
A Comprehensive Bioprocessing Approach to Foster Cheese Whey Valorization: On-Site β-Galactosidase Secretion for Lactose Hydrolysis and Sequential Bacterial Cellulose Production

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Strains and Media
2.2. Crude Enzyme Production and Cheese Whey Hydrolysis
2.3. Submerged Fermentation and Bacterial Cellulose (BC) Production
2.4. Analytical Methods
2.5. Statistical Analysis
3. Results and Discussion
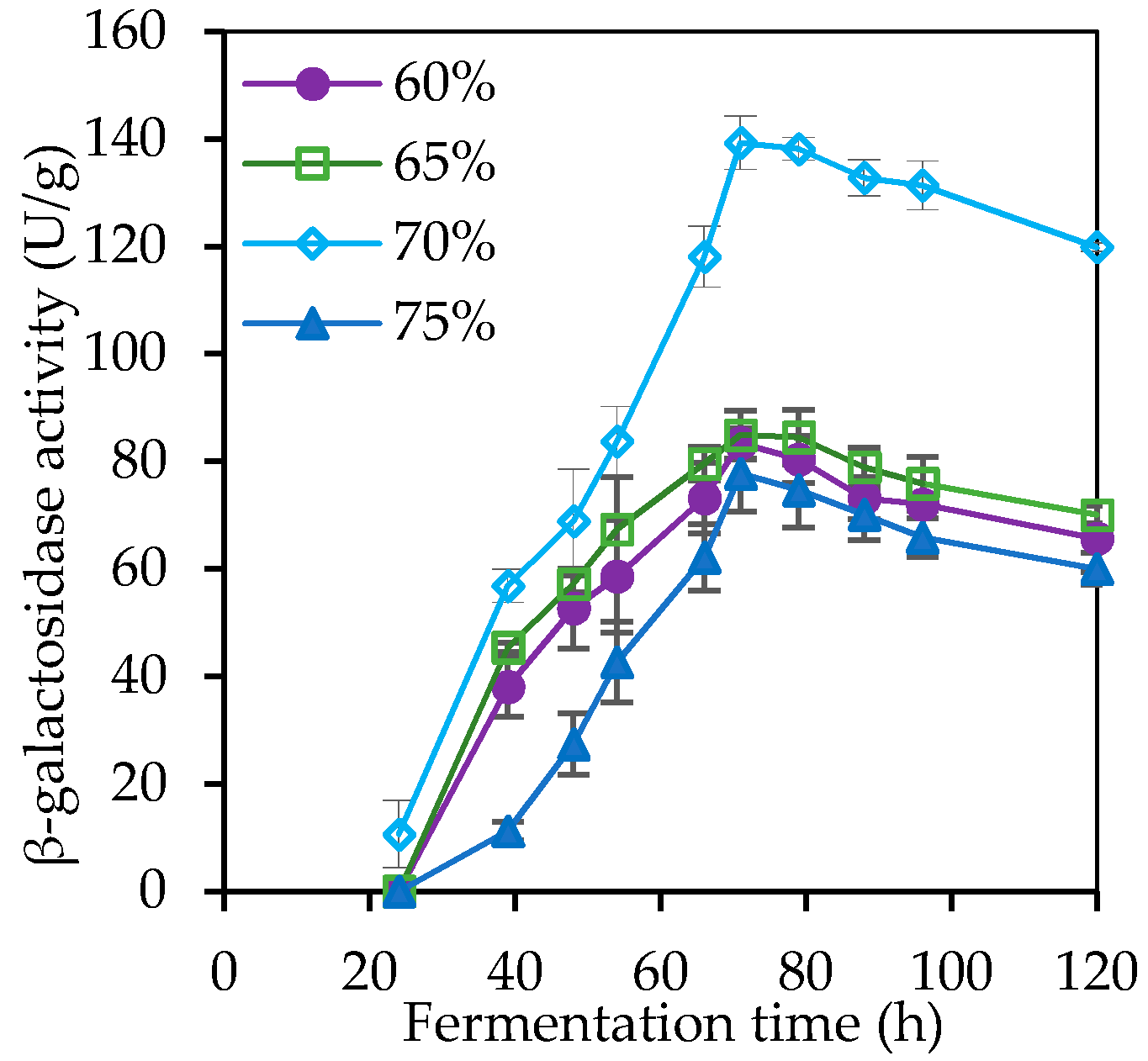
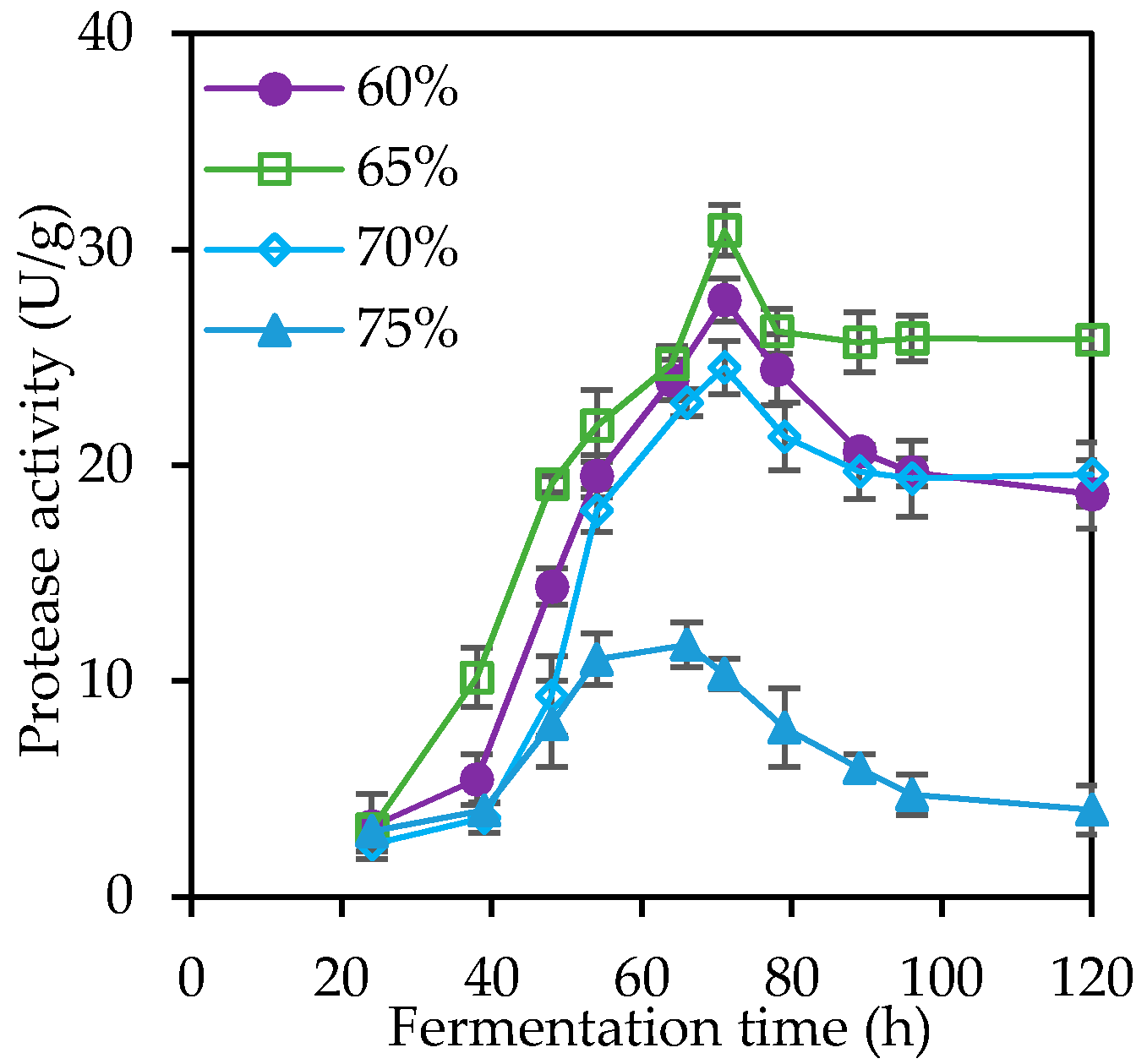
3.1. Solid State Fermentation (SSF) and Crude β-Galactosidase Production
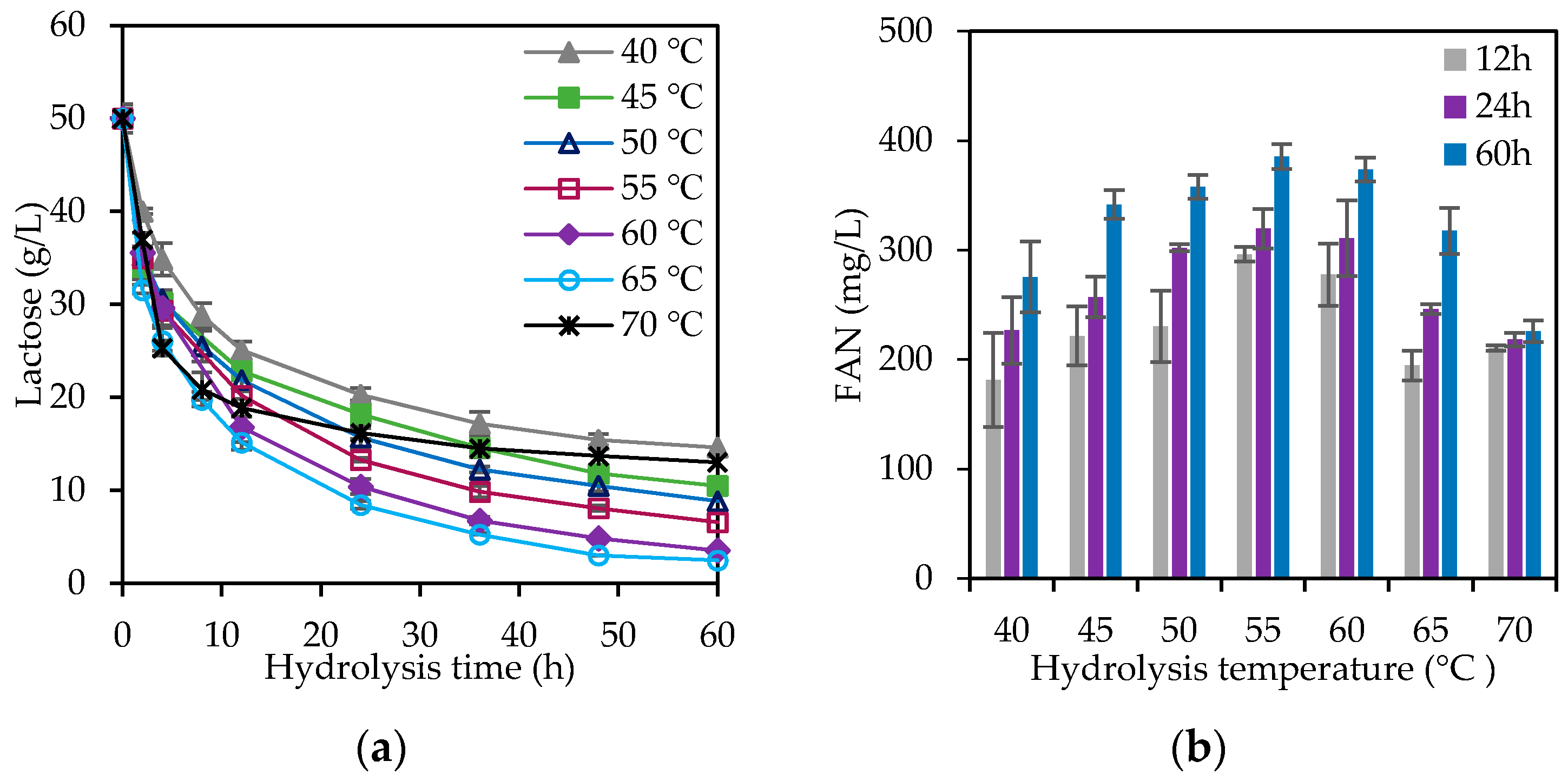
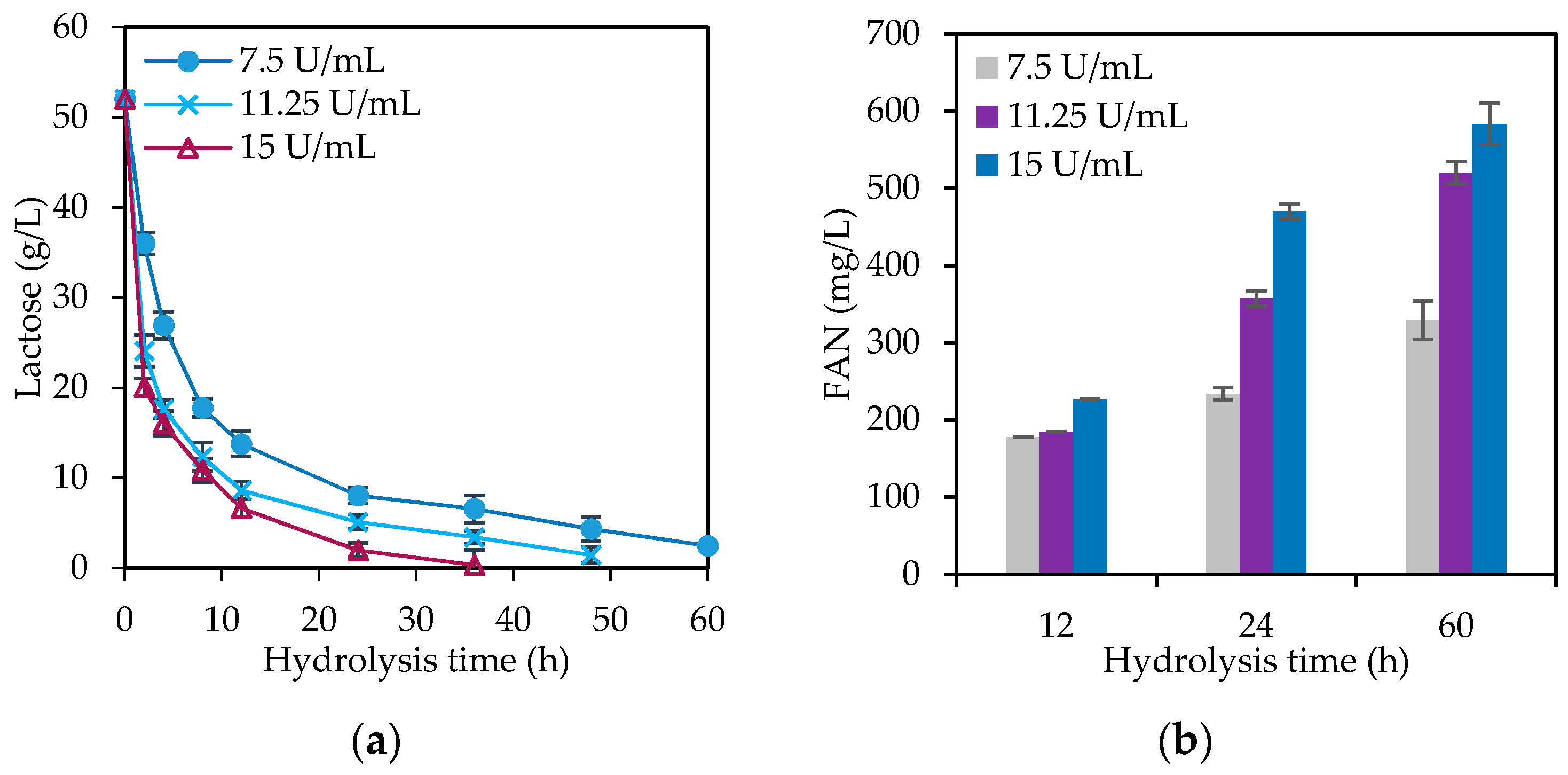
3.2. Cheese Whey Hydrolysis Study
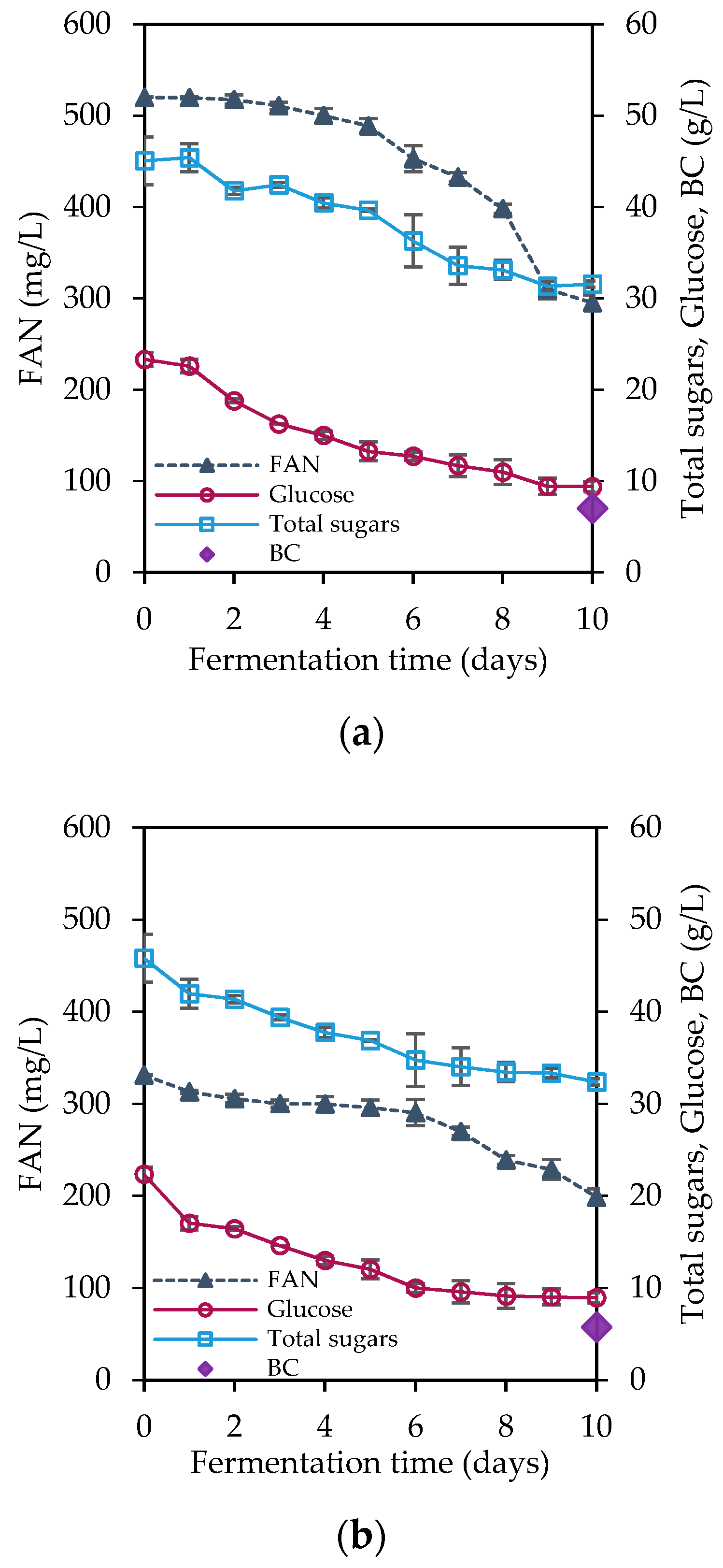
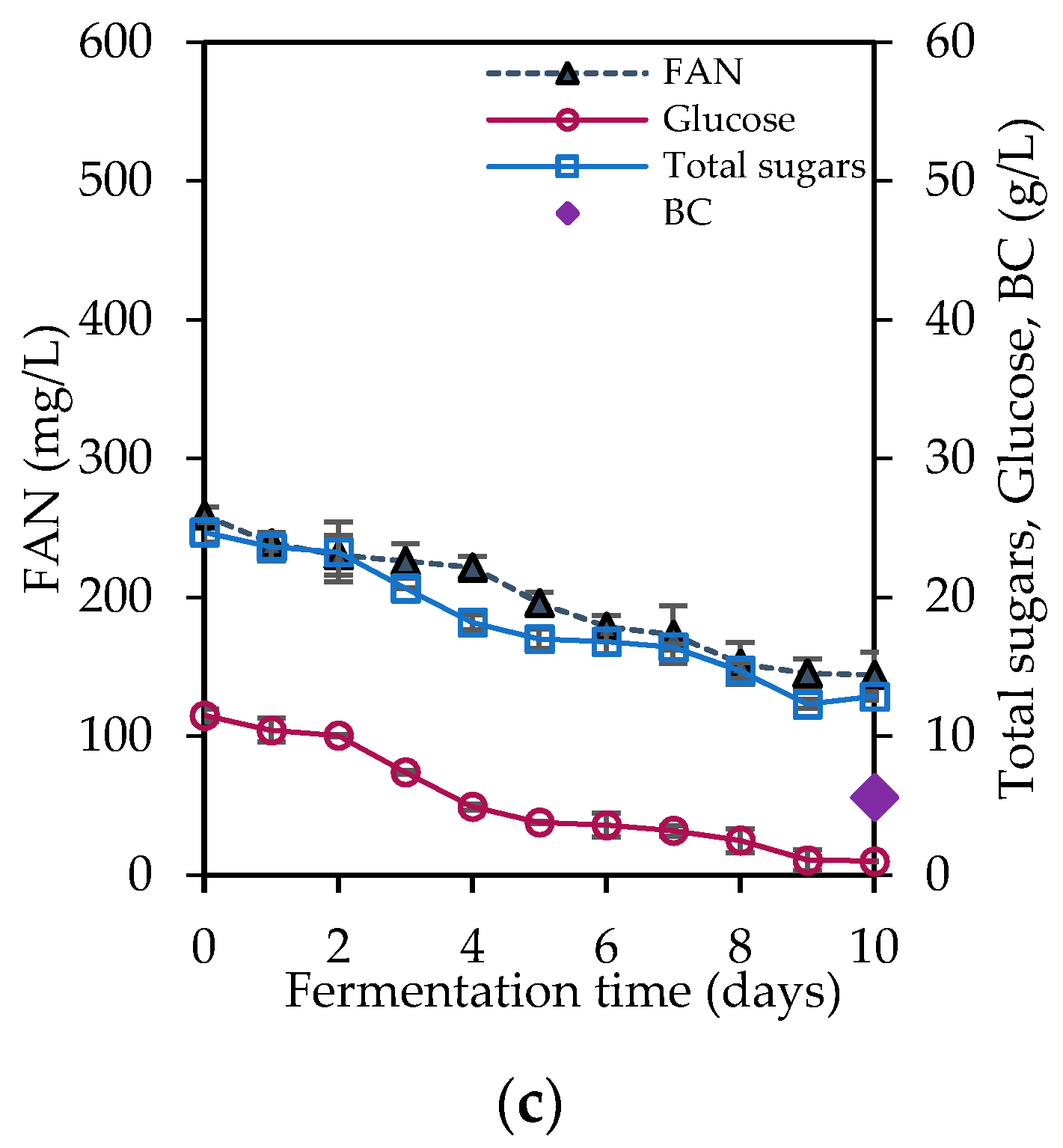
3.3. Bacterial Cellulose Production
3.4. Technological Consideration of the Study
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prazeres, A.R.; Carvalho, F.; Rivas, J. Cheese whey management: A review. J. Environ. Manag. 2012, 110, 48–68. [Google Scholar] [CrossRef]
- Yadav, J.S.S.; Yan, S.; Pilli, S.; Kumar, L.; Tyagi, R.D.; Surampalli, R.Y. Cheese whey: A potential resource to transform into bioprotein, functional/nutritional proteins and bioactive peptides. Biotechnol. Adv. 2015, 33, 756–774. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Nakagawa, S.; Kanawaku, R.; Kawamura, S. Ethanol production from cheese whey and expired milk by the brown rot fungus Neolentinus lepideus. Fermentation 2019, 5, 49. [Google Scholar] [CrossRef] [Green Version]
- Mehri, D.; Perendeci, N.A.; Goksungur, Y. Utilization of whey for red pigment production by Monascus purpureus in submerged fermentation. Fermentation 2021, 7, 75. [Google Scholar] [CrossRef]
- Marcus, J.F.; DeMarsh, T.A.; Alcaine, S.D. Upcycling of whey permeate through yeast- and mold-driven fermentations under anoxic and oxic conditions. Fermentation 2021, 7, 16. [Google Scholar] [CrossRef]
- Costa, S.; Summa, D.; Semeraro, B.; Zappaterra, F.; Rugiero, I.; Tamburini, E. Fermentation as a strategy for bio-transforming waste into resources: Lactic acid production from agri-food residues. Fermentation 2021, 7, 3. [Google Scholar] [CrossRef]
- Ghosh, B.C.; Prasad, L.N.; Saha, N.P. Enzymatic hydrolysis of whey and its analysis. J. Food Sci. Technol. 2017, 54, 1476–1483. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Xie, J.; Liu, J.; Ouyang, J. A novel thermostable β-galactosidase from Bacillus coagulans with excellent hydrolysis ability for lactose in whey. Int. J. Dairy Sci. 2019, 102, 9740–9748. [Google Scholar] [CrossRef] [PubMed]
- Saqib, S.; Akram, A.; Halim, S.A.; Tassaduq, R. Sources of β-galactosidase and its applications in food industry. 3 Biotech 2017, 7, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Xu, M.; Ji, D.; Agyei, D. Optimization of β-galactosidase production by batch cultures of Lactobacillus leichmannii 313 (ATCC 7830™). Fermentation 2020, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Martarello, R.D.; Cunha, L.; Cardoso, S.L.; de Freitas, M.M.; Silveira, D.; Fonseca-Bazzo, Y.M.; Homem-de-Mello, M.; Filho, E.X.F.; Magalhães, P.O. Optimization and partial purification of beta-galactosidase production by Aspergillus niger isolated from Brazilian soils using soybean residue. AMB Express 2019, 9, 81. [Google Scholar] [CrossRef] [Green Version]
- FDA. Partial List of Microorganisms and Microbial-Derived Ingredients Used in Foods. 2015. Available online: http://www.fda.gov/Food/IngredientsPackagingLabeling/GRAS/MicroorganismsMicrobialDerivedIngredients/default.htm (accessed on 20 June 2021).
- Cardoso, B.B.; Silvério, S.C.C.; Abrunhosa, L.; Teixeira, J.A.; Rodrigues, L.R. β-galactosidase from Aspergillus lacticoffeatus: A promising biocatalyst for the synthesis of novel prebiotics. Int. J. Food Microbiol. 2017, 257, 67–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidya, C.H.; Kumar, B.S.; Chinmayee, C.V.; Singh, S.A. Purification, characterization and specificity of a new GH family 35 galactosidase from Aspergillus awamori. Int. J. Biol. Macromol. 2020, 156, 885–895. [Google Scholar] [CrossRef]
- Koutinas, A.A.; Arifeen, N.; Wang, R.; Webb, C. Cereal-based biorefinery development: Integrated enzyme production for cereal flour hydrolysis. Biotechnol. Bioeng. 2007, 97, 61–72. [Google Scholar] [CrossRef]
- Tsakona, S.; Kopsahelis, N.; Chatzifragkou, A.; Papanikolaou, S.; Kookos, I.K.; Koutinas, A.A. Formulation of fermentation media from flour-rich waste streams for microbial lipid production by Lipomyces starkeyi. J. Biotechnol. 2014, 189, 36–45. [Google Scholar] [CrossRef]
- de Sousa Paredes, S.R.; de Barros, R.R.; Inoue, H.; Yano, S.; Bon, E.P. Production of xylanase, α-l-arabinofuranosidase, β-xylosidase, and β-glucosidase by Aspergillus awamori using the liquid stream from hot-compressed water treatment of sugarcane bagasse. Biomass Conv. Bioref. 2015, 5, 299–307. [Google Scholar] [CrossRef]
- Nishida, V.S.; de Oliveira, R.F.; Brugnari, T.; Correa, R.C.G.; Rosely, A.; Peralta, R.A.; Castoldi, R.; de Souza, C.G.; Bracht, A.; Peralta, R.M. Immobilization of Aspergillus awamori β-glucosidase on commercial gelatin: An inexpensive and efficient process. Int. J. Biol. Macromol. 2018, 111, 1206–1213. [Google Scholar] [CrossRef] [PubMed]
- Lappa, I.K.; Papadaki, A.; Kachrimanidou, V.; Terpou, A.; Koulougliotis, D.; Eriotou, E.; Kopsahelis, N. Cheese whey processing: Integrated biorefinery concepts and emerging food applications. Foods 2019, 8, 347. [Google Scholar] [CrossRef] [Green Version]
- Azeredo, H.M.C.; Barud, H.; Farinas, C.S.; Vasconcellos, V.M.; Claro, A.M. Bacterial cellulose as a raw material for food and food packaging applications. Front. Sustain. Food Syst. 2019, 3. [Google Scholar] [CrossRef] [Green Version]
- Fijałkowski, K.; Peitler, D.; Rakoczy, R.; Żywicka, A. Survival of probiotic lactic acid bacteria immobilized in different forms of bacterial cellulose in simulated gastric juices and bile salt solution. LWT Food Sci. Technol. 2016, 68, 322–328. [Google Scholar] [CrossRef]
- Bayazidi, P.; Almasi, H.; Aslm, A.K. Immobilization of lysozyme on bacterial cellulose nanofibers: Characteristics, antimicrobial activity and morphological properties. Int. J. Biol. Macromol. 2018, 107, 2544–2551. [Google Scholar] [CrossRef] [PubMed]
- Mohammadalinejhad, S.; Almasi, H.; Moradi, M. Immobilization of Echium amoenum anthocyanins into bacterial cellulose film: A novel colorimetric pH indicator for freshness/spoilage monitoring of shrimp. Food Control 2020, 113, 107169. [Google Scholar] [CrossRef]
- Cacicedo, M.L.; Castro, M.C.; Servetas, I.; Bosnea, L.; Boura, K.; Tsafrakidou, P.; Dima, A.; Terpou, A.; Koutinas, A.; Castro, G.R. Progress in bacterial cellulose matrices for biotechnological applications. Bioresour. Technol. 2016, 213, 172–180. [Google Scholar] [CrossRef]
- Revin, V.; Liyaskina, E.; Nazarkina, M.; Bogatyreva, A.; Shchankin, M. Cost-effective production of bacterial cellulose using acidic food industry by-products. Braz. J. Microbiol. 2018, 49, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Abol-Fotouh, D.; Hassan, M.A.; Shokry, H.; Roig, A.; Azab, M.S.; Kashyout, A.B. Bacterial nanocellulose from agro-industrial wastes: Low-cost and enhanced production by Komagataeibacter saccharivorans MD1. Sci. Rep. 2020, 10, 3491. [Google Scholar] [CrossRef] [Green Version]
- Son, H.J.; Kim, H.G.; Kim, K.K.; Kim, H.S.; Kim, Y.G.; Lee, S.J. Increased production of bacterial cellulose by Acetobacter sp. V6 in synthetic media under shaking culture conditions. Bioresour. Technol. 2003, 86, 215–219. [Google Scholar] [CrossRef]
- Paximada, P.; Tsouko, E.; Kopsahelis, N.; Koutinas, A.A.; Mandala, I. Bacterial cellulose as stabilizer of o/w emulsions. Food Hydrocoll. 2016, 53, 225–232. [Google Scholar] [CrossRef]
- Raol, G.G.; Raol, B.V.; Prajapati, V.S.; Bhavsar, N.H. Utilization of agro-industrial waste for β-galactosidase production under solid state fermentation using halotolerant Aspergillus tubingensis GR1 isolate. 3 Biotech 2015, 5, 411–421. [Google Scholar] [CrossRef] [Green Version]
- Nizamuddin, S.; Sridevi, A.; Narasimha, G. Production of β-galactosidase by Aspergillus oryzae in solid-state fermentation. Afr. J. Biotech. 2008, 7, 1096–1100. [Google Scholar]
- Żywicka, A.; Wenelska, K.; Junka, A.; Chodaczek, G.; Szymczyk, P.; Fijałkowski, K. Immobilization pattern of morphologically different microorganisms on bacterial cellulose membranes. World J. Microbiol. Biotechnol. 2019, 35, 11. [Google Scholar] [CrossRef]
- Tsouko, E.; Kourmentza, C.; Ladakis, D.; Kopsahelis, N.; Mandala, I.; Papanikolaou, S.; Paloukis, F.; Alves, V.; Koutinas, A. Bacterial cellulose production from industrial waste and by-product streams. Int. J. Mol. Sci. 2015, 16, 14832–14849. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, A.; Papapostolou, H.; Alexandri, M.; Kopsahelis, N.; Papanikolaou, S.; de Castro, A.M.; Freire, D.M.G.; Koutinas, A.A. Fumaric acid production using renewable resources from biodiesel and cane sugar production processes. Environ. Sci. Pollut. Res. 2018, 25, 35960–35970. [Google Scholar] [CrossRef] [PubMed]
- Kachrimanidou, V.; Kopsahelis, N.; Chatzifragkou, A.; Papanikolaou, S.; Yanniotis, S.; Kookos, I.; Koutinas, A.A. Utilisation of by-products from sunflower-based biodiesel production processes for the production of fermentation feedstock. Waste Biomass Valorization 2013, 4, 529–537. [Google Scholar] [CrossRef]
- Lie, S. The EBC-ninhydrin method for determination of free alpha amino nitrogen. J. Inst. Brew. 1973, 79, 37–41. [Google Scholar] [CrossRef]
- Hatzinikolaou, D.G.; Katsifas, E.; Mamma, D.; Karagouni, A.D.; Christakopoulos, P.; Kekos, D. Modeling of the simultaneous hydrolysis–ultrafiltration of whey permeate by a thermostable β-galactosidase from Aspergillus niger. Biochem. Eng. J. 2005, 24, 161–172. [Google Scholar] [CrossRef]
- Merali, Z.; Collins, S.R.A.; Elliston, A.; Wilson, D.R.; Käsper, A.; Waldron, K.W. Characterization of cell wall components of wheat bran following hydrothermal pretreatment and fractionation. Biotechnol. Biofuels 2015, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.R.; Madamwar, D. Xylanase production under solid-state fermentation and its characterization by an isolated strain of Aspergillus foetidus in India. World J. Microbiol. Biotechnol. 2005, 21, 233–243. [Google Scholar] [CrossRef]
- Wang, J.; Tavakoli, J.; Tang, Y. Bacterial cellulose production, properties and applications with different culture methods—A review. Carbohydr. Polym. 2019, 219, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Godoy, L.C.; Shaarani, S.M.; Melikoglu, M.; Koutinas, A.; Webb, C. Improving wheat flour hydrolysis by an enzyme mixture from solid state fungal fermentation. Enzyme Microb. Technol. 2009, 44, 223–228. [Google Scholar] [CrossRef]
- Ansari, S.A.; Husain, Q. Lactose hydrolysis from milk/whey in batch and continuous processes by concanavalin A-Celite 545 immobilized Aspergillus oryzae β galactosidase. Food Bioprod. Process. 2012, 90, 351–359. [Google Scholar] [CrossRef]
- Geiger, B.; Nguyen, H.M.; Wenig, S.; Nguyen, H.A.; Lorenz, C.; Kittl, R.; Mathiesen, G.; Eijsink, V.G.H.; Haltrich, D.; Nguyen, T.H. From by-product to valuable components: Efficient enzymatic conversion of lactose in whey using β-galactosidase from Streptococcus thermophilus. Biochem. Eng. J. 2016, 116, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampaio, F.C.; de Faria, J.T.; da Silva, M.F.; de Souza Oliveira, R.P.; Converti, A. Cheese whey permeate fermentation by Kluyveromyces lactis: A combined approach to wastewater treatment and bioethanol production. Environmen. Technol. 2019, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Silvério, S.C.; Macedo, E.A.; Teixeira, J.A.; Rodrigues, L.R. New β-galactosidase producers with potential for prebiotic synthesis. Bioresour. Technol. 2018, 250, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, P.; Mamo, G.; Karlsson, E.N. Potential and utilization of thermophiles and thermostable enzymes in biorefining. Microb. Cell Factories 2007, 6, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigoldi, F.; Donini, S.; Redaelli, A.; Parisini, E.; Gautieri, A. Review: Engineering of thermostable enzymes for industrial applications. APL Bioeng. 2018, 2, 011501. [Google Scholar] [CrossRef] [Green Version]
- Park, A.R.; Oh, D.K. Effects of galactose and glucose on the hydrolysis reaction of a thermostable β-galactosidase from Caldicellulosiruptor saccharolyticus. Appl. Microbiol. Biotechnol. 2010, 85, 1427–1435. [Google Scholar] [CrossRef]
- Gosling, A.; Stevens, G.W.; Barber, A.R.; Kentish, S.E.; Gras, S.L. Recent advances refining galactooligosaccharide production from lactose. Food Chem. 2010, 121, 307–318. [Google Scholar] [CrossRef]
- Kim, C.S.; Ji, E.S.; Oh, D.K. A new kinetic model of recombinant β-galactosidase from Kluyveromyces lactis for both hydrolysis and transgalactosylation reactions. Biochem. Biophys. Res. Commun. 2004, 316, 738–743. [Google Scholar] [CrossRef]
- Neri, D.F.; Balcão, V.M.; Carneiro-da-Cunha, M.G.; Carvalho, L.B.; Teixeira, J.A. Immobilization of β-galactosidase from Kluyveromyces lactis onto a polysiloxane–polyvinyl alcohol magnetic (mPOS–PVA) composite for lactose hydrolysis. Catal. Commun. 2008, 9, 2334–2339. [Google Scholar] [CrossRef] [Green Version]
- Ansari, S.A.; Husain, Q. Lactose hydrolysis by β galactosidase immobilized on concanavalin A-cellulose in batch and continuous mode. J. Mol. Catal. B Enzym Enzymatic 2010, 63, 68–74. [Google Scholar] [CrossRef]
- Dutra Rosolen, M.; Gennari, A.; Volpato, G.; Volken de Souza, C.F. Lactose hydrolysis in milk and dairy whey using microbial β-galactosidases. Enzyme Res. 2015, 806240. [Google Scholar] [CrossRef] [Green Version]
- Akgül, F.B.; Demirhan, E.; Özbek, B. A Modelling study on skimmed milk lactose hydrolysis and β-galactosidase stability using three reactor types. Int. J. Dairy Technol. 2012, 65, 217–231. [Google Scholar] [CrossRef]
- Horner, T.W.; Dunn, M.L.; Eggett, D.L.; Ogden, L.V. β-Galactosidase activity of commercial lactase samples in raw and pasteurized milk at refrigerated temperatures. J. Dairy Sci. 2011, 94, 3242–3249. [Google Scholar] [CrossRef] [Green Version]
- Semjonovs, P.; Ruklisha, M.; Paegle, L.; Saka, M.; Treimane, R.; Skute, M.; Rozenberga, L.; Vikele, L.; Sabovics, M.; Cleenwerck, I. Cellulose synthesis by Komagataeibacter rhaeticus strain P 1463 isolated from Kombucha. Appl. Microbiol. Biotechnol. 2017, 101, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Bekatorou, A.; Plioni, I.; Sparou, K.; Maroutsiou, R.; Tsafrakidou, P.; Petsi, T.; Kordouli, E. Bacterial cellulose production using the Corinthian currant finishing Side-Stream and Cheese Whey: Process Optimization and Textural Characterization. Foods 2019, 8, 193. [Google Scholar] [CrossRef] [Green Version]
- Salari, M.; Khiabani, M.S.; Mokarram, R.R.; Ghanbarzadeh, B.; Kafil, H.S. Preparation and characterization of cellulose nanocrystals from bacterial cellulose produced in sugar beet molasses and cheese whey media. Int. J. Biol. Macromol. 2019, 122, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Battad-Bernardo, E.; McCrindle, S.L.; Couperwhite, I.; Neilan, B.A. Insertion of an E. coli lacZ gene in Acetobacter xylinus for the production of cellulose in whey. FEMS Microbiol. Lett. 2004, 231, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Sharma, D.K.; Sandhu, P.P.; Jadaun, J.; Sangwan, R.S.; Yadav, S.K. Sustainable process for the production of cellulose by an Acetobacter pasteurianus RSV-4 (MTCC 25117) on whey medium. Cellulose 2021, 28, 103–116. [Google Scholar] [CrossRef]
- Carreira, P.; Mendes, J.A.; Trovatti, E.; Serafim, L.S.; Freire, C.S.; Silvestre, A.J.; Neto, C.P. Utilization of residues from agro-forest industries in the production of high value bacterial cellulose. Bioresour. Technol. 2011, 102, 7354–7360. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Flanagan, B.; Gidley, M.J. Characterization of cellulose production by a Gluconacetobacter xylinus strain from Kombucha. Curr. Microbiol. 2008, 57, 449. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Media | Initial Total Sugars (g/L) | Initial Glucose (g/L) | Residual Glucose (g/L) | Initial FAN (mg/L) | FAN Consumption (mg/L) | BC Production * (g/L) | BC Productivity (g/L/d) |
|---|---|---|---|---|---|---|---|
| Hydrolysate A | 45.04 ± 2.60 | 23.31 ± 0.77 | 9.40 ± 0.24 | 520.05 ± 0.34 | 224.79 ± 8.39 | 7.05 ± 0.14 A | 0.71 |
| Hydrolysate B | 45.80 ± 0.77 | 22.35 ± 0.46 | 8.93 ± 0.19 | 331.36 ± 12.96 | 132.64 ± 5.21 | 5.78 ± 0.35 A,B | 0.58 |
| Hydrolysate C | 24.68 ± 0.69 | 11.48 ± 0.48 | 1.00 + 0.08 | 259.25 ± 5.70 | 114.94 ± 4.73 | 5.59 ± 0.22 B | 0.56 |
| Cheese whey A | 50.00 ± 1.22 | 2.51 ± 0.23 | - | 56.00 ± 3.35 | 19.49 ± 2.78 | 0.58 ± 0.01 a | 0.06 |
| Cheese whey B | 24.45 ± 1.03 | 1.28 ± 0.10 | - | 22.98 ± 5.70 | 22.98 ± 0.00 | 0.71 ± 0.05 a,b | 0.07 |
| Cheese whey C | 24.29 ± 1.18 | 1.39 ± 0.14 | - | 250.00 ± 10.06 | 81.33 ± 4.65 | 1.07 ± 0.09 b | 0.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lappa, I.K.; Kachrimanidou, V.; Papadaki, A.; Stamatiou, A.; Ladakis, D.; Eriotou, E.; Kopsahelis, N. A Comprehensive Bioprocessing Approach to Foster Cheese Whey Valorization: On-Site β-Galactosidase Secretion for Lactose Hydrolysis and Sequential Bacterial Cellulose Production. Fermentation 2021, 7, 184. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030184
Lappa IK, Kachrimanidou V, Papadaki A, Stamatiou A, Ladakis D, Eriotou E, Kopsahelis N. A Comprehensive Bioprocessing Approach to Foster Cheese Whey Valorization: On-Site β-Galactosidase Secretion for Lactose Hydrolysis and Sequential Bacterial Cellulose Production. Fermentation. 2021; 7(3):184. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030184
Chicago/Turabian StyleLappa, Iliada K., Vasiliki Kachrimanidou, Aikaterini Papadaki, Anthi Stamatiou, Dimitrios Ladakis, Effimia Eriotou, and Nikolaos Kopsahelis. 2021. "A Comprehensive Bioprocessing Approach to Foster Cheese Whey Valorization: On-Site β-Galactosidase Secretion for Lactose Hydrolysis and Sequential Bacterial Cellulose Production" Fermentation 7, no. 3: 184. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030184