
Analysis of the Composition of Substrate for Industrial Fermentation of Agaricus bisporus Based on Secondary and Tertiary Fermentation Mode Composition Analysis of Industrial Fermentation Substrates of A. bisporus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Secondary Fermentation Method
2.2. Tertiary Fermentation Method
2.3. Experimental Design
2.4. GC–MS Analysis
2.5. Data Analysis
3. Results and Discussion
3.1. Analysis and Results
3.1.1. Identification of Metabolites (GC–MS Data Analysis)
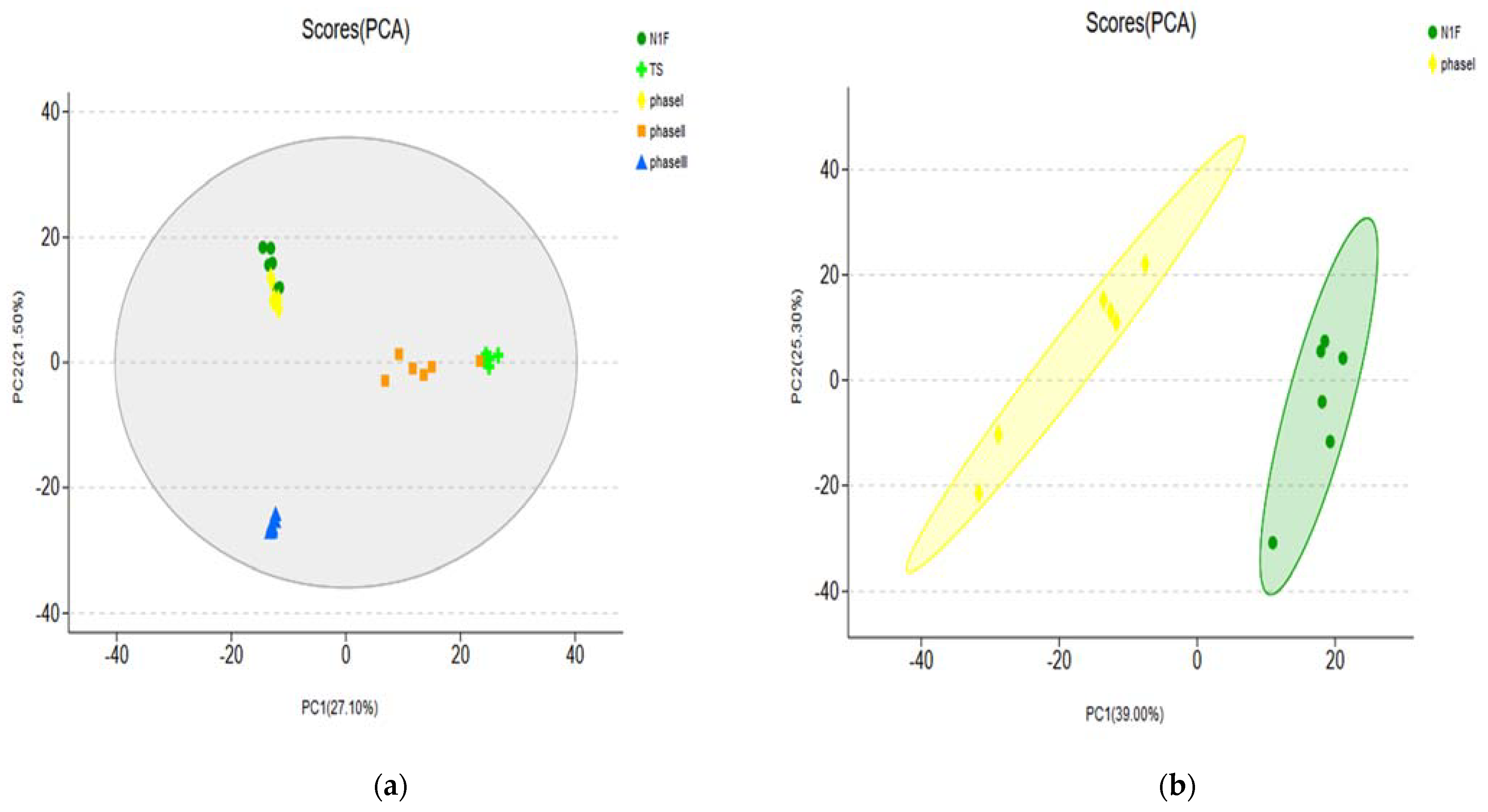
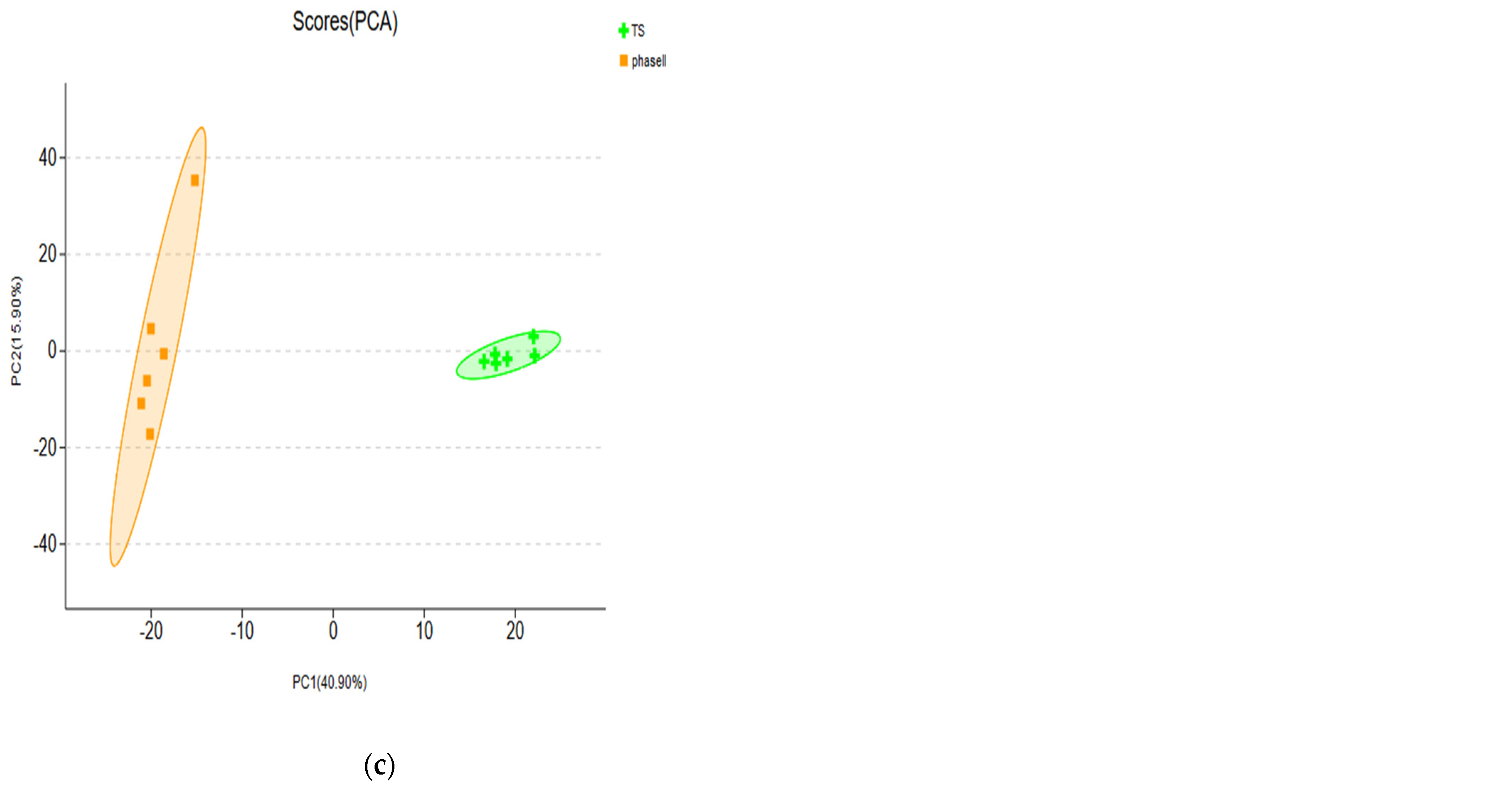
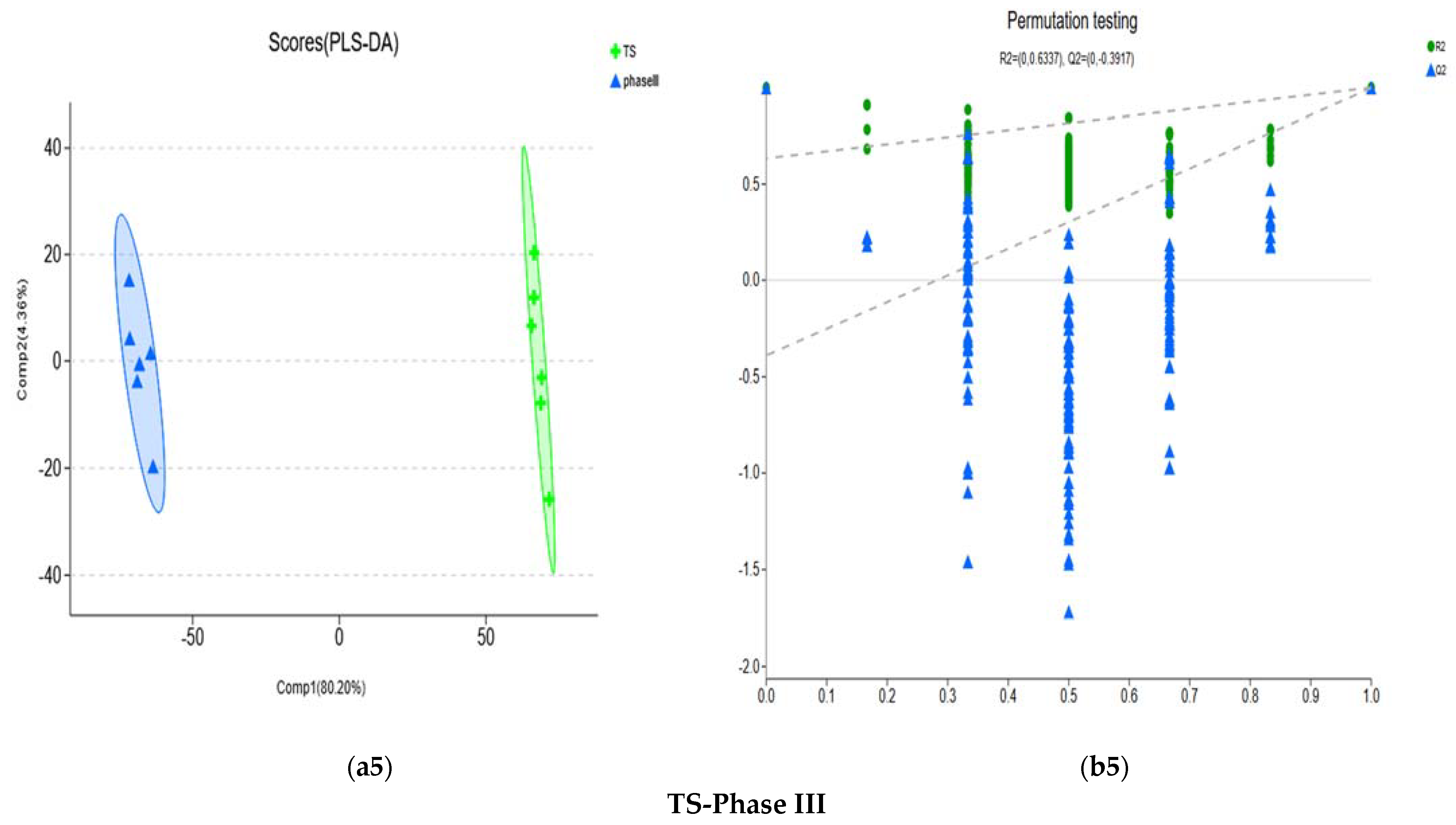
3.1.2. PCA of Metabolites
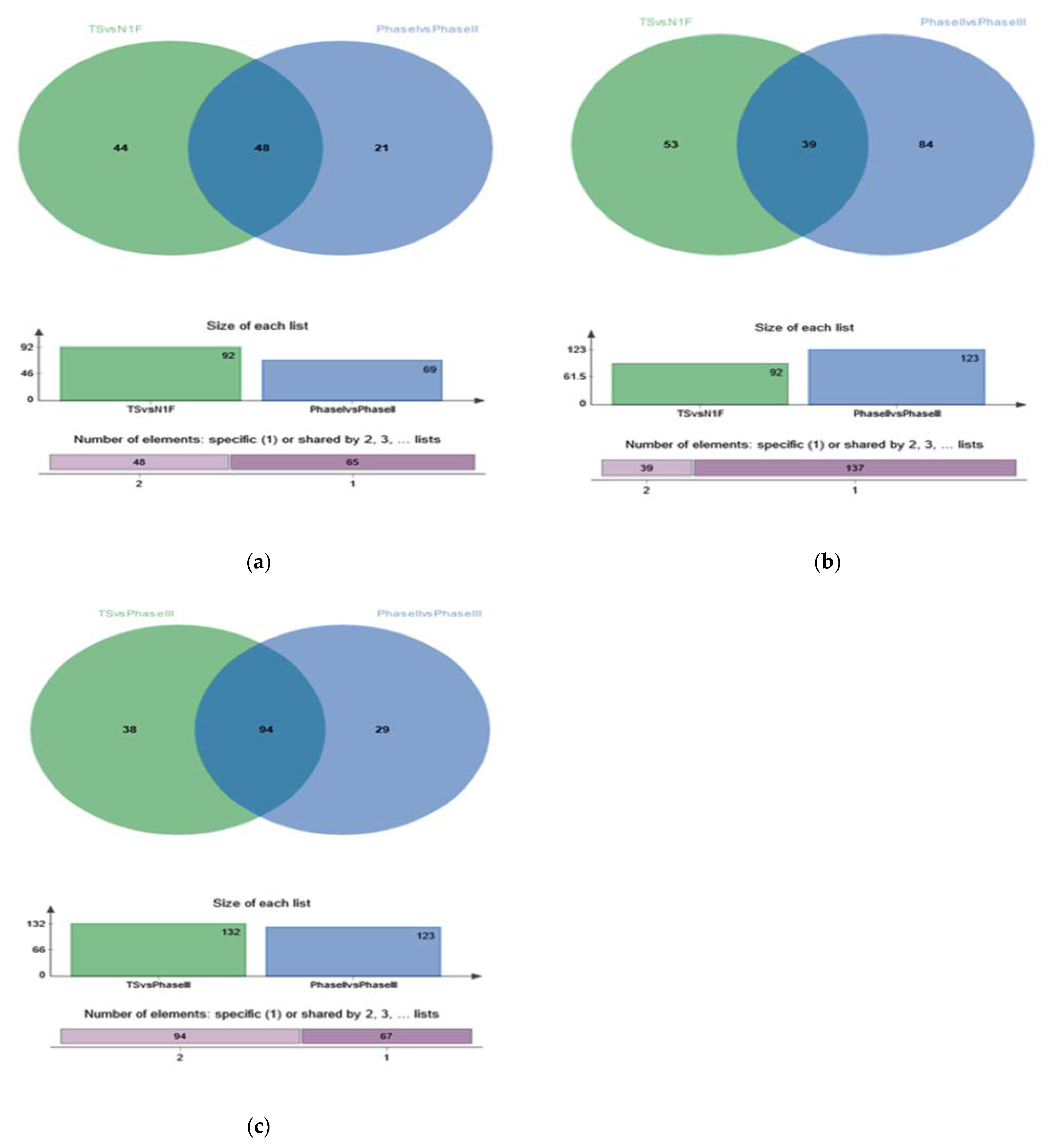
3.1.3. Differential Metabolite Analysis
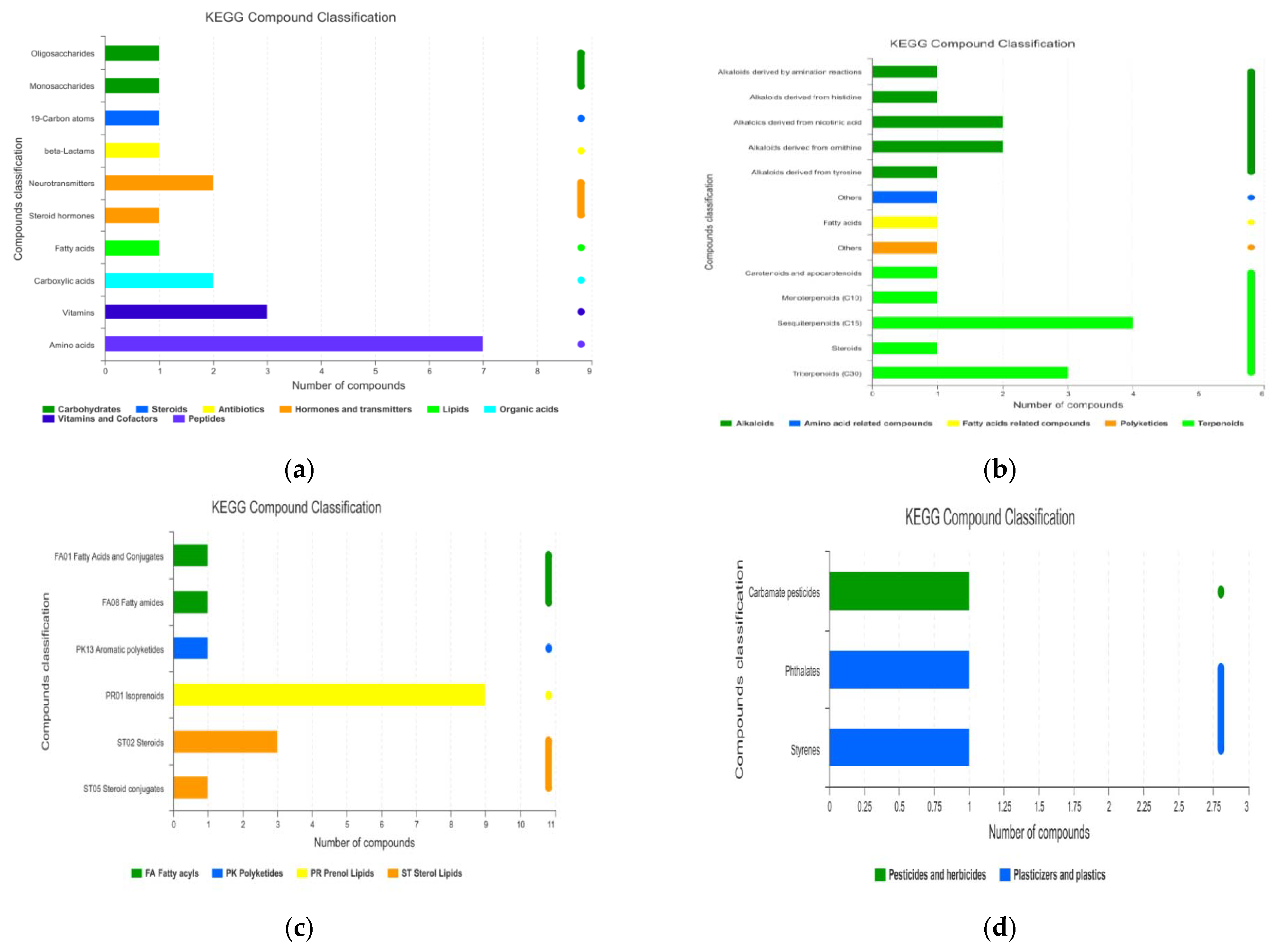
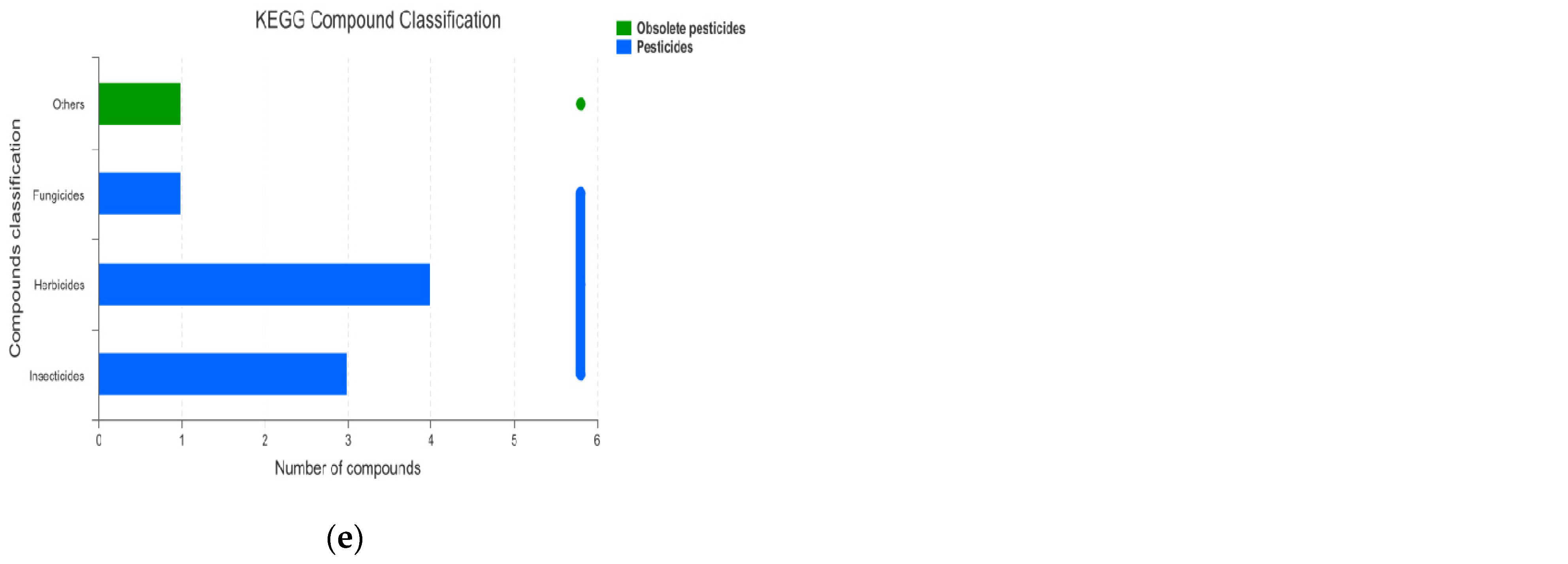
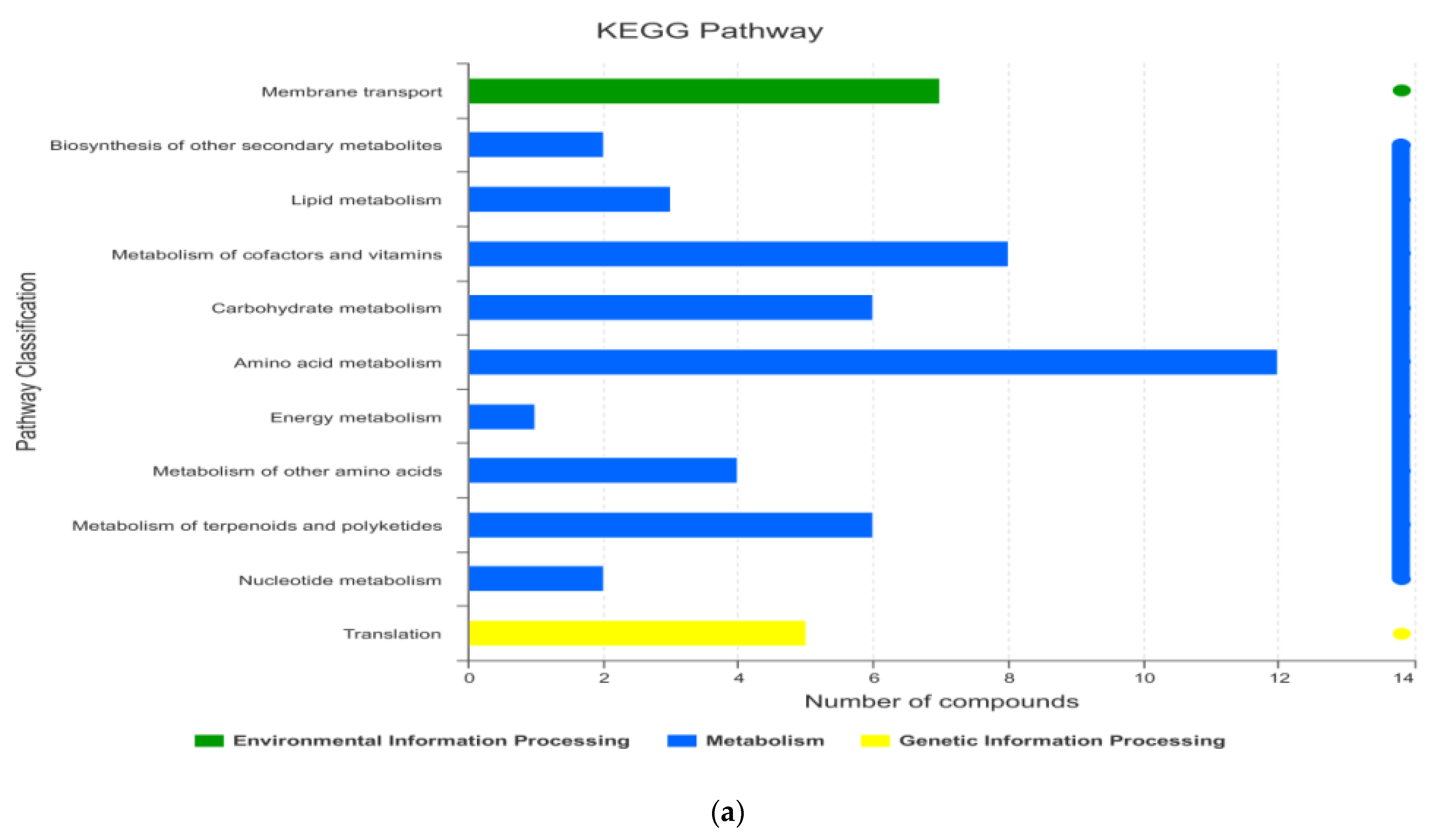
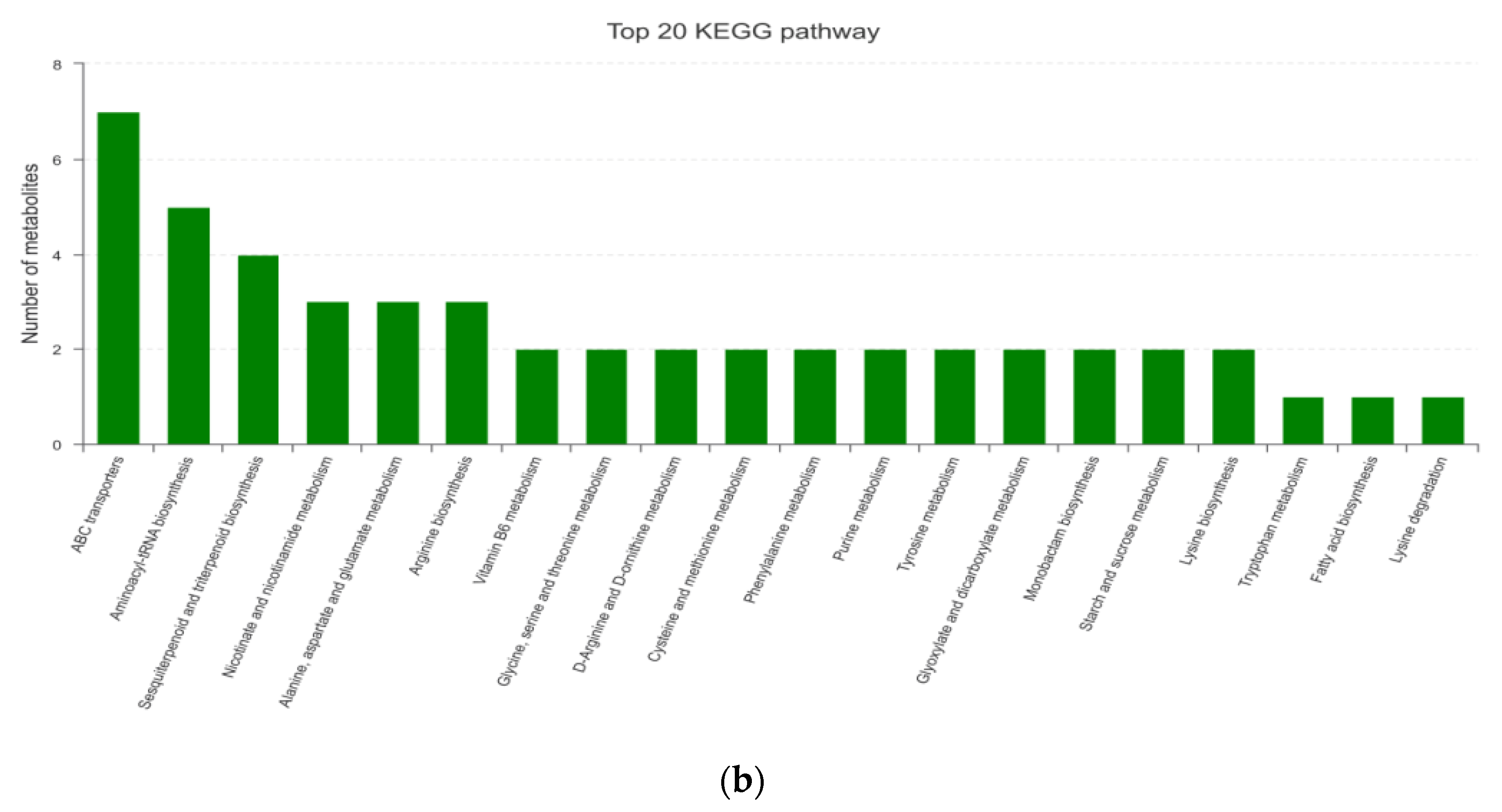
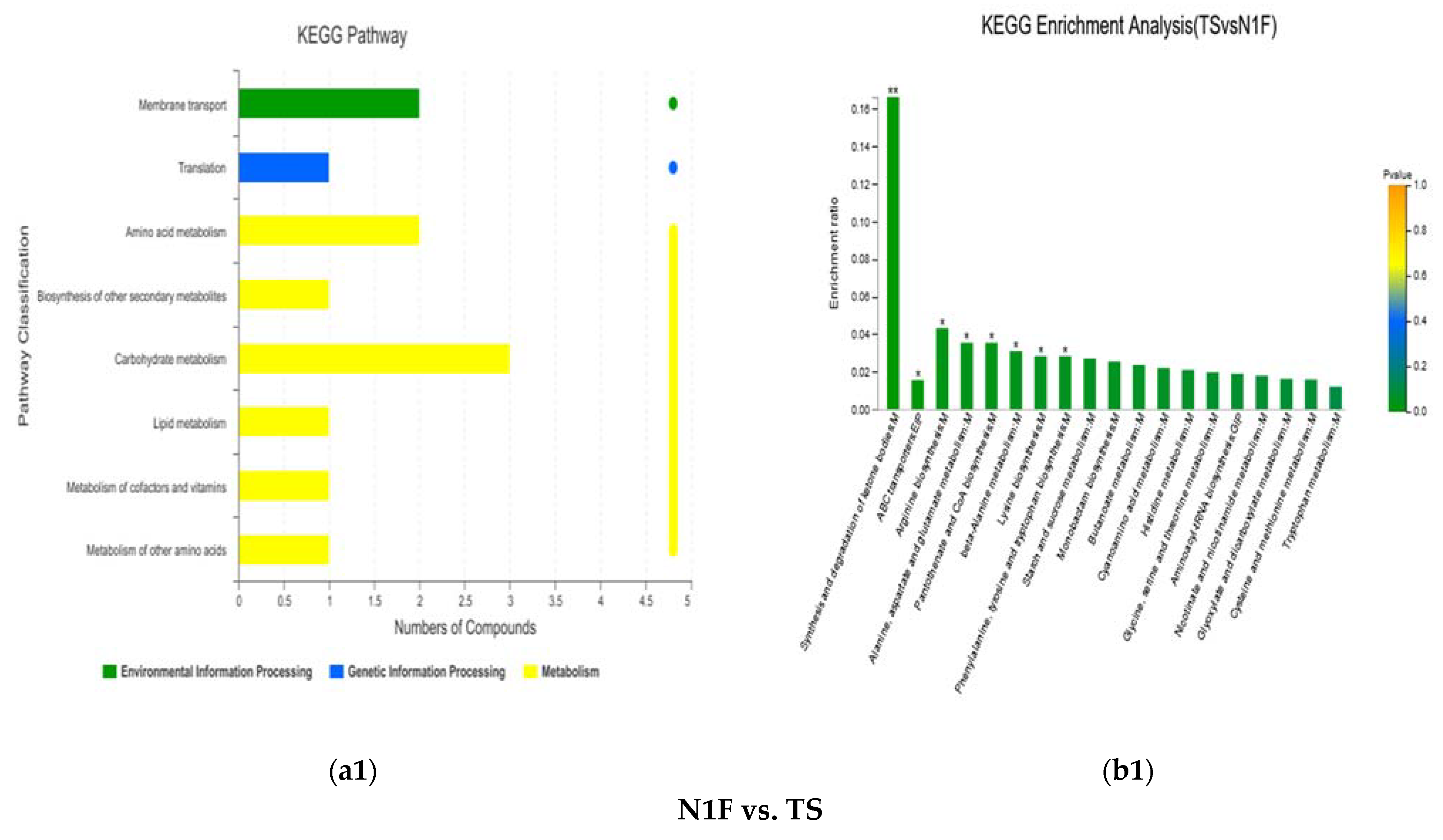
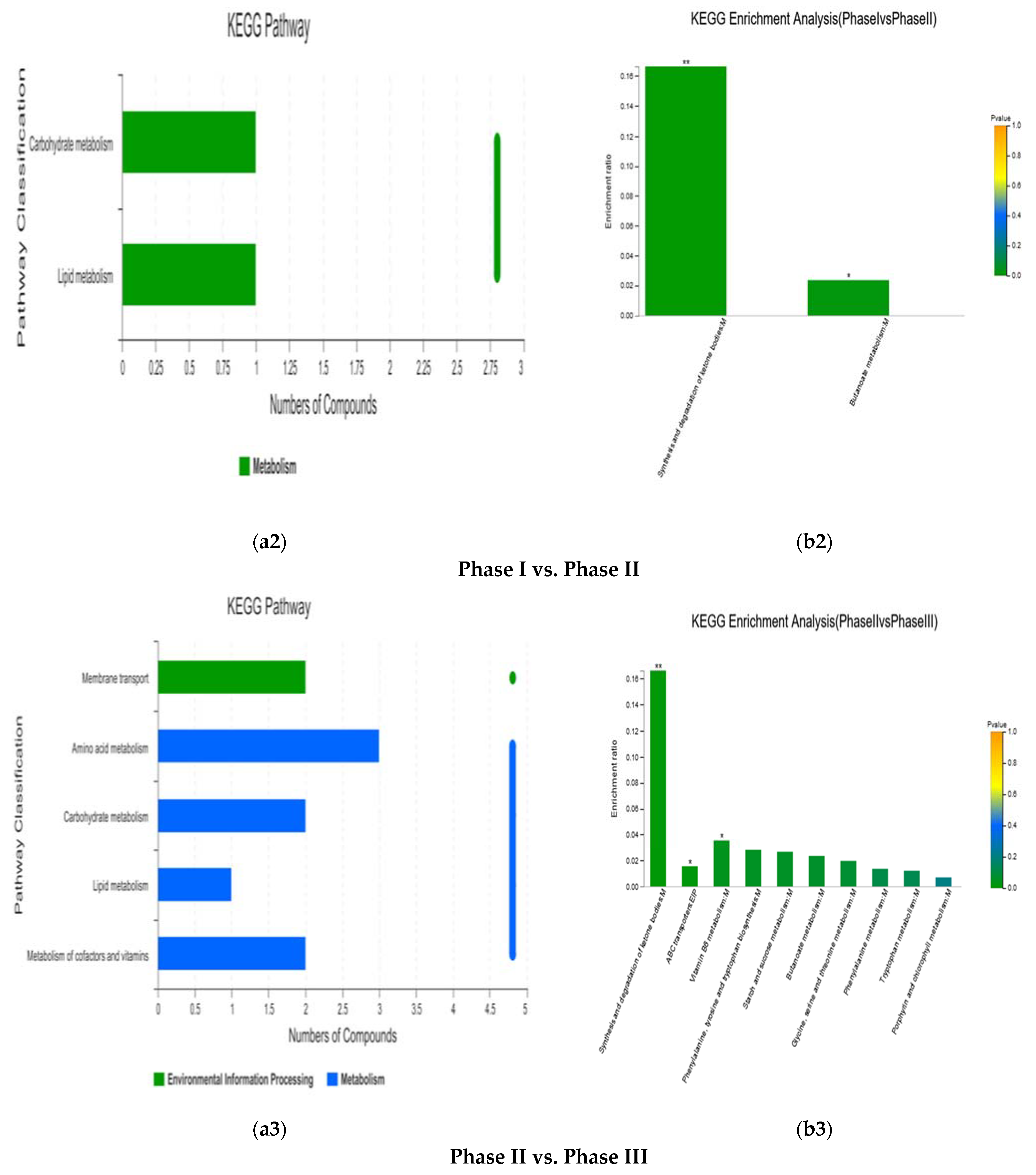
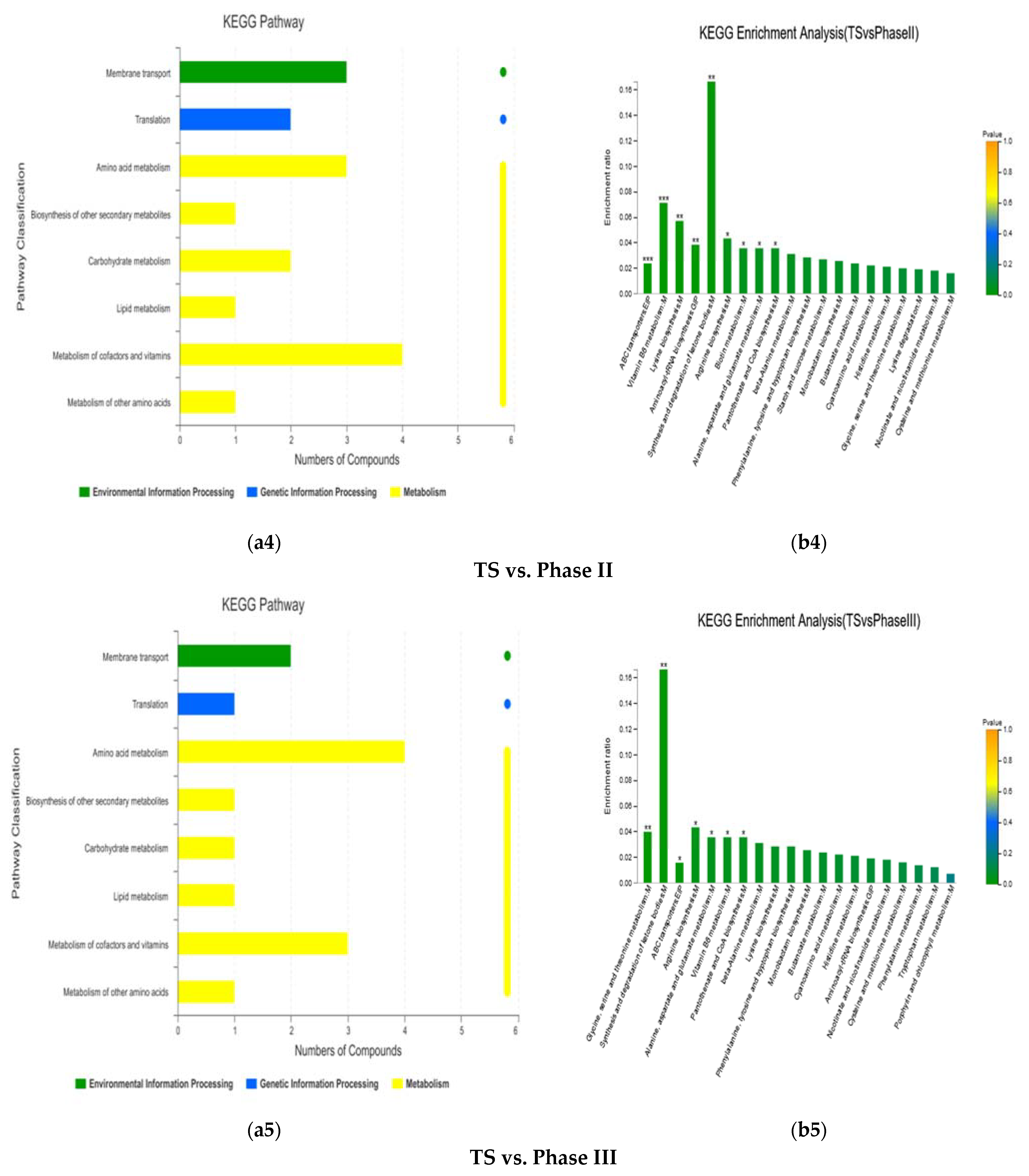
3.1.4. KEGG Enrichment Analysis and Main Differential Metabolites
3.2. Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.Q.; Geng, W.; Shen, Y.Q.; Wang, Y.L.; Dai, Y.C. Edible mushroom cultivation for food security and rural development in China: Bio-innovation, technological dissemination and marketing. Sustainability 2014, 6, 2961–2973. [Google Scholar] [CrossRef] [Green Version]
- Royse, D.J.; Singh, M. A Global Perspective on the High Five: Agaricus, Pleurotus, Lentinula, Auricularia & Flammulina. In Proceedings of the 8th International Conference on Mushroom Biology & Mushroom Products, New Delhi, India, 19–22 November 2014; CABI: Wallingford, UK, 2014; pp. 1–6. [Google Scholar]
- Van Griensven, L.J.L.D. The cultivation of mushrooms. Nature 1988, 142, 416. [Google Scholar] [CrossRef]
- Sharma, H.S.; Kilpatrick, M. Mushroom (Agaricus bisporus) compost quality factors for predicting potential yield of fruiting bodies. Can. J. Microbiol. 2000, 46, 515–519. [Google Scholar] [CrossRef] [PubMed]
- De Andrade, M.C.; de Jesus, J.P.; Vieira, F.R.; Viana, S.R.; Spoto, M.H.; de Almeida Minhoni, M.T. Dynamics of the chemical composition and productivity of composts for the cultivation of Agaricus bisporus strains. Braz. J. Microbiol. 2013, 44, 1139–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zied, D.C.; Pardo-González, J.E.; Minhoni, M.T.A.; Pardo-Giménez, A. A reliable quality index for mushroom cultivation. J. Agric. Sci. 2011, 3, 50–61. [Google Scholar] [CrossRef]
- Royse, D.J.; Chalupa, W. Effects of spawn, supplement and phase II compost additions and time of re-casing second break compost on mushroom (Agaricus bisporus) yield and biological efficiency. Bioresour. Technol. 2009, 100, 5277–5282. [Google Scholar] [CrossRef]
- Zhang, H.-L.; Wei, J.-K.; Wang, Q.-H.; Yang, R.; Gao, X.-J.; Sang, Y.-X.; Cai, P.-P.; Zhang, G.-Q.; Chen, Q.-J. Lignocellulose utilization and bacterial communities of millet straw based mushroom (Agaricus bisporus) production. Sci. Rep. 2019, 9, 1151. [Google Scholar] [CrossRef] [Green Version]
- Altieri, R.; Esposito, A.; Parati, F.; Lobianco, A.; Pepi, M. Performance of olive mill solid waste as a constituent of the substrate in commercial cultivation of Agaricus bisporus. Int. Biodeterior. Biodegrad. 2009, 63, 993–997. [Google Scholar] [CrossRef]
- Vos, A.M.; Heijboer, A.; Boschker, H.T.S.; Bonnet, B.; Lugones, L.G.; Wösten, H.A.B. Microbial biomass in compost during colonization of Agaricus bisporus. AMB Express 2017, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhong, Y.; Yang, S.; Zhang, W.; Xu, M.; Ma, A.; Zhuang, G.; Chen, G.; Liu, W. Diversity and dynamics of the microbial community on decomposing wheat straw during mushroom compost production. Bioresour. Technol. 2014, 170, 183–195. [Google Scholar] [CrossRef]
- Wang, L.; Mao, J.; Zhao, H.; Li, M.; Wei, Q.; Zhou, Y.; Shao, H. Comparison of characterization and microbial communities in rice straw- and wheat straw-based compost for Agaricus bisporus production. J. Ind. Microbiol. Biotechnol. 2016, 43, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Vieira, F.R.; Pecchia, J.A.; Segato, F.; Polikarpov, I. Exploring oyster mushroom (Pleurotus ostreatus) substrate preparation by varying phase I composting time: Changes in bacterial communities and physicochemical composition of biomass impacting mushroom yields. J. Appl. Microbiol. 2019, 126, 931–944. [Google Scholar] [CrossRef] [PubMed]
- Kertesz, M.A.; Thai, M. Compost bacteria and fungi that influence growth and development of Agaricus bisporus and other commercial mushrooms. Appl. Microbiol. Biotechnol. 2018, 102, 1639–1650. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Yuan, X.; Wang, S.; Sun, F.; Hou, Z.; Hu, Q.; Zhai, L.; Cui, Z.; Zou, Y. Methane production and characteristics of the microbial community in the co-digestion of spent mushroom substrate with dairy manure. Bioresour. Technol. 2018, 250, 611–620. [Google Scholar] [CrossRef]
- Vieira, F.R.; de Andrade, M.C. Optimization of substrate preparation for oyster mushroom (Pleurotus ostreatus) cultivation by studying different raw materials and substrate preparation conditions (composting: Phases I and II). World J. Microbiol. Biotechnol. 2016, 32, 190. [Google Scholar] [CrossRef] [PubMed]
- Jeznabadi, E.K.; Jafarpour, M.; Eghbalsaied, S. King oyster mushroom production using various sources of agricultural wastes in Iran. Int. J. Recycl. Org. Waste Agric. 2016, 5, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.T.; Gong, G.Z.; Li, Q.; Weng, H.D.; Cai, K.D. Stack fermentation technology of Agdcus bisporus compost. Edible Fungi China 2009, 28, 29–30, 35. [Google Scholar]
- Fiehn, O.; Kopka, J.; Dörmann, P.; Altmann, T.; Trethewey, R.N.; Willmitzer, L. Metabolite profiling for plant functional genomics. Nat. Biotechnol. 2000, 18, 1157–1161. [Google Scholar] [CrossRef]
- Trivedi, D.K.; Hollywood, K.A.; Goodacre, R. Metabolomics for the masses: The future of metabolomics in a personalized world. New Horiz. Transl. Med. 2017, 3, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-M.; Wu, W.-W.; Li, G.-K. A GC—MS study of the volatile organic composition of straw and oyster mushrooms during maturity and its relation to antioxidant activity. J. Chromatogr. Sci. 2008, 46, 690–696. [Google Scholar] [CrossRef] [Green Version]
- O’Gorman, A.; Barry-Ryan, C.; Frías, J. Evaluation and identification of markers of damage in mushrooms (Agaricus bisporus) postharvest using a GC/MS metabolic profiling approach. Metabolomics 2011, 8, 120–132. [Google Scholar] [CrossRef]
- Qiu, Z.; Wu, X.; Zhang, J.; Huang, C. High-temperature induced changes of extracellular metabolites in Pleurotus ostreatus and their positive effects on the growth of Trichoderma asperellum. Front. Microbiol. 2018, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, F.; Huang, F.; Xie, G.; Wei, R.; Chen, T.; Liu, J.; Zhao, A.; Jia, W. Metabolomics analysis reveals variation in Schisandra Chinensis cetabolites from different origins. J. Sep. Sci. 2014, 37, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Vieira, F.R.; Pecchia, J.A. An exploration into the bacterial community under different pasteurization conditions during substrate preparation (composting-phase II) for Agaricus bisporus cultivation. Microb. Ecol. 2018, 75, 318–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurak, E.; Punt, A.M.; Arts, W.; Kabel, M.A.; Gruppen, H. Fate of carbohydrates and lignin during composting and mycelium growth of Agaricus bisporus on wheat straw based compost. PLoS ONE 2015, 10, e0138909. [Google Scholar] [CrossRef] [PubMed]
- Straatsma, G.; Gerrits, J.P.G.; Thissen, J.T.N.M.; Amsing, J.G.M.; Loeffen, H.; Van Griensven, L.J.L.D. Adjustment of the composting process for mushroom cultivation based on initial substrate composition. Bioresour. Technol. 2000, 72, 67–74. [Google Scholar] [CrossRef]
- Knowles, J.R. The mechanism of biotin-dependent enzymes. Annu. Rev. Biochem. 1989, 58, 195–221. [Google Scholar] [CrossRef]
- Hymes, J.; Wolf, B. Biotinidase and its roles in biotin metabolism. Clin. Chim. Acta 1996, 255, 1–11. [Google Scholar] [CrossRef]
- Tang, Q.; Li, X.; Zou, T.; Zhang, H.; Wang, Y.; Gao, R.; Li, Z.; He, J.; Feng, Y. Mycobacterium smegmatis BioQ defines a new regulatory network for biotin metabolism. Mol. Microbiol. 2014, 94, 1006–1023. [Google Scholar] [CrossRef]
- Fitzpatrick, T.B.; Amrhein, N.; Kappes, B.; Macheroux, P.; Tews, I.; Raschle, T. Two independent routes of de novo vitamin B6 biosynthesis: Not that different after all. Biochem. J. 2007, 407, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Colinas, M.; Fitzpatrick, T.B. Interaction between vitamin B6 metabolism, nitrogen metabolism and autoimmunity. Plant Signal. Behav. 2016, 11, e1161876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selhub, J. Homocysteine metabolism. Annu. Rev. Nutr. 1999, 19, 217–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, K.L.; Olson, B.; Bakun, P.; Dallal, G.E.; Selhub, J.; Rosenberg, I.H. Breakfast cereal fortified with folic acid, vitamin B-6, and vitamin B-12 increases vitamin concentrations and reduces homocysteine concentrations: A randomized trial. Am. J. Clin. Nutr. 2004, 79, 805–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toney, M.D. Reaction specificity in pyridoxal phosphate enzymes. Arch. Biochem. Biophys. 2005, 433, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Manegold, C.; Hoffmann, G.F.; Degen, I.; Ikonomidou, H.; Knust, A.; Laass, M.W.; Pritsch, M.; Wilichowski, E.; Hörster, F. Aromatic L-amino acid decarboxylase deficiency: Clinical features, drug therapy and follow-up. J. Inherit. Metab. Dis. 2009, 32, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Wang, Z.; Cheng, X.; Gao, J.; Zhang, Z.; Wang, L. 5-Aminolevulinic acid promotes anthocyanin accumulation in Fuji apples. Plant Growth Regul. 2013, 69, 295–303. [Google Scholar] [CrossRef]
- Akram, N.A.; Ashraf, M. Regulation in plant stress tolerance by a potential plant growth regulator, 5-aminolevulinic acid. J. Plant Growth Regul. 2013, 32, 663–679. [Google Scholar] [CrossRef]
- McCormick, D.B.; Chen, H. Update on interconversions of vitamin B-6 with its coenzyme. J. Nutr. 1999, 129, 325–327. [Google Scholar] [CrossRef]
- Carrasco, J.; Preston, G.M. Growing edible mushrooms: A conversation between bacteria and fungi. Environ. Microbiol. 2020, 22, 858–872. [Google Scholar] [CrossRef] [Green Version]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
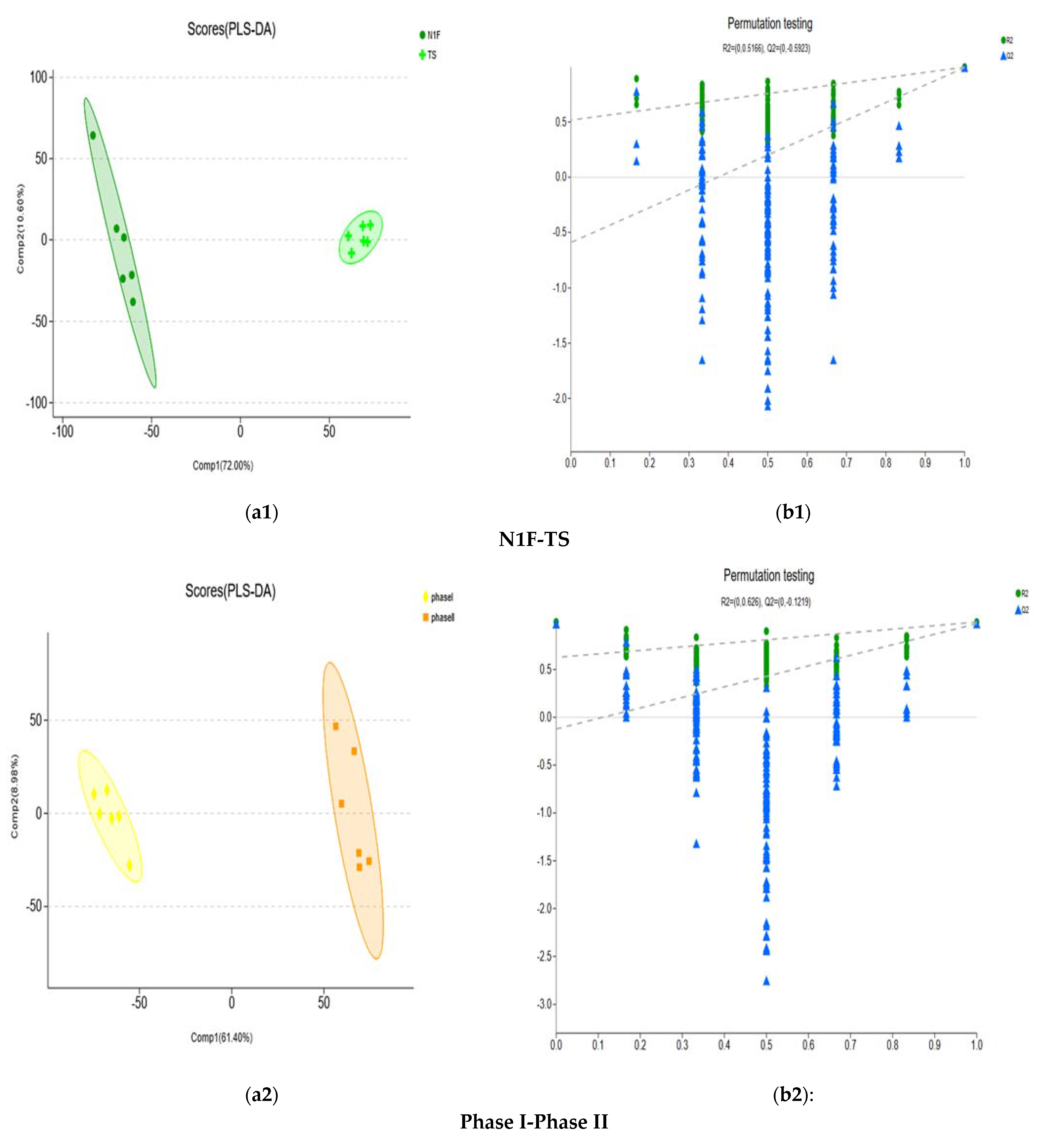
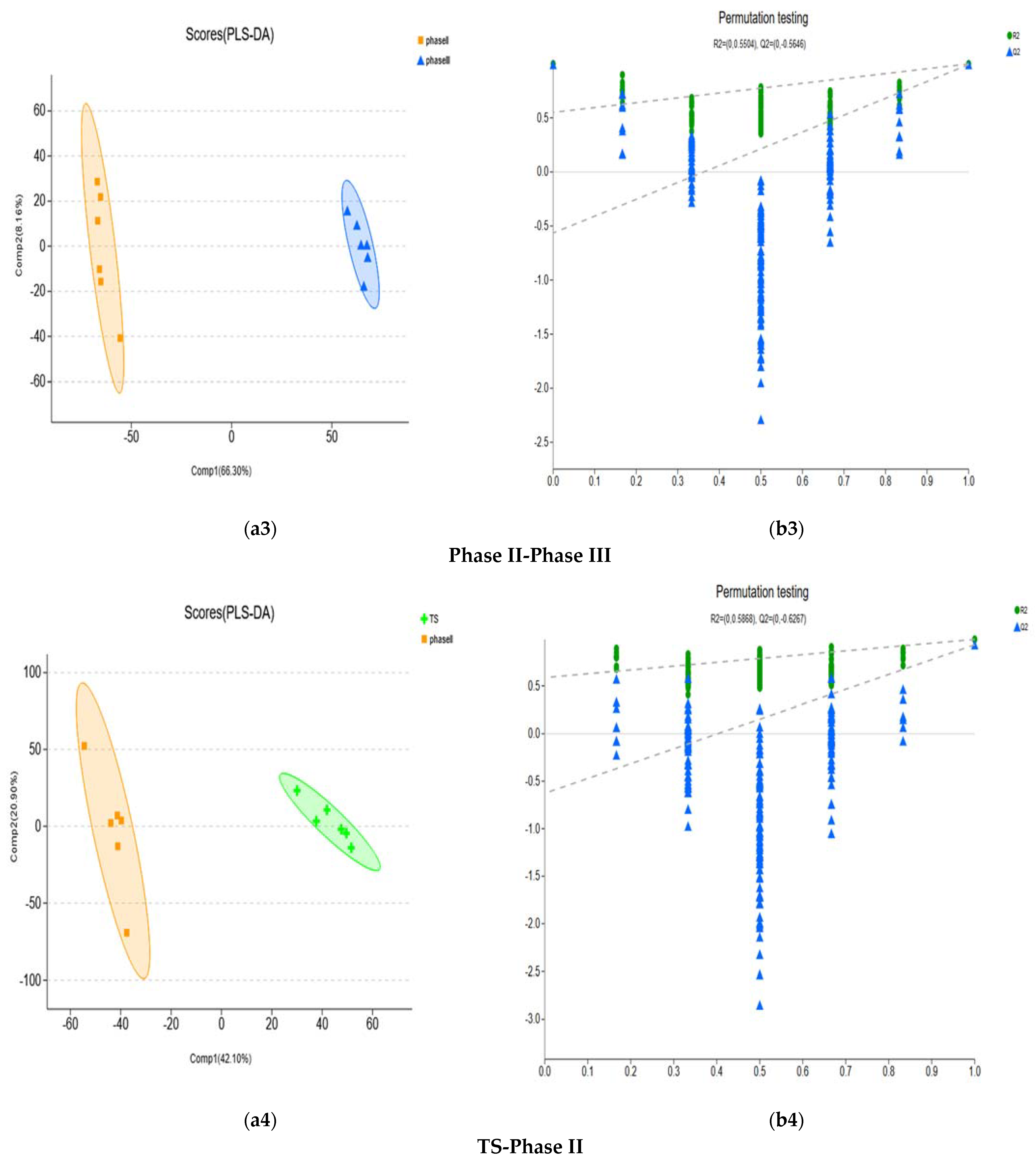
| Metabolic Set | R2X (cum) | R2Y (cum) | Q2 (cum) |
|---|---|---|---|
| N1F-TS | 0.72 | 0.992 | 0.99 |
| Phase I-Phase II | 0.614 | 0.991 | 0.98 |
| Phase II-Phase III | 0.663 | 0.997 | 0.991 |
| TS-Phase II | 0.421 | 0.977 | 0.91 |
| TS-Phase III | 0.802 | 0.998 | 0.997 |
| Metabolic Set | Different Metabolite | VIP_PLS-DA | FC(Y/X) | p-Value | KEGG Compound First Category | KEGG Compound Second Category | KEGG Pathway Description |
|---|---|---|---|---|---|---|---|
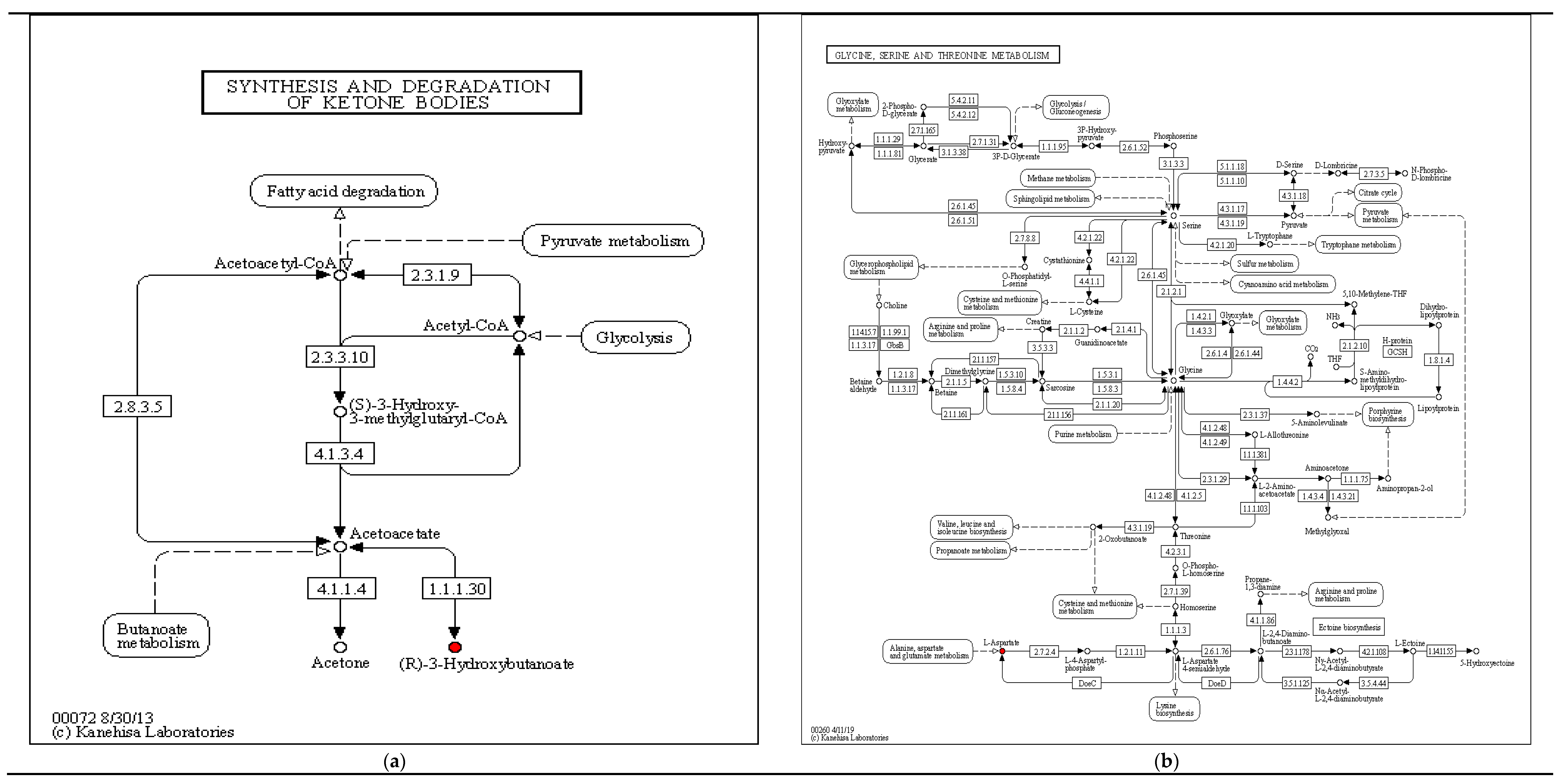
| N1F vs. TS | (R)-3-Hydroxybutyric acid | 2.6707 | 73.8734 | 6.59 × 10−5 | Organic acids | Carboxylic acids | Synthesis and degradation of ketone bodies; |
| Indole | 2.9484 | 1.8832 | 2.78 × 10−4 | - | - | Phenylalanine, tyrosine and tryptophan biosynthesis | |
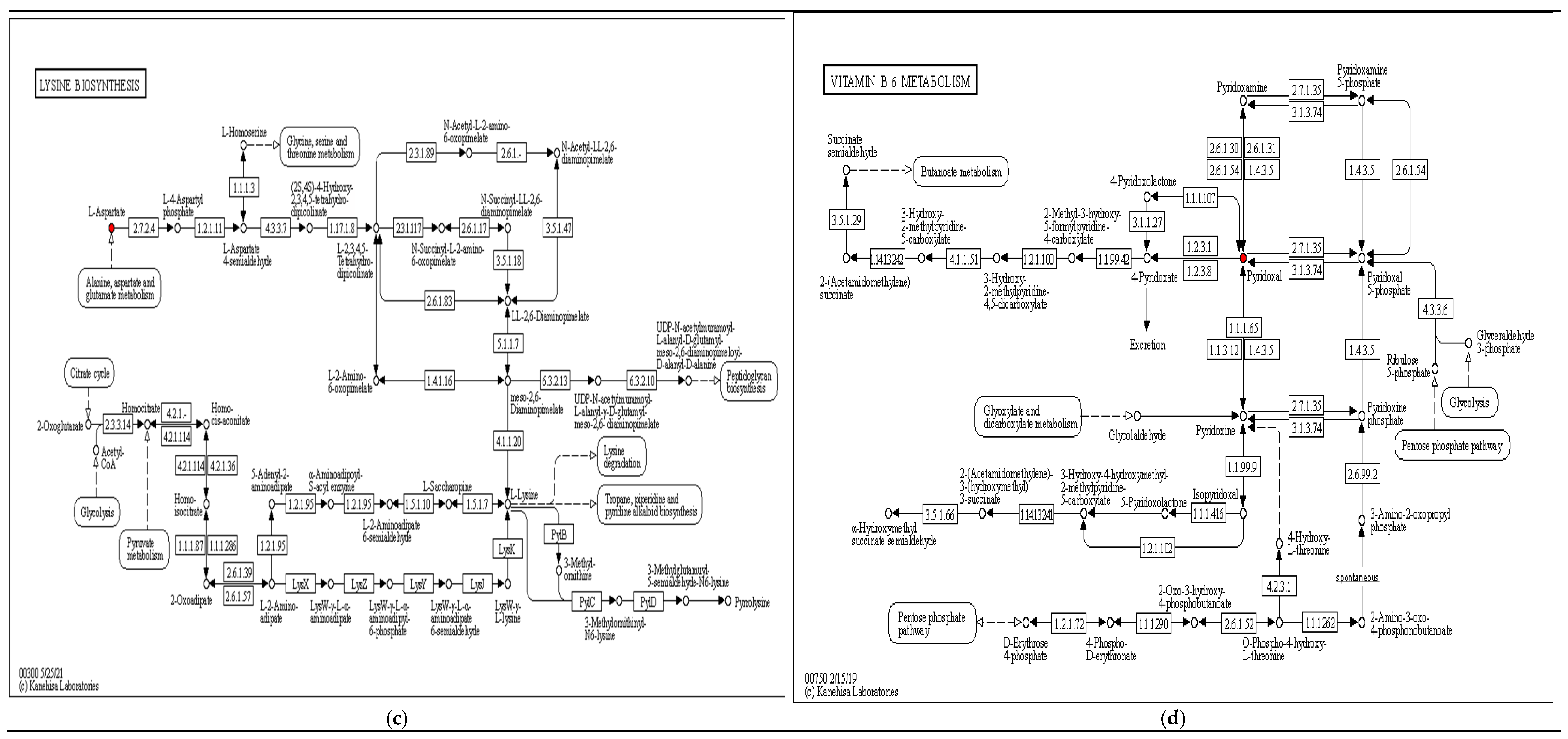
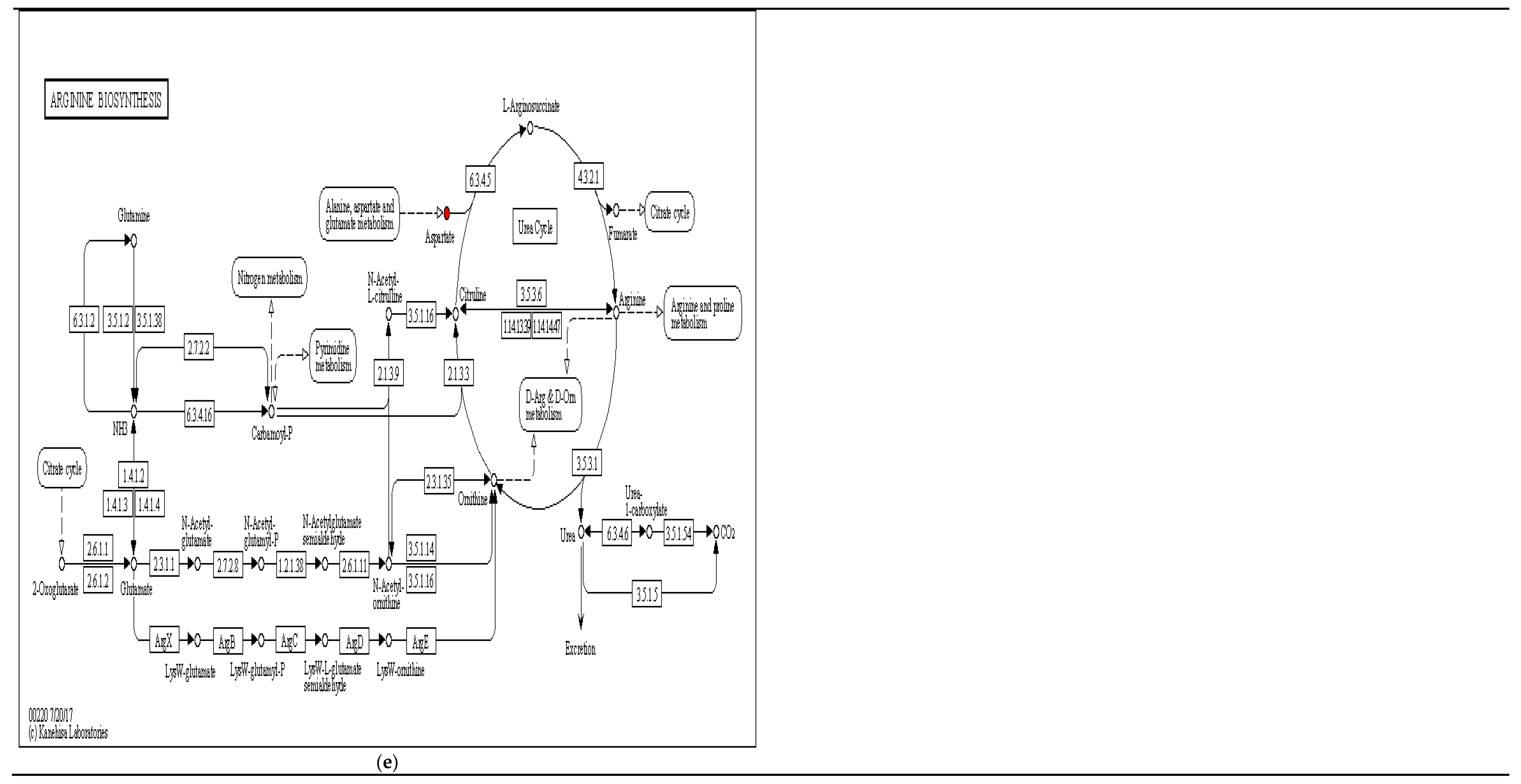
| L-Aspartic acid | 1.1376 | 17.3029 | 4.85 × 10−5 | Peptides; Hormones and transmitters | Amino acids; Neurotransmitters | Lysine biosynthesis; beta-Alanine metabolism; Alanine, aspartate and glutamate metabolism; Pantothenate and CoA biosynthesis; Arginine biosynthesis; ABC transporters | |
| Maltose | 1.0904 | 0.3311 | 1.27 × 10−5 | Carbohydrates | Oligosaccharides | ABC transporters | |
| Phase I vs. Phase II | (R)-3-Hydroxybutyric acid | 1.4858 | 0.0645 | 1.514 × 10−3 | Organic acids | Carboxylic acids | Synthesis and degradation of ketone bodies; Butanoate metabolism |
| Phase II vs. Phase III | (R)-3-Hydroxybutyric acid | 1.626 | 16.8538 | 5.85 × 10−4 | Organic acids | Carboxylic acids | Synthesis and degradation of ketone bodies |
| 5-Aminolevulinic acid | 1.9879 | 7.1494 | 9.28 × 10−8 | - | - | ABC transporters | |
| Maltose | 1.0102 | 0.3005 | 1.178 × 10−2 | Carbohydrates | Oligosaccharides | ABC transporters | |
| Pyridoxal | 1.5561 | 0.1447 | 6.98 × 10−4 | Vitamins and Cofactors | Vitamins | Vitamin B6 metabolism | |
| TS vs. Phase II | (R)-3-Hydroxybutyric acid | 4.1648 | 45.0585 | 7.14 × 10−5 | Organic acids | Carboxylic acids | Synthesis and degradation of ketone bodies |
| L-Aspartic acid | 1.7843 | 17.6814 | 4.8 × 10−5 | Peptides; Hormones and transmitters | Amino acids; Neurotransmitters | Arginine biosynthesis; Alanine, aspartate and glutamate metabolism; Pantothenate and CoA biosynthesis; Lysine biosynthesis; Aminoacyl-tRNA biosynthesis; ABC transporters | |
| L-Lysine | 1.0874 | 5.2084 | 3.31 × 10−4 | Peptides | Amino acids | Biotin metabolism; Lysine biosynthesis; Aminoacyl-tRNA biosynthesis; ABC transporters | |
| Maltose | 1.4443 | 0.3362 | 1.729 × 10−2 | Carbohydrates | Oligosaccharides | ABC transporters | |
| Pyridoxal | 4.1648 | 45.0585 | 7.14 × 10−5 | Vitamins and Cofactors | Vitamins | Vitamin B6 metabolism | |
| Pyridoxamine 5-phosphate | 1.0562 | 0.4609 | 5.03 × 10−5 | - | - | Vitamin B6 metabolism | |
| TS vs. Phase III | (R)-3-Hydroxybutyric acid | 1.9603 | 0.374 | 3.762 × 10−3 | Organic acids | Carboxylic acids | Synthesis and degradation of ketone bodies |
| 5-Aminolevulinic acid | 1.9642 | 13.1007 | 1.56 × 10−9 | - | - | Glycine, serine and threonine metabolism; ABC transporters | |
| L-Aspartic acid | 1.042 | 0.1787 | 2.12 × 10−4 | Peptides; Hormones and transmitters | Amino acids; Neurotransmitters | Alanine, aspartate and glutamate metabolism; Pantothenate and CoA biosynthesis; Arginine biosynthesis; Glycine, serine and threonine metabolism; ABC transporters | |
| Pyridoxal | 2.4917 | 0.0638 | 3.5 × 10−7 | Vitamins and Cofactors | Vitamins | Vitamin B6 metabolism |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Juan, J.; Wang, Q.; Gao, Z.; Xiao, T.; Chen, H.; Zhang, J.; Song, X.; Huang, J. Analysis of the Composition of Substrate for Industrial Fermentation of Agaricus bisporus Based on Secondary and Tertiary Fermentation Mode Composition Analysis of Industrial Fermentation Substrates of A. bisporus. Fermentation 2022, 8, 222. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050222
Juan J, Wang Q, Gao Z, Xiao T, Chen H, Zhang J, Song X, Huang J. Analysis of the Composition of Substrate for Industrial Fermentation of Agaricus bisporus Based on Secondary and Tertiary Fermentation Mode Composition Analysis of Industrial Fermentation Substrates of A. bisporus. Fermentation. 2022; 8(5):222. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050222
Chicago/Turabian StyleJuan, Jiaxiang, Qian Wang, Zhaoliang Gao, Tingting Xiao, Hui Chen, Jinjing Zhang, Xiaoxia Song, and Jianchun Huang. 2022. "Analysis of the Composition of Substrate for Industrial Fermentation of Agaricus bisporus Based on Secondary and Tertiary Fermentation Mode Composition Analysis of Industrial Fermentation Substrates of A. bisporus" Fermentation 8, no. 5: 222. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050222