
Changes in Bioactive Compounds of Coffee Pulp through Fermentation-Based Biotransformation Using Lactobacillus plantarum TISTR 543 and Its Antioxidant Activities


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Microorganisms
2.2. Starter Culture Preparation
2.3. Fermentation
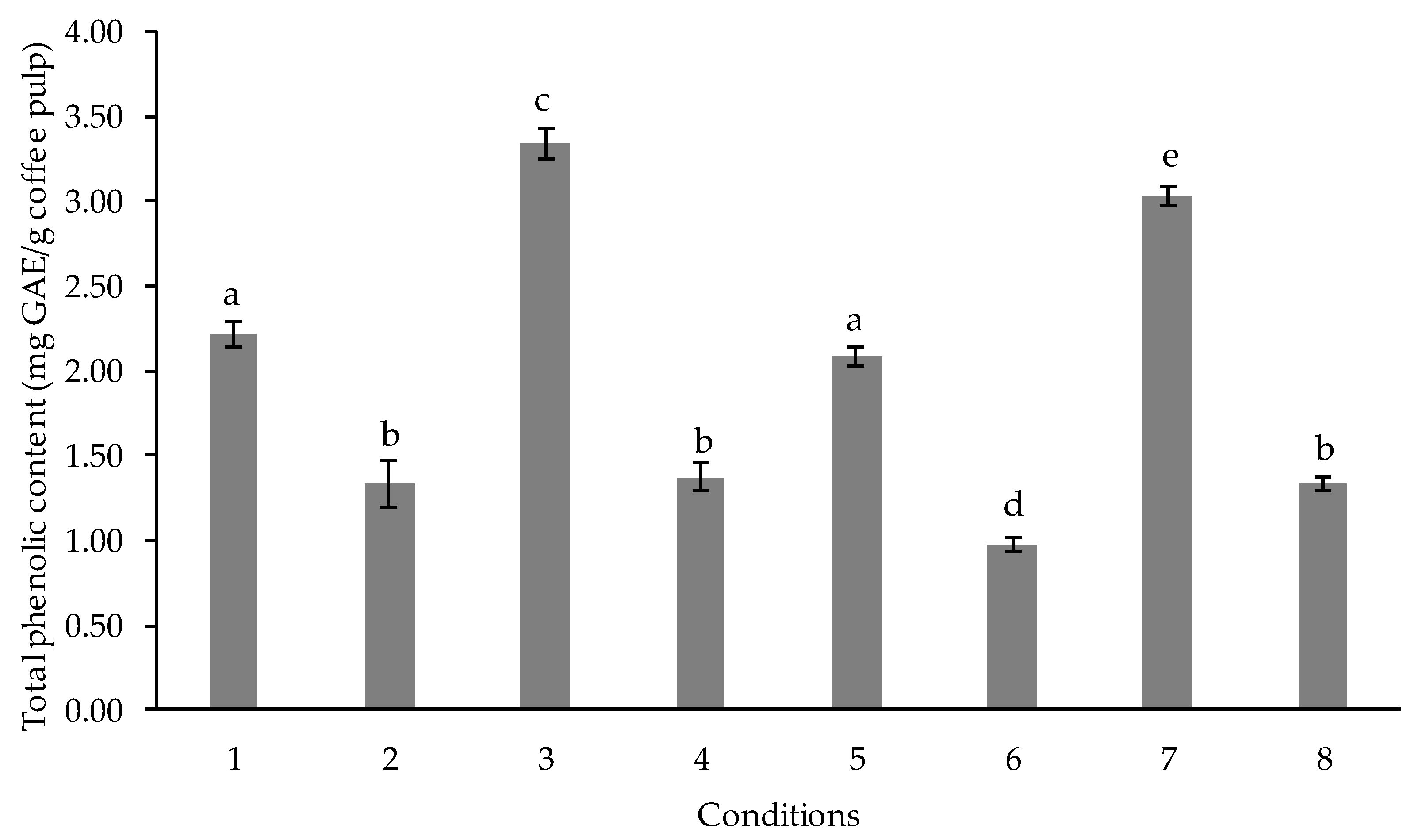
2.4. Determination of the Optimum Fermentation Time
2.5. Microbial Enumeration and pH Determination
2.6. Determination of Total Phenolic Content
2.7. Determination of Total Flavonoid Content
2.8. Determination of Tannin Content
2.9. Antioxidant Activity Assays
2.9.1. 1,1-diphenyl-2-picryl-hydrazil (DPPH) Radical Scavenging Assay
2.9.2. 2,2′-azino-bis-3-ethylbenzthiazoline-6-sulphonic Acid (ABTS) Radical Scavenging Assay
2.9.3. Ferric Reducing Antioxidant Power Assay (FRAP)
2.10. Determination of Bioactive Compounds by LC-QQQ
2.11. Statistical Analysis
3. Results and Discussion
3.1. Fermentation
3.2. Determination of the Optimum Fermentation Time
3.2.1. pH Measurement and Microbial Enumeration
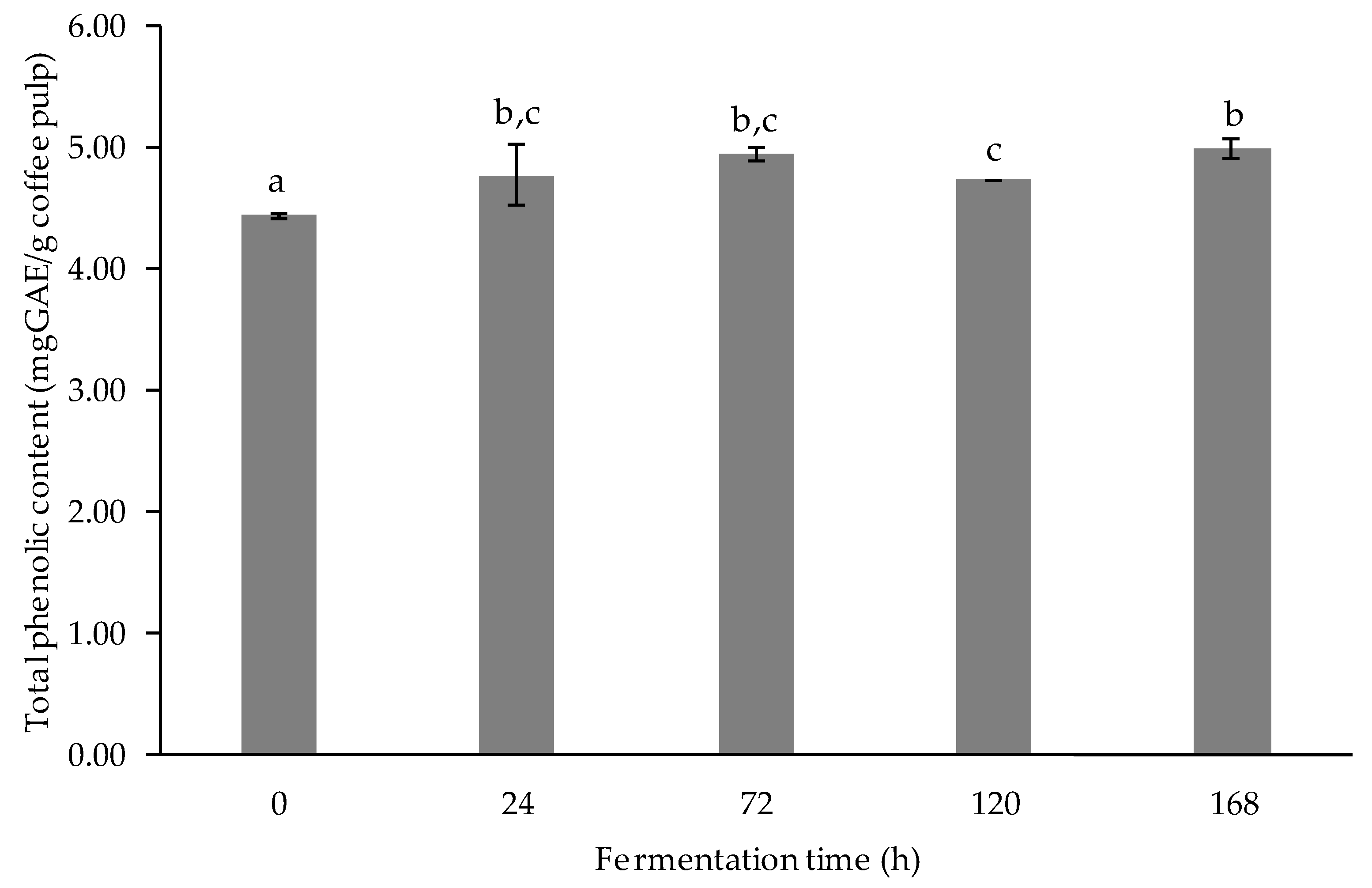
3.2.2. Determination of Total Phenolic Content
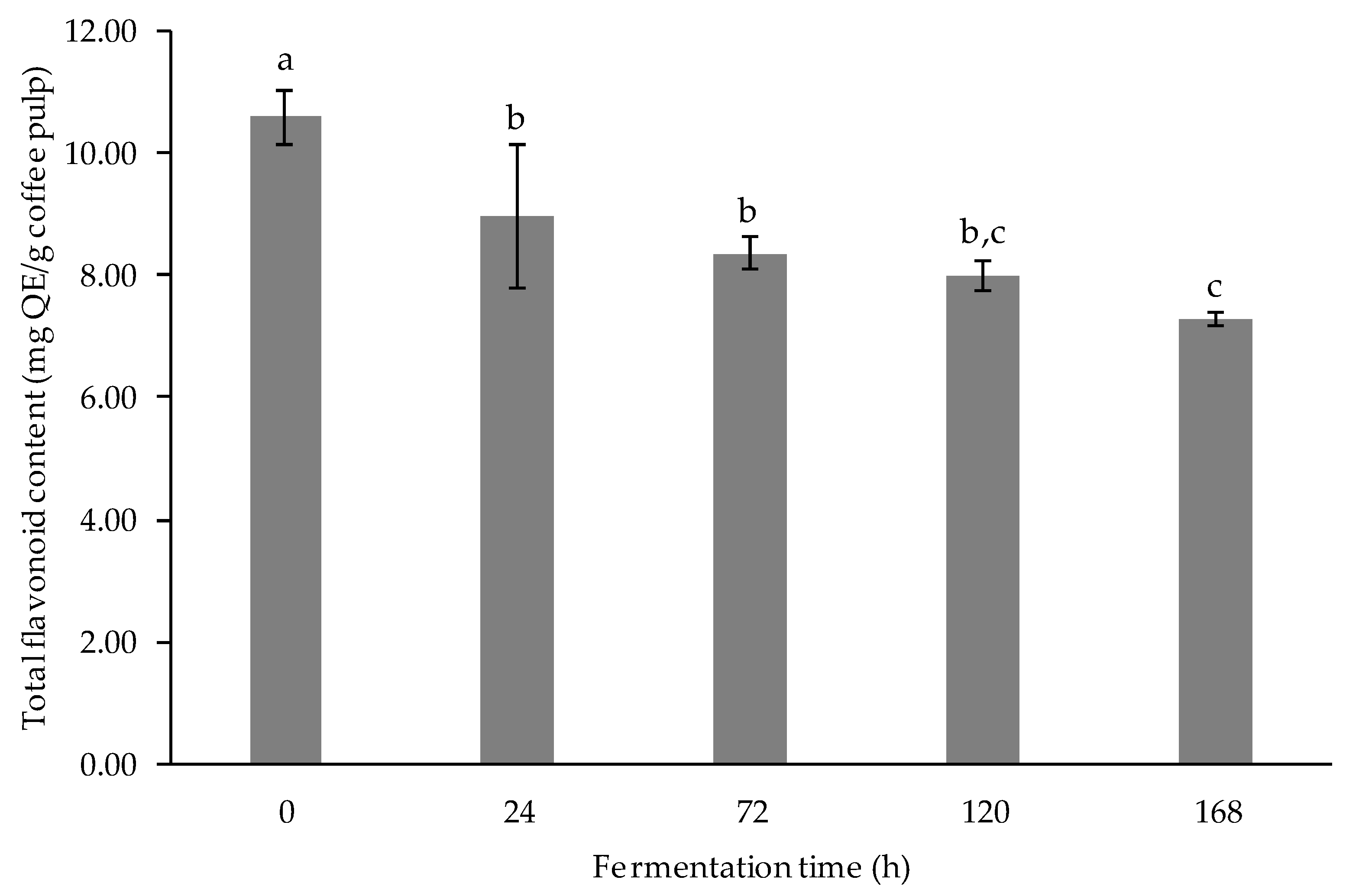
3.2.3. Determination of Total Flavonoid Content
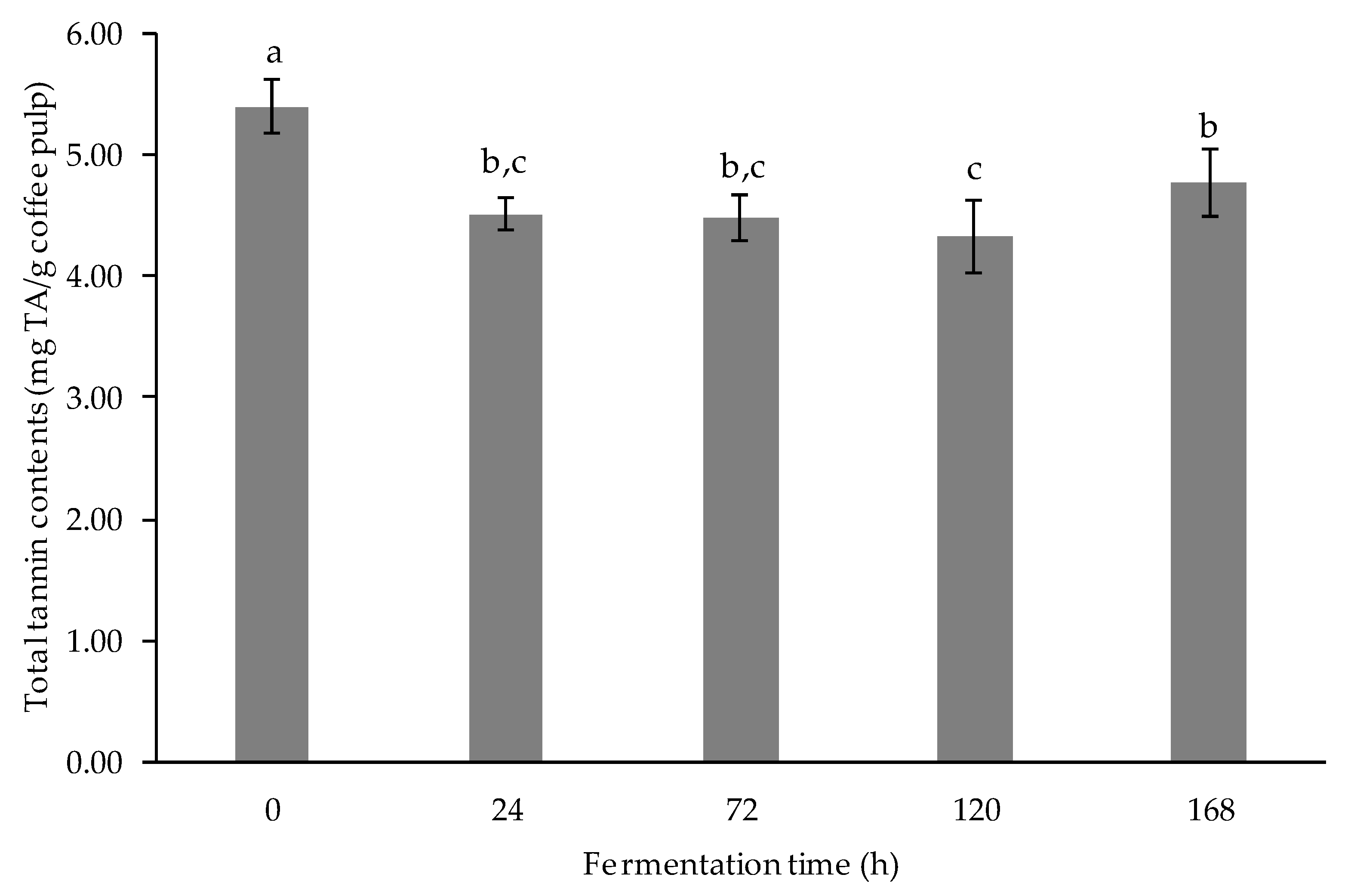
3.2.4. Determination of Tannin Content
3.2.5. Antioxidant Activities
3.2.6. Determination of Bioactive Compounds by LC-QQQ
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adam, A.; Hakim, M.N.; Oktaviani, L.; Inderaja, B.M.; Manurung, R.; Putra, R.-a.E.; Abduh, M.Y. Techno-economic evaluation for integrated cultivation of coffee and stingless bees in West Java, Indonesia: Integrated cultivation of coffee and stingless bees. Biol. Nat. Resour. Environ. J. 2020, 3, 28–36. [Google Scholar] [CrossRef]
- Gamonal, L.E.; Vallejos-Torres, G.; López, L.A. Sensory analysis of four cultivars of coffee (Coffea arabica L.), grown at different altitudes in the San Martin region—Peru. Cienc. Rural. 2017, 47. [Google Scholar] [CrossRef] [Green Version]
- Murthy, P.S.; Madhava Naidu, M. Sustainable management of coffee industry by-products and value addition—A review. Resour. Conserv. Recycl. 2012, 66, 45–58. [Google Scholar] [CrossRef]
- Bakker, R.R.C. Availability of Lignocellulosic Feedstocks for Lactic Acid Production—Feedstock Availability, Lactic Acid Production Potential and Selection Criteria; Wageningen UR Food & Biobased Research: Wageningen, The Netherlands, 2013. [Google Scholar]
- Menezes, E.G.; do Carmo, J.R.; Alves, J.G.; Menezes, A.G.; Guimaraes, I.C.; Queiroz, F.; Pimenta, C.J. Optimization of alkaline pretreatment of coffee pulp for production of bioethanol. Biotechnol. Prog. 2014, 30, 451–462. [Google Scholar] [CrossRef]
- Corro, G.; Paniagua, L.; Pal, U.; Bañuelos, F.; Rosas, M. Generation of biogas from coffee-pulp and cow-dung co-digestion: Infrared studies of postcombustion emissions. Energy. Convers. Manag. 2013, 74, 471–481. [Google Scholar] [CrossRef]
- Pandey, A.; Soccol, C.R.; Nigam, P.; Brand, D.; Mohan, R.; Roussos, S. Biotechnological potential of coffee pulp and coffee husk for bioprocesses. Biochem. Eng. J. 2000, 6, 153–162. [Google Scholar] [CrossRef]
- Heeger, A.; Kosińska-Cagnazzo, A.; Cantergiani, E.; Andlauer, W. Bioactives of coffee cherry pulp and its utilisation for production of Cascara beverage. Food Chem. 2017, 221, 969–975. [Google Scholar] [CrossRef]
- Bonilla-Hermosa, V.A.; Duarte, W.F.; Schwan, R.F. Utilization of coffee by-products obtained from semi-washed process for production of value-added compounds. Bioresour. Technol. 2014, 166, 142–150. [Google Scholar] [CrossRef]
- Rodriguez-Duran, L.V.; Ramirez-Coronel, M.A.; Aranda-Delgado, E.; Nampoothiri, K.M.; Favela-Torres, E.; Aguilar, C.N.; Saucedo-Castaneda, G. Soluble and bound hydroxycinnamates in coffee pulp (Coffea arabica) from seven cultivars at three ripening stages. J. Agric. Food Chem. 2014, 62, 7869–7876. [Google Scholar] [CrossRef]
- Ramirez-Coronel, M.A.; Marnet, N.; Kolli, V.S.; Roussos, S.; Guyot, S.; Augur, C. Characterization and estimation of proanthocyanidins and other phenolics in coffee pulp (Coffea arabica) by thiolysis-high-performance liquid chromatography. J. Agric. Food Chem. 2004, 52, 1344–1349. [Google Scholar] [CrossRef]
- Matsui, T.; Ueda, T.; Oki, T.; Sugita, K.; Terahara, N.; Matsumoto, K. α-Glucosidase inhibitory action of natural acylated anthocyanins. 1. Survey of natural pigments with potent inhibitory activity. J. Agric. Food Chem. 2001, 49, 1948–1951. [Google Scholar] [CrossRef]
- Collins, A.M.; Kennedy, M.J. Biotransformations and bioconversions in New Zealand: Past endeavours and future potential. Australas. Biotechnol. 1999, 9, 86–94. [Google Scholar]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, R.; de las Rivas, B.; López de Felipe, F.; Reverón, I.; Santamaría, L.; Esteban-Torres, M.; Curiel, J.A.; Rodríguez, H.; Landete, J.M. Biotransformation of phenolics by Lactobacillus plantarum in fermented foods. In Fermented Foods in Health and Disease Prevention; Frias, J., Martinez-Villaluenga, C., Peñas, E., Eds.; Academic Press: Boston, MA, USA, 2017; Chapter 4; pp. 63–83. [Google Scholar]
- Rodríguez, H.; Landete, J.M.; Rivas, B.d.l.; Muñoz, R. Metabolism of food phenolic acids by Lactobacillus plantarum CECT 748T. Food Chem. 2008, 107, 1393–1398. [Google Scholar] [CrossRef] [Green Version]
- Darby, T.M.; Jones, R.M. Beneficial influences of Lactobacillus plantarum on human health and disease. In The Microbiota in Gastrointestinal Pathophysiology; Floch, M.H., Ringel, Y., Allan, W., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 109–117. [Google Scholar]
- Saelee, M.; Sivamaruthi, B.S.; Sirilun, S.; Sirithunyalug, J.; Peerajan, S.; Chaiyasut, C. The influence of pasteurization and starter culture on methanol content and bio-profile of fermented Morinda citrifolia linn. (noni) fruit juice. Food Sci. Technol. 2020, 40, 621–628. [Google Scholar] [CrossRef] [Green Version]
- Haile, M.; Kang, W.H. Antioxidant activity, total polyphenol, flavonoid and tannin contents of fermented green coffee beans with selected yeasts. Fermentation 2019, 5, 29. [Google Scholar] [CrossRef] [Green Version]
- Peerajan, S.; Chaiyasut, C.; Sirilun, S.; Chaiyasut, K.; Kesika, P.; Sivamaruthi, B.S. Enrichment of nutritional value of Phyllanthus emblica fruit juice using the probiotic bacterium, Lactobacillus paracasei HII01 mediated fermentation. Food Sci. Technol. 2016, 36, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Saftić, L.; Peršurić, Ž.; Kraljević Pavelić, S. LC-QQQ and LC-QTOF MS methods for comprehensive detection of potential allergens in various propolis extracts. Eur. Food Res. Technol. 2019, 245, 1981–1995. [Google Scholar] [CrossRef]
- Pandis, N.; Walsh, T.; Polychronopoulou, A.; Katsaros, C.; Eliades, T. Factorial designs: An overview with applications to orthodontic clinical trials. Eur. J. Orthod. 2013, 36, 314–320. [Google Scholar] [CrossRef] [Green Version]
- Filannino, P.; Bai, Y.; Di Cagno, R.; Gobbetti, M.; Gänzle, M.G. Metabolism of phenolic compounds by Lactobacillus spp. during fermentation of cherry juice and broccoli puree. Food Microbiol. 2015, 46, 272–279. [Google Scholar] [CrossRef]
- Starzyńska-Janiszewska, A.; Stodolak, B. Effect of inoculated lactic acid fermentation on antinutritional and antiradical properties of grass pea (Lathyrus Sativus ‘Krab’) flour. Pol. J. Food Nutr. Sci. 2011, 61, 245–249. [Google Scholar] [CrossRef] [Green Version]
- Sandhya, M.V.S.; Yallappa, B.S.; Varadaraj, M.C.; Puranaik, J.; Rao, L.J.; Janardhan, P.; Murthy, P.S. Inoculum of the starter consortia and interactive metabolic process in enhancing quality of cocoa bean (Theobroma cacao) fermentation. LWT Food Sci. Technol. 2016, 65, 731–738. [Google Scholar] [CrossRef]
- Sripo, K.; Phianmongkhol, A.; Wirjantoro, T.I. Effect of inoculum levels and final pH values on the antioxidant properties of black glutinous rice solution fermented by Lactobacillus bulgaricus. Int. Food Res. J. 2016, 23, 2207–2213. [Google Scholar]
- Zubaidah, E.; Arum, S.M.; Widyaningsih, D.T.; Rahayu, P.A. Sauerkraut with the addition of Lactobacillus casei: Effects of salt and sugar concentrations on fermentation and antioxidant activity. Curr. Nutr. Food Sci. 2020, 16, 1265–1269. [Google Scholar] [CrossRef]
- Dewi, E.; Septiningrum, S.; Rianingsih, L.; Riyadi, P. Optimization of carbon source and concentration for Lactobacillus acidophilus growth, phenolic production and antioxidant activity in fermented seaweed extract. Int. J. Emerg. Technol. 2020, 11, 495–500. [Google Scholar]
- Kaprasob, R.; Kerdchoechuen, O.; Laohakunjit, N.; Thumthanaruk, B.; Shetty, K. Changes in physico-chemical, astringency, volatile compounds and antioxidant activity of fresh and concentrated cashew apple juice fermented with Lactobacillus plantarum. J. Food Sci. Technol. 2018, 55, 3979–3990. [Google Scholar] [CrossRef] [PubMed]
- Thongruck, K.; Saelao, S.; Sumpavapo, P.; Benjaku, S.; Maneerat, S. Monitoring of changes in lactic acid bacteria during production of Thai traditional fermented shrimp (Kung-Som) by culturing method and PCR-DGGE technique. Songklanakarin J. Sci. Technol. 2017, 39, 41–47. [Google Scholar]
- Ernanin Dyah, W.; Nur Candra Eka, S.; Jean Patricia, C. Effect of lactic acid fermentation on total phenolic content and antioxidant activity of fig fruit juice (Ficus carica). In Proceedings of the Health Science International Conference (HSIC 2017), Faculty of Health Science, University of Muhammadiyah Malang, Kota Malang, Indonesia, 4–5 October 2017; pp. 282–289. [Google Scholar]
- Escudero-Lopez, B.; Cerrillo, I.; Herrero-Martin, G.; Hornero-Mendez, D.; Gil-Izquierdo, A.; Medina, S.; Ferreres, F.; Berna, G.; Martin, F.; Fernandez-Pachon, M.S. Fermented orange juice: Source of higher carotenoid and flavanone contents. J. Agric. Food Chem. 2013, 61, 8773–8782. [Google Scholar] [CrossRef]
- Hernandez, T.; Estrella, I.; Perez-Gordo, M.; Alegria, E.G.; Tenorio, C.; Ruiz-Larrrea, F.; Moreno-Arribas, M.V. Contribution of malolactic fermentation by Oenococcus oeni and Lactobacillus plantarum to the changes in the nonanthocyanin polyphenolic composition of red wine. J. Agric. Food Chem. 2007, 55, 5260–5266. [Google Scholar] [CrossRef]
- Zubaidah, E.; Susanti, I.; Yuwono, S.; Rahayu, A.P.; Srianta, I.; Blanc, P. Effect of Lactobacillus plantarum and Leuconostoc mesenteroides starter cultures in lower salt concentration fermentation on the sauerkraut quality. Food Res. 2020, 4, 1038–1044. [Google Scholar] [CrossRef]
- Wijayanti, E.D.; Setiawan, N.C.E. The effect of lactic acid fermentation on fig (Ficus carica) fruit flavonoid. Berk. Penelit. Hayati 2017, 23. [Google Scholar] [CrossRef]
- Li, Z.; Teng, J.; Lyu, Y.; Hu, X.; Zhao, Y.; Wang, M. Enhanced antioxidant activity for apple juice fermented with Lactobacillus plantarum ATCC14917. Molecules 2018, 24, 51. [Google Scholar] [CrossRef] [Green Version]
- Adetuyi, F.O.; Ibrahim, T.A. Effect of fermentation time on the phenolic, flavonoid and vitamin C contents and antioxidant activities of okra (Abelmoschus esculentus) seeds. Niger. Food J. 2014, 32, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.F.; Cao, H.; Ma, Y.L.; Zhang, C.; Ma, F.; Wang, C.X.; Ni, X.L.; Lee, W.J.; Wei, Z.J. Effect of lactic acid bacteria fermentation on tannins removal in Xuan Mugua fruits. Food Chem. 2019, 274, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; Rodriguez, H.; De las Rivas, B.; Munoz, R. High-added-value antioxidants obtained from the degradation of wine phenolics by Lactobacillus plantarum. J. Food Prot. 2007, 70, 2670–2675. [Google Scholar] [CrossRef]
- Amorati, R.; Valgimigli, L. Advantages and limitations of common testing methods for antioxidants. Free Radic. Res. 2015, 49, 633–649. [Google Scholar] [CrossRef]
- Sawangwan, T.; Porncharoennop, C.; Nimraksa, H. Antioxidant compounds from rice bran fermentation by lactic acid bacteria. AIMS Agric. Food. 2021, 6, 578–587. [Google Scholar] [CrossRef]
- Ryu, J.Y.; Kang, H.R.; Cho, S.K. Changes over the fermentation period in phenolic compounds and antioxidant and anticancer activities of blueberries fermented by Lactobacillus plantarum. J. Food Sci. 2019, 84, 2347–2356. [Google Scholar] [CrossRef]
- Adebo, O.A.; Gabriela Medina-Meza, I. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [Green Version]
- Ashihara, H. Metabolism of alkaloids in coffee plants. Braz. J. Plant Physiol. 2006, 18, 1–8. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Code | Unit | Coded Levels | |
|---|---|---|---|---|
| (−1) | (+1) | |||
| Substrate ratio of substrate per 10 parts of water | X1 | − | 3 | 7 |
| Sugar ratio of sugar per 10 parts of water | X2 | − | 1 | 3 |
| Starter | X3 | % | 5 | 15 |
| Standard No. | Run No. | X1: Substrate Ratio of Substrate per 10 Parts of Water | X2: Sugar Ratio of Sugar per 10 Parts of Water | X3: Starter (%) |
|---|---|---|---|---|
| 1 | 4 | −1 | −1 | −1 |
| 2 | 1 | 1 | −1 | −1 |
| 3 | 6 | −1 | 1 | −1 |
| 4 | 2 | 1 | 1 | −1 |
| 5 | 3 | −1 | −1 | 1 |
| 6 | 7 | 1 | −1 | 1 |
| 7 | 8 | −1 | 1 | 1 |
| 8 | 5 | 1 | 1 | 1 |
| Bioactive Compounds | Retention Time (RT, min) | Ionization Mode of the Analytes | Precursor Ion (m/z, [M − H]−) | Product Ion (m/z) |
|---|---|---|---|---|
| Phenolic acids | ||||
| Protocatechuic acid | 2.627 | Negative | 153.25 | 109.05 |
| Chlorogenic acid | 3.710 | Negative | 352.95 | 191.05 |
| Neochlorogenic acid | 2.547 | Negative | 352.95 | 191.05 |
| Caffeic acid | 4.566 | Negative | 178.80 | 135.20 |
| Gallic acid | 1.639 | Negative | 169.10 | 125.20 |
| o-Coumaric acid | 5.864 | Negative | 163.00 | 119.00 |
| p-Coumaric acid | 5.529 | Negative | 162.85 | 119.00 |
| p-hydroxybenzoic acid | 6.263 | Negative | 137.05 | 92.95 |
| Flavonoids | ||||
| Quercetin-3-O-rutinoside | 5.487 | Negative | 609.05 | 300.20, 271.10 |
| 4′,5,7-trihydroxyflavone | 6.279 | Negative | 269.10 | 117.00 |
| Alkaloids | ||||
| Caffeine | 3.617 | Positive | 195.00 | 138.15, 110.10 |
| Trigonelline | 0.917 | Positive | 138.10 | 92.10, 94.05, 78.05 |
| Theobromine | 1.763 | Positive | 181.10 | 163.10, 138.10, 110.05 |
| Variables | Type III Sum of Squares | df | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| Corrected Model | 15.682 | 7 | 2.240 | 358.897 | <0.001 * |
| Intercept | 1.959 | 1 | 92.123 | 14,758.693 | <0.001 * |
| A: Substrate | −0.707 | 1 | 11.996 | 1921.813 | <0.001 * |
| B: Sugar | 0.308 | 1 | 2.273 | 364.133 | <0.001 * |
| C: Starter | −0.105 | 1 | 0.265 | 42.456 | <0.001 * |
| AB | −0.209 | 1 | 1.046 | 167.503 | <0.001 * |
| AC | 0.007 | 1 | 0.001 | 0.209 | 0.654 |
| BC | 0.018 | 1 | 0.007 | 1.190 | 0.291 |
| ABC | 0.062 | 1 | 0.093 | 14.976 | 0.001 * |
| Error | 0.100 | 16 | 0.006 | ||
| Total | 107.905 | 24 | |||
| Corrected Total | 15.781 | 23 | |||
| R-Squared | 0.994 | ||||
| Adj. R-Squared | 0.991 |
| Fermentation Time (h) | DPPH (mg TE/g Sample) | ABTS (mg TE/g Sample) | FRAP (mg FeSO4/g Sample) |
|---|---|---|---|
| 0 | 3.51 ± 0.68 a | 3.99 ± 0.10 a | 6.89 ± 0.45 a |
| 24 | 4.57 ± 0.25 b | 4.91 ± 0.08 b | 9.09 ± 0.13 b |
| 72 | 4.35 ± 0.36 b,c | 4.91 ± 0.40 b | 8.64 ± 0.14 b,c |
| 120 | 4.16 ± 0.33 a,b,c | 3.79 ± 0.14 a,c | 7.72 ± 0.31 d |
| 168 | 3.65 ± 0.10 a,c | 3.59 ± 0.08 c | 7.88 ± 0.78 c,d |
| Compounds | Fermented Coffee Pulp (g/mL) | % Change from the Initial | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 h | 24 h | 72 h | 120 h | 168 h | 24 h | 72 h | 120 h | 168 h | |
| Phenolic acids | |||||||||
| Protocatechuic acid | 1.17 ± 0.05 a | 1.34 ± 0.04 b | 1.12 ± 0.07 a,c | 1.01 ± 0.09 c | 1.00 ± 0.12 c | 17.68% | −4.98% | −15.34% | −16.59% |
| Chlorogenic acid | 12.81 ± 0.22 a | 12.73 ± 0.03 a | 11.71 ± 0.04 b | 11.29 ± 0.12 c | 10.70 ± 0.04 d | −0.58% | −8.49% | −11.73% | −6.31% |
| Neochlorogenic acid | N.A. | N.A. | N.A. | N.A. | N.A. | −9.21% | −12.51% | −10.12% | −12.56% |
| Caffeic acid | N.D. | N.D. | N.D. | N.D. | N.D. | 25.25% | −32.19% | −42.50% | −4.60% |
| Gallic acid | N.D. | N.D. | N.D. | N.D. | N.D. | −2.75% | 40.81% | v22.12% | 29.42% |
| o-Coumaric acid | N.D. | N.D. | N.D. | N.D. | N.D. | 17.67% | 77.16% | 18.11% | 26.82% |
| p-Coumaric acid | N.D. | N.D. | N.D. | N.D. | N.D. | 47.14% | −28.45% | −3.32% | 2.97% |
| p-hydroxybenzoic acid | N.A. | N.A. | N.A. | N.A. | N.A. | 54.68% | −82.24% | −28.77% | −26.74% |
| Flavonoids | |||||||||
| Quercetin-3-O-rutinoside | N.A. | N.A. | N.A. | N.A. | N.A. | −3.57% | −27.86% | −16.52% | −16.29% |
| 4′,5,7-trihydroxyflavone | N.A. | N.A. | N.A. | N.A. | N.A. | −24.23% | 4.87% | 36.43% | 61.77% |
| Alkaloids | |||||||||
| Caffeine | 96.50 ± 0.50 ns | 94.50 ± 1.32 ns | 95.67 ± 1.61 ns | 95.33 ± 0.29 ns | 94.17 ± 0.29 ns | −2.22% | −1.14% | −1.37% | −2.49% |
| Trigonelline | 65.00 ± 1.73 a | 65.33 ± 0.58 a | 62.67 ± 2.31 a,c | 58.67 ± 1.53 b | 60.00 ± 1.00 b,c | −4.69% | −7.39% | −8.95% | −7.38% |
| Theobromine | 1.52 ± 0.15 ns | 1.50 ± 0.09 ns | 1.29 ± 0.09 ns | 1.39 ± 0.04 ns | 1.38 ± 0.13 ns | −1.34% | −13.97% | −8.02% | −8.25% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Myo, H.; Nantarat, N.; Khat-Udomkiri, N. Changes in Bioactive Compounds of Coffee Pulp through Fermentation-Based Biotransformation Using Lactobacillus plantarum TISTR 543 and Its Antioxidant Activities. Fermentation 2021, 7, 292. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7040292
Myo H, Nantarat N, Khat-Udomkiri N. Changes in Bioactive Compounds of Coffee Pulp through Fermentation-Based Biotransformation Using Lactobacillus plantarum TISTR 543 and Its Antioxidant Activities. Fermentation. 2021; 7(4):292. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7040292
Chicago/Turabian StyleMyo, Hla, Nara Nantarat, and Nuntawat Khat-Udomkiri. 2021. "Changes in Bioactive Compounds of Coffee Pulp through Fermentation-Based Biotransformation Using Lactobacillus plantarum TISTR 543 and Its Antioxidant Activities" Fermentation 7, no. 4: 292. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7040292