
Comparative Transcriptome Analysis Reveals Different Low-Nitrogen-Responsive Genes in Pepper Cultivars



 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Screening of Pepper Cultivars with Different Low-N Tolerance
2.2. RNA Preparation and Sequencing
2.3. Transcriptome De Novo Assembly, Gene Functional Annotation, and Differentially Expressed Genes (DEGs) Analyses
2.4. Quantitative Real-Time PCR (qRT-PCR) Analysis
3. Results
3.1. Screening for Pepper Cultivars with Contrasting Low-N Tolerance
3.2. RNA-Seq Analysis of Two Pepper Cultivars with Contrasting Low-N Tolerances
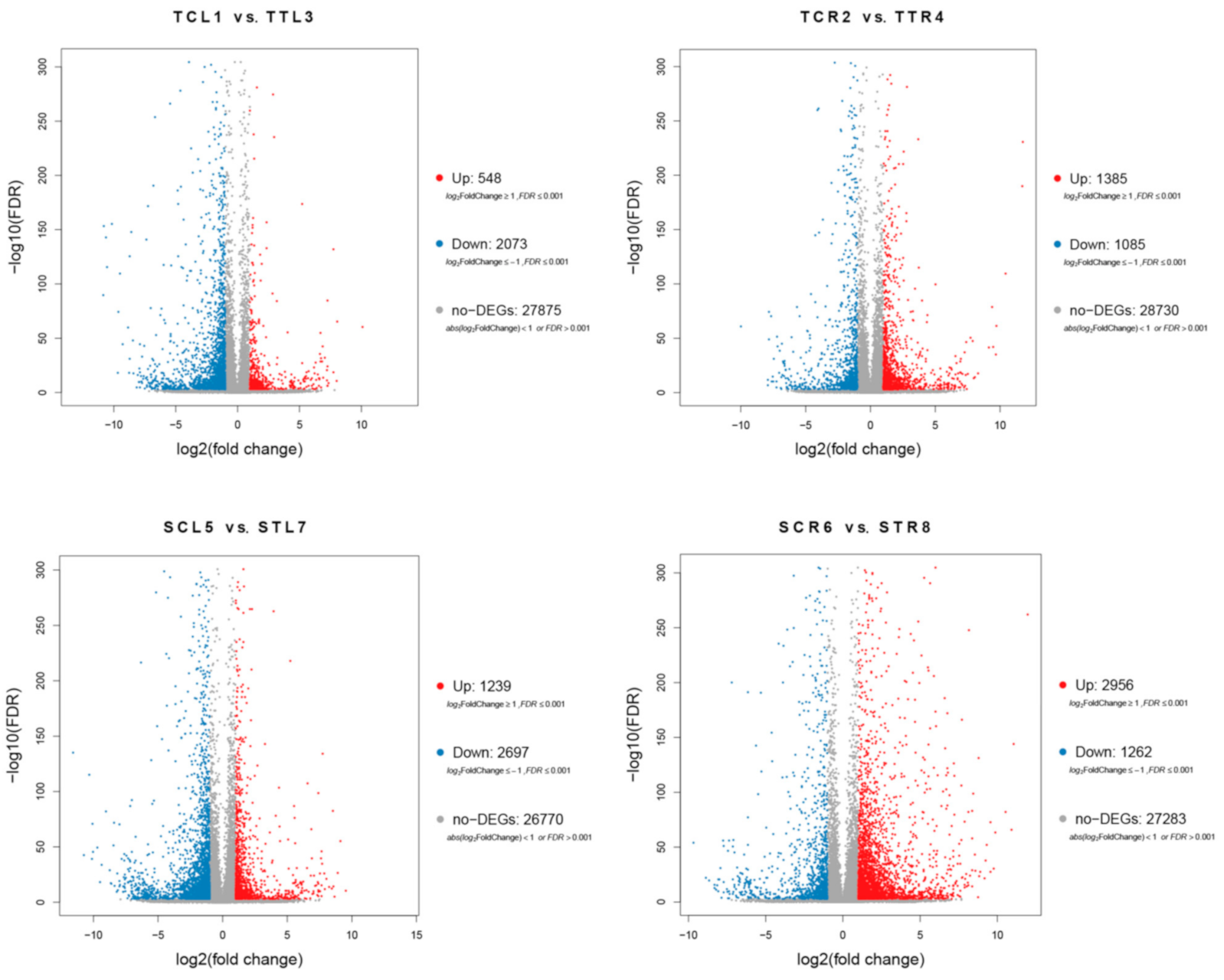
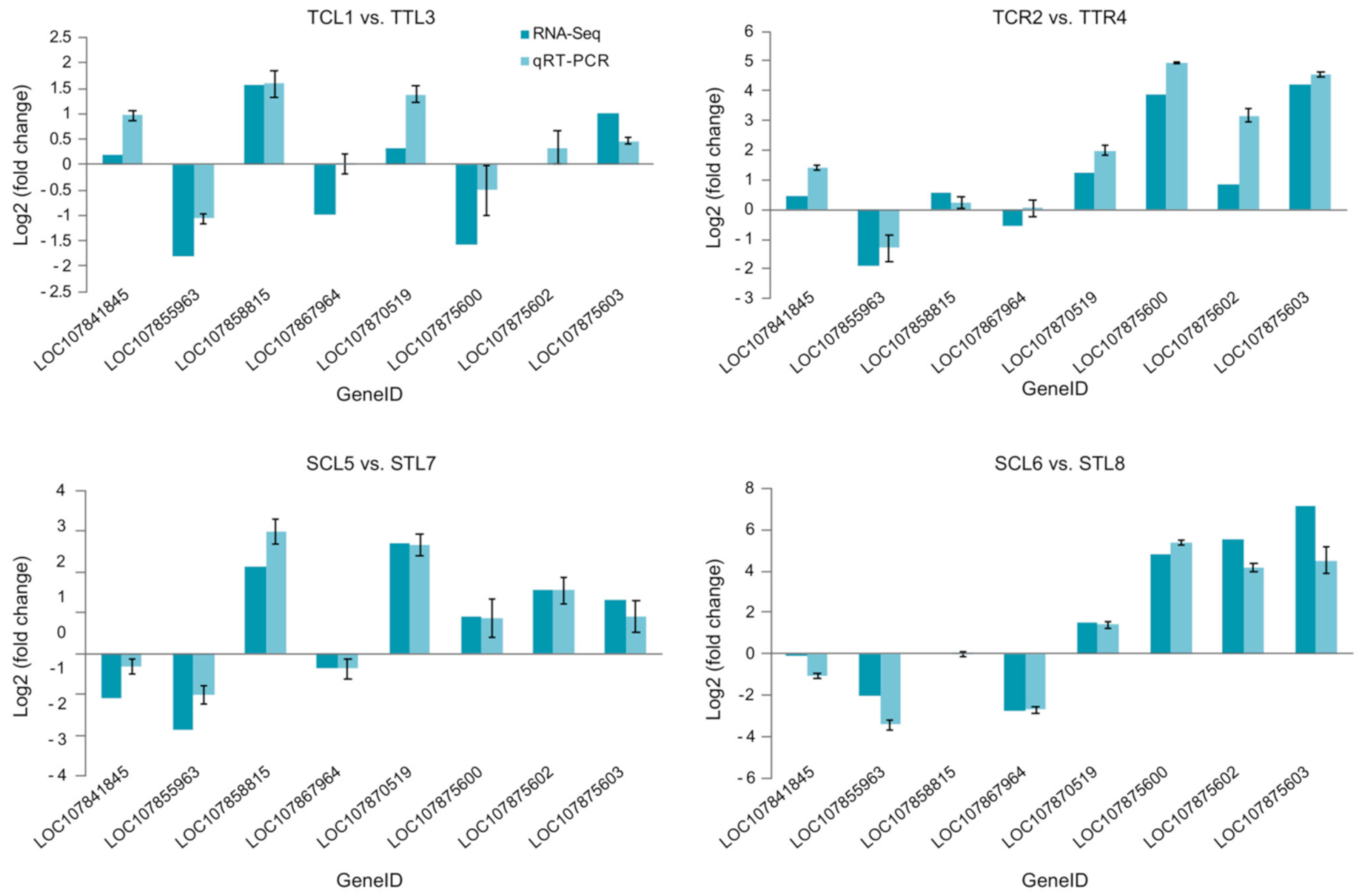
3.3. Identification of DEGs and Validation of RNA-Seq by qRT-PCR
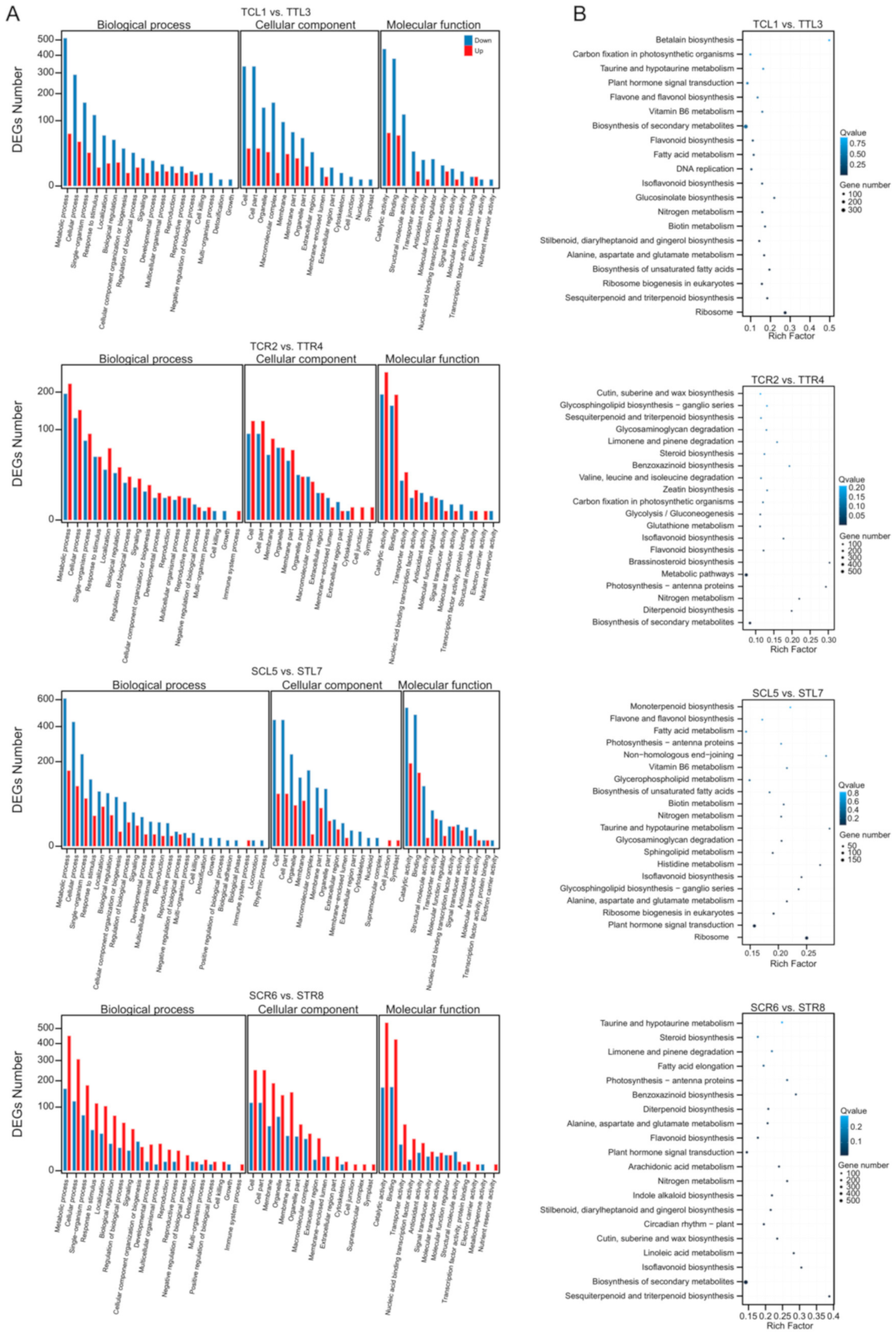
3.4. GO and KEGG Enrichment Analysis of DEGs
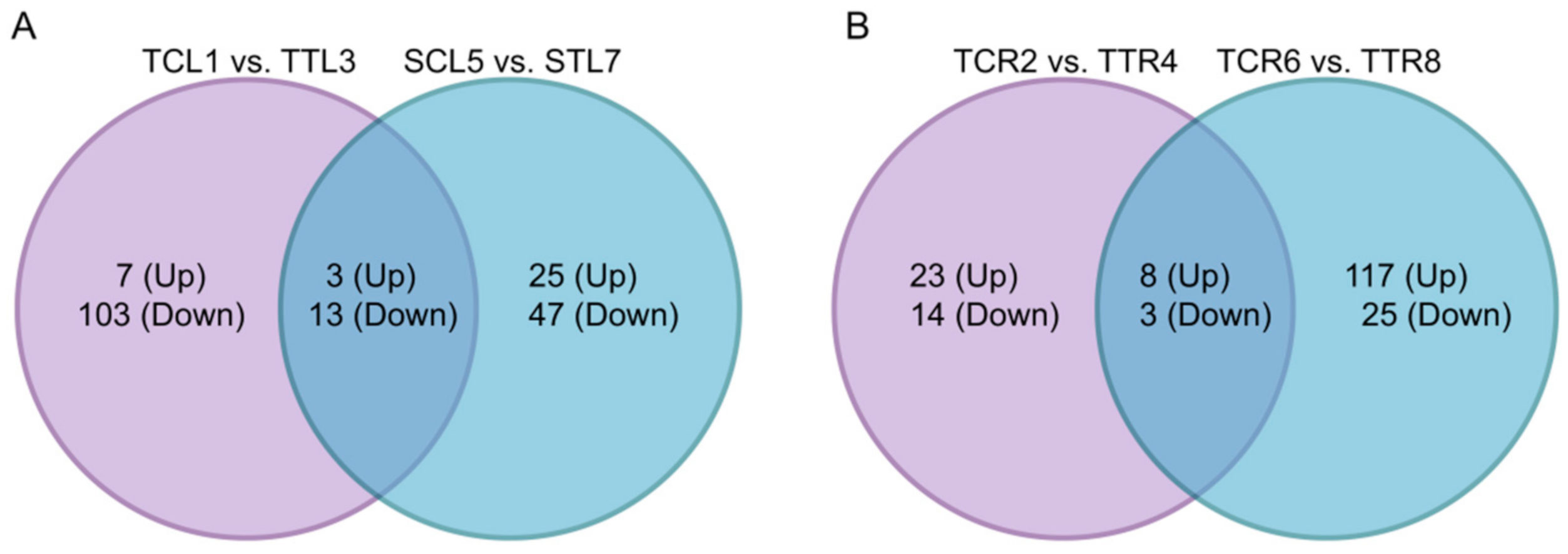
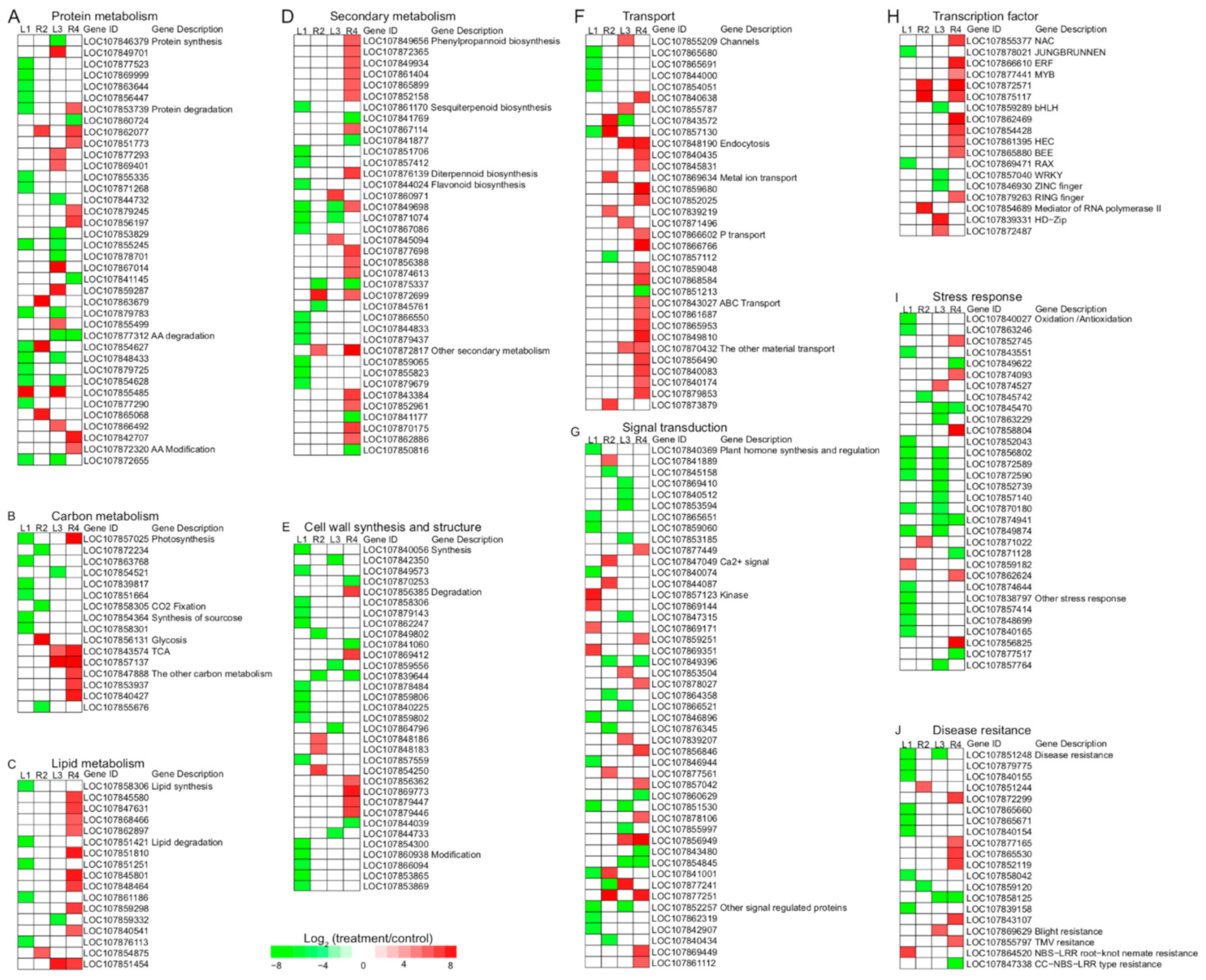
3.5. Different DEGs between Two Cultivars
3.5.1. Protein Metabolism
3.5.2. Carbon and Lipid Metabolism
3.5.3. Secondary Metabolism
3.5.4. Cell Wall Synthesis and Structure
3.5.5. Transport
3.5.6. Signal Transduction and Transcription Factor
3.5.7. Stress Response and Disease Resistance
4. Discussion
4.1. Unique DEGs in the Low-N-Tolerant Genotype
4.2. Unique DEGs in the Low-N Sensitive Genotype
4.3. P and N Crosstalk
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Q.; Chen, X.; Wu, K.; Fu, X. Nitrogen signaling and use efficiency in plants: What’s new? Curr. Opin. Plant Biol. 2015, 27, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Krapp, A.; David, L.C.; Chardin, C.; Girin, T.; Marmagne, A.; Leprince, A.S.; Chaillou, S.; Ferrario-Méry, S.; Meyer, C.; Daniel-Vedele, F. Nitrate transport and signalling in Arabidopsis. J. Exp. Bot. 2014, 65, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, M.; Bi, Y.M.; Zhu, T.; Rothstein, S.J. Genome-wide analysis of Arabidopsis responsive transcriptome to nitrogen limitation and its regulation by the ubiquitin ligase gene NLA. Plant Mol. Biol. 2007, 65, 775–797. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Yang, T.; Zhang, Z.; Li, F.; Chen, Q.; Sun, J.; Shi, C.; Deng, W.; Tao, M.; Tai, Y.; et al. Identification and characterization of cationic amino acid transporters (CATs) in tea plant (Camellia sinensis). Plant Growth Regul. 2018, 84, 57–69. [Google Scholar] [CrossRef]
- Liu, S.; Wang, D.; Mei, Y.; Xia, T.; Xu, W.; Zhang, Y.; You, X.; Zhang, X.; Li, L.; Wang, N.N. Overexpression of GmAAP6a enhances tolerance to low nitrogen and improves seed nitrogen status by optimizing amino acid partitioning in soybean. Plant Biotechnol. J. 2020, 18, 1749–1762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Zhang, Z. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayodele, O.J.; Alabi, E.O.; Aluko, M. Nitrogen fertilizer effects on growth, yield and chemical composition of hot pepper (rodo). Int. J. Agric. Sci. 2015, 8, 666–673. [Google Scholar]
- Wahocho, N.A.; Zeshan Ahmed, S.; Jogi, Q.; Talpur, K.H.; Leghari, S.J. Growth and productivity of chilli (Capsicum annuum L.) under various nitrogen levels. Sci. Int. Lahore 2016, 28, 1321–1326. [Google Scholar]
- Zhu, J.H.; Li, X.L.; Christie, P.; Li, J.L. Environmental implications of low nitrogen use efficiency in excessively fertilized hot pepper (Capsicum frutescens L.) cropping systems. Agric. Ecosyst. Environ. 2005, 111, 70–80. [Google Scholar] [CrossRef] [Green Version]
- Amare, T.; Nigussie, D.; Kebede, W.T. Performance of hot pepper (Capsicum annuum) varieties as influenced by nitrogen and phosphorus fertilizers at Bure, Upper Watershed of the Blue Nile in Northwestern Ethiopia. Int. J. Agric. Sci. 2013, 3, 599–608. [Google Scholar]
- Sachiko, N.; Toriyama, K.; Yoshimichi, F. Genetic variations in dry matter production and physiological nitrogen use efficiency in rice (Oryza sativa L.) varieties. Breed. Sci. 2009, 59, 269–276. [Google Scholar]
- Anbessa, Y.; Juskiw, P.; Good, A.; Nyachiro, J.; Helm, J. Genetic variability in nitrogen use efficiency of spring barley. Crop Sci. 2009, 49, 1259–1269. [Google Scholar] [CrossRef]
- Górny, A.G.; Banaszak, Z.; Ługowska, B.; Ratajczak, D. Inheritance of the efficiency of nitrogen uptake and utilization in winter wheat (Triticum aestivum L.) under diverse nutrition levels. Euphytica 2010, 177, 191–206. [Google Scholar] [CrossRef] [Green Version]
- Bouchet, A.-S.; Laperche, A.; Bissuel-Belaygue, C.; Snowdon, R.; Nesi, N.; Stahl, A. Nitrogen use efficiency in rapeseed. Agron. Sustain. Dev. 2016, 36, 38. [Google Scholar] [CrossRef]
- Rodrigues, M.C.; Rezende, W.M.; Silva, M.E.J.; Faria, S.V.; Zuffo, L.T.; Galvão, J.C.C.; DeLima, R.O. Genotypic variation and relationships among nitrogen-use efficiency and agronomic traits in tropical maize inbred lines. Genet. Mol. Res. 2017, 16, gmr16039757. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Fu, X.; Wang, X.; Gui, H.; Dong, Q.; Pang, N.; Wang, Z.; Zhang, X.; Song, M. Identification and screening of nitrogen-efficient cotton genotypes under low and normal nitrogen environments at the seedling stage. J. Cotton. Res. 2018, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Qian, Q.; Wu, K.; Luo, J.; Wang, S.; Zhang, C.; Fu, X. Heterotrimeric G proteins regulate nitrogen-use efficiency in rice. Nat. Genet. 2014, 46, 652–656. [Google Scholar] [CrossRef]
- Hu, B.; Wang, W.; Ou, S.; Tang, J.; Li, H.; Che, R.; Chu, C. Variation in NRT1.1B contributes to nitrate-use divergence between rice subspecies. Nat. Genet. 2015, 47, 834–838. [Google Scholar] [CrossRef]
- Fan, X.; Tang, Z.; Tan, Y.; Zhang, Y.; Luo, B.; Yang, M.; Lian, X.; Shen, Q.; Miller, A.; Xu, G. Overexpression of a pH-sensitive nitrate transporter in rice increases crop yields. Proc. Natl. Acad. Sci. USA 2016, 113, 7118–7123. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Nian, J.; Xie, X.; Yu, H.; Zhang, J.; Bai, J.; Zuo, J. Genetic variations in ARE1 mediate grain yield by modulating nitrogen utilization in rice. Nat. Commun. 2018, 9, 735. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Hu, B.; Yuan, D.; Liu, Y.; Che, R.; Hu, Y.; Chu, C. Expression of the nitrate transporter gene OsNRT1.1A/OsNPF6.3 confers high yield and early maturation in rice. Plant Cell 2018, 30, 638–651. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Ye, J.; Yao, X.; Zhao, P.; Xuan, W.; Tian, Y.; Wan, J. Genome-wide associated study identifies NAC42-activated nitrate transporter conferring high nitrogen use efficiency in rice. Nat. Commun. 2019, 10, 5279. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Yang, X.; Yu, H.; Jiang, W.; Sun, N.; Liu, X.; Gu, X. RNA-Seq-based transcriptome profiling of early nitrogen deficiency response in cucumber seedlings provides new insight into the putative nitrogen regulatory network. Plant Cell Physiol. 2015, 56, 455–467. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Liang, Z.; Ge, M.; Qi, W.; Zhang, T.; Lin, F.; Peng, Z.; Zhao, H. Genome-wide identification and functional prediction of nitrogen-responsive intergenic and intronic long non-coding RNAs in maize (Zea mays L.). BMC Genom. 2016, 17, 350. [Google Scholar] [CrossRef] [Green Version]
- Curci, P.L.; Cigliano, R.A.; Zuluaga, D.L.; Janni, M.; Sanseverino, W.; Sonnante, G. Transcriptomic response of durum wheat to nitrogen starvation. Sci. Rep. 2017, 7, 1176. [Google Scholar] [CrossRef]
- Kuang, Q.; Zhang, S.; Wu, P.; Chen, Y.; Li, M.; Jiang, H.; Wu, G. Global gene expression analysis of the response of physic nut (Jatropha curcas L.) to medium- and long-term nitrogen deficiency. PLoS ONE 2017, 12, e0182700. [Google Scholar]
- Xin, W.; Zhang, L.; Zhang, W.; Gao, J.; Yi, J.; Zhen, X.; Li, Z.; Zhao, Y.; Peng, C.; Zhao, C. An integrated analysis of the rice transcriptome and metabolome reveals differential regulation of carbon and nitrogen metabolism in response to nitrogen availability. Int. J. Mol. Sci. 2019, 20, 2349. [Google Scholar] [CrossRef] [Green Version]
- Hao, Q.N.; Zhou, X.A.; Sha, A.H.; Wang, C.; Zhou, R.; Chen, S.L. Identification of genes associated with nitrogen use efficiency by genome-wide transcriptional analysis of two soybean genotypes. BMC Genom. 2011, 12, 525. [Google Scholar] [CrossRef] [Green Version]
- Gelli, M.; Duo, Y.D.; Konda, A.R.; Zhang, C.; Holding, D.; Dweikat, I. Identification of differentially expressed genes between sorghum genotypes with contrasting nitrogen stress tolerance by genome-wide transcriptional profiling. BMC Genom. 2014, 15, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, X.; Zeng, J.; Ye, L.; Chen, G.; Han, Z.; Shah, J.; Zhang, G. Transcriptome profiling analysis for two Tibetan wild barley genotypes in responses to low nitrogen. BMC Plant Biol. 2016, 16, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Xiang, F.; Zhong, M.; Zhou, L.; Liu, H.; Li, S.; Wang, X. Transcriptome and metabolite analysis identifies nitrogen utilization genes in tea plant (Camellia sinensis). Sci. Rep. 2017, 7, 1693. [Google Scholar]
- Sinha, S.K.; Sevanthi, V.A.M.; Chaudhary, S.; Tyagi, P.; Venkadesan, S.; Rani, M.; Mandal, P.K. Transcriptome analysis of two rice varieties contrasting for nitrogen use efficiency under chronic N starvation reveals differences in chloroplast and starch metabolism-related genes. Genes 2018, 9, 206. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, A.; Dong, Q.; Wang, X.R.; Gui, H.P.; Zhang, H.H.; Zhang, X.L.; Song, M.Z. Transcriptome analysis reveals differences in key genes and pathways regulating carbon and nitrogen metabolism in cotton genotypes under N starvation and resupply. Int. J. Mol. Sci. 2020, 21, 1500. [Google Scholar]
- Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/genome/10896?genome_assembly_id=227510 (accessed on 3 August 2017).
- Available online: ftp://ftp.ncbi.nih.gov/blast/db (accessed on 3 August 2017).
- Available online: http://www.geneontology.org/ (accessed on 3 August 2017).
- Available online: http://www.genome.jp/kegg/ (accessed on 3 August 2017).
- Albrech, C.; Russinova, E.; Kemmerling, B.; Kwaaitaal, M.; de Vries, S.C. Arabidopsis somatic embryogenesis receptor kinase proteins serve brassinosteroid-dependent and -independent signaling pathways. Plant Physiol. 2008, 148, 611–619. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yu, X.; Song, L.; An, C. ABI4 activates DGAT1 expression in Arabidopsis seedlings during nitrogen deficiency. Plant Physiol. 2011, 156, 873–883. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.Y.; Burgos, A.; Ma, L.; Zhang, Q.; Tang, D.; Ruan, J. Lipidomics analysis unravels the effect of nitrogen fertilization on lipid metabolism in tea plant (Camellia sinensis L.). BMC Plant Biol. 2017, 17, 165. [Google Scholar] [CrossRef] [Green Version]
- D’Hondt, K.; Heese-Peck, A.; Riezman, H. Protein and lipid requirements for endocytosis. Annu. Rev. Genet. 2000, 34, 255–295. [Google Scholar] [CrossRef]
- Chen, X.; Irani, N.G.; Friml, J. Clathrin-mediated endocytosis: The gateway into plant cells. Curr. Opin. Plant Biol. 2011, 14, 674–682. [Google Scholar] [CrossRef]
- Fan, L.; Li, R.; Pan, J.; Ding, Z.; Lin, J. Endocytosis and its regulation in plants. Trends Plant Sci. 2015, 20, 388–397. [Google Scholar] [CrossRef]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef] [Green Version]
- Khare, S.; Singh, N.B.; Singh, A.; Hussain, I.; Niharika, K.; Yadav, V.; Bano, C.; Yadav, R.K.; Amist, N. Plant secondary metabolites synthesis and their regulations under biotic and abiotic constraints. J. Plant Biol. 2020, 63, 203–216. [Google Scholar] [CrossRef]
- Yazaki, K. ABC transporters involved in the transport of plant secondary metabolites. FEBS Lett. 2006, 580, 1183–1191. [Google Scholar] [CrossRef] [Green Version]
- Lv, H.; Li, J.; Wu, Y.; Garyali, S.; Wang, Y. Transporter and its engineering for secondary metabolites. Appl. Microbiol. Biotechnol. 2016, 100, 6119–6130. [Google Scholar] [CrossRef]
- Larbat, R.; Robin, C.; Lillo, C.; Drengstig, T.; Ruoff, P. Modeling the diversion of primary carbon flux into secondary metabolism under variable nitrate and light/dark conditions. J. Theor. Biol. 2016, 402, 144–157. [Google Scholar] [CrossRef]
- Peng, M.; Hannam, C.; Gu, H.; Bi, Y.M.; Bothstein, S.J. A mutation in NLA, wich encodes a RING-type ubiquitin ligase, disrupts the adaptability of Arabidopsis to nitrogen limitation. Plant J. 2007, 50, 320–337. [Google Scholar] [CrossRef]
- Kant, S.; Peng, M.; Rothstein, S.J. Genetic regulation by NLA and microRNA827 for maintaining nitrate-dependent phosphate homeostasis in Arabidopsis. PLoS Genet. 2011, 7, e1002021. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.Y.; Huang, T.K.; Chiou, T.J. Nitrogen limitation adaptation, a target of microRNA827, mediates degradation of plasma membrane-localized phosphate transporters to maintain phosphate homeostasis in Arabidopsis. Plant Cell 2013, 25, 4061–4074. [Google Scholar] [CrossRef] [Green Version]
- Park, B.S.; Seo, J.S.; Chua, N.H. Nitrogen limitation adaptation recruits phosphate2 to target the phosphate transporter PT2 for degradation during the regulation of Arabidopsis phosphate homeostasis. Plant Cell 2014, 26, 454–464. [Google Scholar] [CrossRef] [Green Version]
- Yue, W.; Ying, Y.; Wang, C.; Zhao, Y.; Dong, C.; Whelan, J.; Shou, H. OsNLA1, a RING-type ubiquitin ligase, maintains phosphate homeostasis in Oryza sativa via degradation of phosphate transporters. Plant J. 2017, 90, 1040–1051. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.H.; Yan, W.; Yang, S.P.; Wang, A.; Gai, J.Y.; Zhu, Y.L. Overexpression of rice phosphate transporter gene OsPT2 enhances tolerance to low phosphorus stress in soybean. J. Agric. Sci. Technol. 2015, 17, 469–482. [Google Scholar]
- Zhu, W.; Yang, L.; Yang, S.; Gai, J.; Zhu, Y. Overexpression of rice phosphate transporter gene OsPT2 enhances nitrogen fixation and ammonium assimilation in transgenic soybean under phosphorus deficiency. J. Plant Biol. 2016, 59, 172–181. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Mean | Range | CV (%) |
|---|---|---|---|
| Relative value of leaf area | 0.63 | 0.38–0.95 | 18.9 |
| Relative value of shoot dry weight | 0.68 | 0.41–0.99 | 16.8 |
| Relative value of root dry weight | 1.08 | 0.66–1.61 | 15.6 |
| Relative value of plant dry weight | 0.73 | 0.44–1.03 | 15.4 |
| Cluster | Relative Value of Leaf Area | Relative Value of Shoot Dry Weight | Relative Value of Root Dry Weight | Relative Value of Plant Dry Weight | ||||
|---|---|---|---|---|---|---|---|---|
| Mean | Range | Mean | Range | Mean | Range | Mean | Range | |
| Group I | 0.86a | 0.75–0.95 | 0.90a | 0.81–0.99 | 1.46a | 1.31–1.61 | 0.97a | 0.89–1.03 |
| Group II | 0.44c | 0.38–0.48 | 0.47c | 0.41–0.52 | 0.85c | 0.66–1.03 | 0.52c | 0.44–0.57 |
| Group III | 0.63b | 0.41–0.91 | 0.68b | 0.52–0.94 | 1.08b | 0.67–1.44 | 0.73b | 0.57–0.99 |
| Sample | Total Clean Reads | Total Mapping Ratio | Uniquely Mapping Ratio | Total Gene Number |
|---|---|---|---|---|
| TCL1 | 66,496,250 | 92.38% | 70.18% | 29,167 |
| TCR2 | 66,186,556 | 92.03% | 72.58% | 29,817 |
| TTL3 | 65,608,344 | 92.54% | 69.65% | 28,970 |
| TTR4 | 65,388,728 | 92.26% | 73.41% | 29,981 |
| SCL5 | 65,498,816 | 91.75% | 69.00% | 29,330 |
| SCR6 | 65,052,664 | 91.53% | 72.37% | 29,814 |
| STL7 | 66,211,026 | 93.55% | 72.20% | 29,078 |
| STR8 | 66,062,774 | 93.09% | 75.68% | 30,335 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Li, Y.; Bai, W.; Yang, X.; Wu, H.; Lei, K.; Huang, R.; Zhang, S.; Huang, Q.; Lin, Q. Comparative Transcriptome Analysis Reveals Different Low-Nitrogen-Responsive Genes in Pepper Cultivars. Horticulturae 2021, 7, 110. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7050110
Wang C, Li Y, Bai W, Yang X, Wu H, Lei K, Huang R, Zhang S, Huang Q, Lin Q. Comparative Transcriptome Analysis Reveals Different Low-Nitrogen-Responsive Genes in Pepper Cultivars. Horticulturae. 2021; 7(5):110. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7050110
Chicago/Turabian StyleWang, Chunping, Yifei Li, Wenqin Bai, Xiaomiao Yang, Hong Wu, Kairong Lei, Renzhong Huang, Shicai Zhang, Qizhong Huang, and Qing Lin. 2021. "Comparative Transcriptome Analysis Reveals Different Low-Nitrogen-Responsive Genes in Pepper Cultivars" Horticulturae 7, no. 5: 110. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7050110