
Increased Varietal Aroma Diversity of Marselan Wine by Mixed Fermentation with Indigenous Non-Saccharomyces Yeasts


Abstract
:1. Introduction
2. Materials and Methods
2.1. Marselan Must Samples
2.2. Yeast Strains
2.3. Chemicals
2.4. HS-SPME-GC-MS Analysis
2.5. Fermentation Methods
2.6. Sensory Analysis
2.7. Detection of Physicochemical Parameters
2.8. Statistical Analysis
3. Results
3.1. Aroma Compounds of Marselan Must Detected by GC-MS
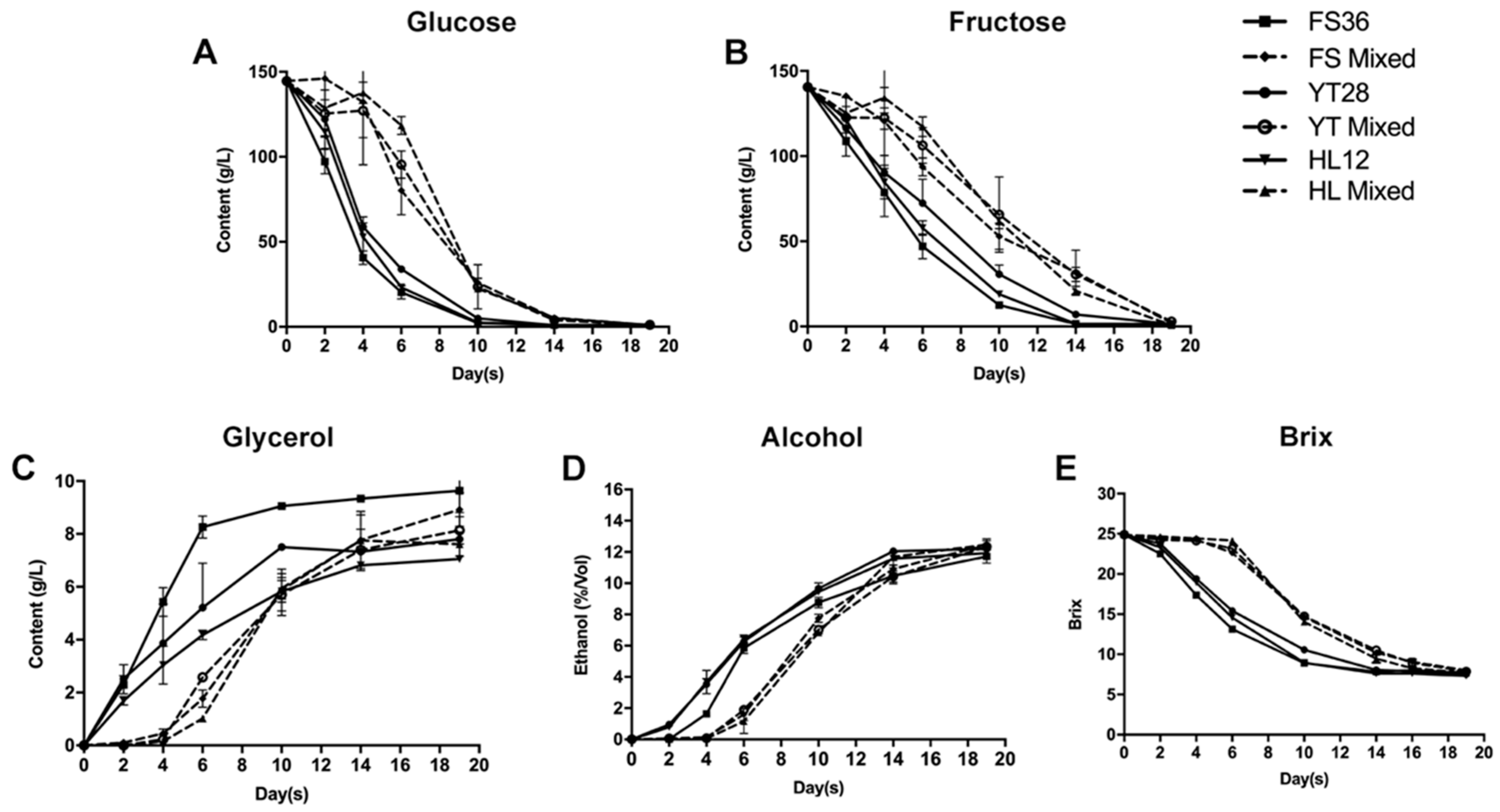
3.2. Basic Parameters of Marselan Wine
3.3. Identification of Aroma Compounds in Marselan Wine by GC-MS
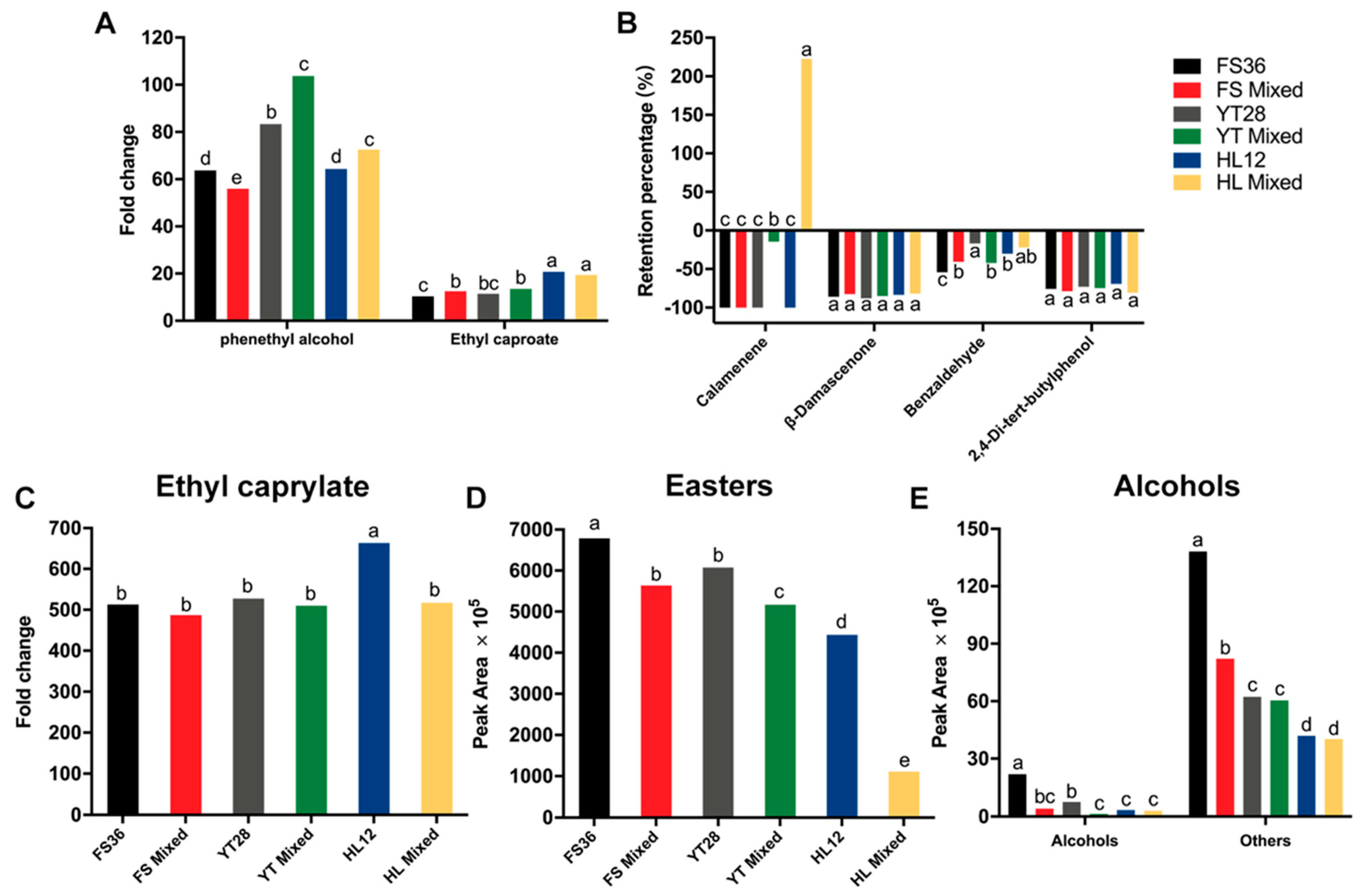
3.4. Comparison of Aroma Compounds between Marselan Must and Wine
3.5. The PCA Analysis of Marselan Wine
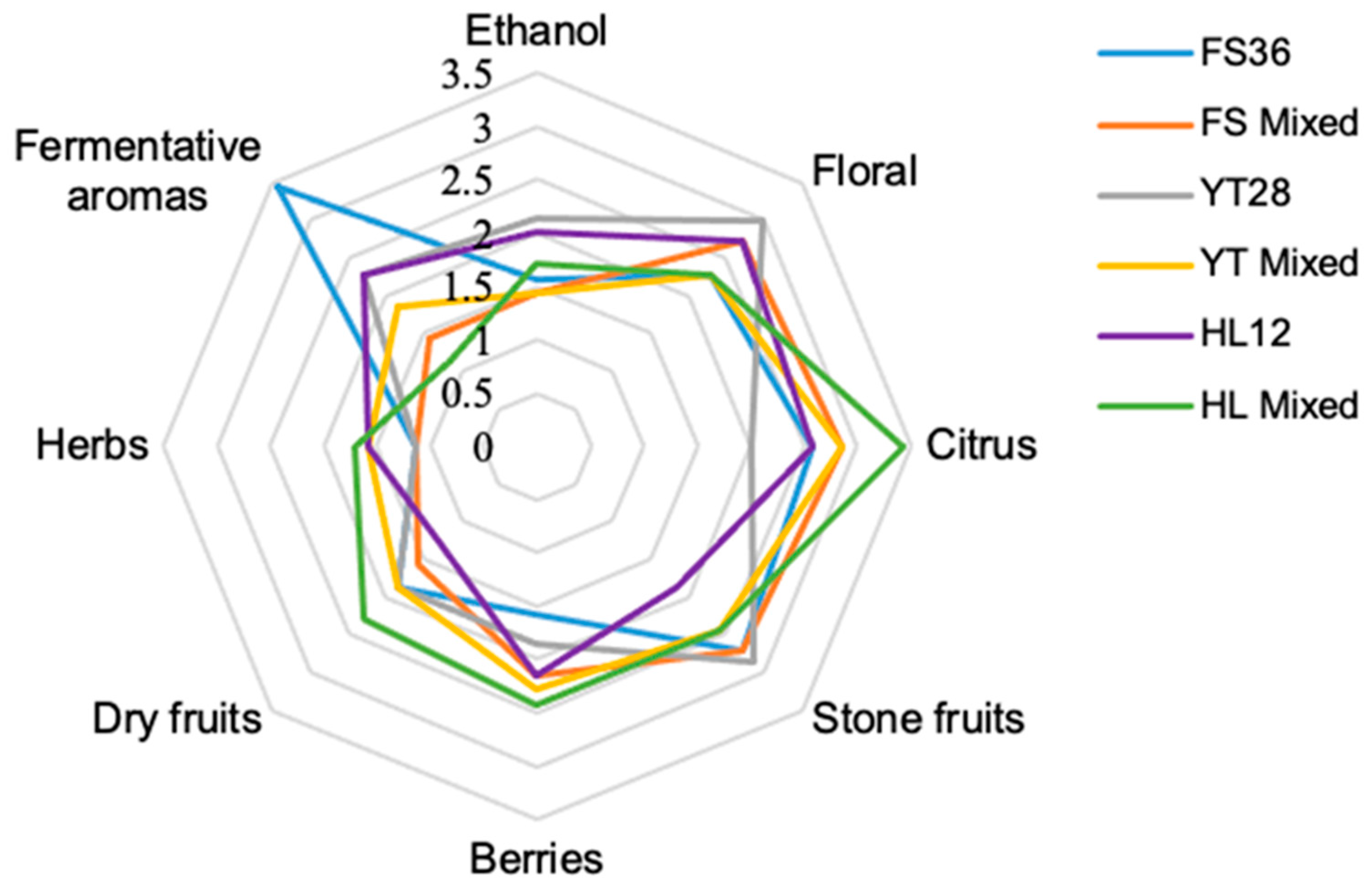
3.6. The Sensory Analysis of Marselan Wine
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Styger, G.; Prior, B.; Bauer, F.F. Wine flavor and aroma. J. Ind. Microbiol. Biotechnol. 2011, 38, 1145–1159. [Google Scholar] [CrossRef]
- Van Wyk, N.; Grossmann, M.; Wendland, J.; Von Wallbrunn, C.; Pretorius, I.S. The Whiff of Wine Yeast Innovation: Strategies for Enhancing Aroma Production by Yeast during Wine Fermentation. J. Agric. Food Chem. 2019, 67, 13496–13505. [Google Scholar] [CrossRef]
- Borren, E.; Tian, B. The Important Contribution of Non-Saccharomyces Yeasts to the Aroma Complexity of Wine: A Review. Foods 2021, 10, 13. [Google Scholar] [CrossRef]
- Alperstein, L.; Gardner, J.M.; Sundstrom, J.F.; Sumby, K.M.; Jiranek, V. Yeast bioprospecting versus synthetic biology—Which is better for innovative beverage fermentation? Appl. Microbiol. Biotechnol. 2020, 104, 1–15. [Google Scholar] [CrossRef]
- Meier-Dörnberg, T.; Kory, O.I.; Jacob, F.; Michel, M.; Hutzler, M. Saccharomyces cerevisiae variety diastaticus friend or foe? Spoilage potential and brewing ability of different Saccharomyces cerevisiae variety diastaticus yeast isolates by genetic, phenotypic and physiological characterization. FEMS Yeast Res. 2018, 18, foy023. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.M.-H.; Boss, P.K.; Walker, M.E.; Sumby, K.M.; Grbin, P.R.; Jiranek, V. Evaluation of indigenous Non-Saccharomyces yeasts isolated from a South Australian vineyard for their potential as wine starter cultures. Int. J. Food Microbiol. 2020, 312, 108373. [Google Scholar] [CrossRef] [PubMed]
- Binati, R.L.; Lemos, J.W.J.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of Non-Saccharomyces yeasts to wine volatile and sensory diversity: A study on Lachancea thermotolerans, Metschnikowia spp. and Starmerella bacillaris strains isolated in Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef]
- Escribano-Viana, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aroma evolution throughout alcoholic fermentation sequentially inoculated with Non-Saccharomyces/Saccharomyces yeasts. Food Res. Int. 2018, 112, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.-T.; Lu, L.; Duan, C.-Q.; Yan, G.-L. The contribution of indigenous Non-Saccharomyces wine yeast to improved aromatic quality of Cabernet Sauvignon wines by spontaneous fermentation. LWT 2016, 71, 356–363. [Google Scholar] [CrossRef]
- Morata, A.; Escott, C.; Bañuelos, M.A.; Loira, I.; Del Fresno, J.M.; González, C.; Suárez-lepe, J.A. Contribution of Non-Saccharomyces yeasts to wine freshness. A review. Biomolecules 2020, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected Non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial Resources and Enological Significance: Opportunities and Benefits. Front. Microbiol. 2017, 8, 995. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Escott, C.; Loira, I.; del Fresno, J.M.; Morata, A.; Tesfaye, W.; Calderon, F.; Suárez-Lepe, J.A.; Han, S.; Benito, S. Use of Non-Saccharomyces yeasts and oenological tannin in red winemaking: Influence on colour, aroma and sensorial properties of young wines. Food Microbiol. 2018, 69, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Carrau, F.M.; Rivas-Gonzalo, J.C.; Alcalde-eon, C.; Boido, E.; Dellacassa, E. Pigment Profiles in Monovarietal Wines Produced in Uruguay. Am. J. Enol. Vitic. 2006, 57, 449–459. [Google Scholar]
- Robinson, J.; Harding, J.; Vouillamoz, J. Wine Grapes: A Complete Guide to 1368 Vine Varieties, Including Their Origins and Flavours; Penguin: London, UK, 2013. [Google Scholar]
- Sagratini, G.; Maggi, F.; Caprioli, G.; Cristalli, G.; Ricciutelli, M.; Torregiani, E.; Vittori, S. Comparative study of aroma profile and phenolic content of Montepulciano monovarietal red wines from the Marches and Abruzzo regions of Italy using HS-SPME–GC–MS and HPLC–MS. Food Chem. 2012, 132, 1592–1599. [Google Scholar] [CrossRef] [PubMed]
- Suzzi, G.; Arfelli, G.; Schirone, M.; Corsetti, A.; Perpetuini, G.; Tofalo, R. Effect of grape indigenous Saccharomyces cerevisiae strains on Montepulciano d’Abruzzo red wine quality. Food Res. Int. 2012, 46, 22–29. [Google Scholar] [CrossRef]
- Sun, Y.; Li, E.; Qi, X.; Liu, Y. Changes of diversity and population of yeasts during the fermentations by pure and mixed inoculation of Saccharomyces cerevisiae strains. Ann. Microbiol. 2015, 65, 911–919. [Google Scholar] [CrossRef]
- Lyu, J.; Ma, Y.; Xu, Y.; Nie, Y.; Tang, K. Characterization of the Key Aroma Compounds in Marselan Wine by Gas Chromatography-Olfactometry, Quantitative Measurements, Aroma Recombination, and Omission Tests. Molecules 2019, 24, 2978. [Google Scholar] [CrossRef] [Green Version]
- Conde, C.; Agasse, A.; Glissant, D.; Tavares, R.; Gerós, H.; Delrot, S. Pathways of Glucose Regulation of Monosaccharide Transport in Grape Cells. Plant Physiol. 2006, 141, 1563–1577. [Google Scholar] [CrossRef] [Green Version]
- Tronchoni, J.; Gamero, A.; Arroyo-López, F.N.; Barrio, E.; Querol, A. Differences in the glucose and fructose consumption profiles in diverse Saccharomyces wine species and their hybrids during grape juice fermentation. Int. J. Food Microbiol. 2009, 134, 237–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loureiro, V.; Malfeito-Ferreira, M. Spoilage yeasts in the wine industry. Int. J. Food Microbiol. 2003, 86, 23–50. [Google Scholar] [CrossRef]
- Kelebek, H.; Kesen, S.; Sonmezdag, A.S.; Cetiner, B.; Kola, O.; Selli, S. Characterization of the key aroma compounds in tomato pastes as affected by hot and cold break process. J. Food Meas. Charact. 2018, 12, 2461–2474. [Google Scholar] [CrossRef]
- Comuzzo, P.; Tat, L.; Tonizzo, A.; Battistutta, F. Yeast derivatives (extracts and autolysates) in winemaking: Release of volatile compounds and effects on wine aroma volatility. Food Chem. 2006, 99, 217–230. [Google Scholar] [CrossRef]
- González-Barreiro, C.; Rial-Otero, R.; Cancho-Grande, B.; Simal-Gándara, J. Wine Aroma Compounds in Grapes: A Critical Review. Crit. Rev. Food Sci. Nutr. 2015, 55, 202–218. [Google Scholar] [CrossRef] [PubMed]
- Rapp, A.; Versini, G. Influence of nitrogen compounds in grapes on aroma compounds of wines. Dev. Food Sci. 1995, 37, 1659–1694. [Google Scholar] [CrossRef]
- Culleré, L.; Cacho, J.; Ferreira, V. An assessment of the role played by some oxidation-related aldehydes in wine aroma. J. Agric. Food Chem. 2007, 55, 876–881. [Google Scholar] [CrossRef]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J.C. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot berries. Am. J. Enol. Vitic. 2002, 53, 171–182. [Google Scholar] [CrossRef]
- Hernández-Orte, P.; Cersosimo, M.; Loscos, N.; Cacho, J.; Garcia-Moruno, E.; Ferreira, V. The development of varietal aroma from non-floral grapes by yeasts of different genera. Food Chem. 2008, 107, 1064–1077. [Google Scholar] [CrossRef]
- Ewart, A.; Brien, C.; Soderlund, R.; Smart, R. The effects of light pruning, irrigation and improved soil management on wine quality of the Vitis vinifera cv. Riesling. Vitis Geilweilerhof. 1985, 24, 209–217. [Google Scholar]
- Ebeler, S.E.; Thorngate, J.H. Wine chemistry and flavor: Looking into the crystal glass. J. Agric. Food Chem. 2009, 57, 8098–8108. [Google Scholar] [CrossRef]
- King, A.; Dickinson, J.R. Biotransformation of monoterpene alcohols by Saccharomyces cerevisiae, Torulaspora delbrueckii and Kluyveromyces lactis. Yeast 2000, 16, 499–506. [Google Scholar] [CrossRef]
- Mateo, J.J.; Jiménez, M. Monoterpenes in grape juice and wines. J. Chromatogr. A 2000, 881, 557–567. [Google Scholar] [CrossRef]
- Pogorzelski, E.; Wilkowska, A. Flavour enhancement through the enzymatic hydrolysis of glycosidic aroma precursors in juices and wine beverages: A review. Flavour Fragr. J. 2010, 22, 251–254. [Google Scholar] [CrossRef]
- Liang, Z.; Fang, Z.; Pai, A.; Luo, J.; Gan, R.; Gao, Y.; Lu, J.; Zhang, P. Glycosidically bound aroma precursors in fruits: A comprehensive review. Crit. Rev. Food Sci. Nutr. 2020, 1–29. [Google Scholar] [CrossRef]
- Yang, Y.; Jin, G.-J.; Wang, X.-J.; Kong, C.-L.; Liu, J.; Tao, Y.-S. Chemical profiles and aroma contribution of terpene compounds in Meili (Vitis vinifera L.) grape and wine. Food Chem. 2019, 284, 155–161. [Google Scholar] [CrossRef]
- Holt, S.; Miks, M.H.; De Carvalho, B.T.; Foulquié-Moreno, M.R.; Thevelein, J.M. The molecular biology of fruity and floral aromas in beer and other alcoholic beverages. FEMS Microbiol. Rev. 2019, 43, 193–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alem, H.; Rigou, P.; Schneider, R.; Ojeda, H.; Torregrosa, L. Impact of agronomic practices on grape aroma composition: A review. J. Sci. Food Agric. 2019, 99, 975–985. [Google Scholar] [CrossRef] [PubMed]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [Green Version]
- Padilla, B.; Zulian, L.; Ferreres, À.; Pastor, R.; Esteve-zarzoso, B.; Beltran, G.; Mas, A. Sequential Inoculation of Native Non- Saccharomyces and Saccharomyces cerevisiae Strains for Wine Making. Front. Microbiol. 2017, 8, 1293. [Google Scholar] [CrossRef]
- Renault, P.; Coulon, J.; de Revel, G.; Barbe, J.C.; Bely, M. Increase of fruity aroma during mixed T. delbrueckii/S. cerevisiae wine fermentation is linked to specific esters enhancement. Int. J. Food Microbiol. 2015, 207, 40–48. [Google Scholar] [CrossRef]
- Tofalo, R.; Patrignani, F.; Lanciotti, R.; Perpetuini, G.; Schirone, M.; Di Gianvito, P.; Pizzoni, D.; Arfelli, G.; Suzzi, G. Aroma profile of montepulciano d’abruzzo wine fermented by single and co-culture starters of autochthonous Saccharomyces and Non-Saccharomyces yeasts. Front. Microbiol. 2016, 7, 610. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef]
- Terrell, E.; Cliff, M.A.; Van Vuuren, H.J.J. Functional characterization of individual- and mixed-Burgundian Saccharomyces cerevisiae isolates for fermentation of pinot noir. Molecules 2015, 20, 5112–5136. [Google Scholar] [CrossRef]
- Hu, K.; Jin, G.-J.; Mei, W.-C.; Li, T.; Tao, Y.-S. Increase of medium-chain fatty acid ethyl ester content in mixed H. uvarum/S. cerevisiae fermentation leads to wine fruity aroma enhancement. Food Chem. 2018, 15, 495–501. [Google Scholar] [CrossRef]
- Du Toit, M.; Pretorius, I.S. Microbial Spoilage and Preservation of Wine: Using Weapons from Nature’s Own Arsenal. S. Afr. J. Enol. Vitic. 2000, 21, 74–76. [Google Scholar] [CrossRef] [Green Version]
- Minnaar, P.P.; Du Plessis, H.W.; Jolly, N.P.; Van Der Rijst, M.; Du Toit, M. Non-Saccharomyces yeast and lactic acid bacteria in Co-inoculated fermentations with two Saccharomyces cerevisiae yeast strains: A strategy to improve the phenolic content of Syrah wine. Food Chem. 2019, 11, 100070. [Google Scholar] [CrossRef] [PubMed]
- Vilela, A. Lachancea thermotolerans, the Non-Saccharomyces yeast that reduces the volatile acidity of wines. Fermentation 2018, 4, 56. [Google Scholar] [CrossRef] [Green Version]
- Ivit, N.N.; Kemp, B. The impact of Non-Saccharomyces yeast on traditional method sparkling Wine. Fermentation 2018, 4, 73. [Google Scholar] [CrossRef] [Green Version]
- Dutraive, O.; Benito, S.; Fritsch, S.; Beisert, B.; Patz, C.-D.; Rauhut, D. Effect of sequential inoculation with Non-Saccharomyces and Saccharomyces yeasts on Riesling wine chemical composition. Fermentation 2019, 5, 79. [Google Scholar] [CrossRef] [Green Version]
- Rantsiou, K.; Dolci, P.; Giacosa, S.; Torchio, F.; Tofalo, R.; Torriani, S.; Suzzi, G.; Rolle, L.; Cocolin, L. Candida zemplinina can reduce acetic acid produced by Saccharomyces cerevisiae in sweet wine fermentations. Appl. Environ. Microbiol. 2012, 78, 1987–1994. [Google Scholar] [CrossRef] [Green Version]
- Domizio, P.; Liu, Y.; Bisson, L.F.; Barile, D. Cell wall polysaccharides released during the alcoholic fermentation by SchizoSaccharomyces pombe and S. japonicus: Quantification and characterization. Food Microbiol. 2017, 61, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Di Gianvito, P.; Perpetuini, G.; Tittarelli, F.; Schirone, M.; Arfelli, G.; Piva, A.; Patrignani, F.; Lanciotti, R.; Olivastri, L.; Suzzi, G. Impact of Saccharomyces cerevisiae strains on traditional sparkling wines production. Food Res. Int. 2018, 109, 552–560. [Google Scholar] [CrossRef]
- Coloretti, F.; Zambonelli, C.; Tini, V. Characterization of flocculent Saccharomyces interspecific hybrids for the production of sparkling wines. Food Microbiol. 2006, 23, 672–676. [Google Scholar] [CrossRef]
- Kutyna, D.R.; Varela, C.; Stanley, G.A.; Borneman, A.R.; Henschke, P.A.; Chambers, P.J. Adaptive evolution of Saccharomyces cerevisiae to generate strains with enhanced glycerol production. Appl. Microbiol. Biotechnol. 2012, 93, 1175–1184. [Google Scholar] [CrossRef]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its Importance to Wine Aroma-A Review. S. Afr. J. Enol. Vitic. 2019, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with Non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Zhang, B.Q.; Luan, Y.; Duan, C.Q.; Yan, G.L. Use of Torulaspora delbrueckii Co-fermentation with two Saccharomyces cerevisiae Strains with different aromatic characteristic to improve the diversity of red wine aroma profile. Front. Microbiol. 2018, 5, 606. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Jin, G.-J.; Xu, Y.-H.; Tao, Y.-S. Wine aroma response to different participation of selected Hanseniaspora uvarum in mixed fermentation with Saccharomyces cerevisiae. Food Res. Int. 2018, 108, 119–127. [Google Scholar] [CrossRef]
- Zhang, B.-Q.; Shen, J.-Y.; Duan, C.-Q.; Yan, G.-L. Use of indigenous Hanseniaspora vineae and Metschnikowia pulcherrima co-fermentation with Saccharomyces cerevisiae to improve the aroma diversity of Vidal Blanc icewine. Front. Microbiol. 2018, 9, 2303. [Google Scholar] [CrossRef]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Region | Species | Property |
|---|---|---|---|
| FS36 | Fanghshan, Beijing | Saccharomyces cerevisiae | Saccharomyces |
| FS31 | Fanghshan, Beijing | Kazachstania exigua | Non-Saccharomyces |
| YT28 | Yantai, Shandong | Saccharomyces cerevisiae | Saccharomyces |
| YT2 | Yantai, Shandong | Candida glabrata | Non-Saccharomyces |
| HL12 | Huailai, Hebei | Saccharomyces cerevisiae | Saccharomyces |
| HL9 | Huailai, Hebei | Hanseniaspora opuntiae | Non-Saccharomyces |
| Fermentation Trails | Yeasts | Method |
|---|---|---|
| FS36 | FS36 alone | / |
| FS Mixed | FS31 + FS36 | FS36 was inoculated on 4th day |
| YT28 | YT28 alone | / |
| YT Mixed | YT2 + YT28 | YT28 was inoculated on 4th day |
| HL12 | HL12 alone | / |
| HL Mixed | HL9 + HL12 | HL12 was inoculated on 4th day |
| 2017 HL | 2018 FS | 2018 YT | 2018 HL | |||||
|---|---|---|---|---|---|---|---|---|
| Aroma Compound | Peak Area * 106 | RPA% | Peak Area * 106 | RPA% | Peak Area * 106 | RPA% | Peak Area * 106 | RPA% |
| C13-Norisoprenoids | ||||||||
| Violine | N.D. | 0.00 | 5.42 ± 0.12 a | 1.10 | 2.35 ± 0.09 b | 0.24 | 2.17 ± 0.01 b | 0.24 |
| Calamenene | 1.37 ± 0.33 d | 0.42 | 3.45 ± 0.10 a | 0.72 | 2.28 ± 0.39 b | 0.23 | 1.77 ± 0.22 c | 0.20 |
| β-Damascone | 18.35 ± 0.42 d | 5.60 | 32.82 ± 1.01 a | 6.83 | 30.69 ± 0.59 b | 3.16 | 29.29 ± 0.72 c | 3.25 |
| Terpenes | ||||||||
| Linalol | N.D. | 0.00 | N.D. | 0.00 | N.D. | 0.00 | 0.53 ± 0.01 | 0.06 |
| 4-terpineol | N.D. | 0.00 | N.D. | 0.00 | N.D. | 0.00 | 0.57 ± 0.03 | 0.06 |
| α-terpene | N.D. | 0.00 | N.D. | 0.00 | 0.91 ± 0.01 | 0.09 | N.D. | 0.00 |
| β-myrcene | N.D. | 0.00 | N.D. | 0.00 | N.D. | 0.00 | 2.81 ± 0.13 | 0.31 |
| Semenene | 1.65 ± 0.11 c | 0.50 | 2.35 ± 0.30 b | 0.49 | 2.40 ± 0.14 b | 0.25 | 3.34 ± 0.14 a | 0.37 |
| Alcohols | ||||||||
| Phenylethyl alcohol | 4.46 ± 0.97 a | 1.36 | 2.44 ± 0.26 ab | 0.51 | 1.40 ± 0.10 b | 0.14 | 1.19 ± 0.15 b | 0.13 |
| 1-hexanol | N.D. | 0.00 | N.D. | 0.00 | 659.27 ± 2.14 a | 67.80 | 555.95 ± 3.67 b | 61.70 |
| 1-hexene-3-ol | N.D. | 0.00 | N.D. | 0.00 | N.D. | 0.00 | 6.59 ± 0.33 | 0.73 |
| Esters | ||||||||
| Hexyl acetate | N.D. | 0.00 | N.D. | 0.00 | 4.45 ± 0.12 a | 0.37 | 2.48 ± 0.04 a | 0.28 |
| Ethyl octoate | 1.50 ± 0.28 a | 0.31 | 0.63 ± 0.01 a | 0.13 | 0.40 ± 0.06 a | 0.04 | 0.24 ± 0.00 a | 0.03 |
| Phenethyl acetate | 1.42 ± 0.02 | 0.43 | N.D. | 0.00 | N.D. | 0.00 | N.D. | 0.00 |
| Ethyl benzoate | 0.52 ± 0.07 a | 0.16 | 0.50 ± 0.07 a | 0.10 | 0.34 ± 0.05 a | 0.03 | 0.28 ± 0.01 a | 0.03 |
| Ethyl hexanoate | 4.33 ± 0.21 a | 0.99 | 0.82 ± 0.19 a | 0.17 | 0.50 ± 0.01 a | 0.05 | 0.53 ± 0.09 a | 0.06 |
| Ethyl phthalate | N.D. | 0.00 | 0.14 ± 0.00 a | 0.03 | 0.07 ± 0.00 b | 0.01 | N.D. | 0.00 |
| Ethyl myristate | 1.88 ± 0.83 a | 0.57 | 0.28 ± 0.05 a | 0.06 | 0.09 ± 0.00 a | 0.01 | N.D. | 0.00 |
| Ethyl palmitate | 1.85 ± 0.02 a | 0.56 | 0.33 ± 0.11 b | 0.07 | 0.14 ± 0.02 a | 0.01 | N.D. | 0.00 |
| Ethyl laurate | 5.93 ± 0.31 a | 1.81 | 1.56 ± 0.37 a | 0.32 | 1.11 ± 0.01 a | 0.11 | 0.36 ± 0.00 a | 0.04 |
| Others | ||||||||
| n-hexanal | 152.28 ± 2.54 a | 46.50 | 81.57 ± 0.06 b | 17.00 | 53.60 ± 1.25 c | 5.51 | 94.70 ± 0.22 b | 10.50 |
| 2-hexenal | N.D. | 0.00 | 129.77 ± 2.34 | 27.00 | N.D. | 0.00 | N.D. | 0.00 |
| Benzaldehyde | 6.01 ± 0.56 b | 1.83 | 15.65 ± 0.22 a | 3.26 | 14.02 ± 0.57 ab | 1.44 | 2.73 ± 0.03 b | 0.30 |
| Nonanal | 4.17 ± 0.03 a | 0.97 | 1.77 ± 0.03 b | 0.37 | 1.16 ± 0.16 c | 0.12 | 1.00 ± 0.00 c | 0.11 |
| 2-decanone | N.D. | 0.00 | 0.93 ± 0.01 a | 0.19 | 1.06 ± 0.05 a | 0.11 | 1.02 ± 0.00 a | 0.11 |
| 2,4-di-tert-butylphenol | 30.08 ± 0.37 a | 9.18 | 13.41 ± 0.07 b | 2.79 | 10.34 ± 0.05 b | 1.06 | 10.17 ± 1.20 b | 1.13 |
| FS36 | YT28 | HL12 | FS Mixed | YT Mixed | HL Mixed | |
|---|---|---|---|---|---|---|
| Aroma Compound | Peak Area * 106 | Peak Area * 106 | Peak Area * 106 | Peak Area * 106 | Peak Area * 106 | Peak Area * 106 |
| Norisoprenes | ||||||
| Calamenene | N.D. | N.D. | N.D. | N.D. | 1.17 ± 0.01 b | 4.42 ± 0.03 a |
| β-Damascone | 2.60 ± 0.20 c | 2.20 ± 0.09 d | 3.03 ± 0.15 b | 3.18 ± 0.08 ab | 2.77 ± 0.06 c | 3.34 ± 0.18 a |
| Alcohols | ||||||
| Phenethyl alcohol | 284.27 ± 3.75 de | 371.74 ± 1.97 c | 287.04 ± 0.91 de | 249.47 ± 0.89 e | 462.46 ± 0.29 a | 323.63 ± 1.02 d |
| Decyl alcohol | 2.07 ± 0.07 a | N.D. | 1.84 ± 0.07 b | N.D. | N.D. | N.D. |
| Trans β-farnesol | 6.82 ± 0.70 a | 3.68 ± 0.59 b | N.D. | 1.33 ± 0.16 c | N.D. | 1.06 ± 0.11 d |
| β-bisabolol | 4.02 ± 0.81 a | 1.28 ± 0.23 b | 1.04 ± 0.21 b | 2.02 ± 0.11 ab | 1.32 ± 0.27 b | 1.40 ± 0.21 b |
| Cis-α-bisabolol | 0.70 ± 0.15 a | 0.54 ± 0.10 a | 0.50 ± 0.16 a | 0.88 ± 0.07 a | 0.11 ± 0.02 a | 0.66 ± 0.32 a |
| Esters | ||||||
| Ethyl hexanoate | 33.62 ± 6.22 b | 37.08 ± 2.62 b | 67.43 ± 1.20 a | 40.74 ± 0.76 b | 43.93 ± 0.94 b | 63.21 ± 1.39 a |
| Hexyl acetate | N.D. | N.D. | N.D. | 2.45 ± 0.44 a | 2.55 ± 0.87 a | 3.40 ± 0.30 a |
| Ethyl octanoate | 771.21 ± 5.13 b | 792.82 ± 31.12 b | 996.70 ± 8.57 a | 732.79 ± 14.52 b | 766.20 ± 1.45 b | 777.31 ± 7.79 b |
| 7-Ethyl caprylate | 3.38 ± 0.78 b | 3.11 ± 0.05 b | 3.00 ± 0.85 b | 2.52 ± 0.56 b | 5.70 ± 0.82 a | 4.56 ± 0.13 ab |
| 3-Methylbutyl octanoate | 3.24 ± 0.79 b | 7.29 ± 1.42 a | 6.27 ± 0.18 ab | 4.95 ± 0.36 ab | 6.20 ± 1.42 ab | 3.87 ± 0.40 b |
| Ethyl benzoate | 0.72 ± 0.13 b | 1.31 ± 0.15 ab | 3.42 ± 0.31 a | 1.07 ± 0.23 ab | 1.88 ± 0.38 ab | 1.68 ± 0.44 ab |
| Diethyl succinate | 2.07 ± 0.12 bc | 1.83 ± 0.34 bc | 3.06 ± 0.06 ab | 1.27 ± 0.20 c | 2.34 ± 0.44 b | 3.56 ± 0.09 a |
| Phenethyl acetate | 39.51 ± 6.42 c | 51.78 ± 1.70 bc | 40.29 ± 3.12 c | 43.98 ± 0.75 c | 72.26 ± 1.03 a | 56.46 ± 0.56 b |
| 2-Methyl propyl caprylate | 2.03 ± 0.08 a | 2.03 ± 0.12 a | 2.05 ± 0.15 a | 1.81 ± 0.40 a | 2.04 ± 0.14 a | 2.49 ± 0.06 a |
| Ethyl decanoate | 3493.96 ± 43.58 a | 3455.40 ± 23.77 a | 3154.25 ± 24.12 ab | 3051.76 ± 7.57 ab | 2577.69 ± 19.87 b | 2520.97 ± 15.39 b |
| Ethyl 9-decanoate | 2670.03 ± 28.46 a | 2093.40 ± 18.67 b | 990.66 ± 7.37 c | 1992.78 ± 17.91 b | 2261.45 ± 3.09 ab | 850.23 ± 8.75 c |
| Ethyl oleate | 23.73 ± 1.82 a | 6.22 ± 0.54 bc | 4.82 ± 1.38 bc | 11.11 ± 0.81 b | 3.21 ± 1.26 c | 3.39 ± 0.25 c |
| Ethyl propionate | N.D. | N.D. | N.D. | 11.96 ± 1.20 a | 3.28 ± 1.26 a | 3.83 ± 1.40 a |
| Trans ethyl 4-decanoate | 16.98 ± 3.64 a | 16.65 ± 2.52 a | 6.70 ± 1.07 b | 11.88 ± 0.96 ab | 4.10 ± 1.61 b | N.D. |
| Isoamyl caprylate | 52.56 ± 6.71 a | 48.57 ± 1.07 a | N.D. | 42.43 ± 2.70 a | N.D. | N.D. |
| Propyl decanoate | 4.91 ± 0.22 a | 2.91 ± 0.48 b | 2.08 ± 0.29 c | 4.21 ± 2.74 a | 2.35 ± 0.80 bc | 1.67 ± 0.75 c |
| Ethyl undecanoate | 2.48 ± 0.46 ab | 2.29 ± 0.60 b | 0.97 ± 0.02 c | 2.90 ± 0.34 a | 2.03 ± 0.09 b | 0.93 ± 0.14 c |
| Ethyl laurate | 65.84 ± 0.96 ab | 51.71 ± 0.08 b | 33.36 ± 0.84 c | 71.22 ± 0.10 a | 41.28 ± 0.15 bc | 21.37 ± 0.23 c |
| Ethyl palmitate | 149.85 ± 6.42 a | 158.53 ± 1.18 a | 68.69 ± 1.24 b | 156.30 ± 1.62 a | 129.55 ± 0.37 a | 46.82 ± 1.17 b |
| Others | ||||||
| Benzaldehyde | 2.75 ± 0.21 b | 4.99 ± 0.07 ab | 4.20 ± 0.15 b | 3.57 ± 0.04 b | 3.47 ± 0.05 b | 4.67 ± 0.06 ab |
| Phenylacetaldehyde | 2.14 ± 0.05 b | 5.39 ± 0.16 a | 4.80 ± 0.04 a | 5.55 ± 0.18 a | 4.73 ± 0.08 a | 3.80 ± 0.21 ab |
| 16-octadecenal | 1.63 ± 0.23 | N.D. | N.D. | N.D. | N.D. | N.D. |
| Ethyl benzaldehyde | 25.42 ± 0.91 a | 11.71 ± 1.03 b | 12.17 ± 0.67 b | 10.21 ± 0.12 b | 10.66 ± 0.30 b | 8.49 ± 0.08 b |
| Curcumene | 4.63 ± 0.10 a | 5.10 ± 0.07 a | N.D. | 5.50 ± 0.58 a | 4.30 ± 0.09 a | 6.15 ± 0.18 a |
| Cyclohexene | 31.26 ± 1.21 a | N.D. | N.D. | 16.76 ± 0.20 b | N.D. | N.D. |
| 2,4-Di-tert-butylphenol | 7.26 ± 0.05 b | 8.11 ± 0.14 b | 9.23 ± 0.13 b | 6.35 ± 0.11 b | 7.54 ± 0.01 b | 5.82 ± 0.07 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xi, X.; Xin, A.; You, Y.; Huang, W.; Zhan, J. Increased Varietal Aroma Diversity of Marselan Wine by Mixed Fermentation with Indigenous Non-Saccharomyces Yeasts. Fermentation 2021, 7, 133. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030133
Xi X, Xin A, You Y, Huang W, Zhan J. Increased Varietal Aroma Diversity of Marselan Wine by Mixed Fermentation with Indigenous Non-Saccharomyces Yeasts. Fermentation. 2021; 7(3):133. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030133
Chicago/Turabian StyleXi, Xiaomin, Aili Xin, Yilin You, Weidong Huang, and Jicheng Zhan. 2021. "Increased Varietal Aroma Diversity of Marselan Wine by Mixed Fermentation with Indigenous Non-Saccharomyces Yeasts" Fermentation 7, no. 3: 133. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030133