pombe has also been documented [50,61]. This resistance to ethanol could be explained the highest metabolic activity of native Z. bailii CLI 622 and S. pombe CLI 1085 strains in mixed cultures respect to the control since the presence of non-Saccharomyces and Saccharomyces at the same time during the fermentation promotes an increase in the persistence of non-Saccharomyces yeasts [87,88].
Finally, two autochthonous yeast strains previously well-studied [18,25,26,56,89] were selected to fermentations at higher volume under high osmotic pressure. The strain S. pombe CLI 1085 has shown good resistance to three stress factors tested in the current work; besides, it was a low producer of acetic acid, consumer of malic acid and producer of lactic acid [56] that are favorable characteristics in warm winemaking areas. The pure culture C. stellata CLI 920 have presented an osmotic pressure and ethanol tolerance similar to S. cerevisiae; other study showed it as high glycerol producer, its pure culture was the most appreciated by trained tasters from IMIDRA Institute and this strain was proposed by using in sweet wine elaboration [56].
It is interesting to note that C. stellata CLI 920 strain produced the highest level of ethanol under osmotic stress situations. This non-Saccharomyces species is typically associated with the fermentation of botrytized wines and wines produced from overripe grapes in cooked musts [18]. Besides, various authors have mentioned that C. stellata has better ability than S. cerevisiae to grow in high-sugar fermentations [90,91]. A strain-dependent characteristic described in S. cerevisiae [92,93] denoted that this yeast may possess the capacity to overcome osmotic stress and to yield ethanol by fermentation of musts with high sugar proportion in winemaking. It could be possible C. stellata CLI 920 showed this ability, although more studies are needed.
After hyperosmotic shock, yeast cells produce and accumulate osmoprotectants as glycerol and trehalose [5]. This statement is in agreement with our results where these two compounds are elevated in fermentations under high osmotic pressure. However, the production of glycerol often linked to increased acetic acid yield in S. cerevisiae [94,95]. Nevertheless, it seems that many non-Saccharomyces yeasts have different metabolic responses to osmotic stress, producing low levels of acetic acid [14,96,97]. Several authors have stated an elevated acetic acid production as the main consequence of the use of S. pombe in alcoholic beverages elaboration [98,99,100,101,102]. The S. pombe CLI 1085 used in this study produced acetic acid values very low (0.02 g/L under control situations; 0.10 g/L under osmotic stress). This same strain produced 0.33 g/L of acetic acid in pure culture in Malvar white wines fermentation [56]. These values are similar to those reported by Du Plessis et al. [103] ranged between 0.07 g/L and 0.35 g/L acetic acid. Ciani et al. [104] described the C. stellata DBVPG 3827 strain as low acetic acid producer in both aerobic (0.12 g/L of acetic acid) and anaerobic (0.10 g/L of acetic acid) conditions regarding S. cerevisiae monoculture. This result is in agreement with our data where the acetic acid of p-Cs and p-Cs30% cultures were below detection limits. The mixed cultures completed the fermentation and had high glycerol and acetic acid levels very similar to controls (p-Sc and p-Sc30%); these fermentations seem to be dominated by S. cerevisiae strain due to the high similarity of their parameters at final of fermentation.
5. Conclusions
Most non-Saccharomyces yeasts tested have been capable of growing under different stress conditions, but better in the presence of oxygen. The most resistant species were L. fermentati, L. thermotolerans, and S. pombe under osmotic pressure, ethanol, and pH stresses. The use of mixed combinations has improved the resistance against stress situations by S. pombe, T. delbrueckii, and Z. bailii under osmotic pressure and by C. stellata, S. pombe, and Z. bailii under high ethanol content regarding S. cerevisiae pure culture. However, the mixed cultures between C. stellata and S. pombe with S. cerevisiae have shown a favorable increase of glycerol content but the adverse rise of acetic acid. Maybe one solution to the problem of excessive volatile acidity was to use a higher concentration of non-Saccharomyces strains in mixed combinations.
Therefore, the resistance presented by these non-Saccharomyces to stresses inherent to fermentation make them good candidates for winemaking in warm areas. However, more studies should be carried out to wine elaboration at a higher scale.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/fermentation7030124/s1, Figure S1: Phenotypic microarray analysis (% of redox signal intensity) of non-Saccharomyces strains in aerobic (AE) and anaerobic (AN) condition after 96 h in YNB medium.
Author Contributions
M.G.: analyzed results, discussion of results and wrote the manuscript; J.C. and J.M.C.: analyzed results and participated in the revision of the manuscript; T.A.: designed experiments, performed experiments, analyzed results, discussion of results and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the research project RTI2018-101085-R-C31-MINOVID (MICINN/AEI/ERDF, EU).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Battaglini, A.; Barbeau, G.; Bindi, M.; Badeck, F.W. European winegrowers’ perceptions of climate change impact and options for adaptation. Reg. Environ. Chang. 2009, 9, 61–73. [Google Scholar] [CrossRef]
- Crespo, J.; Rigou, P.; Romero, V.; García, M.; Arroyo, T.; Cabellos, J.M. Effect of seasonal climate fluctuations on the evolution of glycoconjugates along the ripening period of grapevine cv. Muscat a petits grains blancs berries. J. Sci. Food Agric. 2018, 98, 1803–1812. [Google Scholar] [CrossRef] [PubMed]
- Drappier, J.; Thibon, C.; Rabot, A.; Geny-Denis, L. Relationship between wine composition and temperature: Impact on Bordeaux wine typicity in the context of global warming—Review. Crit. Rev. Food Sci. Nutr. 2019, 59, 14–30. [Google Scholar] [CrossRef] [PubMed]
- Berbegal, C.; Fragasso, M.; Russo, P.; Bimbo, F.; Grieco, F.; Spano, G.; Capozzi, V. Climate changes and food quality: The potential of microbial activities as mitigating strategies in the wine sector. Fermentation 2019, 5, 85. [Google Scholar] [CrossRef] [Green Version]
- Hohmann, S.; Mager, W.H. (Eds.) Yeast Stress Responses, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2003; p. 389. ISBN 3540456112. [Google Scholar]
- Bauer, E.F.; Pretorius, L.S. Yeast stress response and fermentation efficiency: How to survive the making of wine—A review. S. Afr. J. Enol. Vitic. 2000, 21, 27–51. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [Green Version]
- Benito, Á.; Calderón, F.; Benito, S. The influence of non-Saccharomyces species on wine fermentation quality parameters. Fermentation 2019, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Escribano, R.; González-Arenzana, L.; Garijo, P.; Berlanas, C.; López-Alfaro, I.; López, R.; Gutiérrez, A.R.; Santamaría, P. Screening of enzymatic activities within different enological non-Saccharomyces yeasts. J. Food Sci. Technol. 2017, 54, 1555–1564. [Google Scholar] [CrossRef] [Green Version]
- Berbegal, C.; Spano, G.; Tristezza, M.; Grieco, F.; Capozzi, V. Microbial resources and innovation in the wine production sector. S. Afr. J. Enol. Vitic. 2017, 38, 156–166. [Google Scholar] [CrossRef]
- Kuchen, B.; Maturano, Y.P.; Mestre, M.V.; Combina, M.; Toro, M.E.; Vazquez, F. Selection of native non-Saccharomyces yeasts with biocontrol activity against spoilage yeasts in order to produce healthy regional wines. Fermentation 2019, 5, 60. [Google Scholar] [CrossRef] [Green Version]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces commercial starter cultures: Scientific trends, recent patents and innovation in the wine sector. Recent Pat. Food Nutr. Agric. 2019, 10, 27–39. [Google Scholar] [CrossRef]
- Ramírez, M.; Velázquez, R. The yeast Torulaspora delbrueckii: An interesting but difficult-to-use tool for winemaking. Fermentation 2018, 4, 94. [Google Scholar] [CrossRef] [Green Version]
- Bely, M.; Stoeckle, P.; Masneuf-Pomarède, I.; Dubourdieu, D. Impact of mixed Torulaspora delbrueckii-Saccharomyces cerevisiae culture on high-sugar fermentation. Int. J. Food Microbiol. 2008, 122, 312–320. [Google Scholar] [CrossRef]
- Benito, S. The impact of Torulaspora delbrueckii yeast in winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 3081–3094. [Google Scholar] [CrossRef]
- Morata, A.; Benito, S.; Loira, I.; Palomero, F.; González, M.C. Formation of pyranoanthocyanins by Schizosaccharomyces pombe during the fermentation of red must. Int. J. Food Microbiol. 2012, 159, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Loira, I.; Morata, A.; Palomero, F.; González, C.; Suárez-Lepe, J.A. Schizosaccharomyces pombe: A promising biotechnology for modulating wine composition. Fermentation 2018, 4, 70. [Google Scholar] [CrossRef] [Green Version]
- García, M.; Esteve-Zarzoso, B.; Cabellos, J.M.; Arroyo, T. Advances in the study of Candida stellate. Fermentation 2018, 4, 74. [Google Scholar] [CrossRef] [Green Version]
- Martin, V.; Jose Valera, M.; Medina, K.; Boido, E.; Carrau, F. Oenological impact of the Hanseniaspora/Kloeckera yeast genus on wines—A review. Fermentation 2018, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Aranda, A. Enological repercussions of non-Saccharomyces species. Fermentation 2019, 5, 68. [Google Scholar] [CrossRef] [Green Version]
- Morata, A.; Loira, I.; Escott, C.; del Fresno, J.M.; Bañuelos, M.A.; Suárez-Lepe, J.A. Applications of Metschnikowia pulcherrima in wine biotechnology. Fermentation 2019, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Ciani, M.; Comitini, F. Yeast interactions in multi-starter wine fermentation. Curr. Opin. Food Sci. 2015, 1, 1–6. [Google Scholar] [CrossRef]
- De Kock, M.C. Investigating Osmotic Stress in Mixed Yeast Cultures and Its Effects on Wine Composition. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2015. [Google Scholar]
- Tello, J.; Cordero-Bueso, G.; Aporta, I.; Cabellos, J.M.; Arroyo, T. Genetic diversity in commercial wineries: Effects of the farming system and vinification management on wine yeasts. J. Appl. Microbiol. 2012, 112, 302–315. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Bueso, G.; Esteve-Zarzoso, B.; Cabellos, J.M.; Gil-Díaz, M.; Arroyo, T. Biotechnological potential of non-Saccharomyces yeasts isolated during spontaneous fermentations of Malvar (Vitis vinifera cv. L.). Eur. Food Res. Technol. 2013, 236, 193–207. [Google Scholar] [CrossRef]
- García, M.; Greetham, D.; Wimalasena, T.T.; Phister, T.G.; Cabellos, J.M.; Arroyo, T. The phenotypic characterization of yeast strains to stresses inherent to wine fermentation in warm climates. J. Appl. Microbiol. 2016, 121, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Bochner, B.R.; Gadzinski, P.; Panomitros, E. Phenotype microarrays for high-throughput phenotypic testing and assay of gene function. Genome Res. 2001, 11, 1246–1255. [Google Scholar] [CrossRef] [Green Version]
- Borglin, S.; Joyner, D.; DeAngelis, K.M.; Khudyakov, J.; D’haeseleer, P.; Joachimiak, M.P.; Hazen, T. Application of phenotypic microarrays to environmental microbiology. Curr. Opin. Biotechnol. 2012, 23, 41–48. [Google Scholar] [CrossRef] [Green Version]
- DeNittis, M.; Querol, A.; Zanoni, B.; Minati, J.L.; Ambrosoli, R. Possible use of Biolog methodology for monitoring yeast presence in alcoholic fermentation for wine-making. J. Appl. Microbiol. 2010, 108, 1199–1206. [Google Scholar] [CrossRef]
- Greetham, D. Phenotype microarray technology and its application in industrial biotechnology. Biotechnol. Lett. 2014, 36, 1153–1160. [Google Scholar] [CrossRef]
- Berridge, M.V.; Herst, P.M.; Tan, A.S. Tetrazolium dyes as tools in cell biology: New insights into their cellular reduction. Biotechnol. Annu. Rev. 2005, 11, 127–152. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Bueso, G.; Arroyo, T.; Serrano, A.; Tello, J.; Aporta, I.; Vélez, M.D.; Valero, E. Influence of the farming system and vine variety on yeast communities associated with grape berries. Int. J. Food Microbiol. 2011, 145, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Ungar, S. Knowledge, ignorance and the popular culture: Climate change versus the ozone hole. Public Underst. Sci. 2000, 9, 297–312. [Google Scholar] [CrossRef]
- Ivorra, C.; Pérez-Ortín, J.E.; Del Olmo, M. An inverse correlation between stress resistance and stuck fermentations in wine yeasts. A molecular study. Biotechnol. Bioeng. 1999, 64, 698–708. [Google Scholar] [CrossRef]
- Zuzuarregui, A.; del Olmo, M. Analyses of stress resistance under laboratory conditions constitute a suitable criterion for wine yeast selection. Antonie Van Leeuwenhoek 2004, 85, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Esteve-Zarzoso, B.; Gostıncar, A.; Bobet, R. Selection and molecular characterization of wine yeasts isolated from the “El Penedès” area (Spain). Food Microbiol. 2000, 17, 553–562. [Google Scholar] [CrossRef]
- Shekhawat, K.; Patterton, H.; Bauer, F.F.; Setati, M.E. RNA-seq based transcriptional analysis of Saccharomyces cerevisiae and Lachancea thermotolerans in mixed-culture fermentations under anaerobic conditions. BMC Genom. 2019, 20, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visser, W.; Scheffers, W.A.; Batenburg-Van der Vegte, W.H.; Van Dijken, J.P. Oxygen requirements of yeasts. Appl. Environ. Microbiol. 1990, 56, 3785–3792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, E.H.; Nissen, P.; Sommer, P.; Nielsen, J.C.; Arneborg, N. The effect of oxygen on the survival of non-Saccharomyces yeasts during mixed culture fermentations of grape juice with Saccharomyces cerevisiae. J. Appl. Microbiol. 2001, 91, 541–547. [Google Scholar] [CrossRef]
- Brandam, C.; Lai, Q.P.; Julien-Ortiz, A.; Taillandier, P. Influence of oxygen on alcoholic fermentation by a wine strain of Torulaspora delbrueckii: Kinetics and carbon mass balance. Biosci. Biotechnol. Biochem. 2013, 77, 1848–1853. [Google Scholar] [CrossRef] [Green Version]
- Quirós, M.; Rojas, V.; Gonzalez, R.; Morales, P. Selection of non-Saccharomyces yeast strains for reducing alcohol levels in wine by sugar respiration. Int. J. Food Microbiol. 2014, 181, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Shekhawat, K.; Bauer, F.F.; Setati, M.E. Impact of oxygenation on the performance of three non-Saccharomyces yeasts in co-fermentation with Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2016, 101, 2479–2491. [Google Scholar] [CrossRef]
- Sun, S.Y.; Gong, H.S.; Jiang, X.M.; Zhao, Y.P. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae on alcoholic fermentation behaviour and wine aroma of cherry wines. Food Microbiol. 2014, 44, 15–23. [Google Scholar] [CrossRef]
- Morales, P.; Rojas, V.; Quirós, M.; Gonzalez, R. The impact of oxygen on the final alcohol content of wine fermented by a mixed starter culture. Appl. Microbiol. Biotechnol. 2015, 99, 3993–4003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.A.; Curtin, C.; Varela, C. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, M.; Pretorius, I.S. Microbial spoilage and preservation of wine: Using weapons from nature’s own arsenal—A review. S. Afr. J. Enol. Vitic. 2019, 21, 74–96. [Google Scholar] [CrossRef] [Green Version]
- Walker, G.M. Pichia anomala: Cell physiology and biotechnology relative to other yeasts. Antonie Van Leeuwenhoek 2011, 99, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krantz, M.; Nordlander, B.; Valadi, H.; Johansson, M.; Gustafsson, L.; Hohmann, S. Anaerobicity prepares Saccharomyces cerevisiae cells for faster adaptation to osmotic shock. Eukaryot. Cell 2004, 3, 1381–1390. [Google Scholar] [CrossRef] [Green Version]
- Porter, T.J.; Divol, B.; Setati, M.E. Lachancea yeast species: Origin, biochemical characteristics and oenological significance. Food Res. Int. 2019, 119, 378–389. [Google Scholar] [CrossRef]
- Suárez-Lepe, J.A.; Palomero, F.; Benito, S.; Calderón, F.; Morata, A. Oenological versatility of Schizosaccharomyces spp. Eur. Food Res. Technol. 2012, 235, 375–383. [Google Scholar] [CrossRef]
- Benito, S.; Palomero, F.; Morata, A.; Calderón, F.; Palmero, D.; Suárez-Lepe, J.A. Physiological features of Schizosaccharomyces pombe of interest in making of white wines. Eur. Food Res. Technol. 2013, 236, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Palomero, F.; Morata, A.; Benito, S.; Calderón, F.; Suárez-Lepe, J.A. New genera of yeasts for over-lees aging of red wine. Food Chem. 2009, 112, 432–441. [Google Scholar] [CrossRef]
- Pina, C.; Santos, C.; Couto, J.A.; Hogg, T. Ethanol tolerance of five non-Saccharomyces wine yeasts in comparison with a strain of Saccharomyces cerevisiae-Influence of different culture conditions. Food Microbiol. 2004, 21, 439–447. [Google Scholar] [CrossRef]
- García, M.; Esteve-Zarzoso, B.; Cabellos, J.M.; Arroyo, T. Sequential non-Saccharomyces and Saccharomyces cerevisiae fermentations to reduce the alcohol content in wine. Fermentation 2020, 6, 60. [Google Scholar] [CrossRef]
- Cominiti, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- García, M.; Arroyo, T.; Crespo, J.; Cabellos, J.M.; Esteve-Zarzoso, B. Use of native non-Saccharomyces strain: A new strategy in D.O. “Vinos de Madrid” (Spain) wines elaboration. Eur. J. Food Sci. Technol. 2017, 5, 1–31. [Google Scholar]
- Combina, M.; Elía, A.; Mercado, L.; Catania, C.; Ganga, A.; Martinez, C. Dynamics of indigenous yeast populations during spontaneous fermentation of wines from Mendoza, Argentina. Int. J. Food Microbiol. 2005, 99, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Sousa, M.J.; Miranda, L.; Côrte-Real, M.; Leao, C. Transport of acetic acid in Zygosaccharomyces bailii: Effects of ethanol and their implications on the resistance of the yeast to acidic environments. Appl. Environ. Microbiol. 1996, 62, 3152–3157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratford, M.; James, S.A. Non-alcoholic beverages and yeasts. In Yeasts in Food: Beneficial and Detrimental Aspects; Boekhout, T., Robert, V., Eds.; Behr’s Verlag: Hamburg, Germany, 2003; pp. 309–345. ISBN 9781855737068. [Google Scholar]
- Martorell, P.; Stratford, M.; Steels, H.; Fernández-Espinar, M.T.; Querol, A. Physiological characterization of spoilage strains of Zygosaccharomyces bailii and Zygosaccharomyces rouxii isolated from high sugar environments. Int. J. Food Microbiol. 2007, 114, 234–242. [Google Scholar] [CrossRef]
- Santos, J.; Sousa, M.J.; Cardoso, H.; Inácio, J.; Silva, S.; Spencer-Martins, I.; Leão, C. Ethanol tolerance of sugar transport, and the rectification of stuck wine fermentations. Microbiology 2008, 154, 422–430. [Google Scholar] [CrossRef] [Green Version]
- Stratford, M.; Steels, H.; Nebe-von-Caron, G.; Novodvorska, M.; Hayer, K.; Archer, D.B. Extreme resistance to weak-acid preservatives in the spoilage yeast Zygosaccharomyces bailii. Int. J. Food Microbiol. 2013, 166, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.V.; White, M.A.; Cooper, O.R.; Storchmann, K. Climate change and global wine quality. Clim. Chang. 2005, 73, 319–343. [Google Scholar] [CrossRef]
- Fleet, G.H.; Heard, G.M. Yeast growth during fermentation. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Harwood Academic Publishers: Chur, Switzerland, 1993; pp. 27–54. ISBN 3718651327. [Google Scholar]
- Benito, S. The impacts of Lachancea thermotolerans yeast strains on winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 6775–6790. [Google Scholar] [CrossRef] [Green Version]
- Kapsopoulou, K.; Mourtzini, A.; Anthoulas, M.; Nerantzis, E. Biological acidification during grape must fermentation using mixed cultures of Kluyveromyces thermotolerans and Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2007, 23, 735–739. [Google Scholar] [CrossRef]
- Kapsopoulou, K.; Kapaklis, A.; Spyropoulos, H. Growth and fermentation characteristics of a strain of the wine yeast Kluyveromyces thermotolerans isolated in Greece. World J. Microbiol. Biotechnol. 2005, 21, 1599–1602. [Google Scholar] [CrossRef]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Osburn, K.; Amaral, J.; Metcalf, S.R.; Nickens, D.M.; Rogers, C.M.; Sausen, C.; Caputo, R.; Miller, J.; Li, H.; Tennessen, J.M.; et al. Primary souring: A novel bacteria-free method for sour beer production. Food Microbiol. 2018, 70, 76–84. [Google Scholar] [CrossRef]
- Bellut, K.; Michel, M.; Hutzler, M.; Zarnkow, M.; Jacob, F.; De Schutter, D.P.; Daenen, L.; Lynch, K.M.; Zannini, E.; Arendt, E.K. Investigation into the potential of Lachancea fermentati strain KBI 12.1 for low alcohol beer brewing. J. Am. Soc. Brew. Chem. 2019, 77, 157–169. [Google Scholar] [CrossRef]
- Bellut, K.; Krogerus, K.; Arendt, E.K. Lachancea fermentati strains isolated from kombucha: Fundamental insights, and practical application in low alcohol beer brewing. Front. Microbiol. 2020, 11, 764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozaki, A.; Konishi, R.; Otomo, C.; Kishida, M.; Takayama, S.; Matsumoto, T.; Tanaka, T.; Kondo, A. Metabolic engineering of Schizosaccharomyces pombe via CRISPR-Cas9 genome editing for lactic acid production from glucose and cellobiose. Metab. Eng. Commun. 2017, 5, 60–67. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Palomero, F.; Benito, S.; Ebeler, S.; Hopfer, H. Combine use of selected Schizosaccharomyces pombe and Lachancea thermotolerans yeast strains as an alternative to the traditional malolactic fermentation in red wine production. Molecules 2015, 20, 9510–9523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benito, Á.; Calderón, F.; Benito, S. Mixed alcoholic fermentation of Schizosaccharomyces pombe and Lachancea thermotolerans and its influence on mannose-containing polysaccharides wine composition. AMB Express 2019, 9, 17. [Google Scholar] [CrossRef] [Green Version]
- Esteve-Zarzoso, B.; Manzanares, P.; Ramón, D.; Querol, A. The role of non-Saccharomyces yeasts in industrial winemaking. Int. Microbiol. 1998, 1, 143–148. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, C. The impact of non-Saccharomyces yeasts in the production of alcoholic beverages. Appl. Microbiol. Biotechnol. 2016, 100, 9861–9874. [Google Scholar] [CrossRef] [PubMed]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Torrado, R.; Carrasco, P.; Aranda, A.; Gimeno-Alcañiz, J.; Pérez-Ortín, J.E.; Matallana, E.; Del Olmo, M. Study of the first hours of microvinification by the use of osmotic stress-response genes as probes. Syst. Appl. Microbiol. 2002, 161, 153–161. [Google Scholar] [CrossRef]
- Balikci, E.K.; Tanguler, H.; Jolly, N.P.; Erten, H. Influence of Lachancea thermotolerans on cv. Emir wine fermentation. Yeast 2016, 33, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Nissen, P.; Arneborg, N. Characterization of early deaths of non-Saccharomyces yeasts in mixed cultures with Saccharomyces cerevisiae. Arch. Microbiol. 2003, 180, 257–263. [Google Scholar] [CrossRef]
- Albergaria, H.; Francisco, D.; Gori, K.; Arneborg, N.; Gírio, F. Saccharomyces cerevisiae CCMI 885 secretes peptides that inhibit the growth of some non-Saccharomyces wine-related strains. Appl. Microbiol. Biotechnol. 2010, 86, 965–972. [Google Scholar] [CrossRef]
- Wang, C.; Mas, A.; Esteve-Zarzoso, B. The interaction between Saccharomyces cerevisiae and non-Saccharomyces yeast during alcoholic fermentation is species and strain specific. Front. Microbiol. 2016, 7, 502. [Google Scholar] [CrossRef] [Green Version]
- Legan, J.D.; Voysey, P.A. Yeast spoilage of bakery products and ingredients. J. Appl. Bacteriol. 1991, 70, 361–371. [Google Scholar] [CrossRef]
- Sipiczki, M.; Romano, P.; Lipani, G.; Miklos, I.; Antunovics, Z. Analysis of yeasts derived from natural fermentation in a Tokaj winery. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2001, 79, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Antunovics, Z.; Csoma, H.; Sipiczki, M. Molecular and genetic analysis of the yeast flora of botrytized Tokaj wines. Bull. IOIV 2003, 76, 380–397. [Google Scholar]
- Ciani, M.; Beco, L.; Comitini, F. Fermentation behaviour and metabolic interactions of multistarter wine yeast fermentations. Int. J. Food Microbiol. 2006, 108, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, L.M.; De Nadra, M.C.M.; Farías, M.E. Kinetics and metabolic behavior of a composite culture of Kloeckera apiculata and Saccharomyces cerevisiae wine related strains. Biotechnol. Lett. 2007, 29, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- García, M.; Esteve-Zarzoso, B.; Crespo, J.; Cabellos, J.M.; Arroyo, T. Yeast monitoring of wine mixed or sequential fermentations made by native strains from D.O. “Vinos de Madrid” using real-time quantitative PCR. Front. Microbiol. 2017, 8, 2520. [Google Scholar] [CrossRef]
- Lafon-Lafourcade, S. Wine and brandy. In Biotechnology Vol. V: Food and Feed Production with Microorganisms; Rehm, H.J., Reed, G., Eds.; Verlag Chemie: Weinheim, Germany, 1983; pp. 81–163. [Google Scholar]
- Charoenchai, C.; Fleet, G.H.; Henschke, P.A. Effects of temperature, pH, and sugar concentration on the growth rates and cell biomass of wine yeasts. Am. J. Enol. Vitic. 1998, 49, 283–288. [Google Scholar]
- Zhao, Y.; Lin, Y.H. Growth of Saccharomyces cerevisiae in a chemostat under high glucose conditions. Biotechnol. Lett. 2003, 25, 1151–1154. [Google Scholar] [CrossRef]
- Malacrinò, P.; Tosi, E.; Caramia, G.; Prisco, R.; Zapparoli, G. The vinification of partially dried grapes: A comparative fermentation study of Saccharomyces cerevisiae strains under high sugar stress. Lett. Appl. Microbiol. 2005, 40, 466–472. [Google Scholar] [CrossRef]
- Prior, B.A.; Toh, T.H.; Jolly, N.; Baccari, C.; Mortimer, R.K. Impact of yeast breeding for elevated glycerol production on fermentative activity and metabolite formation in Chardonnay wine. S. Afr. J. Enol. Vitic. 2000, 21, 92–99. [Google Scholar] [CrossRef] [Green Version]
- Pigeau, G.M.; Inglis, D.L. Response of wine yeast (Saccharomyces cerevisiae) aldehyde dehydrogenases to acetaldehyde stress during Icewine fermentation. J. Appl. Microbiol. 2007, 103, 1576–1586. [Google Scholar] [CrossRef]
- Milanovic, V.; Ciani, M.; Oro, L.; Comitini, F. Starmerella bombicola influences the metabolism of Saccharomyces cerevisiae at pyruvate decarboxylase and alcohol dehydrogenase level during mixed wine fermentation. Microb. Cell Fact. 2012, 11, 18. [Google Scholar] [CrossRef] [Green Version]
- Sadoudi, M.; Rousseaux, S.; David, V.; Alexandre, H.; Tourdot-Maréchal, R. Metschnikowia pulcherrima influences the expression of genes involved in PDH bypass and glyceropyruvic fermentation in Saccharomyces cerevisiae. Front. Microbiol. 2017, 8, 1137. [Google Scholar] [CrossRef]
- Mylona, A.E.; Del Fresno, J.M.; Palomero, F.; Loira, I.; Bañuelos, M.A.; Morata, A.; Calderón, F.; Benito, S.; Suárez-Lepe, J.A. Use of Schizosaccharomyces strains for wine fermentation-Effect on the wine composition and food safety. Int. J. Food Microbiol. 2016, 232, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Benito, Á.; Jeffares, D.; Palomero, F.; Calderón, F.; Bai, F.Y.; Bähler, J.; Benito, S. Selected Schizosaccharomyces pombe strains have characteristics that are beneficial for winemaking. PLoS ONE 2016, 11, e0151102. [Google Scholar] [CrossRef] [PubMed]
- Minnaar, P.P.; Jolly, N.P.; Paulsen, V.; Du Plessis, H.W.; Van Der Rijst, M. Schizosaccharomyces pombe and Saccharomyces cerevisiae yeasts in sequential fermentations: Effect on phenolic acids of fermented Kei-apple (Dovyalis caffra L.) juice. Int. J. Food Microbiol. 2017, 257, 232–237. [Google Scholar] [CrossRef]
- Roca-Domènech, G.; Cordero-Otero, R.; Rozès, N.; Cléroux, M.; Pernet, A.; Mira de Orduña, R. Metabolism of Schizosaccharomyces pombe under reduced osmotic stress conditions afforded by fed-batch alcoholic fermentation of white grape must. Food Res. Int. 2018, 113, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Scomparin, A.; Bureik, M. A convenient new method for reproducible fed-batch fermentation of fission yeast Schizosaccharomyces pombe. Biotechnol. Lett. 2020, 42, 937–943. [Google Scholar] [CrossRef]
- Du Plessis, H.W.; du Toit, M.; Hoff, J.W.; Hart, R.S.; Ndimba, B.K.; Jolly, N.P. Characterisation of non-Saccharomyces yeasts using different methodologies and evaluation of their compatibility with malolactic fermentation. S. Afr. J. Enol. Vitic. 2017, 38, 46–63. [Google Scholar] [CrossRef]
- Ciani, M.; Ferraro, L.; Fatichenti, F. Influence of glycerol production on the aerobic and anaerobic growth of the wine yeast Candida stellata. Enzym. Microb. Technol. 2000, 27, 698–703. [Google Scholar] [CrossRef]
Figure 1.
Metabolic output expressed as redox signal intensity of non-Saccharomyces yeast strains under high osmotic pressure. (a) Growth ratio under stress situations generated by 20% of sorbitol medium in aerobic and anaerobic conditions; (b) Growth ratio under stress situations generated by 30% of sorbitol medium in aerobic and anaerobic condition.
Figure 1.
Metabolic output expressed as redox signal intensity of non-Saccharomyces yeast strains under high osmotic pressure. (a) Growth ratio under stress situations generated by 20% of sorbitol medium in aerobic and anaerobic conditions; (b) Growth ratio under stress situations generated by 30% of sorbitol medium in aerobic and anaerobic condition.

Figure 2.
Metabolic output as redox signal intensity of non-Saccharomyces yeast strains under different concentrations of ethanol in AE and AN: (a) 5% of ethanol; (b) 8% of ethanol; and (c) 13% of ethanol.
Figure 2.
Metabolic output as redox signal intensity of non-Saccharomyces yeast strains under different concentrations of ethanol in AE and AN: (a) 5% of ethanol; (b) 8% of ethanol; and (c) 13% of ethanol.

Figure 3.
Metabolic output expressed as redox signal intensity of non-Saccharomyces yeast strains at low pH in aerobic (AE) and anaerobic (AN).
Figure 3.
Metabolic output expressed as redox signal intensity of non-Saccharomyces yeast strains at low pH in aerobic (AE) and anaerobic (AN).

Figure 4.
Metabolic rate as redox signal intensity of non-Saccharomyces strains and S. cerevisiae under high osmotic stress induced by 30% sorbitol in: (a) pure culture and (b) mixed culture 1:1 with S. cerevisiae. Pure culture of S. cerevisiae (p-Sc) was considered as control in both cases.
Figure 4.
Metabolic rate as redox signal intensity of non-Saccharomyces strains and S. cerevisiae under high osmotic stress induced by 30% sorbitol in: (a) pure culture and (b) mixed culture 1:1 with S. cerevisiae. Pure culture of S. cerevisiae (p-Sc) was considered as control in both cases.


Figure 5.
Metabolic rate as redox signal intensity of non-Saccharomyces strains and S. cerevisiae under alcohol degree stress induced by 13% ethanol in: (a) pure culture and (b) mixed culture 1:1 with S. cerevisiae. Pure culture of S. cerevisiae (p-Sc) was considered as control in both cases.
Figure 5.
Metabolic rate as redox signal intensity of non-Saccharomyces strains and S. cerevisiae under alcohol degree stress induced by 13% ethanol in: (a) pure culture and (b) mixed culture 1:1 with S. cerevisiae. Pure culture of S. cerevisiae (p-Sc) was considered as control in both cases.

Figure 6.
Fermentative kinetic of pure and mixed cultures of S. cerevisiae, C. stellata, and S. pombe in: (a) YNB + glucose medium and, (b) YNB + glucose + 30% sorbitol medium.
Figure 6.
Fermentative kinetic of pure and mixed cultures of S. cerevisiae, C. stellata, and S. pombe in: (a) YNB + glucose medium and, (b) YNB + glucose + 30% sorbitol medium.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Yeast strains tested in this study isolated from spontaneous fermentations of grape variety Malvar. Yeast abbreviation included in brackets.
Table 1.
Yeast strains tested in this study isolated from spontaneous fermentations of grape variety Malvar. Yeast abbreviation included in brackets.
| Yeast Code | Yeast Species |
|---|---|
| CLI 920 | Candida stellata (Cs) |
| CLI 1220 | Lachancea fermentati (Lf) |
| CLI 1219 | Lachancea thermotolerans (Lt) |
| CLI 460 | Metschnikowia pulcherrima (Mp) |
| CLI 1217 | Meyerozyma guilliermondii (Mg) |
| CLI 679 | Pichia membranaefaciens (Pm) |
| CLI 2465 | Pichia toletana (Pt) |
| CLI 1221 | Priceomyces carsonii (Pc) |
| CLI 519 | Saccharomyces cerevisiae (Sc) |
| CLI 1085 | Schizosaccharomyces pombe (Sp) |
| CLI 918 | Torulaspora delbrueckii (Td) |
| CLI 1218 | Wickerhamomyces anomalus (Wa) |
| CLI 622 | Zygosaccharomyces bailii (Zb) |
Table 2.
Fermentative parameters produced by pure and mixed cultures with S. cerevisiae, S. pombe and C. stellata in YNB + glucose medium (control) and YNB + glucose + 30% sorbitol (stressful medium). Data are means ± standard deviation (n = 3). Asterisks (*) denoted significant differences regarding the controls, p-Sc, and p-Sc30% (Tukey test; p < 0.05).
Table 2.
Fermentative parameters produced by pure and mixed cultures with S. cerevisiae, S. pombe and C. stellata in YNB + glucose medium (control) and YNB + glucose + 30% sorbitol (stressful medium). Data are means ± standard deviation (n = 3). Asterisks (*) denoted significant differences regarding the controls, p-Sc, and p-Sc30% (Tukey test; p < 0.05).
| Type of Culture | Ethanol (g/L) | Glucose (g/L) | Acetic Acid (g/L) | Glycerol (g/L) | Trehalose (g/L) | pH |
|---|---|---|---|---|---|---|
| p-Sc | 43.79 ± 0.20 | 0.02 ± 0.00 | 0.73 ± 0.10 | 4.29 ± 0.08 | 0.53 ± 0.01 | 2.51 ± 0.02 |
| p-Sp | 12.03 ± 4.58 * | 45.77 ± 8.15 * | 0.02 ± 0.01 * | 1.50 ± 0.76 * | 0.59 ± 0.01 * | 2.99 ± 0.10 * |
| p-Cs | 34.27 ± 1.95 * | 13.53 ± 1.95 * | 0.00 ± 0.00 * | 3.64 ± 0.26 | 0.72 ± 0.01 * | 2.55 ± 0.02 |
| m-Sc/Sp | 44.09 ± 1.17 | 0.00 ± 0.00 | 0.69 ± 0.07 | 4.18 ± 0.11 | 0.56 ± 0.02 * | 2.51 ± 0.01 |
| m-Sc/Cs | 44.54 ± 0.29 | 2.54 ± 0.02 | 0.68 ± 0.10 | 4.23 ± 0.13 | 0.57 ± 0.00 * | 2.55 ± 0.02 |
| p-Sc30% | 33.81 ± 0.17 | 0.02 ± 0.01 | 1.16 ± 0.01 | 11.46 ± 0.11 | 0.92 ± 0.00 | 2.46 ± 0.00 |
| p-Sp30% | 8.99 ± 2.42 * | 35.96 ± 4.52 * | 0.10 ± 0.00 * | 6.83 ± 0.72 * | 0.84 ± 0.00 * | 2.87 ± 0.10 * |
| p-Cs30% | 36.95 ± 0.35 | 0.00 ± 0.00 | 0.00 ± 0.00 * | 9.90 ± 0.59 * | 0.96 ± 0.02 * | 2.48 ± 0.01 |
| m-Sc/Sp30% | 34.19 ± 0.17 | 0.02 ± 0.00 | 1.15 ± 0.06 | 11.59 ± 0.08 | 0.93 ± 0.01 | 2.49 ± 0.01 |
| m-Sc/Cs30% | 34.63 ± 0.06 | 0.03 ± 0.01 | 1.02 ± 0.01 | 11.00 ± 0.03 | 0.93 ± 0.01 | 2.56 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
García, M.; Crespo, J.; Cabellos, J.M.; Arroyo, T. Growth of Non-Saccharomyces Native Strains under Different Fermentative Stress Conditions. Fermentation 2021, 7, 124. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030124
AMA Style
García M, Crespo J, Cabellos JM, Arroyo T. Growth of Non-Saccharomyces Native Strains under Different Fermentative Stress Conditions. Fermentation. 2021; 7(3):124. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030124
Chicago/Turabian StyleGarcía, Margarita, Julia Crespo, Juan Mariano Cabellos, and Teresa Arroyo. 2021. "Growth of Non-Saccharomyces Native Strains under Different Fermentative Stress Conditions" Fermentation 7, no. 3: 124. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030124
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.