
The Use of Baikal Psychrophilic Actinobacteria for Synthesis of Biologically Active Natural Products from Sawdust Waste

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation of Actinobacteria
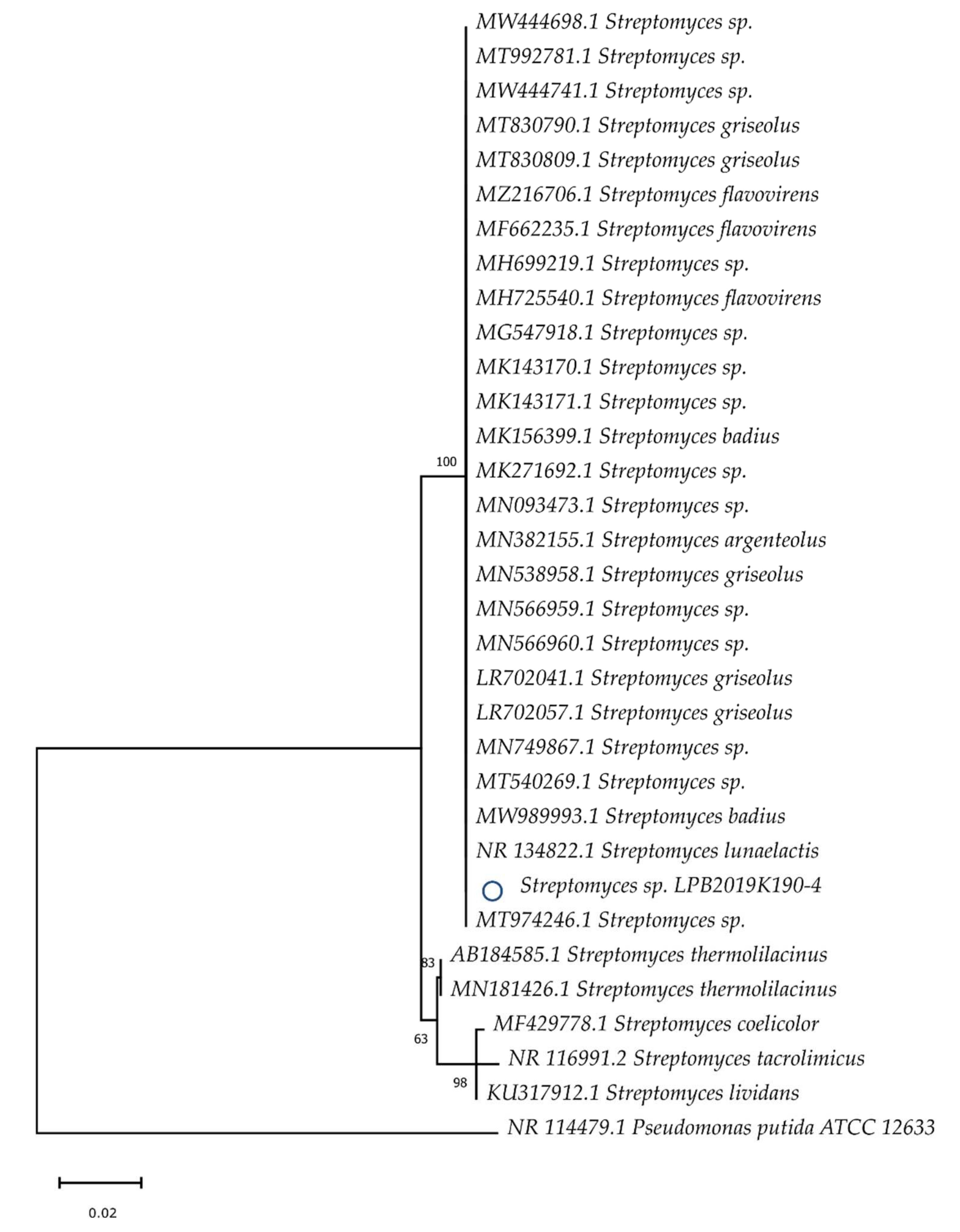
2.2. Identification
2.3. Cultivation
2.4. Extraction of Natural Products
2.5. Assay of Antibiotic Activity of Extracts from Isolated Strains
2.6. Estimation of Biotechnological Potential Using the LC-MS Approach
3. Results
3.1. Identification of Isolated Strains and Their Ability to Grow on Sawdust
3.2. Antibiotic Activity of Isolated Strains
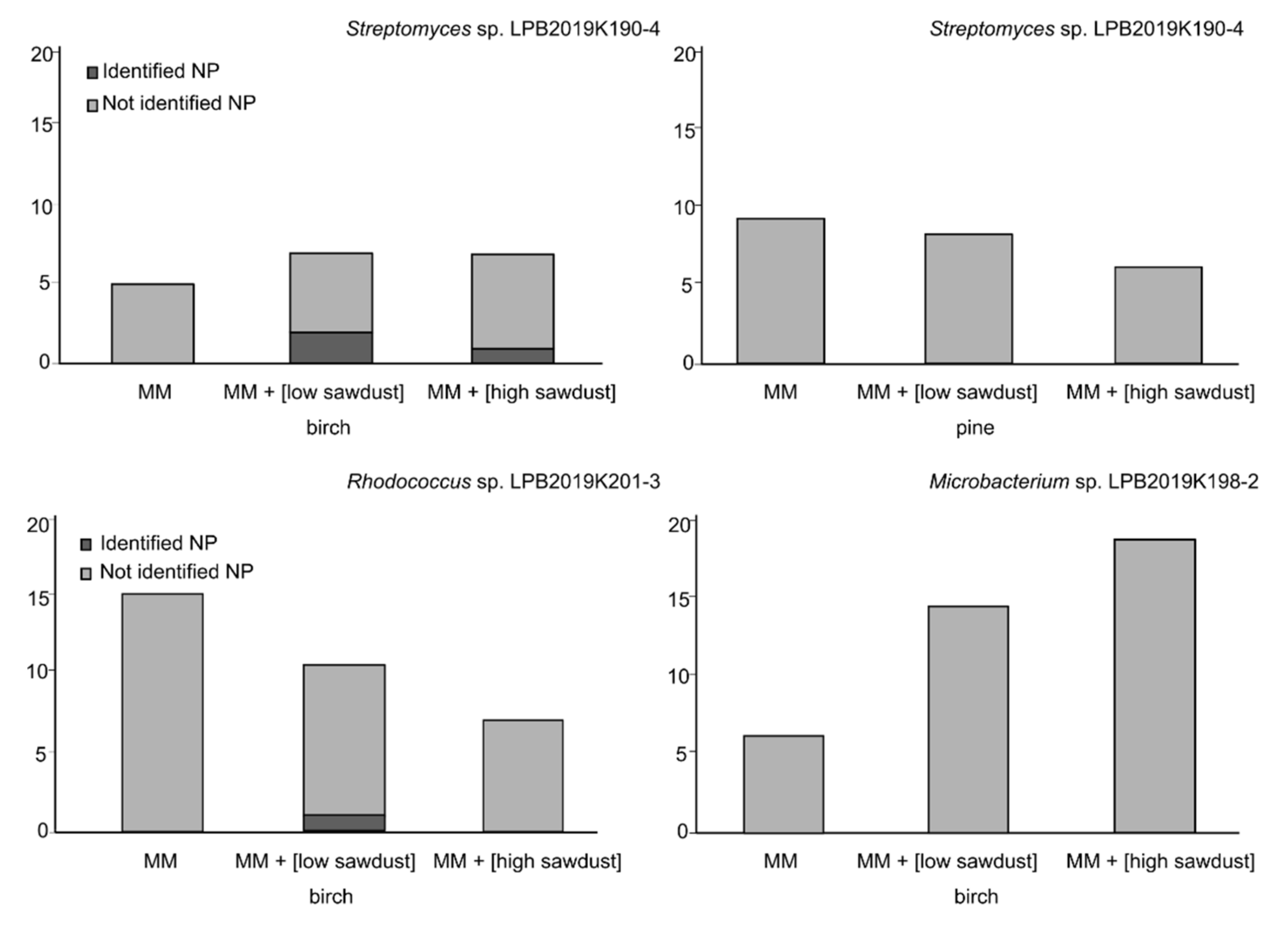
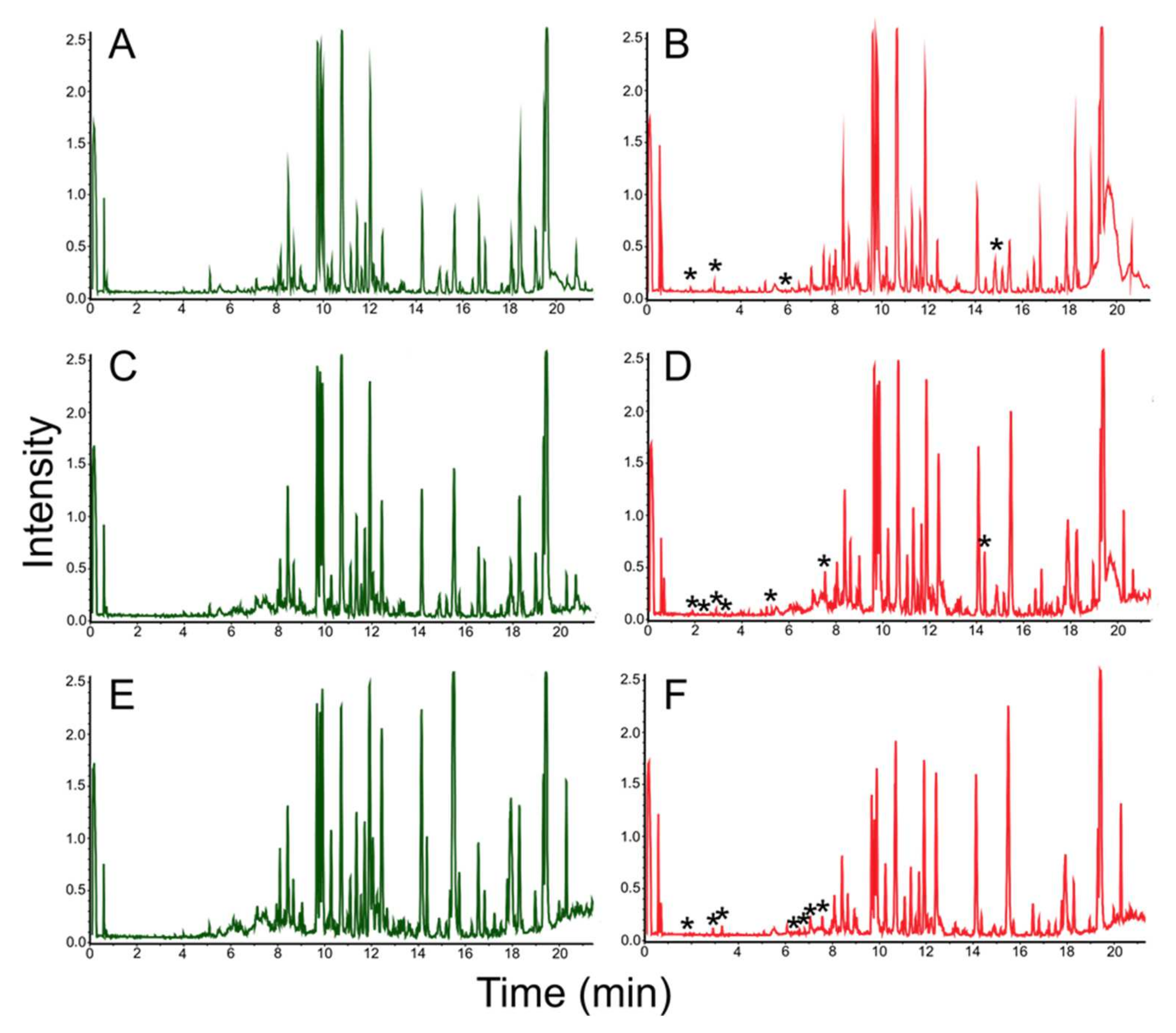
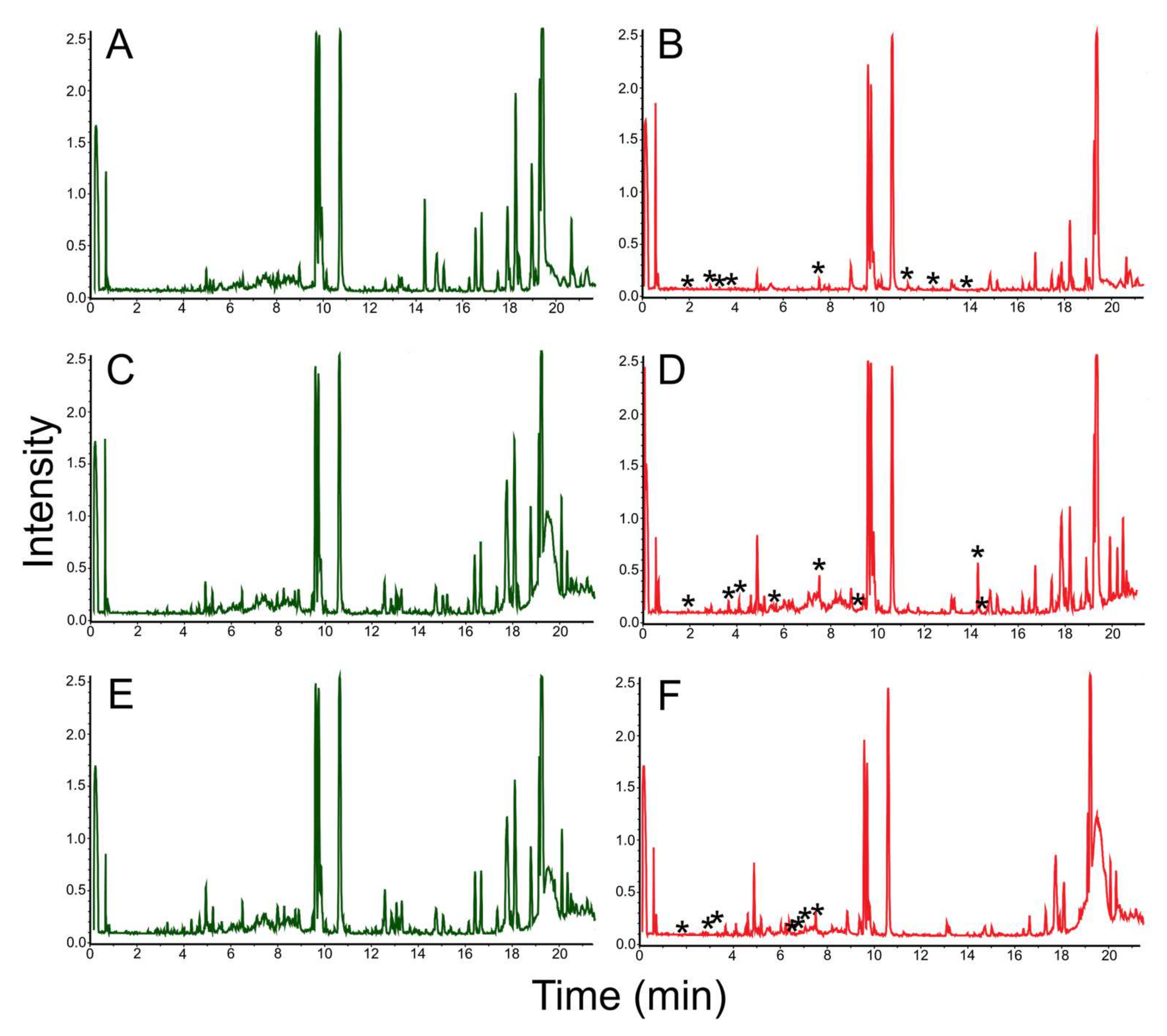
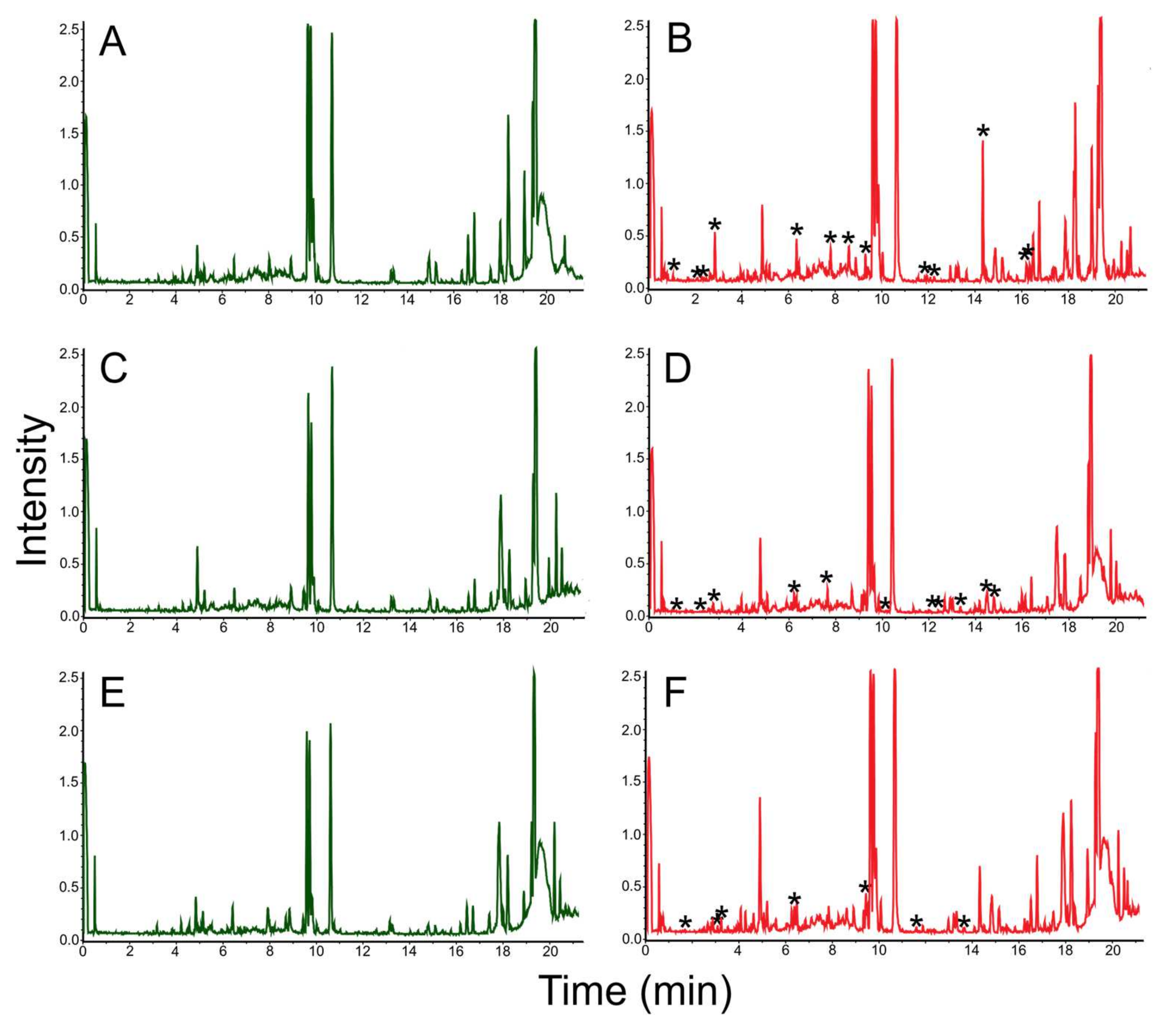
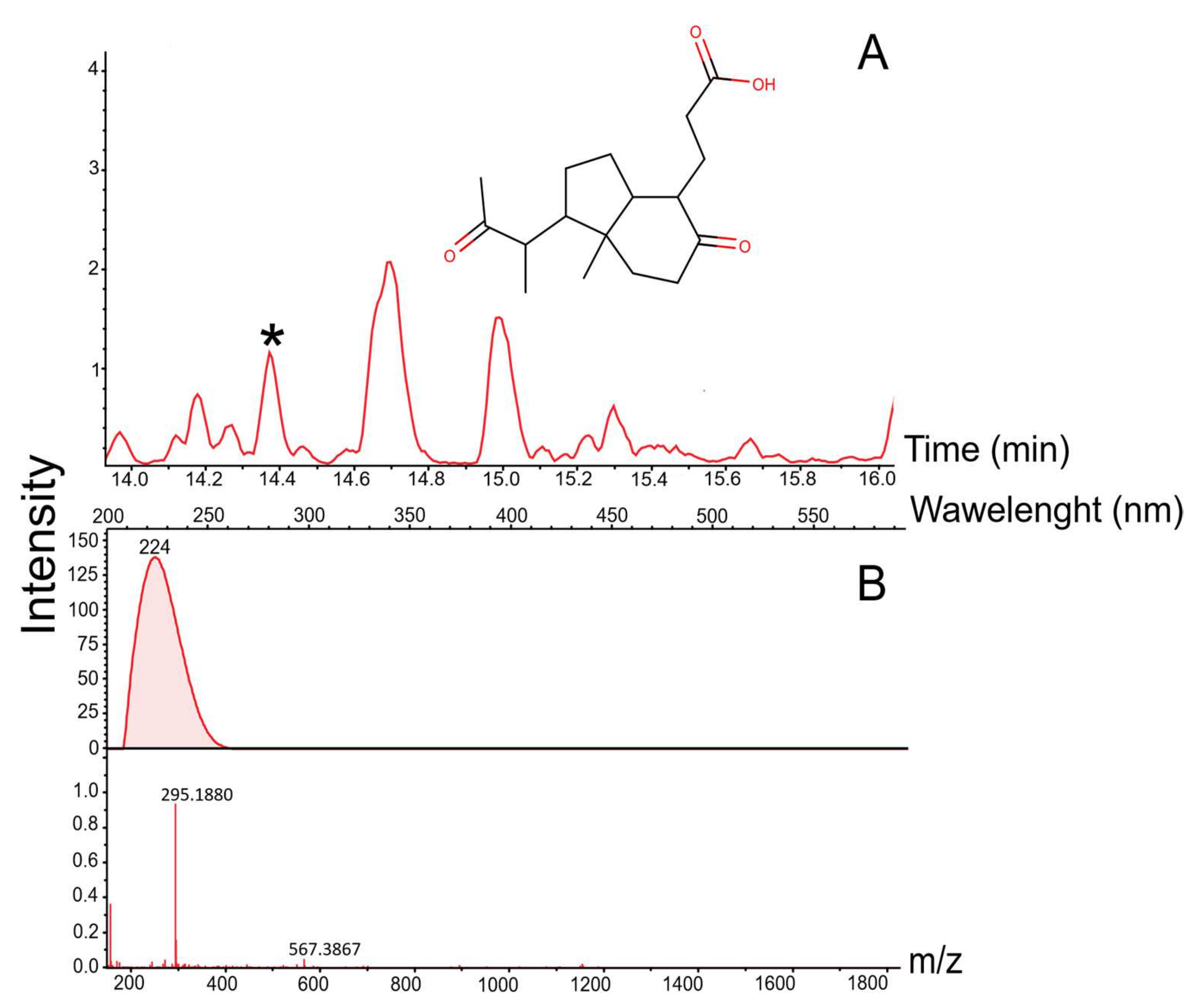
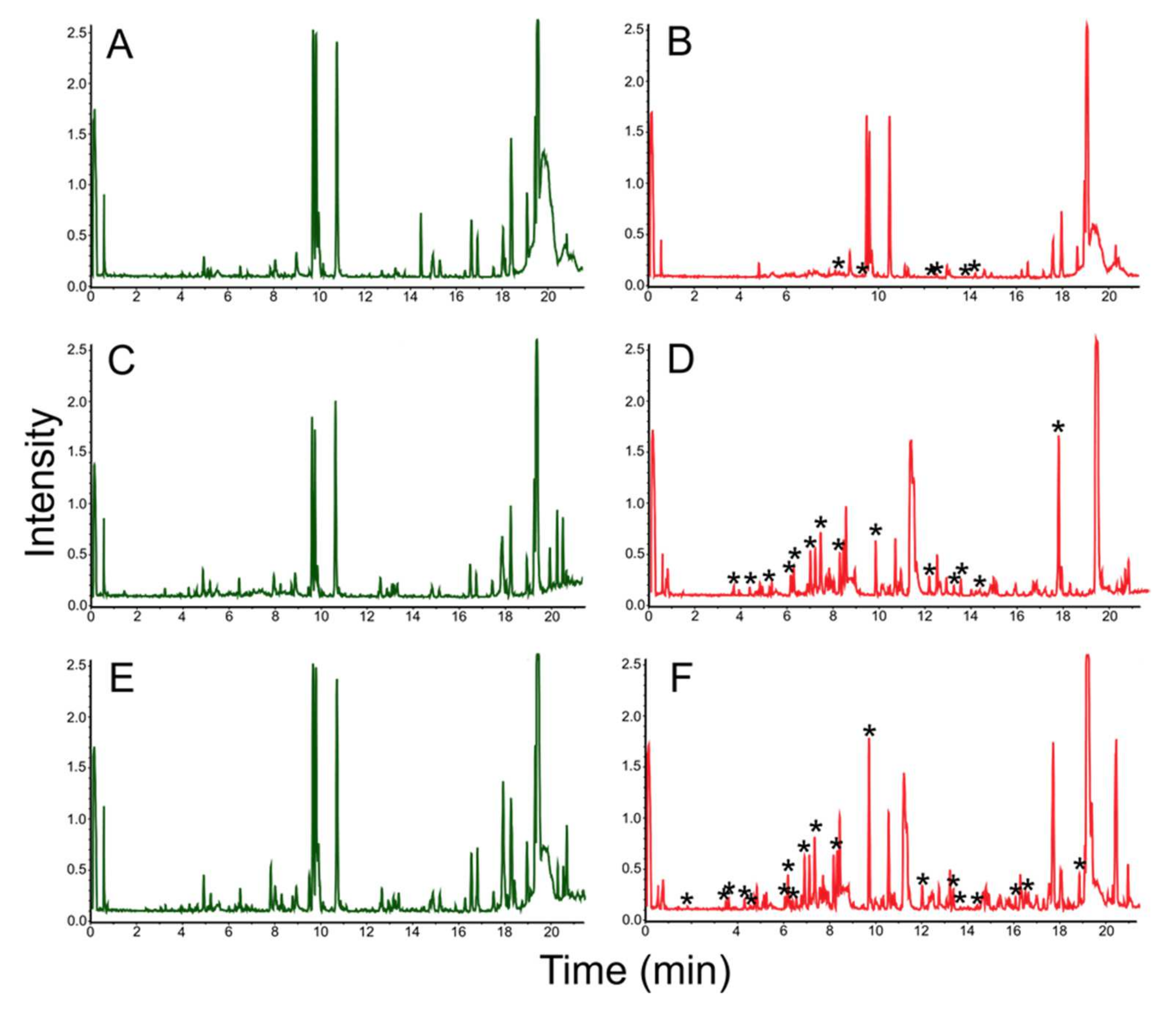
3.3. Mass Spectrometric Estimation of Biotechnological Potential
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kundungal, H.; Gangarapu, M.; Sarangapani, S.; Patchaiyappan, A.; Devipriya, S.P. Efficient Biodegradation of Polyethylene (HDPE) Waste by the Plastic-Eating Lesser Waxworm (Achroia Grisella). Environ. Sci. Pollut. Res. 2019, 26, 18509–18519. [Google Scholar] [CrossRef] [PubMed]
- Molino, A.; Larocca, V.; Chianese, S.; Musmarra, D. Biofuels Production by Biomass Gasification: A Review. Energies 2018, 11, 811. [Google Scholar] [CrossRef] [Green Version]
- Mollica, G.J.G.; Balestieri, J.A.P. Is It Worth Generating Energy with Garbage? Defining a Carbon Tax to Encourage Waste-to-Energy Cycles. Appl. Therm. Eng. 2020, 173, 115195. [Google Scholar] [CrossRef]
- Luo, J.; Donnison, A.; Ross, C.; Bolan, N.; Ledgard, S.; Clark, D.; Qiu, W. Sawdust and Bark to Treat Nitrogen and Faecal Bacteria in Winter Stand-Off Pads on A Dairy Farm. N. Z. J. Agr. Res. 2008, 51, 331–340. [Google Scholar] [CrossRef]
- Zorpas, A.A.; Loizidou, M. Sawdust and Natural Zeolite as A Bulking Agent for Improving Quality of a Composting Product from Anaerobically Stabilized Sewage Sludge. Bioresour. Technol. 2008, 99, 7545–7552. [Google Scholar] [CrossRef]
- Cho, E.J.; Trinh, L.T.P.; Song, Y.; Lee, Y.G.; Bae, H.J. Bioconversion of Biomass Waste into High Value Chemicals. Bioresour. Technol. 2020, 298, 122386. [Google Scholar] [CrossRef]
- Amran, M.A.; Palaniveloo, K.; Fauzi, R.; Satar, N.M.; Mohidin, T.B.M.; Mohan, G.; Seelan, J.S.S. Value-Added Metabolites from Agricultural Waste and Application of Green Extraction Techniques. Sustainability 2021, 13, 11432. [Google Scholar] [CrossRef]
- Hajam, M.E.; Plavan, G.I.; Kandri, N.I.; Dumitru, G.; Nicoara, M.N.; Zerouale, A.; Faggio, C. Evaluation of Softwood and Hardwood Sawmill Wastes Impact on the Common Carp “Cyprinus Carpio” and Its Aquatic Environment: An Oxidative Stress Study. Environ. Toxicol. Pharmacol. 2020, 75, 103327. [Google Scholar] [CrossRef]
- Samarin, E.N.; Kravchenko, N.S.; Zerkal, O.V.; Chernov, M.S.; Rodkina, I.A. Grouting of Waste of the Baikal Pulp and Paper Mill to Reduce the Technogenic Impact to the Baikal Lake Ecosystem. J. Geosci. Environ. Prot. 2020, 8, 112–118. [Google Scholar] [CrossRef]
- Amin, D.H.; Abdallah, N.A.; Abolmaaty, A.; Tolba, S.; Wellington, E.M.H. Microbiological and Molecular Insights on Rare Actinobacteria Harboring Bioactive Prospective. Bull. Natl. Res. Cent. 2020, 44, 5. [Google Scholar] [CrossRef]
- Wurzbacher, C.; Kerr, J.; Grossart, H.P. Aquatic fungi. In The Dynamical Processes of Biodiversity: Evolution and Spatial Distribution; Few Study Cases; Grillo, O., Venora, G., Eds.; InTech Open: Rijeka, Croatia, 2011; pp. 227–258. [Google Scholar]
- Wang, C.; Dong, D.; Wang, H.; Müller, K.; Qin, Y.; Wang, H.; Wu, W. Metagenomic Analysis of Microbial Consortia Enriched from Compost: New Insights into the Role of Actinobacteria in Lignocellulose Decomposition. Biotechnol. Biofuels 2016, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Yanful, E.K.; Bassi, A.S. A Review of Plastic Waste Biodegradation. Crit. Rev. Biotechnol. 2005, 25, 243–250. [Google Scholar] [CrossRef]
- Hazarika, J.; Khwairakpam, M. Evaluation of Biodegradation Feasibility through Rotary Drum Composting Recalcitrant Primary Paper Mill Sludge. Waste Manag. 2018, 76, 275–283. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Kuyukina, M.S.; Krivoruchko, A.V.; Plekhov, O.A.; Naimark, O.B.; Podorozhko, E.A.; Lozinsky, V.I. Biosurfactant-Enhanced Immobilization of Hydrocarbon-Oxidizing Rhodococcus Ruber on Sawdust. Appl. Microbiol. Biotechnol. 2013, 97, 5315–5327. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Vikhareva, E.V.; Richkova, M.I.; Mukhutdinova, A.N.; Karpenko, J.N. Biodegradation of Drotaverine Hydrochloride by Free and Immobilized Cells of Rhodococcus Rhodochrous IEGM 608. World J. Microbiol. Biotechnol. 2012, 28, 2997–3006. [Google Scholar] [CrossRef]
- Baltz, R.H. Natural Product Drug Discovery in the Genomic Era: Realities, Conjectures, Misconceptions, and Opportunities. J. Ind. Microbiol. Biotechnol. 2019, 46, 281–299. [Google Scholar] [CrossRef]
- Zhang, L.; Demain, A.L. Natural Products: Drug Discovery and Therapeutic Medicine; Humana Press: Totowa, NJ, USA, 2005. [Google Scholar]
- Salwan, R.; Sharma, V. Molecular and Biotechnological Aspects of Secondary Metabolites in Actinobacteria. Microbiol. Res. 2020, 231, 126374. [Google Scholar] [CrossRef]
- List of Prokaryotic Names with Standing in Nomenclature. Available online: https://lpsn.dsmz.de/genus/streptomyces (accessed on 11 April 2022).
- Dhakal, D.; Pokhrel, A.R.; Shrestha, B.; Sohng, J.K. Marine Rare Actinobacteria: Isolation, Characterization, and Strategies for Harnessing Bioactive Compounds. Front. Microbiol. 2017, 8, 1106. [Google Scholar] [CrossRef] [Green Version]
- Hui, M.L.Y.; Tan, L.T.H.; Letchumanan, V.; He, Y.W.; Fang, C.M.; Chan, K.G.; Law, J.W.F.; Lee, L.H. The Extremophilic Actinobacteria: From Microbes to Medicine. Antibiotics 2021, 10, 682. [Google Scholar] [CrossRef]
- Lee, L.H.; Goh, B.H.; Chan, K.G. Editorial: Actinobacteria: Prolific Producers of Bioactive Metabolites. Front. Microbiol. 2020, 11, 1612. [Google Scholar] [CrossRef]
- Takhteev, V.V. On the Current State of Taxonomy of the Baikal Lake Amphipods (Crustacea, Amphipoda) and the Typological Ways of Constructing Their System. Arthsel 2019, 28, 374–402. [Google Scholar] [CrossRef]
- Kostrzewa, M. Application of the MALDI Biotyper to Clinical Microbiology: Progress and Potential. Expert Rev. Proteomic 2018, 15, 193–202. [Google Scholar] [CrossRef]
- Schulthess, B.; Bloemberg, G.V.; Zbinden, A.; Mouttet, F.; Zbinden, R.; Böttger, E.C.; Hombach, M. Evaluation of the Bruker MALDI Biotyper for Identification of Fastidious Gram-Negative Rods. J. Clin. Microbiol. 2016, 54, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Stach, J.E.M.; Maldonado, L.A.; Ward, A.C.; Goodfellow, M.; Bull, A.T. New Primers for the Class Actinobacteria: Application to Marine and Terrestrial Environments. Environ. Microbiol. 2003, 5, 828–841. [Google Scholar] [CrossRef] [Green Version]
- Shieh, J.C.; Martin, H.; Millar, J.B.A. Evidence for a Novel MAPKKK-Independent Pathway Controlling the Stress Activated Sty1/Spc1 MAP Kinase in FIssion Yeast. J. Cell Sci. 1998, 111, 799–807. [Google Scholar]
- The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [CrossRef]
- Felsenstein, J. Phylogenies and the Comparative Method. Am. Nat. 1985, 125, 171–185. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Sticher, O. Natural Product Isolation. Nat. Prod. Rep. 2008, 25, 517. [Google Scholar] [CrossRef]
- Sarker, S.D.; Nahar, L. (Eds.) Natural Products Isolation. Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; Volume 864. [Google Scholar] [CrossRef]
- Mahajan, G.B.; Balachandran, L. Sources of Antibiotics: Hot Springs. Biochem. Pharmacol. 2017, 134, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, F.; Bortolotto, V.; Araujo, S.; Poetini, M.; Sehn, C.; Neto, J.É.; Zeni, G.; Prigol, M. Antimicrobial Effect of 2-Phenylethynyl-Butyltellurium in Escherichia Coli and Its Association with Oxidative Stress. J. Microbiol. Biotechnol. 2018, 28, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Voitsekhovskaia, I.; Paulus, C.; Dahlem, C.; Rebets, Y.; Nadmid, S.; Zapp, J.; Axenov-Gribanov, D.; Rückert, C.; Timofeyev, M.; Kalinowski, J.; et al. New Aquayamycin-Type Angucyclines Isolated from Lake Baikal Derived Streptomyces sp. IB201691-2A. Microorganisms 2020, 8, 680. [Google Scholar] [CrossRef] [PubMed]
- Axenov-Gribanov, D.V.; Morgunova, M.M.; Vasilieva, U.A.; Gamaiunov, S.V.; Dmitrieva, M.E.; Pereliaeva, E.V.; Belyshenko, A.Y.; Luzhetskyy, A.N. Composition of Nutrient Media and Temperature of Cultivation Imposes Effect on the Content of Secondary Metabolites of Nocardiopsis Sp. Isolated from a Siberian Cave. 3 Biotech 2021, 11, 386. [Google Scholar] [CrossRef] [PubMed]
- Błajet-Kosicka, A.; Kosicki, R.; Twarużek, M.; Grajewski, J. Determination of Moulds and Mycotoxins in Dry Dog and Cat Food Using Liquid Chromatography with Mass Spectrometry and Fluorescence Detection. Food Addit. Contam. B 2014, 7, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Buckingham, J. (Ed.) Dictionary of Natural Products, Supplement 4; CRC Press: Boca Raton, FL, USA, 1997. [Google Scholar]
- Macherla, V.R.; Liu, J.; Bellows, C.; Teisan, S.; Nicholson, B.; Lam, K.S.; Potts, B.C.M. Glaciapyrroles A, B, and C, Pyrrolosesquiterpenes from a Streptomyces sp. Isolated from an Alaskan Marine Sediment. J. Nat. Prod. 2005, 68, 780–783. [Google Scholar] [CrossRef]
- Burres, N.S.; Premachandran, U.; Hoselton, S.; Cwik, D.; Hochlowski, J.E.; Ye, Q.; Sunga, G.N.; Karwowski, J.P.; Jackson, M.; Whittern, D.N.; et al. Simple Aromatics Identified with a NFAT–lacZ Transcription Assay for the Detection of Immunosuppressants. J. Antibiot. 1995, 48, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni-Almeida, A.A.; Brahma, M.K.; Padmanabhan, P.; Mishra, P.D.; Parab, R.R.; Gaikwad, N.V.; Thakkar, C.S.; Tokdar, P.; Ranadive, P.V.; Nair, A.S.; et al. Fermentation, Isolation, Structure, and Antidiabetic Activity of NFAT-133 Produced by Streptomyces strain PM0324667. AMB Express 2011, 1, 42. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yu, L.; Komaki, H.; Oku, N.; Igarashi, Y. Absolute Configuration of NFAT-133, an Aromatic Polyketide with Immunosuppressive and Antidiabetic Activity from Actinomycetes. J. Antibiot. 2016, 69, 69–71. [Google Scholar] [CrossRef]
- Yi, H.; Li, M.; Huo, X.; Zeng, G.; Lai, C.; Huang, D.; An, Z.; Qin, L.; Liu, X.; Li, B.; et al. Recent Development of Advanced Biotechnology for Wastewater Treatment. Crit. Rev. Biotechnol. 2020, 40, 99–118. [Google Scholar] [CrossRef]
- Tkachuk, N.; Zelena, L. The Impact of Bacteria of The Genus Bacillus Upon the Biodamage/Biodegradation of Some Metals And Extensively Used Petroleum-Based Plastics. Corros. Mater. Degrad. 2021, 2, 531–553. [Google Scholar] [CrossRef]
- Suteu, D.; Zaharia, C. Sawdust as biosorbent for removal of dyes from wastewaters. Kinetic and thermodynamic study. Chem. Bull. Politeh. 2011, 56, 85–88. [Google Scholar]
- Suteu, D.; Zaharia, C.; Badeanu, M. Agriculture wastes used as sorbents for dyes removal from aqueous environments. Seed 2010, 8, 140–145. [Google Scholar]
- Suteu, D.; Bilba, D.; Zaharia, C.; Popescu, A. Removal of dyes from textile wastewater by sorption onto ligno-cellulosic materials. Sci. Study Res. 2008, 9, 293–302. [Google Scholar]
- Yadav, B.; Pandey, A.; Kumar, L.R.; Tyagi, R.D. Bioconversion of Waste (Water)/Residues to Bioplastics- A Circular Bioeconomy Approach. Bioresour. Technol. 2020, 298, 122584. [Google Scholar] [CrossRef] [PubMed]
- Babich, O.; Shevchenko, M.; Ivanova, S.; Pavsky, V.; Zimina, M.; Noskova, S.; Anohova, V.; Chupakhin, E.; Sukhikh, S. Antimicrobial Potential of Microorganisms Isolated from the Bottom Sediments of Lake Baikal. Antibiotics 2021, 10, 927. [Google Scholar] [CrossRef] [PubMed]
- Zemskaya, T.I.; Cabello-Yeves, P.J.; Pavlova, O.N.; Rodriguez-Valera, F. Microorganisms of Lake Baikal—The Deepest and Most Ancient Lake on Earth. Appl. Microbiol. Biotechnol. 2020, 104, 6079–6090. [Google Scholar] [CrossRef]
- Suslova, M.Y.; Pestunova, O.S.; Sukhanova, E.V.; Shtykova, Y.R.; Kostornova, T.Y.; Khanaev, I.V.; Sakirko, M.V.; Parfenova, V.V. Role of Cultured Microorganisms from Biofilms Formed on Rocky Substrates in the Lake Baikal Self-Purification System. Microbiology 2018, 87, 817–824. [Google Scholar] [CrossRef]
- Chauhan, P.S. Role of Various Bacterial Enzymes in Complete Depolymerization of Lignin: A Review. Biocat. Agric. Biotechnol. 2020, 23, 101498. [Google Scholar] [CrossRef]
- Vilanova, C.; Marín, M.; Baixeras, J.; Latorre, A.; Porcar, M. Selecting Microbial Strains from Pine Tree Resin: Biotechnological Applications from a Terpene World. PLoS ONE 2014, 9, e100740. [Google Scholar] [CrossRef] [Green Version]
- Chang, R.; Rohindra, D.; Lata, R.; Kuboyama, K.; Ougizawa, T. Development of Poly(ε-Caprolactone)/Pine Resin Blends: Study of Thermal, Mechanical, and Antimicrobial Properties: Development of Poly(ε-Caprolactone)/Pine Resin Blends. Polym. Eng. Sci. 2019, 59, E32–E41. [Google Scholar] [CrossRef]
- Sakoda, S.; Aisu, K.; Imagami, H.; Matsuda, Y. Comparison of Actinomycete Community Composition on the Surface and Inside of Japanese Black Pine (Pinus Thunbergii) Tree Roots Colonized by the Ectomycorrhizal Fungus Cenococcum Geophilum. Microb. Ecol. 2019, 77, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Satomi, M.; Fukui, Y.; Matsunobu, S.; Morifuku, Y.; Enokida, Y. Streptomyces Abietis Sp. Nov., a Cellulolytic Bacterium Isolated from Soil of a Pine Forest. Int. J. Syst. Evol. Microbiol. 2013, 63, 4754–4759. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ponpandian, L.N.; Kim, H.; Jeon, J.; Hwang, B.S.; Lee, S.K.; Park, S.C.; Bae, H. Distribution and Diversity of Bacterial Endophytes from Four Pinus Species and Their Efficacy as Biocontrol Agents for Devastating Pine Wood Nematodes. Sci. Rep. 2019, 9, 12461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakkar, C.S.; Kate, A.S.; Desai, D.C.; Ghosh, A.R.; Kulkarni-Almeida, A.A. NFAT-133 Increases Glucose Uptake in L6 Myotubes by Activating AMPK Pathway. Eur. J. Pharmacol. 2015, 769, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.P.; Huang, S.H.; Ting, Y.; Hsu, H.Y.; Cheng, K.C. Evaluation of Detoxified Sugarcane Bagasse Hydrolysate by Atmospheric Cold Plasma for Bacterial Cellulose Production. Int. J. Biol. Macromol. 2022, 204, 136–143. [Google Scholar] [CrossRef]
- Lin, S.P.; Kuo, T.C.; Wang, H.T.; Ting, Y.; Hsieh, C.W.; Chen, Y.K.; Cheng, K.C. Enhanced Bioethanol Production Using Atmospheric Cold Plasma-Assisted Detoxification of Sugarcane Bagasse Hydrolysate. Bioresour. Technol. 2020, 313, 123704. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MM + Pine Sawdust | MM + Birch Sawdust |
|---|---|---|
| Streptomyces sp. LPB2019K190-4 | + | + |
| Rhodococcus sp. LPB2019K201-3 | − | + |
| Microbacterium sp. LPB2019K198-2 | − | + |
| Strain | Nutrient Media | Experiment | Control | ||
|---|---|---|---|---|---|
| B. subtilis | P. putida | B. subtilis | P. putida | ||
| Streptomyces sp. LPB2019K190-4 | MM | + | − | − | − |
| MM + low content of pine sawdust | + | − | − | − | |
| MM + high content of pine sawdust | + | − | − | − | |
| MM | − | − | − | − | |
| MM + low content of birch sawdust | − | − | − | − | |
| MM + high content of birch sawdust | − | − | − | − | |
| Rhodococcus sp. LPB2019K201-3 | MM | − | − | − | − |
| MM + low content of birch sawdust | − | − | − | − | |
| MM + high content of birch sawdust | − | − | − | − | |
| Microbacterium sp. LPB2019K198-2 | MM | − | − | − | − |
| MM + low content of birch sawdust | − | + | − | − | |
| MM + high content of birch sawdust | − | + | − | − | |
| № | Retention Time (min) | Mass. [M + H] m/z = [M] −1.0079 | Adducts | CRC-Code | Natural Product Name | Synonym of Natural Product | Mass | Δ[M] | ppm | Importance | Known Biological Source | References |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2.6 | 318.149 | [M + Na] | NRR69-H | N2-[5-(4-Aminophenyl)-2,4-pentadienoyl] glutamine | − | 317.138 | 0.0032 | 10.2 | − | Streptomyces sp. Tu 3946 | [40] |
| 2 | 5.2 | 318.207 | [M + Na] | OOJ07-P | Glaciapyrrol B | − | 317.199 | 0.0001 | 0.3 | Antibacterial activity | Streptomyces sp. NPS008187, isolated from sea water | [41] |
| 3 | 6.5 | 277.18 | [M + Na] | NNX36-X | NFAT 133 | 4-hydroxy-5-[2-(3-hydroxy-1-propenyl)-4-methylphenyl]-3-methyl-2-hexanone | 276.73 | 0.0008 | 3.1 | Blocks NFAT-dependent transcription and shows immunosuppressant props.; exhibits antidiabetic props. | Streptomyces karnatakensis | [42,43,44] |
| 4 | 14.4 | 295.188 | [M + NH4] | GNH63-C | Octahydro-7α-methyl-1-(1-methyl-2-oxopropyl)-5-oxo-1H-indene-4-propanoic acid | 3-[7a-Methyl-5-oxo-1-(1-methyl-2-oxopropyl)-3aH-hexahydroindan-4-yl] propionic acid | 294.18 | 0.0030 | 10.2 | Antifungal properties | Rhodococcus sp. | [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereliaeva, E.V.; Dmitrieva, M.E.; Morgunova, M.M.; Belyshenko, A.Y.; Imidoeva, N.A.; Ostyak, A.S.; Axenov-Gribanov, D.V. The Use of Baikal Psychrophilic Actinobacteria for Synthesis of Biologically Active Natural Products from Sawdust Waste. Fermentation 2022, 8, 213. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050213
Pereliaeva EV, Dmitrieva ME, Morgunova MM, Belyshenko AY, Imidoeva NA, Ostyak AS, Axenov-Gribanov DV. The Use of Baikal Psychrophilic Actinobacteria for Synthesis of Biologically Active Natural Products from Sawdust Waste. Fermentation. 2022; 8(5):213. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050213
Chicago/Turabian StylePereliaeva, Ekaterina V., Maria E. Dmitrieva, Maria M. Morgunova, Alexander Y. Belyshenko, Natalia A. Imidoeva, Alexander S. Ostyak, and Denis V. Axenov-Gribanov. 2022. "The Use of Baikal Psychrophilic Actinobacteria for Synthesis of Biologically Active Natural Products from Sawdust Waste" Fermentation 8, no. 5: 213. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050213