
Growth and Competitive Infection Behaviors of Bradyrhizobium japonicum and Bradyrhizobium elkanii at Different Temperatures


Abstract
:1. Introduction
2. Materials and Methods
2.1. Effect of Temperature on Growth of Bradyrhizobium spp. in Liquid Culture
2.2. Effect of Temperature on Competitive Infection of Bradyrhizobium spp. in Soybean
2.3. Statistical Analysis
3. Results
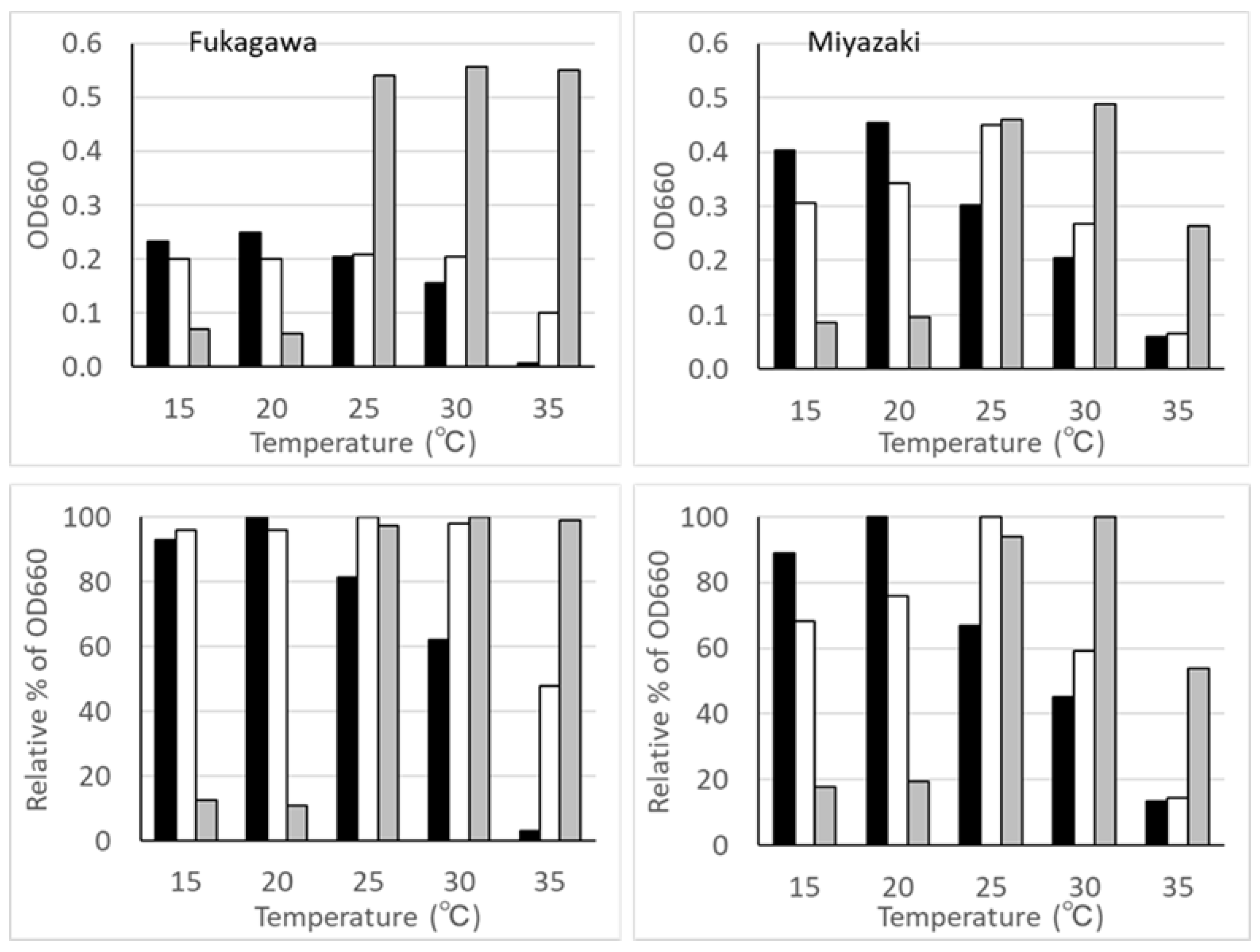
3.1. Effect of Temperature on Growth of Bradyrhizobium spp. Strains in Liquid Culture
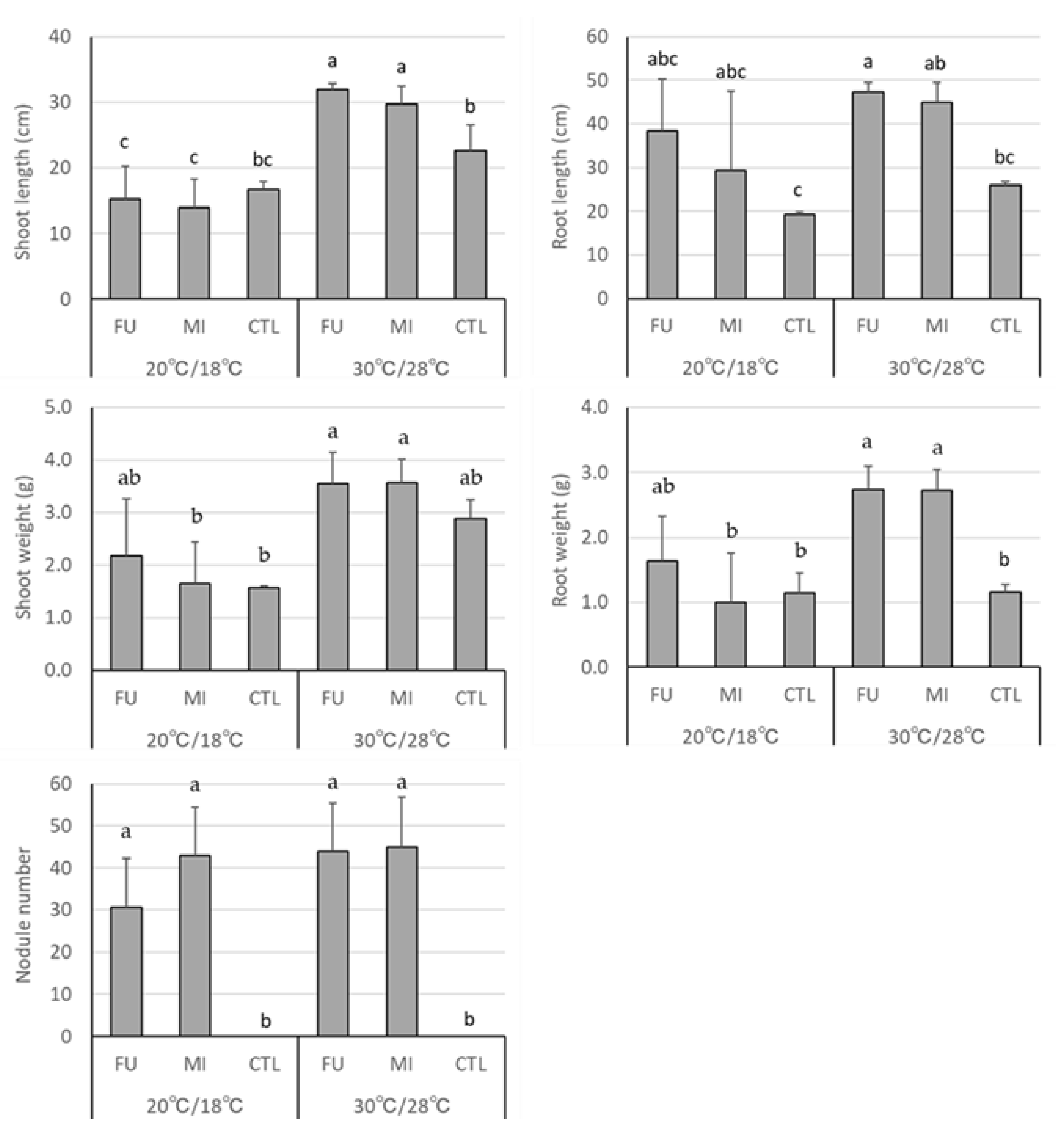
3.2. Effect of Temperature on Growth and Nodule Number of Soybean Inoculated with a Set of Bradyrhizobium spp. Strains
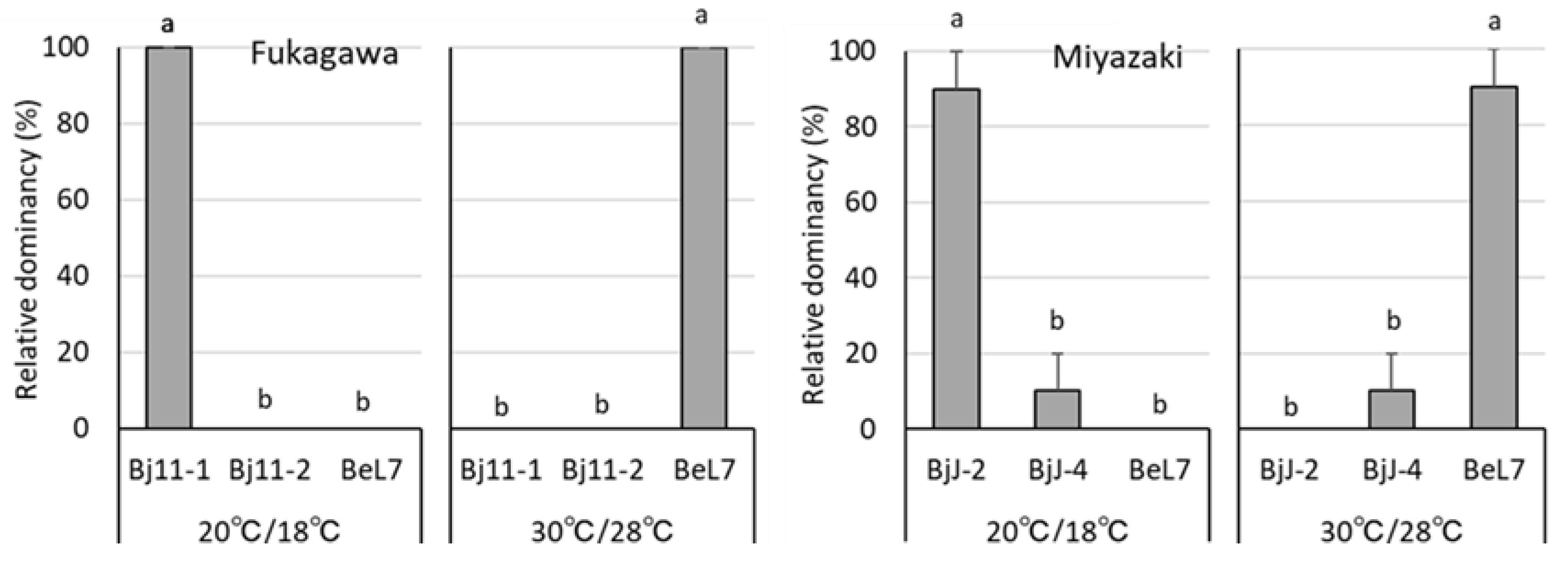
3.3. Effect of Temperature on Soybean Nodule Composition of Inoculated Bradyrhizobium spp. Stains
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hymowitz, T. On the domestication of the soybean. Econ. Bot. 1970, 24, 408–421. [Google Scholar] [CrossRef]
- Hymowitz, T.; Singh, R.J. Taxonomy and speciation. In Soybeans, Improvement, Production and Uses, 2nd. ed.; Wilcox, J.R., Ed.; American Society of Agronomy: Madison, WI, USA, 1987; pp. 23–48. [Google Scholar]
- Hungria, M.; Mendes, I.C.; de Bruijn, F. Nitrogen fixation with soybean: The perfect symbiosis? In Biological Nitrogen Fixation; De Bruijn, F., Ed.; Wiley Publisher: Hoboken, NJ, USA, 2015; Volume 2, pp. 1005–1019. [Google Scholar]
- Jordan, D.C. NOTES: Transfer of Rhizobium japonicum Buchanan 1980 to Bradyrhizobium gen. nov., a Genus of Slow-Growing, Root Nodule Bacteria from Leguminous Plants. Int. J. Syst. Bacteriol. 1982, 32, 136–139. [Google Scholar] [CrossRef]
- Kuykendall, L.D.; Saxena, B.N.; Devine, T.E.; Udell, S.E. Genetic diversity in Bradyrhizobium japonicum Jordan 1982 and a proposal for Bradyrhizobium elkanii sp.nov. Can. J. Microbiol. 1992, 38, 501–505. [Google Scholar] [CrossRef]
- Hungria, M.; Vargas, M.A.T. Environmental factors impacting N2 fixation in legumes grown in the tropics, with an emphasis on Brazil. Field Crop. Res. 2000, 65, 151–164. [Google Scholar] [CrossRef]
- Hafiz, H.R.; Salehin, A.; Adachi, F.; Omichi, M.; Saeki, Y.; Yamamoto, A.; Hayashi, S.; Itoh, K. Latitudinal Characteristic Nodule Composition of Soybean-Nodulating Bradyrhizobia: Temperature-Dependent Proliferation in Soil or Infection? Horticulturae 2021, 7, 22. [Google Scholar] [CrossRef]
- Saeki, Y.; Aimi, N.; Tsukamoto, S.; Yamakawa, T.; Nagatomo, Y.; Akao, S. Diversity and geographical distribution of indigenous soybean-nodulating bradyrhizobia in Japan. Soil Sci. Plant Nutr. 2006, 52, 418–426. [Google Scholar] [CrossRef]
- Shiro, S.; Matsuura, S.; Saiki, R.; Sigua, G.C.; Yamamoto, A.; Umehara, Y.; Hayashi, M.; Saeki, Y. Genetic Diversity and Geographical Distribution of Indigenous Soybean-Nodulating Bradyrhizobia in the United States. Appl. Environ. Microbiol. 2013, 79, 3610–3618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, D.; Kaneto, M.; Itoh, K.; Suyama, K.; Pokharel, B.B.; Gaihre, Y.K. Genetic diversity of soybean-nodulating rhizobia in Nepal in relation to climate and soil properties. Plant Soil 2012, 357, 131–145. [Google Scholar] [CrossRef]
- Kluson, R.A.; Kenworthy, W.J.; Weber, D.F. Soil temperature effects on competitiveness and growth ofRhizobium japonicum and on Rhizobium-induced chlorosis of soybeans. Plant Soil 1986, 95, 201–207. [Google Scholar] [CrossRef]
- Suzuki, Y.; Adhikari, D.; Itoh, K.; Suyama, K. Effects of temperature on competition and relative dominance of Bradyrhizobium japonicum and Bradyrhizobium elkanii in the process of soybean nodulation. Plant Soil 2013, 374, 915–924. [Google Scholar] [CrossRef]
- Shiro, S.; Kuranaga, C.; Yamamoto, A.; Sameshima-Saito, R.; Saeki, Y. Temperature-Dependent Expression of NodC and Community Structure of Soybean-Nodulating Bradyrhizobia. Microbes Environ. 2016, 31, 27–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callow, J.A.; Vincent, J.M. A Manual for the Practical Study of Root-Nodule Bacteria. In IBP Handbook No. 15; Blackwell Scientific Publishers: Oxford, UK, 1970; p. 164. [Google Scholar]
- Leonard, L.T. A Simple Assembly for Use in the Testing of Cultures of Rhizobia. J. Bacteriol. 1943, 45, 523–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. J. Plant. Physiol. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Shiro, S.; Yamamoto, A.; Umehara, Y.; Hayashi, M.; Yoshida, N.; Nishiwaki, A.; Yamakawa, T.; Saeki, Y. Effect ofRjGenotype and Cultivation Temperature on the Community Structure of Soybean-Nodulating Bradyrhizobia. Appl. Environ. Microbiol. 2011, 78, 1243–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeki, Y.; Kaneko, A.; Hara, T.; Suzuki, K.; Yamakawa, T.; Nguyen, M.T.; Nagatomo, Y.; Akao, S. Phylogenetic Analysis of Soybean-Nodulating Rhizobia Isolated from Alkaline Soils in Vietnam. Soil Sci. Plant. Nutr. 2005, 51, 1043–1052. [Google Scholar] [CrossRef] [Green Version]
- Freed, R. MSTAT: A software program for plant breeder. In Principles of Plant Genetics and Breeding, 2nd ed.; Acquaah, G., Ed.; Blackwell Publishing: Malden, MA, USA, 2007; Volume 1, pp. 426–431. [Google Scholar]
- Saeki, Y.; Ozumi, S.; Yamamoto, A.; Umehara, Y.; Hayashi, M.; Sigua, G.C. Changes in Population Occupancy of Bradyrhizobia under Different Temperature Regimes. Microbes Environ. 2010, 25, 309–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoyanova, J. Growth, nodulation and nitrogen fixation in soybean as affected by air humidity and root temperature. Biol. Plant. 1996, 38, 537–544. [Google Scholar] [CrossRef]
- Zhang, F.; Smith, D.L. Effects of low root zone temperatures on the early stages of symbiosis establishment between soybean [Glycine max (L.) Merr.] and Bradyrhizobium japonicum. J. Exp. Bot. 1994, 45, 1467–1473. [Google Scholar] [CrossRef]
- Montañez, A.; Danso, S.; Hardarson, G. The effect of temperature on nodulation and nitrogen fixation by five Bradyrhizobium japonicum strains. Appl. Soil Ecol. 1995, 2, 165–174. [Google Scholar] [CrossRef]

 ); Miyazaki strains: B. japonicum Kh 16-43 (Bj10J-2) (■), B. japonicum Kh 16-64 (Bj10J-4) (□), and B. elkanii Kh 16-7 (BeL7) ( ).
); Miyazaki strains: B. japonicum Kh 16-43 (Bj10J-2) (■), B. japonicum Kh 16-64 (Bj10J-4) (□), and B. elkanii Kh 16-7 (BeL7) ( ).
); Miyazaki strains: B. japonicum Kh 16-43 (Bj10J-2) (■), B. japonicum Kh 16-64 (Bj10J-4) (□), and B. elkanii Kh 16-7 (BeL7) ( ).
); Miyazaki strains: B. japonicum Kh 16-43 (Bj10J-2) (■), B. japonicum Kh 16-64 (Bj10J-4) (□), and B. elkanii Kh 16-7 (BeL7) ( ).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain a | Closest 16 rDNA | ITS Group b | Accession Number c |
|---|---|---|---|
| Hh 16-9 | B. japonicum Bj11 | Bj11-1 | LC582854, LC579849 |
| Hh-16-25 | B. japonicum Bj11 | Bj11-2 | LC582860, LC579855 |
| Hk 16-7 | B. elkanii L7 | BeL7 | LC582891, LC579886 |
| Kh 16-43 | B. japonicum S10J | BjS10J-2 | LC582874, LC579869 |
| Kh 16-64 | B. japonicum S10J | BjS10J-4 | LC582887, LC579882 |
| Kh 16-7 | B. elkanii L7 | BeL7 | LC582901, LC579896 |
| Location | Latitude (°N) | Longitude (°E) | Temperature (°C) a | Rainfall (mm) a |
|---|---|---|---|---|
| Fukagawa | 43.71 | 142.01 | 16–26/16–26 (14–24/17–27) b | 432/243 |
| Miyazaki | 31.82 | 131.41 | 24–32/24–31 (25–32/25–33) | 240/860 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hafiz, M.H.R.; Salehin, A.; Itoh, K. Growth and Competitive Infection Behaviors of Bradyrhizobium japonicum and Bradyrhizobium elkanii at Different Temperatures. Horticulturae 2021, 7, 41. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7030041
Hafiz MHR, Salehin A, Itoh K. Growth and Competitive Infection Behaviors of Bradyrhizobium japonicum and Bradyrhizobium elkanii at Different Temperatures. Horticulturae. 2021; 7(3):41. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7030041
Chicago/Turabian StyleHafiz, Md Hafizur Rahman, Ahsanul Salehin, and Kazuhito Itoh. 2021. "Growth and Competitive Infection Behaviors of Bradyrhizobium japonicum and Bradyrhizobium elkanii at Different Temperatures" Horticulturae 7, no. 3: 41. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7030041