
Impact of Oil Sources on In Vitro Fermentation, Microbes, Greenhouse Gas, and Fatty Acid Profile in the Rumen

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Incubation
2.2. Sampling and Chemical Analysis
2.3. DNA Extraction, Primer Information, and Real-Time PCR Analysis
2.4. Statistical Analysis
3. Results
3.1. Fatty Acid Profiles before Incubation
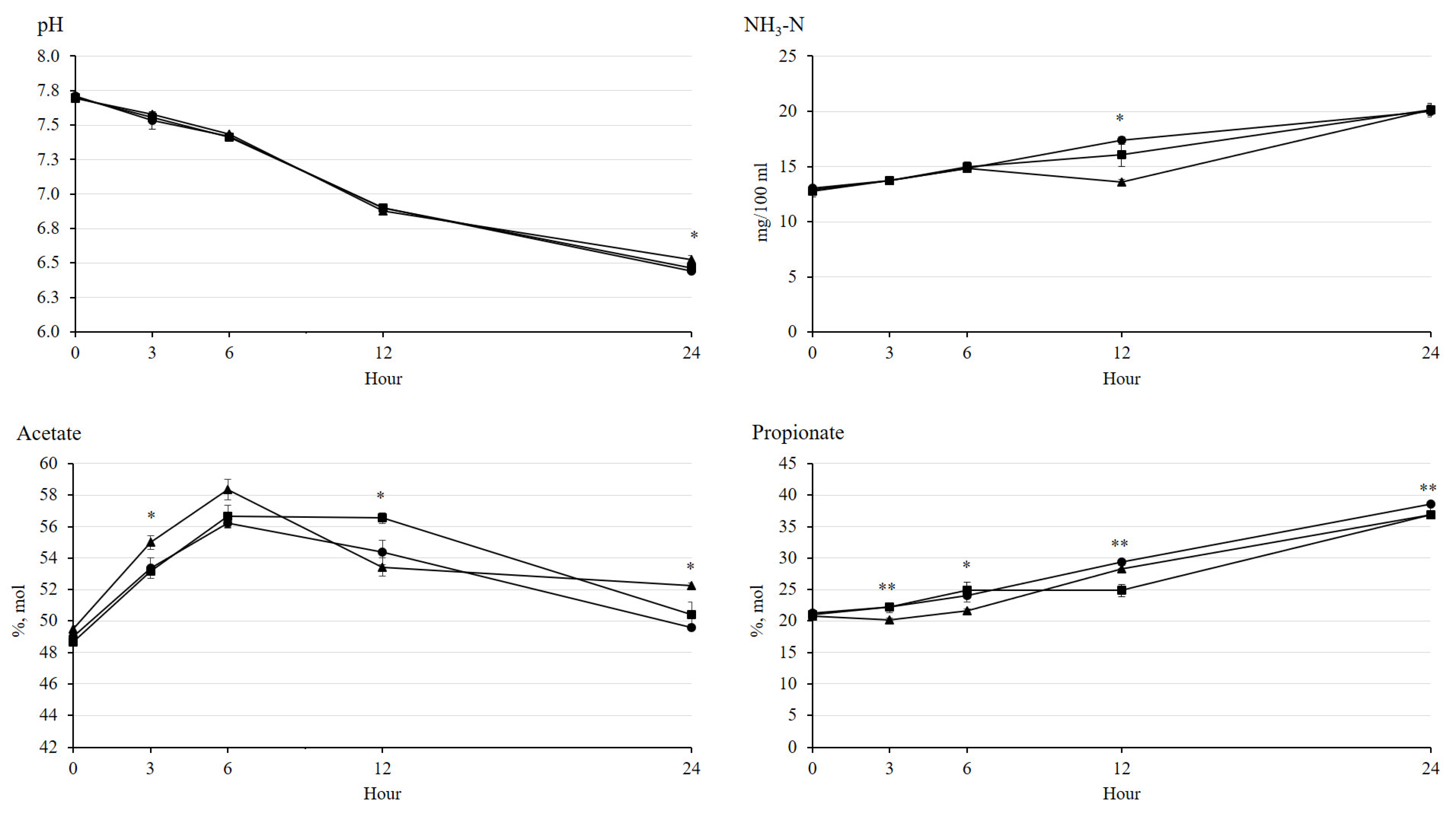
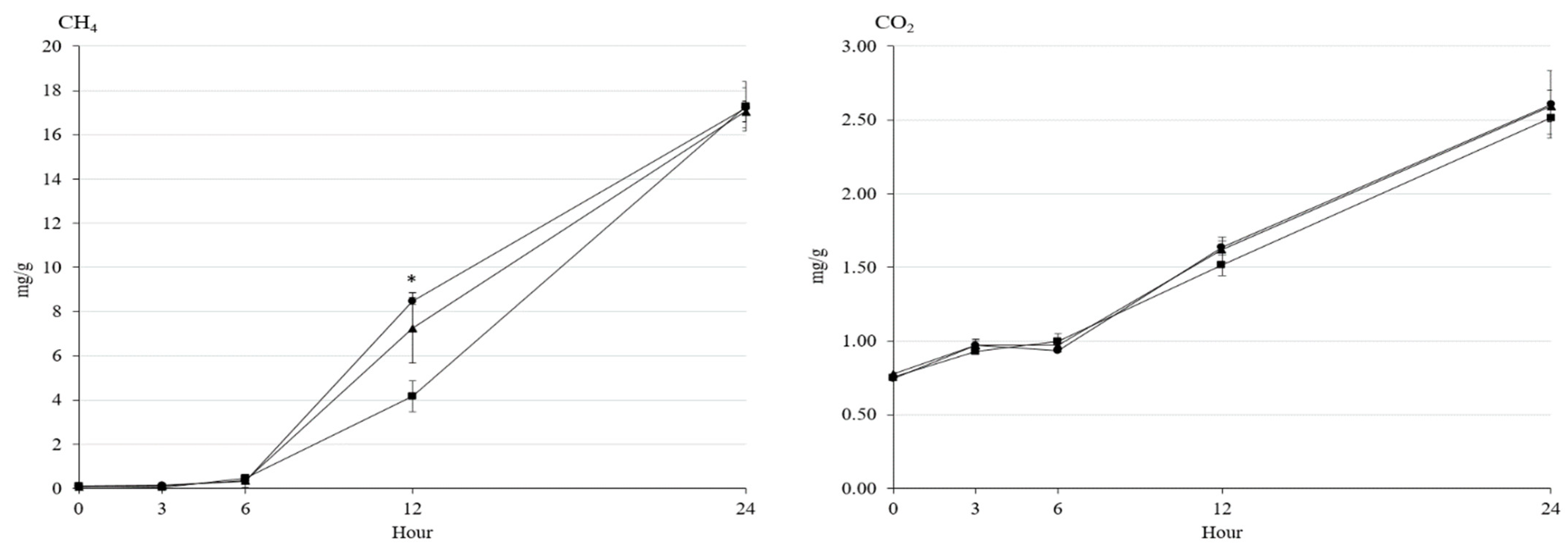
3.2. Fermentation Characteristics and Greenhouse Gas Emission
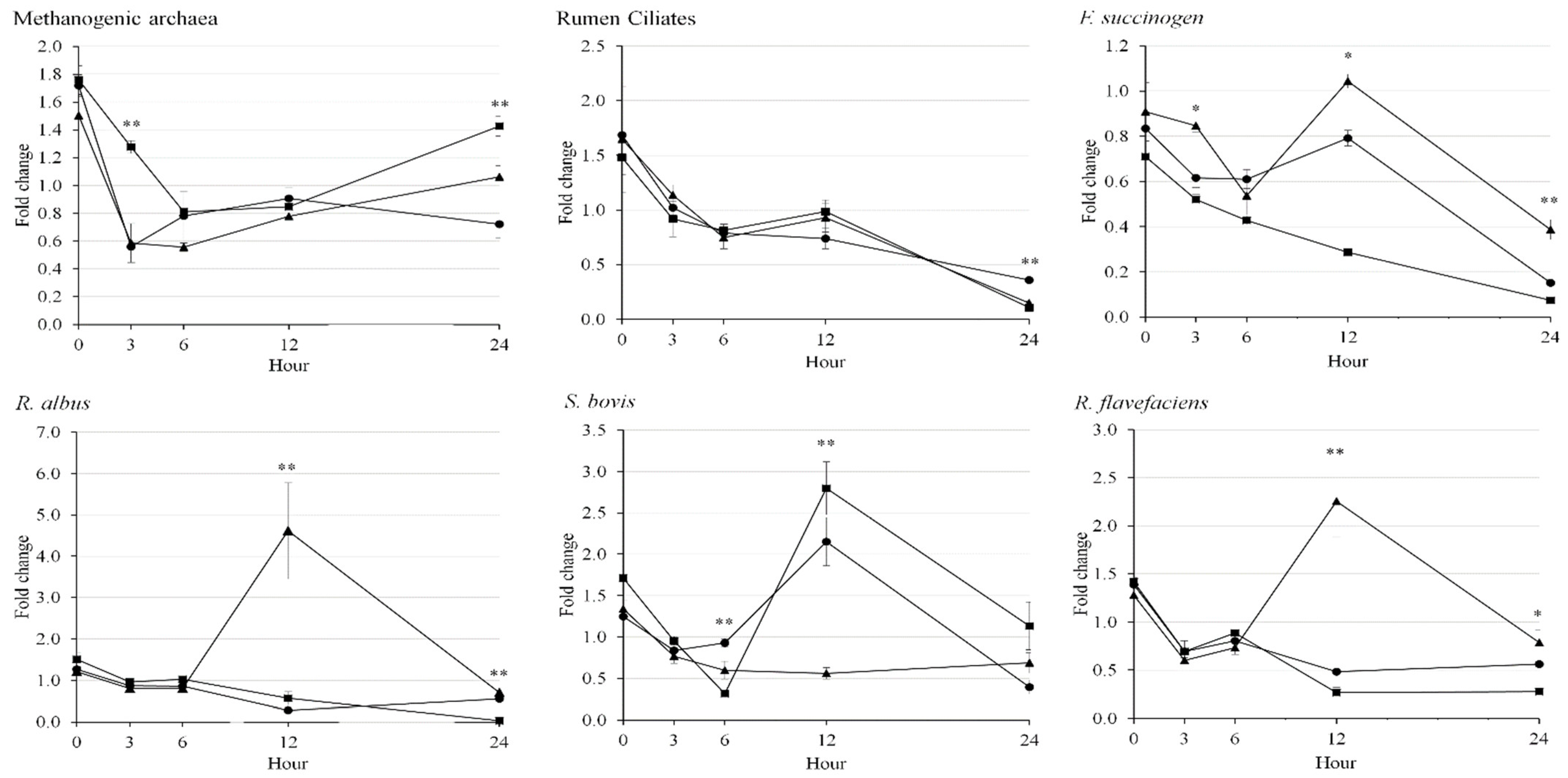
3.3. Rumen Microbes
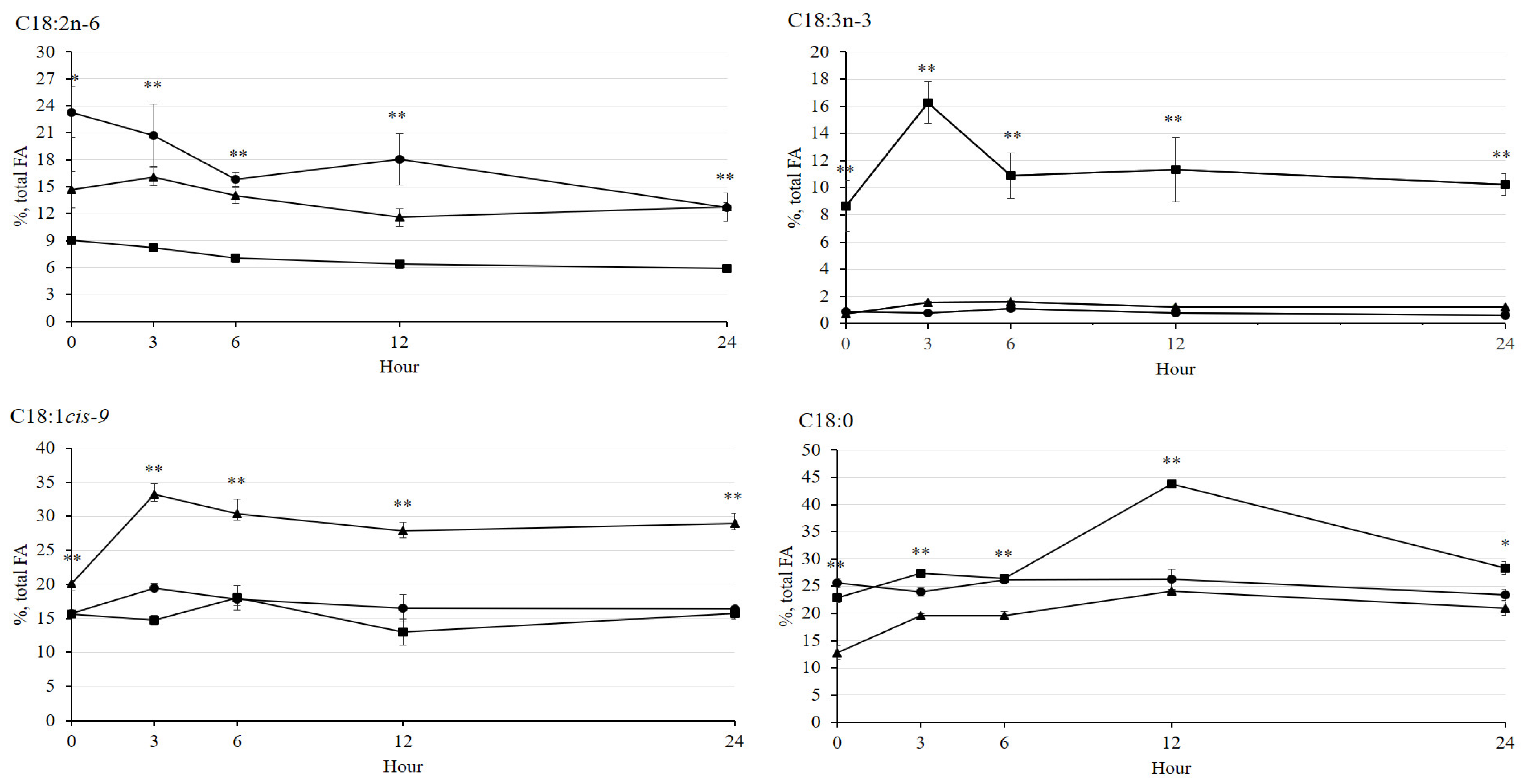
3.4. Fatty Acid Profiles after Incubation
4. Discussion
4.1. Rumen Fermentation Characteristics, Microbial Population, and Methanogenesis
4.2. Rumen Fatty Acid and Profiles
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vahmani, P.; Mapiye, C.; Prieto, N.; Rolland, D.C.; McAllister, T.A.; Aalhus, J.L.; Dugan, M.E.R. The scope for manipulating the polyunsaturated fatty acid content of beef: A review. J. Anim. Sci. Biotechnol. 2015, 6, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanier, J.S.; Cor, B.A. Challenges in enriching milk fat with polyunsaturated fatty acids. J. Anim. Sci. Biotechnol. 2015, 6, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauchemin, K.A.; Kreuzer, M.; O’Mara, F.; McAllister, T.A. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27. [Google Scholar] [CrossRef]
- Patra, A.; Park, A.; Kim, M.; Yu, Z. Rumen methanogens and mitigation of methane emission by anti-methanogenic compounds and substance. J. Anim. Sci. Biotechnol. 2017, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayat, A.R.; Ventto, L.; Kairenius, P.; Stefański, T.; Leskinen, H.; Tapio, I.; Negussie, E.; Vilkki, J.; Shingfield, K.J. Dietary forage to concentrate ratio and sunflower oil supplement alter rumen fermentation, ruminal methane emission, and nutrient utilization in lactating cows. Transl. Anim. Sci. 2017, 1, 277–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, J.E.; Andrés, S.; López-Ferreras, L.; Snelling, T.J.; Yánez-Ruíz, D.R.; Garcia-Estrada, G.; López, S. Dietary supplemental plant oils reduce methanogenesis from anaerobic microbial fermentation in the rumen. Sci. Rep. 2020, 10, 1613. [Google Scholar] [CrossRef] [Green Version]
- Beauchemin, K.A.; McGinn, S.M. Methane emissions from beef cattle: Effects of fumaric acid, essential oil, and canola oil. J. Anim. Sci. 2006, 84, 1489–1496. [Google Scholar] [CrossRef]
- Palmquist, D.L.; Jenkins, T.C. Fat in lactation rations for dairy: A review. J. Dairy Sci. 1980, 63, 1–14. [Google Scholar] [CrossRef]
- Zhang, C.M.; Guo, Y.Q.; Yuan, Z.P.; Wu, Y.M.; Wang, J.K.; Liu, J.X.; Zhu, W.Y. Effect of octadeca carbon fatty acids on microbial fermentation, methanogenesis and microbial flora in vitro. Anim. Feed Sci. Technol. 2008, 146, 259–269. [Google Scholar] [CrossRef]
- Amanullah, S.M.; Kim, D.H.; Paradhipta, D.H.V.; Lee, H.J.; Joo, Y.H.; Lee, S.S.; Kim, E.T.; Kim, S.C. Effects of essential fatty acid supplementation on in vitro fermentation indices, greenhouse gas, microbes, and fatty acid profiles in the rumen. Front. Microbiol. 2021, 12, 401. [Google Scholar] [CrossRef]
- Duarte, A.C.; Durmic, Z.; Vercoe, P.E.; Chaves, A.V. Dose-response effects of dietary pequi oil on fermentation characteristics and microbial population using a rumen simulation technique (Rusitec). Anaerobe 2017, 28, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Bauman, D.E.; Baumgard, L.H.; Corl, B.A.; Griinari, J.M. Biosynthesis of conjugated linoleic acid in ruminants. J. Anim. Sci. 2000, 77, 1–15. [Google Scholar] [CrossRef]
- Wu, Z.; OhajuruLa, O.A.; Palmquist, D.L. Ruminal synthesis, biohydrogenation, and digestibility of fatty acids by dairy cows. J. Dairy Sci. 1991, 74, 3025–3034. [Google Scholar] [CrossRef]
- Doreau, M.; Ferlay, A. Effects of dietary lipids on nitrogen metabolism in the rumen: A review. Livest. Prod. Sci. 1995, 43, 97–110. [Google Scholar] [CrossRef]
- Gillis, M.H.; Duckett, S.K.; Sackmann, J.R. Effects of supplemental rumen-protected conjugated linoleic acid or corn oil on fatty acid composition of adipose tissues in beef cattle. J. Anim. Sci. 2004, 82, 1419–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radcliffe, J.D.; Czajka-Narins, D.M.; Imrhan, V. Fatty acid composition of serum, adipose tissue, and liver in rats fed diets containing corn oil or cottonseed oil. Plant Foods Hum. Nutr. 2004, 59, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Suksombat, W.; Thanh, L.P.; Meeprom, C.; Mirattanaphrai, R. Effect of linseed oil supplementation on performance and milk fatty acid composition in dairy cows. Anim. Sci. J. 2016, 87, 1545–1553. [Google Scholar] [CrossRef] [PubMed]
- Hassanat, F.; Benchaar, C. Corn silage-based diet supplemented with increasing amounts of linseed oil: Effects on methane production, rumen fermentation, nutrient digestibility, nitrogen utilization, and milk production of dairy cows. J. Dairy Sci. 2021, 104, 5375–5390. [Google Scholar] [CrossRef]
- Gadeyne, F.; De Neve, N.; Vlaeminck, B.; Fievez, V. State of the art in rumen lipid protection technologies and emerging interfacial protein cross-linking methods. Eur. J. Lipid Sci. Technol. 2017, 119, 1600345. [Google Scholar] [CrossRef]
- Sukhija, P.S.; Palmquist, D.L. Dissociation of calcium soaps of long-chain fatty acids in rumen fluid. J. Dairy Sci. 1990, 73, 1784–1787. [Google Scholar] [CrossRef]
- Adesogan, A.T.; Krueger, N.K.; Kim, S.C. A novel, wireless, automated system for measuring fermentation gas production kinetics of feeds and its application to feed characterization. Anim. Feed Sci. Technol. 2005, 123–124, 211–223. [Google Scholar] [CrossRef]
- Lee, S.S.; Kim, D.H.; Paradhipta, D.H.V.; Lee, H.J.; Yoon, H.; Joo, Y.H.; Adesogan, A.T.; Kim, S.C. Effects of wormwood (Artemisia montana) essential oil on digestibility, fermentation indices, and microbial diversity in the rumen. Microorganisms 2020, 8, 1605. [Google Scholar] [CrossRef] [PubMed]
- Paradhipta, D.H.V.; Joo, Y.H.; Lee, H.J.; Lee, S.S.; Kwak, Y.G.; Han, O.K.; Kim, D.H.; Kim, S.C. Effects of wild and mutated inoculants on rye silage and its rumen fermentation indices. Asian-Australas. J. Anim. Sci. 2020, 33, 949–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carriquiry, M.; Weber, W.J.; Baumgard, L.H.; Crooker, B.A. In vitro biohydrogenation of four dietary fats. Anim. Feed Sci. Technol. 2008, 141, 339–355. [Google Scholar] [CrossRef]
- Honkanen, A.M.; Griinari, J.M.; Vanhatalo, A.; Ahvenjärvi, S.; Toivonen, V.; Shingfield, K.J. Characterization of the disappearance and formation of biohydrogenation intermediates during incubations of linoleic acid with rumen fluid in vitro. J. Dairy Sci. 2012, 95, 1376–1394. [Google Scholar] [CrossRef] [Green Version]
- Mauricio, R.M.; Mould, F.L.; Dhanoa, M.S.; Owen, E.; Channa, K.S.; Theodorou, M.K. A semi-automated in vitro gas production technique for ruminant feedstuff evaluation. Anim. Feed Sci. Technol. 1999, 79, 321–330. [Google Scholar] [CrossRef]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef]
- Muck, R.E.; Dickerson, J.T. Storage temperature effects on proteolysis in alfalfa silage. Trans. ASAE 1988, 31, 1005–1009. [Google Scholar] [CrossRef]
- Jenkins, T.C.; Thies, E.J.; Mosley, E.E. Direct methylation procedure for converting fatty amides to fatty acid methyl ester in feed and digesta samples. J. Agric. Food Chem. 2001, 49, 2142–2145. [Google Scholar] [CrossRef]
- Kim, E.T.; Kim, C.H.; Min, K.S.; Lee, S.S. Effects of plant extracts on microbial population, methane emission and ruminal fermentation characteristics in in vitro. Asian-Aust. J. Anim. Sci. 2012, 25, 806–811. [Google Scholar] [CrossRef]
- Denman, S.E.; McSweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Koike, S.; Kobayashi, Y. Development and use of competitive PCR assays for the rumen cellulolytic bacteria: Fibrobacter succinogens, Ruminococcus albus and Ruminococcus falvefaciens. FEMS Microbiol. Lett. 2001, 204, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Tajima, K.; Aminov, R.I.; Nagamine, T.; Matsui, H.; Nakamura, M.; Benno, Y. Diet-dependent shifts in the bacterial population of the rumen revealed with Real-Time PCR. Appl. Environ. Microb. 2001, 67, 2766–2774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denman, S.E.; Tomkins, N.W.; McSweeney, C.S. Quantitation and diversity analysis of ruminal methanogenic populations in response to the antimethanogenic compound bromochloromethane. FEMS Microbiol. Ecol. 2007, 62, 313–322. [Google Scholar] [CrossRef] [Green Version]
- Skillman, L.C.; Toovey, A.F.; Williams, A.J.; Wright, A.G. Development and validation of a Real-Time PCR method to quantify rumen protozoa and examination of variability between Endodinium populations in sheep offered a hay-based diet. Appl. Environ. Microb. 2006, 72, 200–206. [Google Scholar] [CrossRef] [Green Version]
- SAS User’s Guide, 8th ed.; SAS Inst., Inc.: Cary, NC, USA, 2002.
- Mao, S.Y.; Zhang, R.Y.; Wang, D.S.; Zhu, W.Y. Impact of subacute ruminal acidosis (SARA) adaptation on rumen microbiota in dairy cattle using pyrosequencing. Anaerobe 2013, 24, 12–19. [Google Scholar] [CrossRef]
- Sutton, J.D.; Knight, R.; McAllan, A.B.; Smith, R.H. Digestion and synthesis in the rumen of sheep given diets supplemented with free and protected oils. Br. J. Nutr. 1983, 49, 419–432. [Google Scholar] [CrossRef]
- Castañeda-Gutiérrez, E.; De Veth, M.J.; Lock, A.L.; Dwyer, D.A.; Murphy, K.D.; Bauman, D.E. Effect of supplementation with calcium salts of fish oil on n−3 fatty acids in milk fat. J. Dairy Sci. 2007, 90, 4149–4156. [Google Scholar] [CrossRef] [Green Version]
- Ferlay, A.; Legay, F.; Bauchart, D.; Poncet, C.; Doreau, M. Effect of a supply of raw or extruded rapeseeds on digestion in dairy cows. J. Anim. Sci. 1992, 70, 915–923. [Google Scholar] [CrossRef]
- Galbraith, H.; Miller, T.B.; Paton, A.M.; Thompson, J.K. Antibacterial activity of long-chain fatty acids and the reversal with calcium, magnesium, ergocalciferol and cholesterol. J. Appl. Bacteriol. 1971, 34, 803–813. [Google Scholar] [CrossRef]
- El-Hag, G.A.; Miller, T.B. Evaluation of whisky distillery by-products. VI. The reduction in digestibility of malt distiller’s grains by fatty acids and the interaction with calcium and other reversal agents. J. Sci. Food Agric. 1972, 23, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Roger, V.; Fonty, G.; Komisarczuk-Bony, S.; Gouet, P. Effects of physicochemical factors on the adhesion to cellulose Avicel of the ruminal bacteria Ruminococcus flavefaciens and Fibrobacter succinogenes subsp. Succinogenes. Appl. Environ. Microbiol. 1990, 56, 3081–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moate, P.J.; Chalupa, W.; Jenkins, T.C.; Boston, R.C. A model to describe ruminalmetabolism and intestinal absorption of long chain fatty acids. Anim. Feed Sci. Technol. 2004, 112, 79–105. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Treatment 1 | ||
|---|---|---|---|
| CO | LSO | Ca-Salt | |
| C10:0 | 0.01 | 0.01 | 0.02 |
| C12:0 | 0.02 | 0.02 | 0.10 |
| C14:0 | 0.06 | 0.06 | 0.28 |
| C14:1 | 0.00 | 0.00 | 0.01 |
| C15:0 | 0.01 | 0.02 | 0.03 |
| C16:0 | 11.5 | 5.30 | 12.4 |
| C16:1 | 0.04 | 0.03 | 0.05 |
| C17:1 | 0.03 | 0.05 | 0.00 |
| C18:0 | 1.94 | 3.84 | 3.59 |
| C18:1cis-9 | 29.7 | 19.2 | 50.8 |
| C18:2n-6 | 55.0 | 15.6 | 28.5 |
| C18:3n-3 | 1.22 | 55.7 | 2.32 |
| C20:0 | 0.40 | 0.04 | 0.53 |
| C20:1 | 0.00 | 0.00 | 0.99 |
| C20:3 | 0.12 | 0.15 | 0.32 |
| Target Organism | Sequence (5′-3′) | Annealing Temp (°C)/Size (bp) | Reference |
|---|---|---|---|
| Bacteria | F-CGGCAACGAGCGCAACCC R-CCATTGTAGCACGTGTGTAGCC | 55.3/130 | Denman and McSweeney [31] |
| Fibrobacter succinogenes | F-GTTCGGAATTACTGGGCGTAAA R-CGCCTGCCCCTGAACTATC | 61.0/121 | Denman and McSweeney [31] |
| Ruminococcus falvefaciens | F-GAACGGAGATAATTTGAGTTTACTTAGG R-CGGTCTCTGTATGTTATGAGGTATTACC | 57.7/132 | Denman and McSweeney [31] |
| Ruminococcus albus | F-CCCTAAAAGCAGTCTTAGTTCG R-CCTCCTTGCGGTTAGAACA | 59.3/175 | Koike and Kobayashi [32] |
| Streptococcus bovis | F-CTAATACCGCATAACAGCAT R-AGAAACTTCCTATCTCTAGG | 57.0/869 | Tajima et al. [33] |
| Methanogenic archaea | F-TTCGGTGGATCDCARAGRGC R-GBARGTCGWAWCCGTAGAATCC | 56.0/140 | Denman et al. [34] |
| Ciliates (Entodinium) | F-GAGCTAATACATGCTAAGGC R-CCCTCACTACAATCGAGATTTAAGG | 55.0/180 | Skillman et al. [35] |
| Treatments 1 | SEM | p-Value | |||
|---|---|---|---|---|---|
| CO | LSO | Ca-Salt | |||
| Total FA, mg/mL | 3.05 | 3.19 | 3.62 | 0.069 | 0.604 |
| C14:0, % total FA | 2.65 | 2.52 | 1.96 | 0.393 | 0.316 |
| C15:0, % total FA | 11.4 ab | 12.6 a | 8.55 b | 0.837 | 0.047 |
| C16:0, % total FA | 21.8 | 23.2 | 20.1 | 1.015 | 0.115 |
| C16:1, % total FA | 0.89 | 1.34 | 0.63 | 0.174 | 0.059 |
| C17:0, % total FA | 0.58 | 0.55 | 0.47 | 0.070 | 0.368 |
| C18:0, % total FA | 24.6 | 24.5 | 25.6 | 0.918 | 0.501 |
| C18:1cis-9, % total FA | 14.7 b | 14.9 b | 21.4 a | 1.360 | 0.026 |
| C18:2n-6, % total FA | 20.0 a | 9.33 b | 18.7 a | 1.643 | 0.013 |
| C18:3n-3, % total FA | 0.92 b | 8.78 a | 1.25 b | 0.851 | 0.004 |
| C20:0, % total FA | 1.76 | 1.52 | 0.85 | 1.011 | 0.686 |
| C22:0, % total FA | 0.64 | 0.64 | 0.48 | 0.087 | 0.245 |
| SFA 2, % total FA | 63.5 | 65.7 | 58.1 | 2.702 | 0.135 |
| MUFA, % total FA | 15.6 b | 16.2 b | 22.0 a | 1.325 | 0.029 |
| PUFA, % total FA | 20.9 | 18.1 | 19.9 | 1.911 | 0.435 |
| SFA: PUFA | 3.07 | 3.65 | 2.92 | 0.436 | 0.347 |
| n-6/n-3 | 21.7 a | 1.06 b | 15.0 a | 2.216 | 0.006 |
| Treatments 1 | SEM | p-Value | |||
|---|---|---|---|---|---|
| CO | LSO | Ca-Salt | |||
| pH | 6.44 b | 6.46 ab | 6.52 a | 0.041 | 0.024 |
| NH3-N, mg/100 mL | 20.0 | 20.2 | 20.2 | 1.157 | 0.976 |
| Total VFA, mmol/L | 58.9 | 60.1 | 57.0 | 1.641 | 0.065 |
| Acetate, % mol | 49.6 b | 50.4 ab | 52.2 a | 0.997 | 0.013 |
| Propionate, % mol | 38.5 a | 36.9 b | 36.9 b | 0.630 | 0.007 |
| Butyrate, % mol | 11.0 | 11.9 | 10.6 | 0.685 | 0.060 |
| Acetate: propionate | 1.28 b | 1.36 ab | 1.41 a | 0.048 | 0.015 |
| Gas volume, mL/g DM | 38.8 | 37.4 | 37.7 | 0.816 | 0.059 |
| CH4, mg/g | 17.2 | 15.2 | 17.1 | 1.171 | 0.209 |
| CO2, mg/g | 2.61 | 2.51 | 2.59 | 0.271 | 0.879 |
| Treatments 1 | SEM | p-Value | |||
|---|---|---|---|---|---|
| CO | LSO | Ca-Salt | |||
| Methanogenic archaea | 0.72 b | 1.43 a | 1.06 b | 0.145 | 0.003 |
| Ciliates | 0.36 a | 0.13 b | 0.22 ab | 0.111 | 0.018 |
| Fibrolytic bacteria | |||||
| F. succinogens | 0.15 b | 0.06 b | 0.45 a | 0.067 | <0.001 |
| R. albus | 0.43 b | 0.04 c | 0.82 a | 0.146 | <0.001 |
| S. bovis | 0.39 | 1.13 | 0.68 | 0.324 | 0.083 |
| R. flavefaciens | 0.48 b | 0.34 b | 1.02 a | 0.276 | 0.005 |
| Treatments 1 | SEM | p-Value | |||
|---|---|---|---|---|---|
| CO | LSO | Ca-salt | |||
| Total FA, mg/mL | 4.04 | 5.02 | 5.51 | 0.076 | 0.086 |
| C14:0, % total FA | 8.27 a | 6.28 b | 5.24 b | 0.494 | 0.007 |
| C15:0, % total FA | 8.00 a | 5.02 b | 5.46 b | 0.785 | 0.035 |
| C16:0, % total FA | 27.8 a | 22.2 b | 23.0 b | 0.876 | 0.005 |
| C16:1, % total FA | 3.04 a | 1.75 b | 2.06 b | 0.161 | 0.003 |
| C17:0, % total FA | 1.06 | 0.80 | 0.81 | 0.082 | 0.058 |
| C18:0, % total FA | 25.4 b | 33.8 a | 20.1 c | 0.938 | 0.002 |
| C18:1cis-9, % total FA | 14.2 b | 15.0 b | 28.2 a | 2.339 | 0.004 |
| C18:2n-6, % total FA | 11.0 a | 5.41 b | 13.1 a | 0.887 | 0.002 |
| C18:3n-3, % total FA | 0.65 c | 9.24 a | 1.24 b | 0.066 | <0.001 |
| C20:0, % total FA | 0.62 ab | 0.53 b | 0.68 a | 0.034 | 0.020 |
| SFA, % total FA | 71.2 a | 68.6 a | 55.3 b | 3.053 | 0.008 |
| MUFA, % total FA | 17.2 b | 16.8 b | 30.3 a | 2.204 | 0.004 |
| PUFA, % total FA | 11.6 b | 14.6 a | 14.4 a | 0.910 | 0.048 |
| SFA: PUFA | 6.14 a | 4.69 b | 3.88 b | 0.626 | 0.016 |
| n-6/n-3 | 16.9 a | 0.58 b | 10.6 a | 1.045 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amanullah, S.M.; Lee, S.-S.; Paradhipta, D.H.V.; Joo, Y.-H.; Kim, D.-H.; Seong, P.-N.; Jeong, S.-M.; Kim, S.-C. Impact of Oil Sources on In Vitro Fermentation, Microbes, Greenhouse Gas, and Fatty Acid Profile in the Rumen. Fermentation 2022, 8, 242. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050242
Amanullah SM, Lee S-S, Paradhipta DHV, Joo Y-H, Kim D-H, Seong P-N, Jeong S-M, Kim S-C. Impact of Oil Sources on In Vitro Fermentation, Microbes, Greenhouse Gas, and Fatty Acid Profile in the Rumen. Fermentation. 2022; 8(5):242. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050242
Chicago/Turabian StyleAmanullah, Sardar Muhammad, Seong-Shin Lee, Dimas Hand Vidya Paradhipta, Young-Hoo Joo, Dong-Hyeon Kim, Pil-Nam Seong, Seung-Min Jeong, and Sam-Churl Kim. 2022. "Impact of Oil Sources on In Vitro Fermentation, Microbes, Greenhouse Gas, and Fatty Acid Profile in the Rumen" Fermentation 8, no. 5: 242. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050242