
Potential of a Techno-Functional Sourdough and Its Application in Sugar-Reduced Soft Buns

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Sourdough Fermentation
2.2.1. Preparation of Sourdough
2.2.2. LAB Development and Acidification of the Sourdough
2.2.3. Quantification of Fructose, Mannitol, and Organic Acids in Sourdoughs
2.3. Dough and Bun Production
2.4. Bun Characteristics
2.5. Determination of Bun Volume, Bun Height, and Slice Area
2.5.1. Crust Texture, Crumb Firmness, and Resilience of Buns
2.5.2. Crust and Crumb Color
2.5.3. Water Activity and Shelf Life of Buns
2.6. Statistical Analysis
3. Results and Discussion
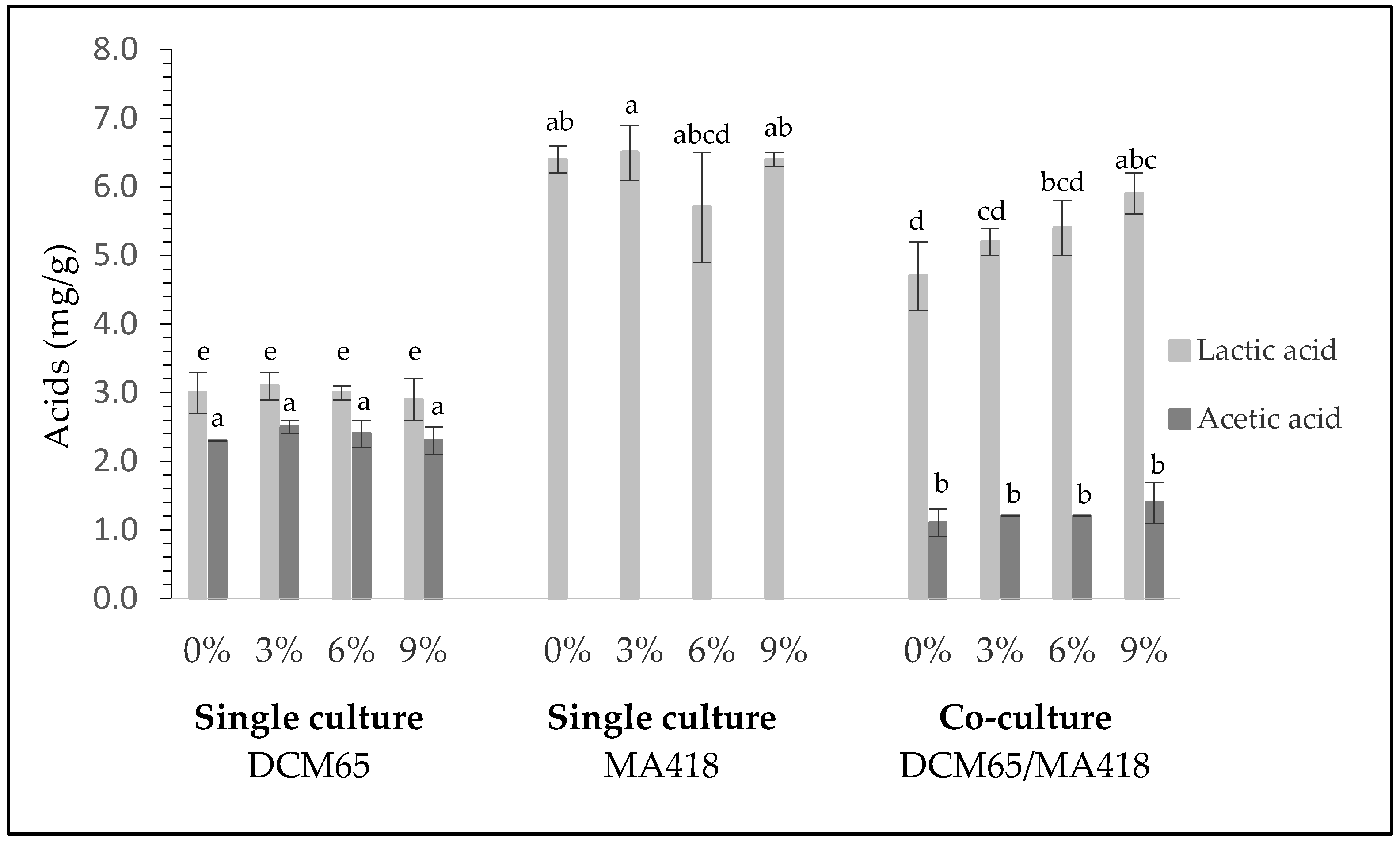
3.1. Characterization of Sourdoughs Inoculated with Single- or Co-Culture
3.2. Physical Properties of the Buns
3.2.1. Crust and Crumb Color of Buns
3.2.2. Water Activity and Shelf Life of Buns
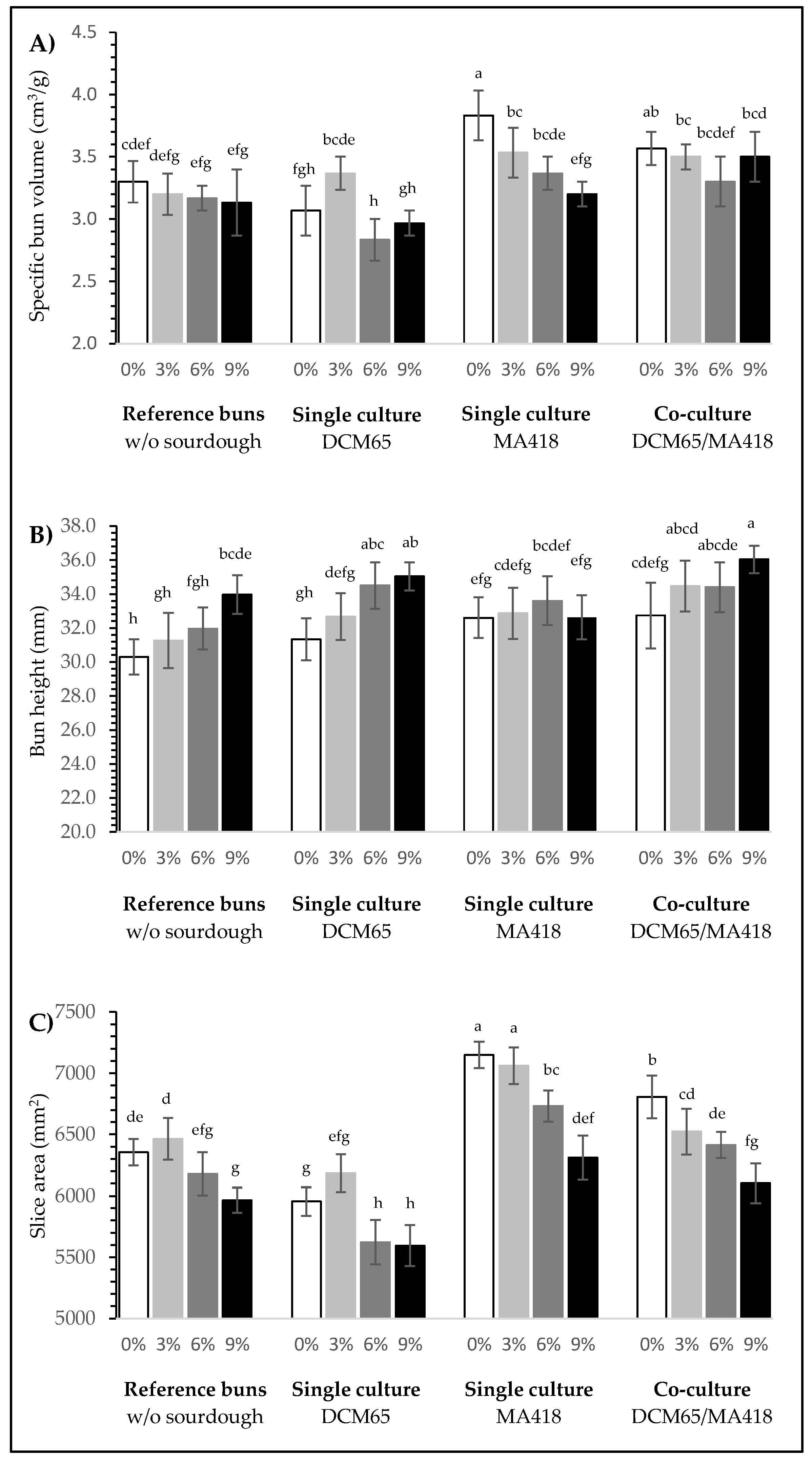
3.2.3. Bun Volume and Shape
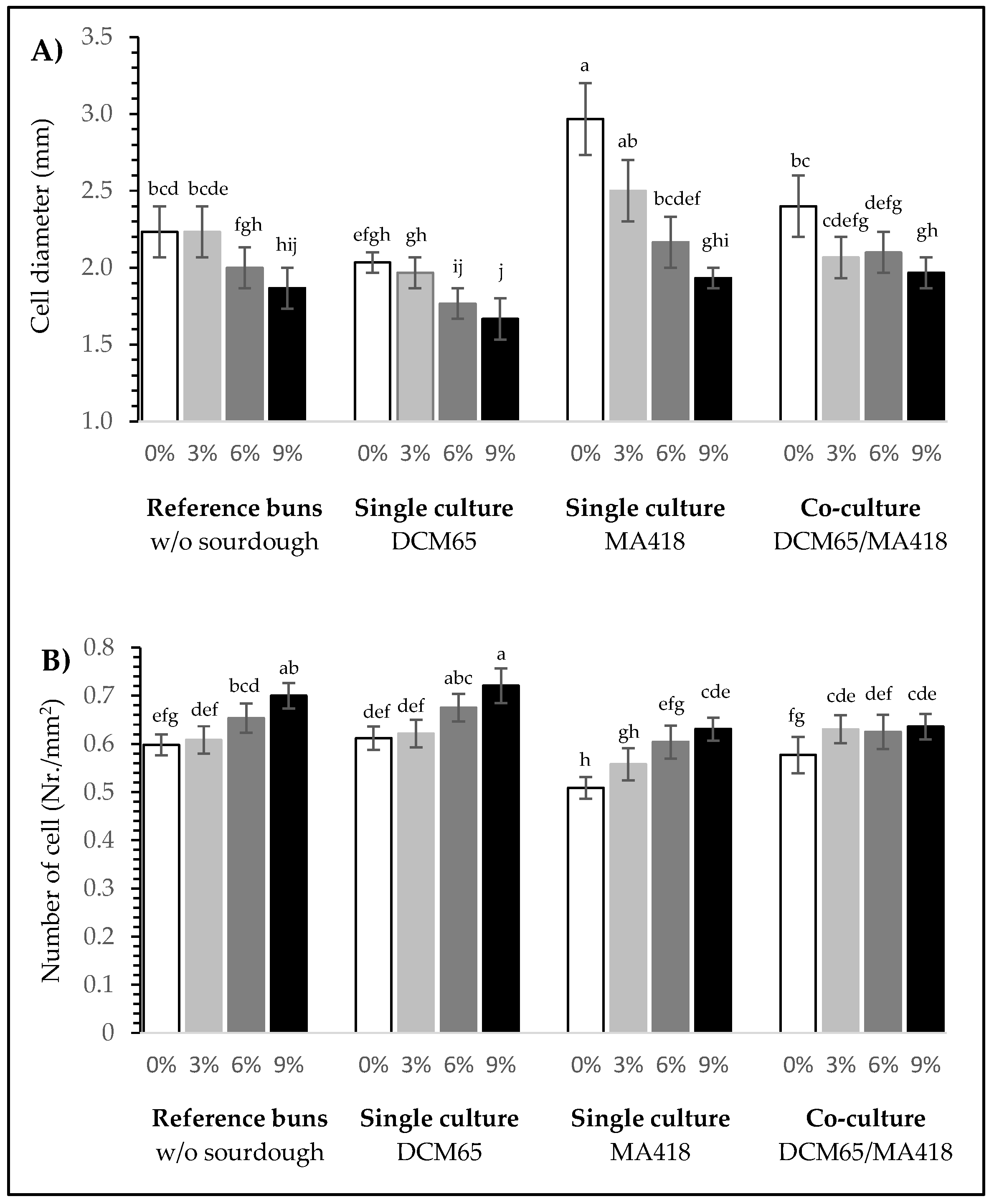
3.2.4. Pore Structure
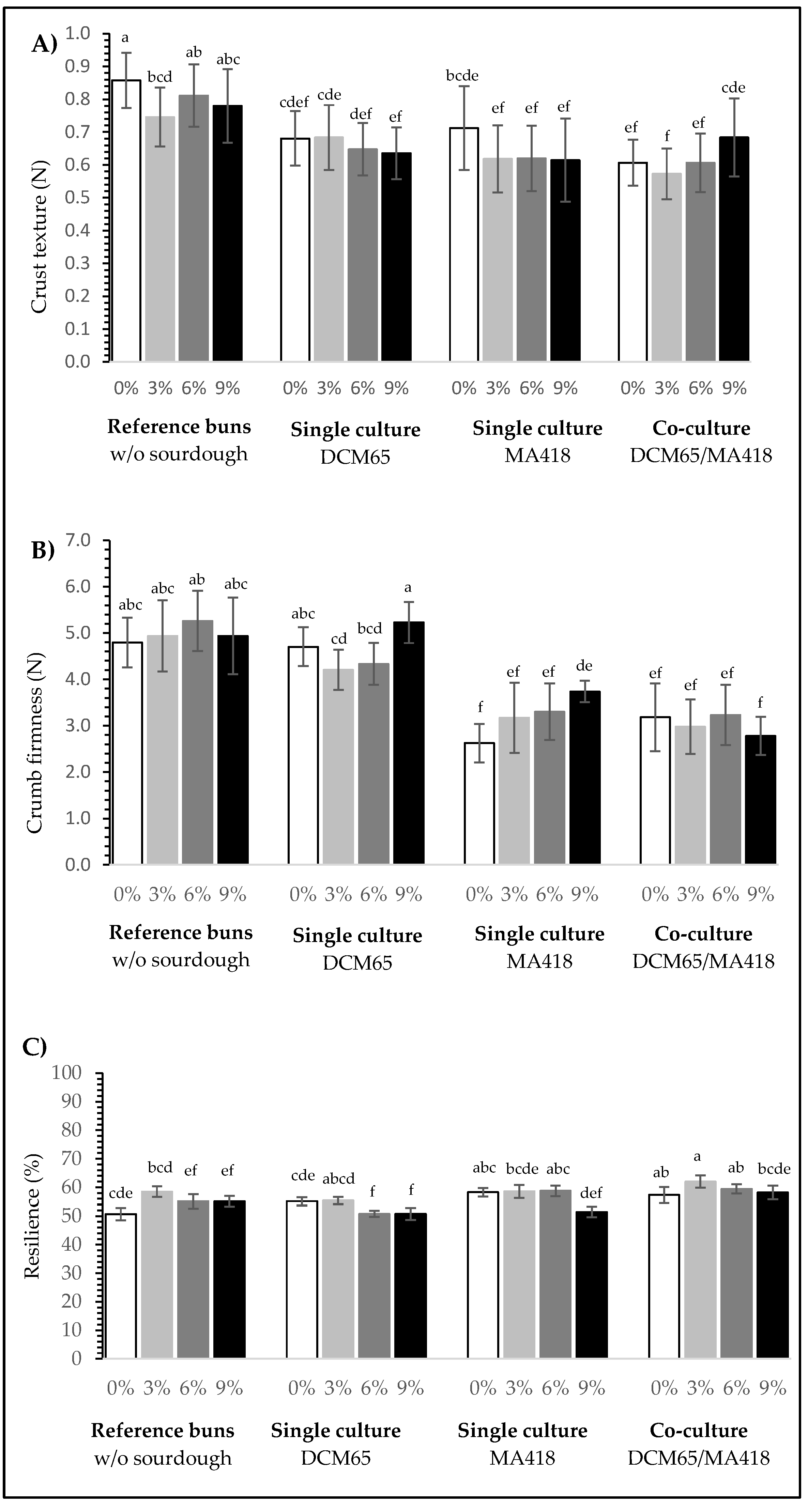
3.2.5. Bun Texture
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gioia, L.C.; Ganancio, J.R.; Steel, C.J. Food additives and processing aids used in breadmaking. In Food Additives; Karunaratne, D.N., Pamunuwa, G., Eds.; InTech: London, UK, 2017; pp. 147–166. [Google Scholar]
- Sahin, A.W.; Axel, C.; Arendt, E.K. Understanding the function of sugar in burger buns: A fundamental Study. Eur. Food Res. Technol. 2017, 243, 1905–1915. [Google Scholar] [CrossRef]
- Bean, M.M.; Yamazaki, W.T. Wheat starch gelatinization in sugar solutions. Sucrose: Microscopy and viscosity effects. Cereal Chem. 1977, 55, 936–944. [Google Scholar]
- Davis, E.A. Functionality of sugars: Physicochemical interactions in food. Am. J. Clin. Nutr. 1995, 62, 170–177. [Google Scholar] [CrossRef]
- Cauvain, S.P. Breadmaking: An Overview. In Breadmaking, 2nd ed.; Cauvain, S.P., Ed.; Woodhead Publishing: Sawston, UK, 2012; pp. 9–31. [Google Scholar]
- Azizi, M.H.; Rajabzadeh, N.; Riahi, E. Effect of mono-diglyceride and lecithin on dough rheological characteristics and quality of flat bread. LWT-Food Sci. Technol. 2003, 36, 189–193. [Google Scholar] [CrossRef]
- Gómez, M.; del Real, S.; Rosell, C.M.; Ronda, F.; Blanco, C.A.; Caballero., P.A. Functionality of different emulsifiers on the performance of breadmaking and wheat bread quality. Eur. Food Res. Technol. 2004, 219, 145–150. [Google Scholar] [CrossRef]
- Legan, J.D. Mould spoilage of bread: The problem and some solutions. Int. Biodeter. Biodegr. 1993, 32, 33–53. [Google Scholar] [CrossRef]
- Marín, S.; Guynot, M.E.; Neira, P.; Bernadó, M.; Sanchis, V.; Ramos, A.J. Risk assessment of the use of sub-optimal levels of weak-acid preservatives in the control of mould growth on bakery products. Int. J. Food Microbiol. 2002, 79, 203–211. [Google Scholar] [CrossRef]
- Samapundo, S.; Devlieghere, F.; Vroman, A.; Eeckhout, M. Antifungal activity of fermentates and their potential to replace propionate in bread. LWT-Food Sci. Technol. 2017, 76, 101–107. [Google Scholar] [CrossRef]
- Passarinho, A.T.P.; Dias, N.F.; Camilloto, G.P.; Cruz, R.S.; Otoni, C.G.; Moraes, A.R.F.; de Soares, N.F.F. Sliced bread preservation through oregano essential oil-containing sachet: Bread preservation through antimicrobial sachet. J. Food Process. Eng. 2014, 37, 53–62. [Google Scholar] [CrossRef]
- Dong, Y.; Karboune, S. A Review of bread qualities and current strategies for bread bioprotection: Flavor, sensory, rheological, and textural attributes. Compr. Rev. Food Sci. Food Saf. 2021, 20, 1937–1981. [Google Scholar] [CrossRef]
- Gänzle, M.G. Enzymatic and bacterial conversions during sourdough fermentation. Food Microbiol. 2014, 37, 2–10. [Google Scholar] [CrossRef]
- Abid, Y.; Azabou, S.; Blecker, C.; Gharsallaoui, A.; Corsaro, M.M.; Besbes, S.; Attia, H. Rheological and emulsifying properties of an exopolysaccharide produced by potential probiotic Leuconostoc citreum-BMS sstrain. Carbohydr. Polym. 2021, 256, 117523. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.C.; Nakamura, L.K. Production of mannitol and lactic acid by fermentation with Lactobacillus intermedius NRRL B-3693. Biotechnol. Bioeng. 2003, 82, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Wisselink, H.W.; Weusthuis, R.A.; Eggink, G.; Hugenholtz, J.; Grobben, G.J. Mannitol production by lactic acid bacteria: A review. Int. Dairy J. 2002, 12, 151–161. [Google Scholar] [CrossRef]
- Petrova, P.; Emanuilova, M.; Petrov, K. Amylolytic Lactobacillus strains from Bulgarian fermented beverage boza. J. Nat. Res. 2010, 65, 218–224. [Google Scholar] [CrossRef]
- Ding, S.; Yang, J. The effects of sugar alcohols on rheological properties, functionalities, and texture in baked products—A review. Trends Food Sci. Technol. 2021, 111, 670–679. [Google Scholar] [CrossRef]
- Sahin, A.W.; Axel, C.; Zannini, E.; Arendt, E.K. Xylitol, mannitol and maltitol as potential sucrose replacers in burger buns. Food Funct. 2018, 9, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Sahin, A.W.; Rice, T.; Zannini, E.; Axel, C.; Coffey, A.; Lynch, K.M.; Arendt, E.K. Leuconostoc citreum TR116: In-situ production of mannitol in sourdough and its application to reduce sugar in burger buns. Int. J. Food Microbiol. 2019, 302, 80–89. [Google Scholar] [CrossRef]
- Galle, S.; Arendt, E.K. Exopolysaccharides from sourdough lactic acid bacteria. Crit. Rev. Food Sci. Nutr. 2014, 54, 891–901. [Google Scholar] [CrossRef]
- Schwab, C.; Mastrangelo, M.; Corsetti, A.; Gänzle, M. Formation of oligosaccharides and polysaccharides by Lactobacillus reuteri LTH5448 and Weissella cibaria 10M in sorghum sourdoughs. Cereal Chem. 2008, 85, 679–684. [Google Scholar] [CrossRef]
- Coda, R.; Xu, Y.; Moreno, D.S.; Mojzita, D.; Nionelli, L.; Rizzello, C.G.; Katina, K. Performance of Leuconostoc citreum FDR241 during wheat flour sourdough Type I propagation and transcriptional analysis of exopolysaccharides biosynthesis genes. Food Microbiol. 2018, 76, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Dlusskaya, E.; Jänsch, A.; Schwab, C.; Gänzle, M.G. Microbial and chemical analysis of a kvass fermentation. Eur. Food Res. Technol. 2008, 227, 261–266. [Google Scholar] [CrossRef]
- Müller, D.C.; Mischler, S.; Schönlechner, R.; Miescher Schwenninger, S. Multiple techno-functional characteristics of Leuconostoc and their potential in sourdough fermentations. Microorganisms 2021, 9, 1633. [Google Scholar] [CrossRef]
- Goesaert, H.; Slade, L.; Levine, H.; Delcour, J.A. Amylases and bread firming—An integrated view. J. Cereal Sci. 2009, 50, 345–352. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Reddy, G.; Altaf, M.d.; Naveena, B.J.; Venkateshwar, M.; Kumar, E.V. Amylolytic bacterial lactic acid fermentation—A review. Biotechnol. Adv. 2008, 26, 22–34. [Google Scholar] [CrossRef] [PubMed]
- Sanni, A.I.; Morlon-Guyot, J.; Guyot, J.P. New efficient amylase-producing strains of Lactobacillus plantarum and L. fermentum isolated from different Nigerian traditional fermented foods. Int. J. Food Microbiol. 2002, 72, 53–62. [Google Scholar] [CrossRef]
- Sadiq, F.A.; Yan, B.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Lactic acid bacteria as antifungal and anti-mycotoxigenic agents: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1403–1436. [Google Scholar] [CrossRef] [Green Version]
- Black, B.A.; Zannini, E.; Curtis, J.M.; Gänzle, M.G. Antifungal hydroxy fatty acids produced during sourdough fermentation: Microbial and enzymatic pathways, and antifungal activity in bread. Appl. Environ. Microbiol. 2013, 79, 1866–1873. [Google Scholar] [CrossRef] [Green Version]
- Dal Bello, F.; Clarke, C.I.; Ryan, L.A.M.; Ulmer, H.; Schober, T.J.; Ström, K.; Sjögren, J.; van Sinderen, D.; Schnürer, J.; Arendt, E.K. Improvement of the quality and shelf life of wheat bread by fermentation with the antifungal strain Lactobacillus plantarum FST 1.7. J. Cereal Sci. 2007, 45, 309–318. [Google Scholar] [CrossRef]
- Axel, C.; Brosnan, B.; Zannini, E.; Peyer, L.C.; Furey, A.; Coffey, A.; Arendt, E.K. Antifungal activities of three different Lactobacillus species and their production of antifungal carboxylic acids in wheat sourdough. Appl. Microbiol. Biotechnol. 2016, 100, 1701–1711. [Google Scholar] [CrossRef] [PubMed]
- Miescher Schwenninger, S.; Freimüller Leischtfeld, S.; Gantenbein-Demarchi, C. High-throughput identification of the microbial biodiversity of cocoa bean fermentation by MALDI-TOF MS. Lett. Appl. Microbiol. 2016, 63, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Quattrini, M.; Liang, N.; Fortina, M.G.; Xiang, S.; Curtis, J.M.; Gänzle, M. Exploiting synergies of sourdough and antifungal organic acids to delay fungal spoilage of bread. Int. J. Food Microbiol. 2019, 302, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Müller, D.C.; Nguyen, H.; Li, Q.; Schönlechner, R.; Miescher Schwenninger, S.; Wismer, W.; Gänzle, M. Enzymatic and microbial conversions to achieve sugar reduction in bread. Food Res. Int. 2021, 9, 110296. [Google Scholar] [CrossRef]
- Markowicz, D.; Monaro, E.; Siguemoto, E.; Sefor, M. Maillard reaction products in processed food: Pros and cons. In Food Industrial Processes—Methods and Equipment; Valdez, B., Ed.; InTech: London, UK, 2012; ISBN 978-953-307-905-9. [Google Scholar]
- Shen, Y.; Chen, G.; Li, Y. Bread characteristics and antioxidant activities of maillard reaction products of white pan bread containing various sugars. LWT 2018, 95, 308–315. [Google Scholar] [CrossRef]
- Martins, S.I.F.S.; Jongen, W.M.F.; van Boekel, M.A.J.S. A review of maillard reaction in food and implications to kinetic modelling. Trends Food Sci. Technol. 2000, 11, 364–373. [Google Scholar] [CrossRef]
- Jouquand, C.; Niquet-Léridon, C.; Jacolot, P.; Petit, N.; Marier, D.; Gadonna-Widehem, P. Effects of maillard reaction products on sensory and nutritional qualities of the traditional French baguette: Food Quality of the French Baguette. J. Food Sci. 2018, 83, 2424–2431. [Google Scholar] [CrossRef]
- Goldfein, K.R.; Slavin, J.L. Why sugar is added to food: Food Science 101: Challenges of labeling added sugars. Compr. Rev. Food Sci. Food Saf. 2015, 14, 644–656. [Google Scholar] [CrossRef] [Green Version]
- Kaditzky, S.; Seitter, M.; Hertel, C.; Vogel, R.F. Performance of Lactobacillus sanfranciscensis TMW 1.392 and its levansucrase deletion mutant in wheat dough and comparison of their impact on bread quality. Eur. Food Res. Technol. 2008, 227, 433–442. [Google Scholar] [CrossRef]
- Purhagen, J.K.; Sjöö, M.E.; Eliasson, A.-C. Starch affecting anti-staling agents and their function in freestanding and pan-baked bread. Food Hydrocoll. 2011, 25, 1656–1666. [Google Scholar] [CrossRef]
- Chen, Y.; Eder, S.; Schubert, S.; Gorgerat, S.; Boschet, E.; Baltensperger, L.; Boschet, E.; Städeli, C.; Kuster, S.; Fischer, P.; et al. Influence of amylase addition on bread quality and bread staling. ACS Food Sci. Technol. 2021, 1, 1143–1150. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sugar Addition (% of Flour) | Sourdough Addition (% of Flour) | Culture Addition | |
|---|---|---|---|
| Reference buns | 9 | - | |
| 6 | - | ||
| 3 | - | - | |
| 0 | - | ||
| Sourdough buns | 9 | 20 | Single culture: Lc. citreum DCM65 |
| 6 | 20 | ||
| 3 | 20 | ||
| 0 | 20 | ||
| 9 | 20 | Single culture: Lpb. plantarum subsp. plantarum MA418 | |
| 6 | 20 | ||
| 3 | 20 | ||
| 0 | 20 | ||
| 9 | 20 | Co-culture: Lc. citreum DCM65 & Lpb. plantarum subsp. plantarum MA418 | |
| 6 | 20 | ||
| 3 | 20 | ||
| 0 | 20 |
| Ingredients (% of Flour) | Reference Bun (w/o Sourdough) | Sourdough Bun (20% Addition) |
|---|---|---|
| Dough ingredients | ||
| Flour | 100 | 80 |
| Tap water | 60–64 (1) | 40–44 |
| Sugar (2) | 0/3/6/9 | 0/3/6/9 |
| Canola oil | 5 | 5 |
| Salt | 1.6 | 1.6 |
| Yeast | 2 | 2 |
| Wheat gluten Sourdough (3) | 0.6 | 0.6 43 |
| Sourdough ingredients | ||
| Flour | 20 | |
| Tap water (including liquid culture) | 20 | |
| Sucrose | 1.5 | |
| Fructose | 1.5 |
| Sourdough Fermentation with Single and Co-Culture: | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DCM65 | MA418 | DCM65 and MA418 | ||||||||||
| Sugar content in dough (% flour) | ||||||||||||
| 0 | 3 | 6 | 9 | 0 | 3 | 6 | 9 | 0 | 3 | 6 | 9 | |
| LAB counts (log CFU/g) | ||||||||||||
| t0 | 8.0 ± 0.2 a | 7.8 ± 0.1 a | 8.0 ± 0.2 a | 7.9 ± 0.4 a | 7.8 ± 0.2 a | 7.8 ± 0.2 a | 7.7 ± 0.1 a | 7.8 ± 0.5 a | 7.6 ± 0.2 a | 7.9 ± 0.1 a | 7.8 ± 0.0 a | 7.5 ± 0.1 a |
| t24 | ± 0.3 ab | ± 0.1 a | ± 0.2 ab | ± 0.1 a | ± 0.3 ab | ± 0.1 ab | 9.1 ± 0.2 ab | 8.8 ± 0.5 ab | 8.8 ± 0.4 ab | ± 0.3 b | 8.7 ± 0.4 ab | 8.9 ± 0.3 ab |
| pH | ||||||||||||
| t0 | ± 0.0 a | ± 0.0 a | ± 0.0 a | ± 0.0 a | ± 0.0 a | ± 0.0 a | ± 0.0 a | ± 0.0 a | a | a | a | ± 0.1 a |
| t24 | ± 0.0 a | ± 0.0 a | ± 0.0 a | ± 0.1 a | ± 0.1 b | ± 0.0 b | ± 0.0 b | ± 0.0 b | ± 0.1 b | ± 0.1 b | ± 0.0 b | ± 0.1 b |
| Fructose (mg/g sourdough) | ||||||||||||
| t0 | 15.7 ± 2.4 ab | a | a | ab | ± 0.6 ab | ± 0.3 ab | ± 3.8 ab | ± 1.3 ab | ab | 10.4 ± 2.2 b | 12.1 ± 0.9 ab | 10.4 ± 2.7 b |
| t24 | 5.5 ± 0.9 e | 6.2 ± 0.3 de | 5.4 ± 1.4 e | 5.5 ± 0.8 e | 11.2 ± 0.3 ab | 12.2 ± 0.1 a | 10.9 ± 2.2 abc | 11.0 ± 0.7 ab | 6.9 ± 1.1 de | 7.7 ± 0.7 de | 9.1 ± 1.2 bcd | 7.9 ± 0.7 cde |
| Mannitol (mg/g sourdough) | ||||||||||||
| t0 | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| t24 | a | ± 0.4 a | a | ± 0.5 a | n.d. | n.d. | n.d. | n.d. | b | 3.3 ± 0.3 b | ± 0.3 b | ± 1.3 b |
| Reference Buns | Sourdough Buns with Starter Cultures: | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| w/o Sourdough | DCM65 | MA418 | DCM65/MA418 | ||||||||||||
| Sugar addition (% flour) | |||||||||||||||
| 0 | 3 | 6 | 9 | 0 | 3 | 6 | 9 | 0 | 3 | 6 | 9 | 0 | 3 | 6 | 9 |
| L*-value crust | |||||||||||||||
| 69.9 ± 4.1 a | 61.1 ± 3.9 bcde | 59.2 ± 4.5 cdef | 58.4 ± 4.5 def | 60.7 ± 2.9 cde | 63.0 ± 4.8 abcde | 64.0 ± 5.4 abcd | 59.7 ± 4.7 cdef | 65.1 ± 3.4 abc | 60.7 ± 3.8 cdef | 58.2 ± 5.6 def | 58.9 ± 6.1 cdef | 67.5 ± 2.4 ab | 58.9 ± 5.3 cdef | 57.5 ± 5.9 ef | 54.1 ± 5.8 f |
| a*-value crust | |||||||||||||||
| 2.0 ± 1.9 d | 8.1 ± 2.7 c | 10.7 ± 2.1 bc | 11.1 ± 2.8 abc | 1.7 ± 1.7 d | 8.9 ± 3.5 c | 8.8 ± 3.7 ± c | 12.2 ± 2.8 ab | 4.0 ± 2.2 d | 8.2 ± 2.5 c | 10.9 ± 3.6 abc | 10.9 ± 3.7 abc | 3.4 ± 1.8 d | 10.2 ± 3.4 bc | 12.7 ± 3.3 ab | 14.0 ± 2.8 a |
| b*-value crust | |||||||||||||||
| 27.3 ± 3.3 d | 32.9 ± 1.4 abc | 34.9 ± 0.8 a | 34.5 ± 1.4 ab | 19.4 ± 2.8 e | 35.0 ± 2.3 a | 34.8 ± 2.7 a | 36.8 ± 1.2 a | 29.0 ± 2.8 cd | 33.5 ± 1.5 ab | 34.7 ± 1.6 ab | 34.9 ± 1.8 a | 30.2 ± 2.6 bcd | 34.2 ± 1.3 ab | 35.6 ± 1.3 a | 34.8 ± 2.0 a |
| L*-value crumb | |||||||||||||||
| 69.1 ± 2.4 cd | 69.8 ± 1.8 bcd | 71.0 ± 1.2 ab | 71.2 ± 1.1 ab | 70.1 ± 1.3 bcd | 72.3 ± 1.4 a | 71.8 ± 2.8 a | 72.4 ± 1.3 a | 68.8 ± 1.4 d | 68.8 ± 1.3 d | 70.6 ± 1.5 bc | 70.8 ± 1.2 bc | 70.4 ± 1.0 bcd | 70.1 ± 1.3 bcd | 71.3 ± 1.3 ab | 71.3 ± 1.2 ab |
| a*-value crumb | |||||||||||||||
| −1.2 ± 0.1 a | −1.2 ± 0.1 a | −1.2 ± 0.1 a | −1.3 ± 0.1 a | −1.1 ± 0.1 a | −1.3 ± 0.1 a | −0.5 ± 2.0 a | −1.3 ± 0.1 a | −1.2 ± 0.1 a | −1.3 ± 0.1 a | −1.2 ± 0.1 a | −1.2 ± 0.1 a | −1.2 ± 0.1 a | −1.2 ± 0.0 a | −1.2 ± 0.1 a | −1.2 ± 0.0 a |
| b*-value crumb | |||||||||||||||
| 16.2 ± 0.5 bcd | 16.5 ± 0.5 bcd | 16.7 ± 0.7 abc | 16.5 ± 0.3 bc | 16.7 ± 0.5 bc | 16.4 ± 0.5 bcd | 18.1 ± 2.6 a | 17.4 ± 0.5 ab | 14.8 ± 0.7 e | 15.1 ± 0.8 de | 15.6 ± 0.7 cde | 16.0 ± 0.4 cde | 15.6 ± 0.7 cde | 15.7 ± 0.7 cde | 15.8 ± 0.8 cde | 14.8 ± 0.5 e |
| aw-value | |||||||||||||||
| 0.933 ± 0.006 ab | 0.931 ± 0.002 abc | 0.927 ± 0.002 bcd | 0.916 ± 0.006 e | 0.934 ± 0.004 ab | 0.919 ± 0.007 de | 0.913 ± 0.010 e | 0.910 ± 0.007 e | 0.936 ± 0.003 a | 0.926 ± 0.005 cd | 0.926 ± 0.005 cd | 0.916 ± 0.006 e | 0.935 ± 0.002 a | 0.929 ± 0.003 abc | 0.916 ± 0.004 e | 0.917 ± 0.003 e |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, D.C.; Schipali, S.; Näf, P.; Kinner, M.; Miescher Schwenninger, S.; Schönlechner, R. Potential of a Techno-Functional Sourdough and Its Application in Sugar-Reduced Soft Buns. Fermentation 2022, 8, 42. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8020042
Müller DC, Schipali S, Näf P, Kinner M, Miescher Schwenninger S, Schönlechner R. Potential of a Techno-Functional Sourdough and Its Application in Sugar-Reduced Soft Buns. Fermentation. 2022; 8(2):42. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8020042
Chicago/Turabian StyleMüller, Denise C., Stefanie Schipali, Patrick Näf, Mathias Kinner, Susanne Miescher Schwenninger, and Regine Schönlechner. 2022. "Potential of a Techno-Functional Sourdough and Its Application in Sugar-Reduced Soft Buns" Fermentation 8, no. 2: 42. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8020042