
Use of Autochthonous Lactic Acid Bacteria as Starter Culture of Pasteurized Milk Adobera Cheese

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lactic-Acid Bacteria Strains
2.1.1. Recovery and Growth
2.1.2. Growth Kinetics and Acidifying Capacity
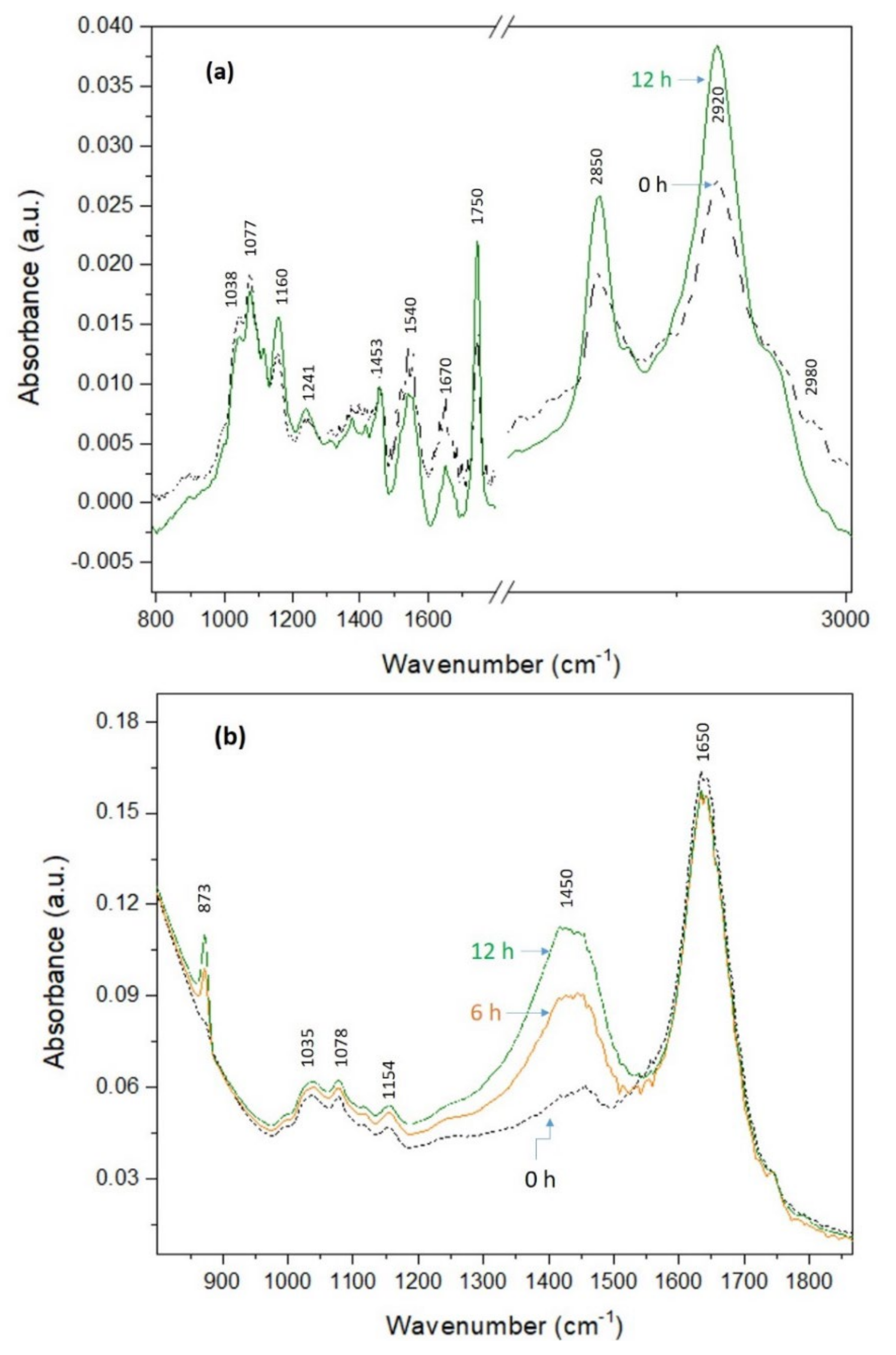
2.1.3. Assessment of Proteolytic and Acidifying Activities by FTIR
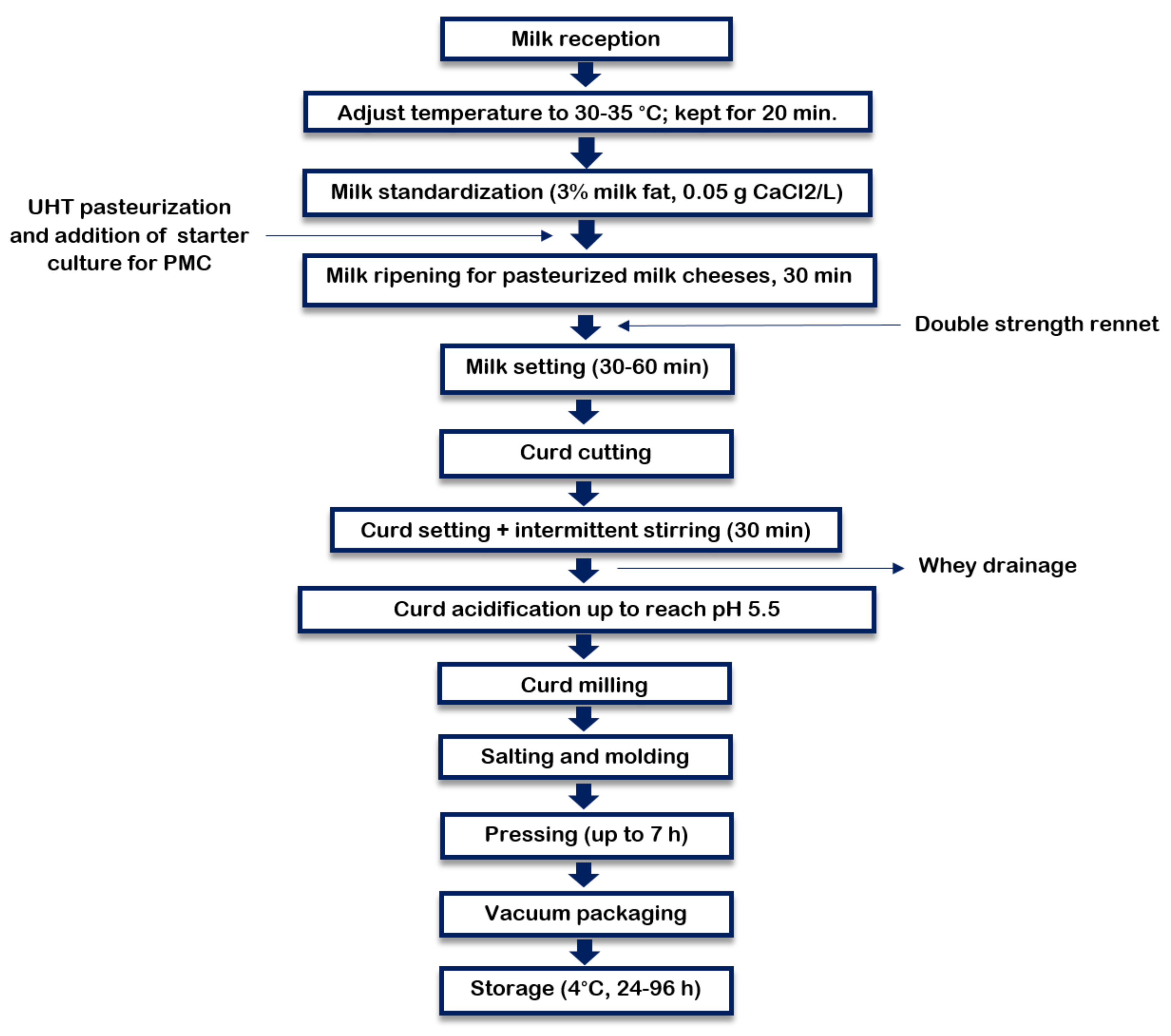
2.2. Experimental Cheeses
2.2.1. Raw Milk Cheeses (RMC)
2.2.2. Pasteurized Milk Cheeses (PMC)
2.3. Physicochemical and Compositional Analysis
Free Amino Acids Content
2.4. Texture Profile Analysis of Cheese
2.5. Cheese Color
2.6. Statistical Analysis
3. Results and Discussion
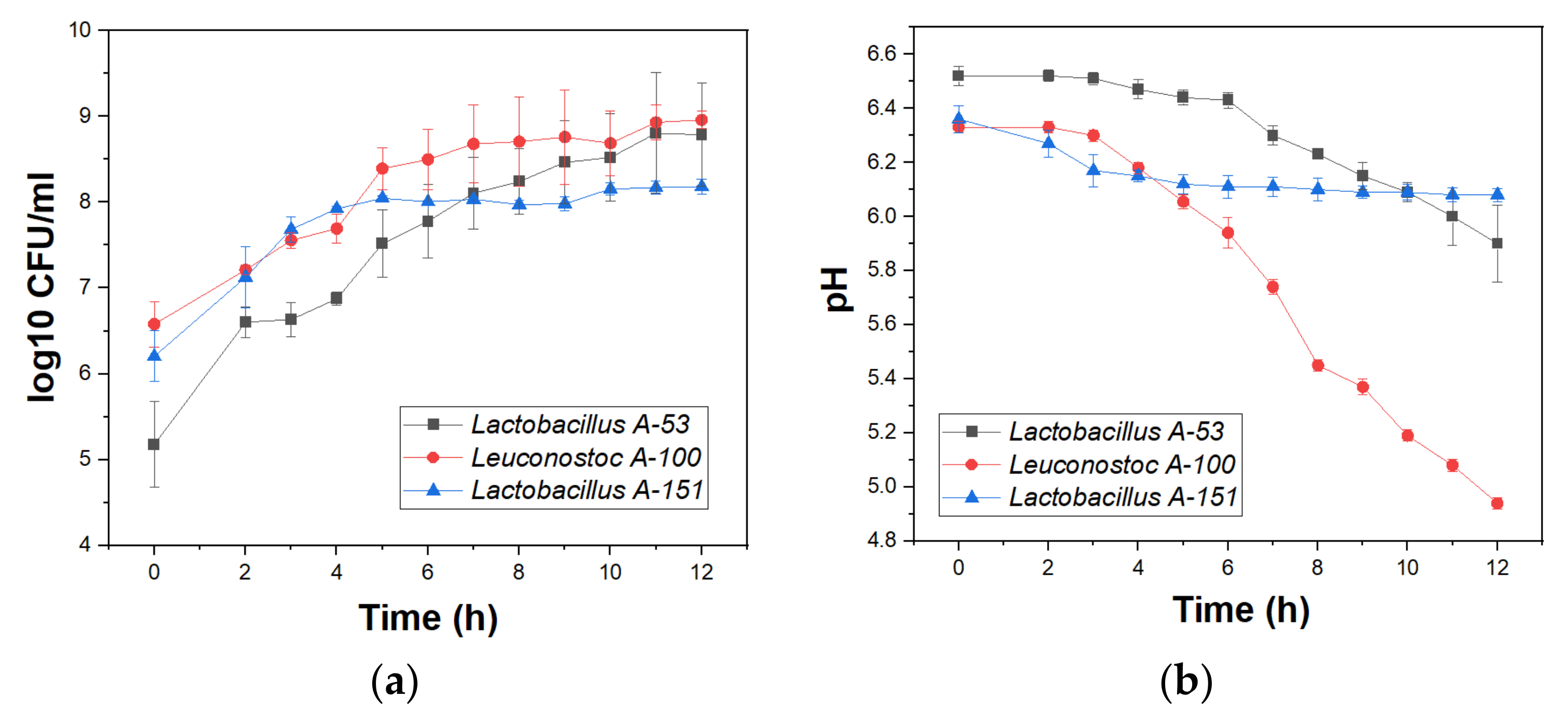
3.1. Strains Selection
3.2. Potential Fermentation Performance of Selected Strains
3.3. Cheese Milk Composition and Yield
3.4. Gross Composition
3.5. Free Amino Acids
3.6. Texture Profile Analysis
3.7. Cheese Color
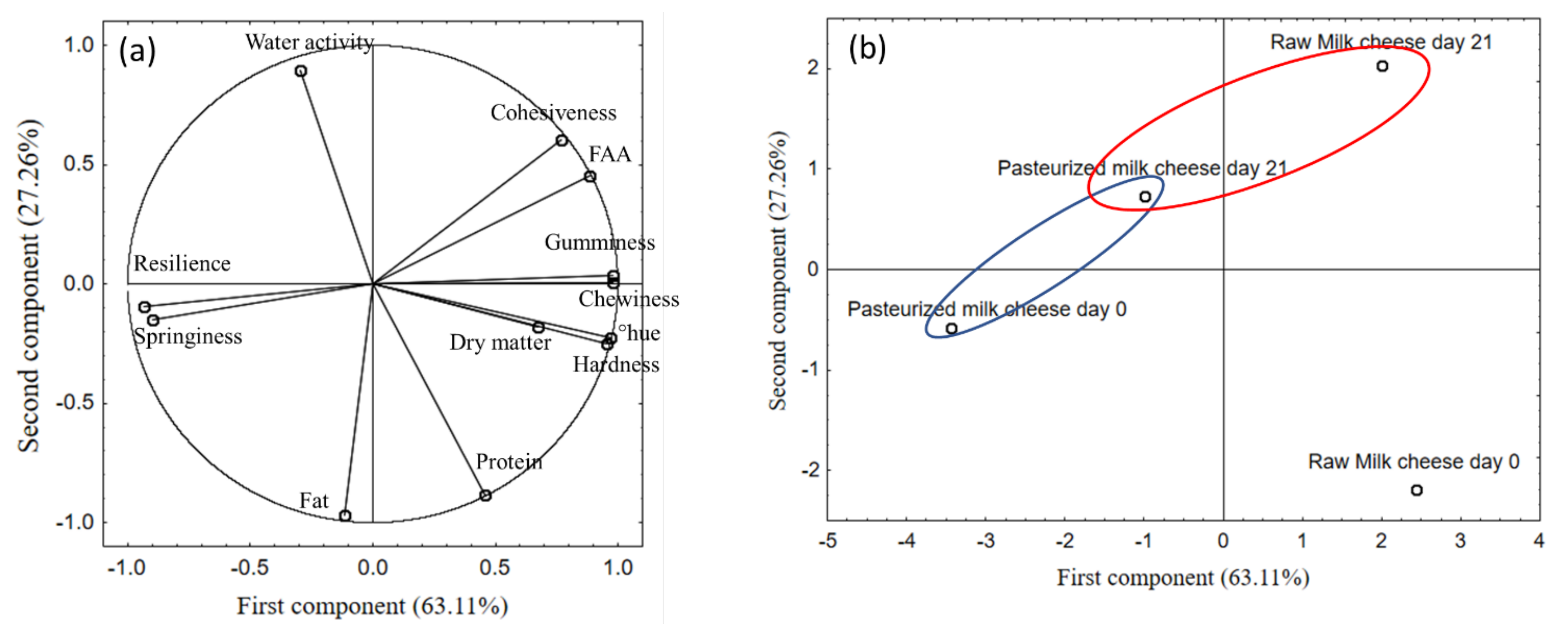
3.8. Principal Component Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Villegas-de Gante, A.; Cervantes-Escoto, F.; Cesín-Vargas, A.; Espinosa-Ortega, A.; Hernández-Montes, A.; Santos-Moreno, A.; Martínez-Campos, A. Atlas de Los Quesos Mexicanos Genuinos, 1st ed.; Colegio de Postgraduados: Texcoco, Mexico, 2014. [Google Scholar]
- González-Córdova, A.F.; Yescas, C.; Ortiz-Estrada, Á.M.; de la Rosa-Alcaraz, M.d.l.Á.; Hernández-Mendoza, A.; Vallejo-Cordoba, B. Invited Review: Artisanal Mexican Cheeses. J. Dairy Sci. 2016, 99, 3250–3262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centeno-Rodríguez, M.A.C.; Gutiérrez-Cárdenas, M.G.; Jaime-Patlán, M.; Meza-Plaza, E.F.; Montecillos-Ramírez, K.E.; Rojas-Salinas, W.B.; Ozuna, C. Genuine Mexican Cheeses: Technological Processes and Manufacturing Parameters. Agro Product. 2020, 13, 9–15. [Google Scholar] [CrossRef]
- Ruvalcaba-Gómez, J.M.; Ruiz-Espinosa, H.; Arteaga-Garibay, R.I.; Rojas-López, M.; Amador-Espejo, G.G.; Anaya-Esparza, L.M.; Delgado-Macuil, R.J. Texture, Physicochemical and Sensory Properties of Artisanal Adobera Cheese from Los Altos de Jalisco, a Genuine Mexican Cheese. Int. J. Dairy Technol. 2020, 73, 411–420. [Google Scholar] [CrossRef]
- Agudelo-López, M.A.; Cervantes-Escoto, F.; Cesín-Vargas, A.; Palacios-Rangel, M.I.; Espinoza-Ortega, A. Impact of Collective Trademarks in Mexican Artisanal Cheesemakers. Cienc. Tecnol. Agropecu. 2021, 22, e-1603. [Google Scholar] [CrossRef]
- Vázquez-Velázquez, R.; Salvador-Figueroa, M.; Adriano-Anaya, L.; DeGyves–Córdova, G.; Vázquez-Ovando, A. Use of Starter Culture of Native Lactic Acid Bacteria for Producing an Artisanal Mexican Cheese Safe and Sensory Acceptable. CYTA J. Food 2018, 16, 460–468. [Google Scholar] [CrossRef] [Green Version]
- Ruvalcaba-Gómez, J.M.; Delgado-Macuil, R.J.; Zelaya-Molina, L.X.; Maya-Lucas, O.; Ruesga-Gutiérrez, E.; Anaya-Esparza, L.M.; Villagrán-de la Mora, Z.; López-de la Mora, D.A.; Arteaga-Garibay, R.I. Bacterial Succession through the Artisanal Process and Seasonal Effects Defining Bacterial Communities of Raw-Milk Adobera Cheese Revealed by High Throughput DNA Sequencing. Microorganisms 2020, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Montes de Oca-Flores, E.; Cruz-Flores, M.A.; Espinoza-Ortega, A. Physicochemical and Microbiological Evaluation of Traditional Queso Molido (Ground Cheese) during Maturation. Agro Product. 2021, 14, 149–153. [Google Scholar] [CrossRef]
- De la Rosa-Alcaraz, M.d.l.Á.; Ortiz-Estrada, Á.M.; Heredia-Castro, P.Y.; Hernández-Mendoza, A.; Reyes-Díaz, R.; Vallejo-Cordoba, B.; González-Córdova, A.F. Poro de Tabasco Cheese: Chemical Composition and Microbiological Quality during Its Artisanal Manufacturing Process. J. Dairy Sci. 2020, 103, 3025–3037. [Google Scholar] [CrossRef]
- Adame-Gómez, R.; Muñoz-Barrios, S.; Castro-Alarcón, N.; Leyva-Vázquez, M.-A.; Toribio-Jiménez, J.; Ramírez-Peralta, A. Prevalence of the Strains of Bacillus Cereus Group in Artisanal Mexican Cheese. Foodborne Pathog. Dis. 2020, 17, 8–14. [Google Scholar] [CrossRef]
- González Ariceaga, C.C.; Afzal, M.I.; Umer, M.; Abbas, S.; Ahmad, H.; Sajjad, M.; Parvaiz, F.; Imdad, K.; Imran, M.; Maan, A.A.; et al. Physicochemical, Sensorial and Microbiological Characterization of PoroCheese, an Artisanal Mexican Cheese Made from Raw Milk. Foods 2019, 8, 509. [Google Scholar] [CrossRef] [Green Version]
- Escobar-Zepeda, A.; Sanchez-Flores, A.; Quirasco Baruch, M. Metagenomic Analysis of a Mexican Ripened Cheese Reveals a Unique Complex Microbiota. Food Microbiol. 2016, 57, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Aldrete-Tapia, A.; Escobar-Ramírez, M.C.; Tamplin, M.L.; Hernández-Iturriaga, M. High-Throughput Sequencing of Microbial Communities in Poro Cheese, an Artisanal Mexican Cheese. Food Microbiol. 2014, 44, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Cervantes-Escoto, F.; Villegas-de Gante, A.; Cesín-Vargas, A. Caracterización de La Cadena Agroindustrial Del Queso Adobera de Los Altos de Jalisco, México. In Proceedings of the 1er Simposio de Los Quesos Mexicanos Genuinos, Texcoco, Mexico State, Mexico, 18 May 2016. [Google Scholar]
- Ruvalcaba-Gómez, J.M.; Arteaga-Garibay, R.I.; Delgado-Macuil, R.J.; Villaseñor-González, F.; Montes-Oceguera, L. Exploración Del Perfil de Inocuidad Microbiológica de Queso Adobera Elaborado Artesanalmente. In Investigaciones en Ciencia e Inocuidad de Alimentos; Orozco-Hernández, L.O., Garay-Martínez, L.E., Torres-Vitela, M.R., Eds.; Universidad de Guadalajara: Guadalajara, Jalisco, México, 2017; Volume 1, pp. 738–741. [Google Scholar]
- Torres-Vitela, M.R.; Mendoza-Bernardo, M.; Castro-Rosas, J.; Gomez-Aldapa, C.A.; Garay-Martinez, L.E.; Navarro-Hidalgo, V.; Villarruel-López, A. Incidence of Salmonella, Listeria monocytogenes, Escherichia coli O157:H7, and Staphylococcal Enterotoxin in Two Types of Mexican Fresh Cheeses. J. Food Prot. 2012, 75, 79–84. [Google Scholar] [CrossRef]
- Havelaar, A.; Grace, D.; Wu, F. Foodborne Diseases from Dairy Products in Developing Countries: Hazards and Health Implications. In Proceedings of the 2019 Annual Meeting of the American Dairy Science Association, Cincinnati, OH, USA, 23–26 June 2019. [Google Scholar]
- Licitra, G.; Caccamo, M.; Lortal, S. Artisanal Products Made With Raw Milk. In Raw Milk; Elsevier: Amsterdam, The Netherlands, 2019; pp. 175–221. [Google Scholar]
- Fox, P.F.; Guinee, T.P.; Cogan, T.M.; McSweeney, P.L.H. Fundamentals of Cheese Science; Springer US: Boston, MA, USA, 2017. [Google Scholar]
- Tarhan, Ö.; Kaya, A. Investigation of the Compositional and Structural Changes in the Proteins of Cow Milk When Processed to Cheese. LWT 2021, 151, 112102. [Google Scholar] [CrossRef]
- Scatassa, M.L.; Gaglio, R.; Cardamone, C.; Macaluso, G.; Arcuri, L.; Todaro, M.; Mancuso, I. Anti-Listeria Activity of Lactic Acid Bacteria in Two Traditional Sicilian Cheeses. Ital. J. Food Saf. 2017, 6, PMC5337783. [Google Scholar] [CrossRef] [Green Version]
- Murugesan, S.; Reyes-Mata, M.P.; Nirmalkar, K.; Chavez-Carbajal, A.; Juárez-Hernández, J.I.; Torres-Gómez, R.E.; Piña Escobedo, A.; Maya, O.; Hoyo-Vadillo, C.; Ramos-Ramírez, E.G.; et al. Profiling of bacterial and fungal communities of Mexican cheeses by high throughput DNA sequencing. Food Res. Int. 2018, 113, 371–381. [Google Scholar] [CrossRef]
- Association of Officiating Analytical Chemists (AOAC). Official Method of Analysis, 18th ed.; Association of Officiating Analytical Chemists (AOAC): Washington, DC, USA, 2005. [Google Scholar]
- Wehr, H.; Frank, J.F. Standard Methods for the Examination of Dairy Products, 17th ed.; American Public Health Association: Washington, DC, USA, 2004. [Google Scholar]
- Wick, C.; Nienaber, U.; Anggraeni, O.; Shellhammer, T.H.; Courtney, P.D. Texture, Proteolysis and Viable Lactic Acid Bacteria in Commercial Cheddar Cheeses Treated with High Pressure. J. Dairy Res. 2004, 71, 107–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.S.; Al-Attabi, Z.H.; Al-Habsi, N.; Al-Khusaibi, M. Measurement of Instrumental Texture Profile Analysis (TPA) of Foods. In Techniques to Measure Food Safety and Quality; Springer International Publishing: Cham, Switzerland, 2021; pp. 427–465. [Google Scholar]
- Hirschler, R. Whiteness, Yellowness, and Browning in Food Colorimetry. A Critical Review. In Color in Food. Technological and Psychophysical Aspects; Luis Caivano, J., del Pilar Buera, M., Eds.; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Boca Raton, FL, USA, 2012; pp. 93–104. [Google Scholar]
- Sánchez-Feria, C.; Salinas Moreno, Y.; Ybarra-Moncada, M.D.C.; González-Hernández, V.A.; Machuca-Sánchez, M.L. Effect of the Dehydration Method of Hibiscus Sabdariffa L. Calyces on the Quality of Their Aqueous Extracts. Emir. J. Food Agric. 2021, 33, 159–170. [Google Scholar]
- Engels, W.; Dusterhoft, E.-M.; Huppertz, T. Starter Cultures for Cheese Manufacture. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Settanni, L.; Moschetti, G. Non-starter lactic acid bacteria used to improve cheese quality and provide health benefits. Food Microbiol. 2010, 27, 691–697. [Google Scholar] [CrossRef]
- Gobbetti, M.; Di Cagno, R.; Calasso, M.; Neviani, E.; Fox, P.F.; De Angelis, M. Drivers that establish and assembly the lactic acid bacteria biota in cheeses. Trends Food Sci. Technol. 2018, 78, 244–254. [Google Scholar] [CrossRef]
- Nieto-Arribas, P.; Seseña, S.; Poveda, J.M.; Palop, L.; Cabezas, L. Genotypic and technological characterization of Leuconostoc isolates to be used as adjunct starters in Manchego cheese manufacture. Food Microbiol. 2010, 27, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Paucean, A.; Vodnar, D.-C.; Socaci, S.-A.; Socaciu, C. Carbohydrate Metabolic Conversions to Lactic Acid and Volatile Derivatives, as Influenced by Lactobacillus plantarum ATCC 8014 and Lactobacillus casei ATCC 393 Efficiency during in Vitro and Sourdough Fermentation. Eur. Food Res. Technol. 2013, 237, 679–689. [Google Scholar] [CrossRef]
- Păucean, A.; Vodnar, D.C.; Mureșan, V.; Fetea, F.; Ranga, F.; Man, S.M.; Muste, S.; Socaciu, C. Monitoring Lactic Acid Concentrations by Infrared Spectroscopy: A New Developed Method for Lactobacillus Fermenting Media with Potential Food Applications. Acta Aliment. 2017, 46, 420–427. [Google Scholar] [CrossRef] [Green Version]
- Ranvir, S.; Sharma, R.; Gandhi, K.; Upadhyay, N.; Mann, B. Assessment of Proteolysis in Ultra-high Temperature Milk Using Attenuated Total Reflectance–Fourier Transform Infrared Spectroscopy. Int. J. Dairy Technol. 2020, 73, 366–375. [Google Scholar] [CrossRef]
- Guinee, T.P.; Mulholland, E.O.; Kelly, J.; Callaghan, D.J.O. Effect of Protein-to-Fat Ratio of Milk on the Composition, Manufacturing Efficiency, and Yield of Cheddar Cheese. J. Dairy Sci. 2007, 90, 110–123. [Google Scholar] [CrossRef]
- Ali, F.; Wang, Z.-X. Effect of Pasteurization on the Enzymatic Cross-Linking of Milk Proteins by Microbial Transglutaminase in View of Milk Fat Globule Membrane Isolation. Food Biosci. 2021, 43, 101100. [Google Scholar] [CrossRef]
- Gharibzahedi, S.M.T.; Chronakis, I.S. Crosslinking of Milk Proteins by Microbial Transglutaminase: Utilization in Functional Yogurt Products. Food Chem. 2018, 245, 620–632. [Google Scholar] [CrossRef]
- Morales-Celaya, M.F.; Lobato-Calleros, C.; Alvarez-Ramirez, J.; Vernon-Carter, E.J. Effect of Milk Pasteurization and Acidification Method on the Chemical Composition and Microstructure of a Mexican Pasta Filata Cheese. LWT-Food Sci. Technol. 2012, 45, 132–141. [Google Scholar] [CrossRef]
- Lucey, J.A. Cheese|Rennet-Induced Coagulation of Milk. In Encyclopedia of Dairy Sciences; Elsevier: Amsterdam, The Netherlands, 2011; pp. 579–584. [Google Scholar]
- D’Incecco, P.; Hogenboom, J.A.; Rosi, V.; Cabassi, G.; Pellegrino, L. Effects of Microbial Coagulants from Rhyzomucor Miehei on Composition, Sensory and Textural Characteristics of Long-Ripened Hard Cheeses. Food Chem. 2022, 370, 131043. [Google Scholar] [CrossRef]
- Guinee, T.P. Effect of High-Temperature Treatment of Milk and Whey Protein Denaturation on the Properties of Rennet–Curd Cheese: A Review. Int. Dairy J. 2021, 121, 105095. [Google Scholar] [CrossRef]
- Pappa, E.C.; Kondyli, E.; Samelis, J. Microbiological and Biochemical Characteristics of Kashkaval Cheese Produced Using Pasteurised or Raw Milk. Int. Dairy J. 2019, 89, 60–67. [Google Scholar] [CrossRef]
- Buffa, M.N.; Trujillo, A.J.; Pavia, M.; Guamis, B. Changes in Textural, Microstructural, and Colour Characteristics during Ripening of Cheeses Made from Raw, Pasteurized or High-Pressure-Treated Goats’ Milk. Int. Dairy J. 2001, 11, 927–934. [Google Scholar] [CrossRef]
- Hickey, D.K.; Kilcawley, K.N.; Beresford, T.P.; Wilkinson, M.G. Lipolysis in Cheddar Cheese Made from Raw, Thermized, and Pasteurized Milks. J. Dairy Sci. 2007, 90, 47–56. [Google Scholar] [CrossRef]
- Grappin, R.; Beuvier, E. Possible Implications of Milk Pasteurization on the Manufacture and Sensory Quality of Ripened Cheese. Int. Dairy J. 1997, 7, 751–761. [Google Scholar] [CrossRef]
- Hougaard, A.B.; Ardö, Y.; Ipsen, R.H. Cheese Made from Instant Infusion Pasteurized Milk: Rennet Coagulation, Cheese Composition, Texture and Ripening. Int. Dairy J. 2010, 20, 449–458. [Google Scholar] [CrossRef]
- Rank, T.C.; Grappin, R.; Olson, N.F. Secondary Proteolysis of Cheese during Ripening: A Review. J. Dairy Sci. 1985, 68, 801–805. [Google Scholar] [CrossRef]
- Folkertsma, B.; Fox, P.F. Use of the Cd-Ninhydrin Reagent to Assess Proteolysis in Cheese during Ripening. J. Dairy Res. 1992, 59, 217–224. [Google Scholar] [CrossRef]
- De Aguiar e Câmara, S.P.; Maduro Dias, C.; Rocha, L.; Dapkevicius, A.; Duarte Rosa, H.J.; de Borba, A.E.S.; Silveira, M.d.G.; Malcata, F.X.; de Lurdes Enes Dapkevicius, M. Assessment of Autochthonous Lactic Acid Bacteria as Starter Cultures for Improved Manufacture of Pico Cheese Using a Cheese Model. Int. Dairy J. 2022, 128, 105294. [Google Scholar] [CrossRef]
- Öründü, S.; Tarakçı, Z. Effects of Different Starter Culture Applications Pre- and Post-Scalding on the Biochemical and Sensory Properties of Pasta Filata Type Cheeses. LWT 2021, 136, 110288. [Google Scholar] [CrossRef]
- Kok, J.; de Vos, W.M. The Proteolytic System of Lactic Acid Bacteria. In Genetics and Biotechnology of Lactic Acid Bacteria; Springer Netherlands: Dordrecht, The Netherlands, 1994; pp. 169–210. [Google Scholar]
- Sánchez-Gamboa, C.; Hicks-Pérez, L.; Gutiérrez-Méndez, N.; Heredia, N.; García, S.; Nevárez-Moorillón, G. Microbiological Changes during Ripening of Chihuahua Cheese Manufactured with Raw Milk and Its Seasonal Variations. Foods 2018, 7, 153. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, S.; Rodríguez, V.; Ruiz, M.E.; Fresno, M. Correlaciones de Textura y Color Instrumental Con La Composición Química de Quesos de Cabra Canarios. Arch. Zootec. 2007, 56, 663–666. [Google Scholar]
- Sandoval-Copado, J.; Orozco-Villafuerte, J.; Pedrero-Fuehrer, D.; Colín-Cruz, M.A. Sensory Profile Development of Oaxaca Cheese and Relationship with Physicochemical Parameters. J. Dairy Sci. 2016, 99, 7075–7084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Creamer, L.K.; Olson, N.F. Rheological Evaluation of Maturing Cheddar Cheese. J. Food Sci. 1982, 47, 631–636. [Google Scholar] [CrossRef]
- Golin Bueno Costa, R.; Caiafa Junior, A.; Gomes da Cruz, A.; Sobral, D.; Gonçalves Costa Júnior, L.C.; Jacinto de Paula, J.C.; Magalhães Machado Moreira, G.; Martins Teodoro, V.A. Effect of Partial Replacement of Sodium Chloride with Potassium Chloride on the Characteristics of Minas Padrão Cheese. Int. Dairy J. 2019, 91, 48–54. [Google Scholar] [CrossRef]
- Tomar, O. The Effects of Probiotic Cultures on the Organic Acid Content, Texture Profile and Sensory Attributes of Tulum Cheese. Int. J. Dairy Technol. 2019, 72, 218–228. [Google Scholar] [CrossRef]
- Sert, D.; Akin, N.; Aktumsek, A. Lipolysis in Tulum Cheese Produced from Raw and Pasteurized Goats’ Milk during Ripening. Small Rumin. Res. 2014, 121, 351–360. [Google Scholar] [CrossRef]
- Khanal, B.K.S.; Bhandari, B.; Prakash, S.; Liu, D.; Zhou, P.; Bansal, N. Modifying Textural and Microstructural Properties of Low Fat Cheddar Cheese Using Sodium Alginate. Food Hydrocoll. 2018, 83, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Diezhandino, I.; Fernández, D.; Sacristán, N.; Combarros-Fuertes, P.; Prieto, B.; Fresno, J.M. Rheological, Textural, Colour and Sensory Characteristics of a Spanish Blue Cheese (Valdeón Cheese). LWT-Food Sci. Technol. 2016, 65, 1118–1125. [Google Scholar] [CrossRef]
- Chudy, S.; Bilska, A.; Kowalski, R.; Teichert, J. Colour of Milk and Milk Products in CIE L*a*b* Space. Med. Weter 2020, 76, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Milovanovic, B.; Djekic, I.; Miocinovic, J.; Djordjevic, V.; Lorenzo, J.M.; Barba, F.J.; Mörlein, D.; Tomasevic, I. What Is the Color of Milk and Dairy Products and How Is It Measured? Foods 2020, 9, 1629. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Raw Milk | Pasteurized Milk |
|---|---|---|
| Fat content (%) | 3.63 | 3.39 |
| Non-fat solids (%) | 8.25 | 8.25 |
| Lactose (%) | 4.54 | 4.53 |
| Protein (%) | 2.51 | 2.43 |
| Salts (%) | 0.67 | 0.67 |
| Density (kg/L) | 1.028 | 1.028 |
| Freezing point (°C) | −0.525 | −0.523 |
| 3% fat standardized milk | ||
| Protein:fat ratio | 0.84 | 0.81 |
| Cheese yield (kg of cheese/100 kg/milk) | 8.45 ± 1.30 | 10.73 ± 1.88 |
| Post-Processing Days | Dry Matter (%) | Water Activity (Aw) | Protein Content (%) | Fat Content (%) | ||||
|---|---|---|---|---|---|---|---|---|
| RMC | PMC | RMC | PMC | RMC | PMC | RMC | PMC | |
| 0 | 51.75 a,A (±7.35) | 49.97 b,A (±0.75) | 0.94 a,A (±0.006) | 0.95 a,A (±0.10) | 46.93 a,A (±4.69) | 43.24 a,A (±5.22) | 46.84 a,A (±5.53) | 43.68 a,A (±4.67) |
| 21 | 51.73 a,A (±4.48) | 48.04 b,B (±2.86) | 0.954 a,A (±0.001) | 0.948 b,A (±0.004) | 42.38 a,A (±2.69) | 42.19 a,A (±1.51) | 37.35 b,B (±8.81) | 42.40 a,A (±0.80) |
| Post-Processing Days | Free Amino Acids Content (FAA) * | |
|---|---|---|
| RMC | PMC | |
| 0 | 0.346 (±0.080) a,B | 0.239 (±0.070) b,C |
| 7 | 0.368 (±0.107) a,B | 0.283 (±0.109) b,B |
| 14 | 0.394 (±0.110) a,A | 0.295 (±0.109) b,B |
| 21 | 0.394 (±0.113) a,A | 0.326 (±0.127) b,A |
| Days | Hardness (kg) | Cohesiveness (-) | Springiness (-) | Chewiness (kg) | Resilience (-) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| RMC | PMC | RMC | PMC | RMC | PMC | RMC | PMC | RMC | PMC | |
| 0 | 2.09 ± 0.89 a,A | 0.698 ± 0.25 b,A | 0.50 ± 0.11 a,B | 0.29 ± 0.11 b,B | 0.84 ± 0.04 a,A | 1.10 ± 0.24 b,A | 0.92 ± 0.51 a,A | 0.20 ± 0.09 b,B | 0.26 ± 0.06 b,A | 0.78 ± 0.48 a,A |
| 7 | 1.90 ± 1.39 a,A | 0.870 ± 0.36 b,A | 0.58 ± 0.06 b,AB | 0.63 ± 0.06 a,A | 0.78 ± 0.04 a,B | 0.80 ± 0.02 a,B | 0.84 ± 0.55 a,A | 0.44 ± 0.17 b,A | 0.26 ± 0.03 a,A | 0.28 ± 0.04 a,B |
| 14 | 1.90 ± 1.16 a,A | 1.068 ± 0.46 b,A | 0.62 ± 0.07 a,A | 0.48 ± 0.17 b,A | 0.79 ± 0.03 b,B | 0.83 ± 0.12 a,B | 0.92 ± 0.58 a,A | 0.43 ± 0.27 b,A | 0.28 ± 0.02 a,A | 0.32 ± 0.12 a,B |
| 21 | 1.69 ± 0.99 a,A | 0.885 ± 0.53 b,A | 0.69 ± 0.05 a,A | 0.55 ± 0.19 b,A | 0.85 ± 0.05 a,A | 0.88 ± 0.08 b,B | 0.92 ± 0.44 a,A | 0.36 ± 0.15 b,AB | 0.30 ± 0.04 a,A | 0.39 ± 0.12 a,B |
| Days | L* | a* | b* | Color (°Hue) | ||||
|---|---|---|---|---|---|---|---|---|
| RMC | PMC | RMC | PMC | RMC | PMC | RMC | PMC | |
| 0 | 87.31 ± 3.41 a,A | 87.87 ± 1.78 a,A | 0.16 ± 0.43 a,AB | −0.89 ± 0.27 b,A | 13.83 ± 2.29 a,AB | 12.14 ± 1.49 b,C | 88.65 ± 0.91 a,A | 85.52 ± 1.21 b,A |
| 7 | 86.15 ± 2.17 b,AB | 87.76 ± 3.47 a,AB | 0.48 ± 0.52 a,A | −1.65 ± 0.95 b,B | 12.47 ± 0.83 b,B | 13.75 ± 1.95 a,AB | 87.35 ± 1.90 a,B | 83.50 ± 2.95 b,B |
| 14 | 85.23 ± 2.21 b,B | 87.32 ± 1.83 a,AB | −0.20 ± 0.84 a,B | −1.08 ± 0.32 b,AB | 14.00 ± 1.25 a,A | 12.83 ± 1.36 b,AC | 86.92 ± 1.12 a,B | 85.11 ± 1.49 b,AB |
| 21 | 84.28 ± 3.08 b,B | 86.79 ± 4.82 a,B | 0.53 ± 0.28 a,A | −0.56 ± 0.60 b,A | 13.21 ± 0.78 b,AB | 13.94 ± 0.94 a,A | 87.66 ± 1.22 a,AB | 86.72 ± 0.24 b,A |
| Chroma | Whiteness | Yellowness | Color | |||||
| Days | RMC | PMC | RMC | PMC | RMC | PMC | RMC | PMC |
| 0 | 14.04 ± 2.30 a,AB | 13.02 ± 1.60 b,B | 81.53 ± 3.81 a,A | 82.76 ± 1.79 a,A | 22.65 ± 4.49 a,A | 19.76 ± 4.75 a,C |  |  |
| 7 | 12.95 ± 0.83 b,B | 17.28 ± 2.04 a,A | 81.30 ± 1.70 a,A | 81.38 ± 3.29 a,AB | 20.69 ± 1.53 b,A | 22.45 ± 3.53 b,AB |  |  |
| 14 | 14.68 ± 1.26 a,A | 14.11 ± 1.35 b,B | 79.62 ± 2.39 b,A | 81.89 ± 1.88 a,A | 23.53 ± 2.58 a,A | 21.01 ± 2.41 b,BC |  |  |
| 21 | 13.57 ± 0.78 b,AB | 14.57 ± 0.94 a,B | 79.38 ± 2.55 a,A | 79.40 ± 3.15 a,B | 22.45 ± 1.81 b,A | 23.36 ± 0.85 a,A |  |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruvalcaba-Gómez, J.M.; Ruiz-Espinosa, H.; Méndez-Robles, M.D.; Arteaga-Garibay, R.I.; Anaya-Esparza, L.M.; Villagrán, Z.; Delgado-Macuil, R.J. Use of Autochthonous Lactic Acid Bacteria as Starter Culture of Pasteurized Milk Adobera Cheese. Fermentation 2022, 8, 234. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050234
Ruvalcaba-Gómez JM, Ruiz-Espinosa H, Méndez-Robles MD, Arteaga-Garibay RI, Anaya-Esparza LM, Villagrán Z, Delgado-Macuil RJ. Use of Autochthonous Lactic Acid Bacteria as Starter Culture of Pasteurized Milk Adobera Cheese. Fermentation. 2022; 8(5):234. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050234
Chicago/Turabian StyleRuvalcaba-Gómez, José Martín, Héctor Ruiz-Espinosa, María Dolores Méndez-Robles, Ramón Ignacio Arteaga-Garibay, Luis Miguel Anaya-Esparza, Zuamí Villagrán, and Raúl Jacobo Delgado-Macuil. 2022. "Use of Autochthonous Lactic Acid Bacteria as Starter Culture of Pasteurized Milk Adobera Cheese" Fermentation 8, no. 5: 234. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050234