
CaHSP18.1a, a Small Heat Shock Protein from Pepper (Capsicum annuum L.), Positively Responds to Heat, Drought, and Salt Tolerance


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. RNA Extraction and Real-Time Fluorescent Quantitative PCR qRT-PCR Analysis
2.3. Subcellular Localization of CaHSP18.1a Protein
2.4. Virus-Induced Gene Silencing of CaHSP18.1a
2.5. Generation of CaHSP18.1a-Overexpression Arabidopsis Lines
2.6. Experimental Treatments and Sample Collection
2.7. Measurement of Physiological Indicators
2.8. Statistical Analyses
3. Results
3.1. Expression of the CaHSP18.1a in Pepper Plants under Abiotic Stress
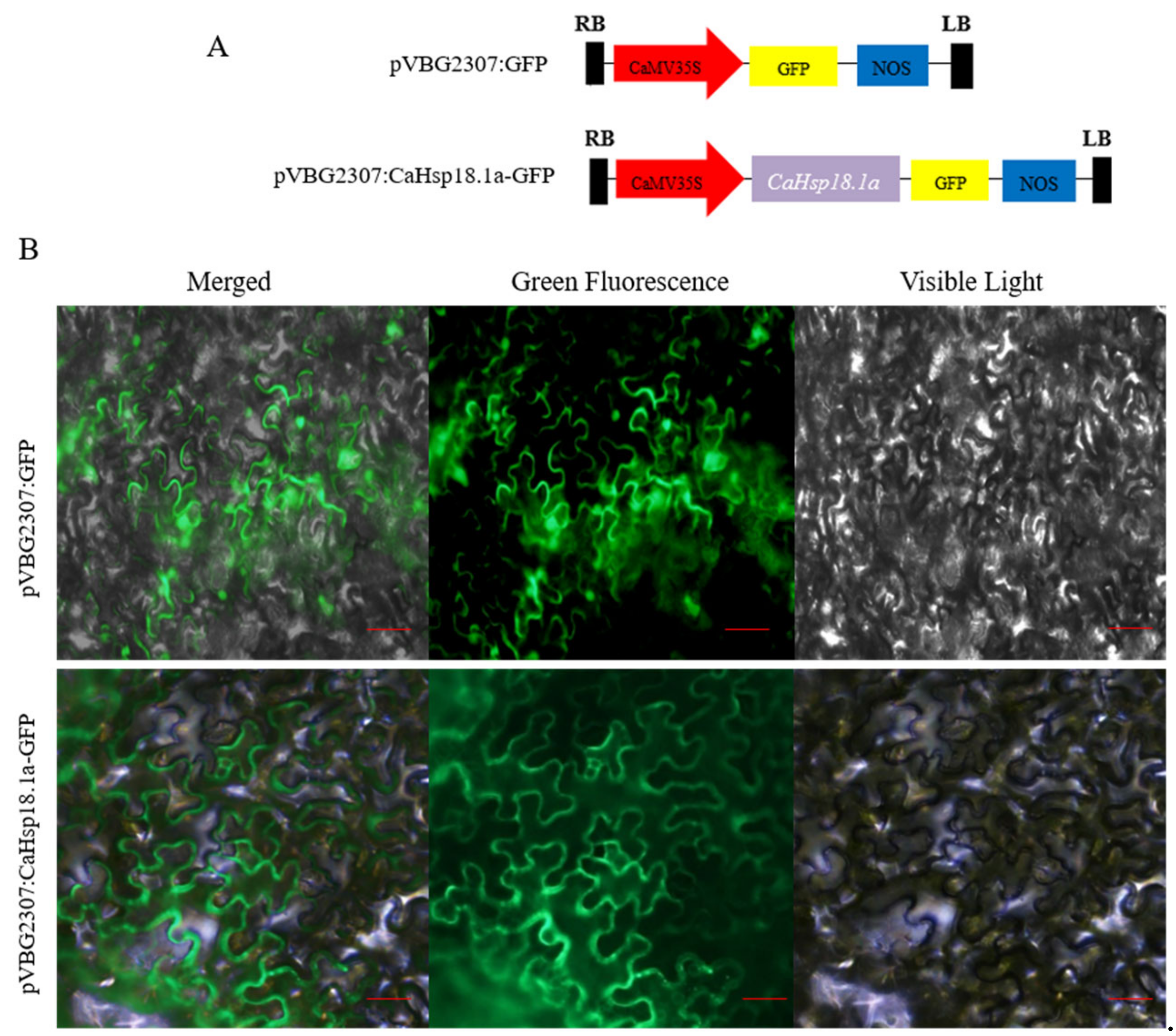
3.2. Subcellular Localization of CaHSP18.1a Protein
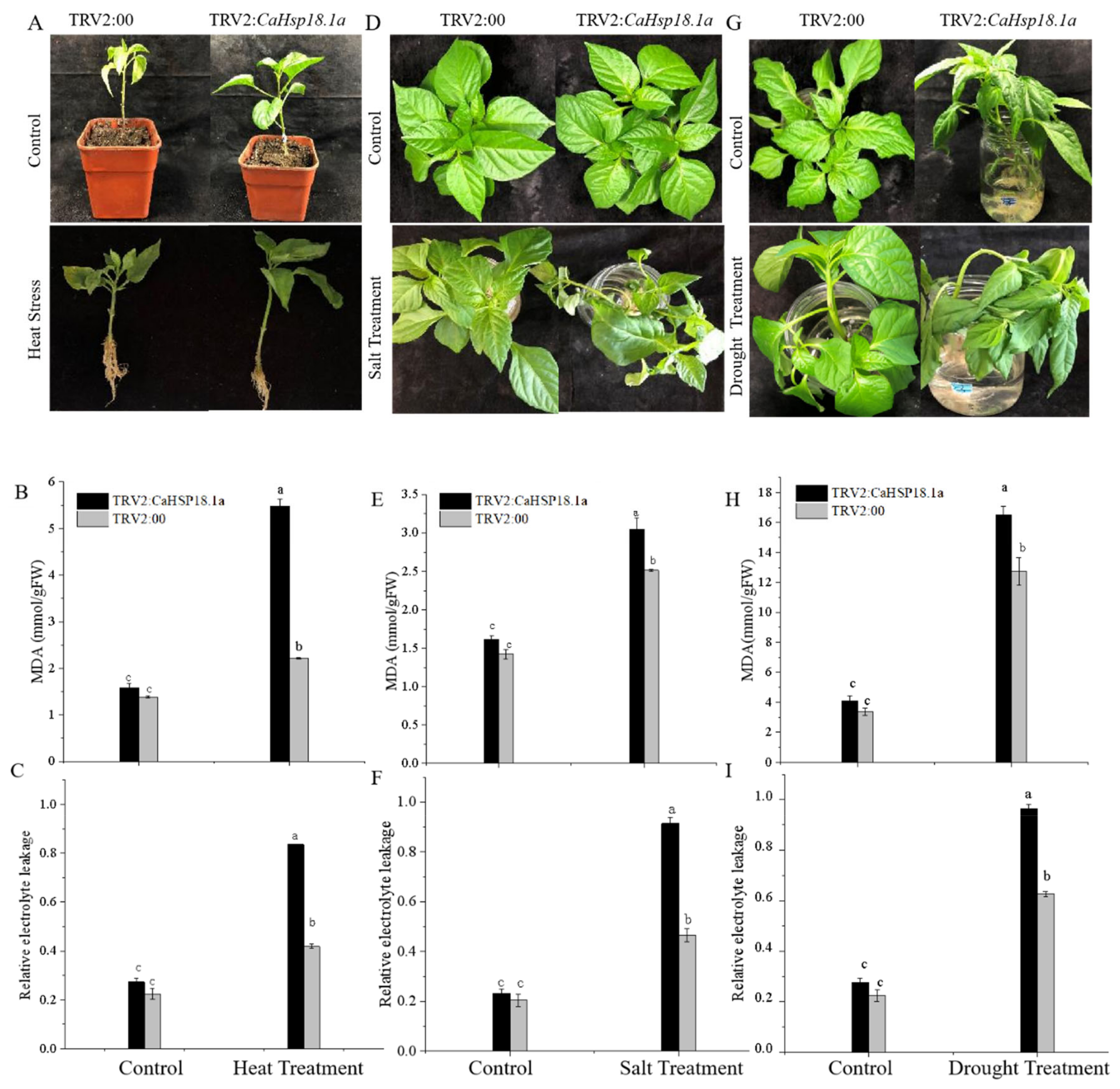
3.3. CaHSP18.1a-Silenced Plants Sensitive to Abiotic Stress
3.4. Effect of CaHSP18.1a Overexpression on Transgenic Arabidopsis
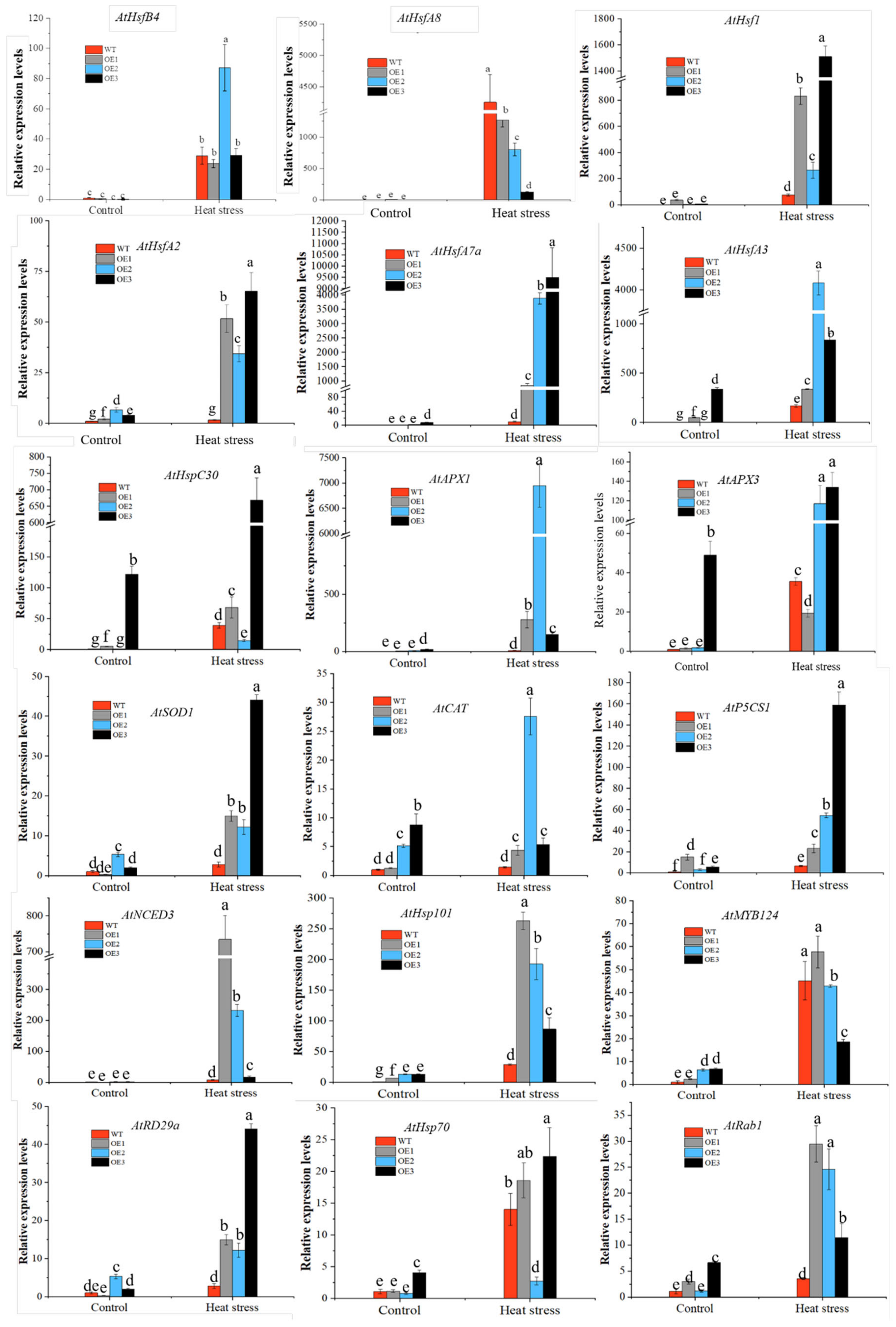
3.4.1. Overexpression of CaHSP18.1a Enhances Plant Tolerance of Heat Stress
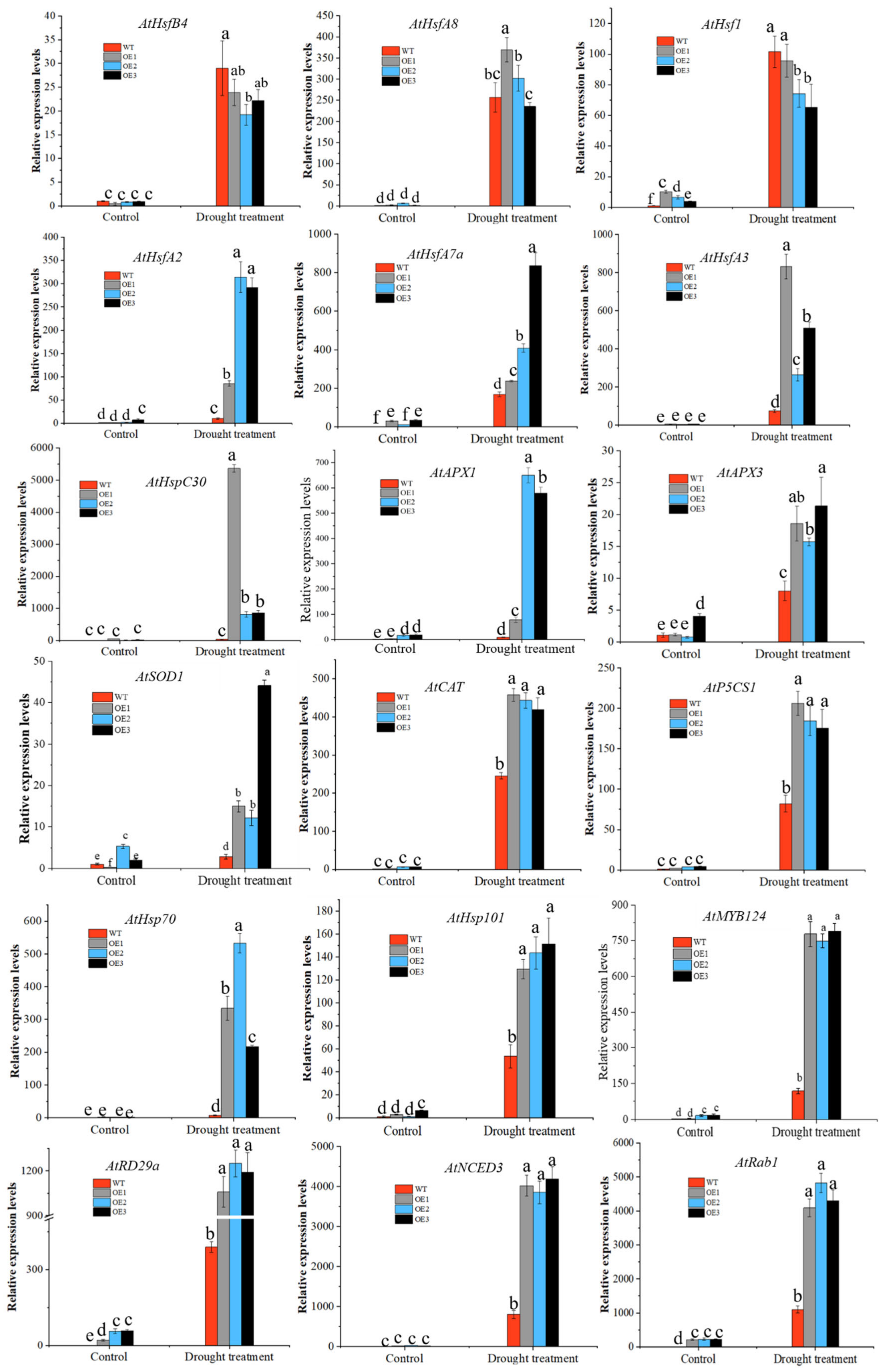
3.4.2. Overexpression of CaHSP18.1a Enhances Plant Tolerance to Drought Stress
3.4.3. Overexpression of CaHSP18.1a Enhances Plant Tolerance to Salt Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACD | alpha-crystallin domain |
| HS | heat stress |
| HSP | heat shock proteins |
| sHSPs | small heat shock proteins |
| REL | relative electrolyte leakage |
| MDA | Malondialdehyde |
| OE | Overexpression |
| OE1 | No. 1 Arabidopsis line with overexpressed CaHSP18.1a |
| OE2 | No. 2 Arabidopsis line with overexpressed CaHSP18.1a |
| OE3 | No. 3 Arabidopsis line with overexpressed CaHSP18.1a |
| R9 | a thermo-tolerant line |
| qRT-PCR | real-time fluorescence quantitative PCR |
| VIGS | virus-induced gene silencing |
| PDS | phytoene desaturase |
| TRV | tobacco rattle virus |
| ROS | reactive oxygen species |
| APX | ascorbate peroxidase |
| CAT | catalase |
| SOD | superoxide dismutase |
| POD | peroxidase |
References
- Ahuja, I.; de Vos, R.C.H.; Bones, A.M.; Hall, R.D. Plant molecular stress responses face climate change. Trends Plant Sci. 2010, 15, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, F.; Basha, E.; Fowler, M.E.; Kim, M.; Bordowitz, J.; Katiyar-Agarwal, S.; Vierling, E. Class I and II Small Heat Shock Proteins Together with HSP101 Protect Protein Translation Factors during Heat Stress. Plant Physiol. 2016, 172, 1221–1236. [Google Scholar] [PubMed] [Green Version]
- Muthusamy, S.K.; Dalal, M.; Chinnusamy, V.; Bansal, K.C. Genome-wide identification and analysis of biotic and abiotic stress regulation of small heat shock protein (HSP20) family genes in bread wheat. J. Plant Physiol. 2017, 211, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Chen, X.; Hui, W.; Xiang, Y.; Yan, M.; Wang, J. Overexpressing heat-shock protein OsHSP50.2 improves drought tolerance in rice. Plant Cell Rep. 2018, 37, 1585–1595. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, J. Research Progress of Heat Shock Protein. Guangdong Seric. 2006, 40, 39–42. [Google Scholar]
- Beck, E.H.; Fettig, S.; Knake, C.; Hartig, K.; Bhattarai, T. Specific and unspecific responses of plants to cold and drought stress. J. Biosci. 2007, 32, 501–510. [Google Scholar] [CrossRef]
- Agrawal, L.; Narula, K.; Basu, S.; Shekhar, S.; Ghosh, S.; Datta, A.; Chakraborty, S. Comparative Proteomics Reveals a Role for Seed Storage Protein AmA1 in Cellular Growth, Development, and Nutrient Accumulation. J. Proteome Res. 2013, 12, 4904–4930. [Google Scholar] [CrossRef]
- Shekhar, S.; Mishra, D.; Gayali, S.; Buragohain, A.K.; Chakraborty, S.; Chakraborty, N. Comparison of proteomic and metabolomic profiles of two contrasting ecotypes of sweetpotato (Ipomoea batata L.). J. Proteom. 2016, 143, 306–317. [Google Scholar] [CrossRef]
- Waters, E.R.; Lee, G.J.; Vierling, E. Evolution, structure and function of the small heat shock proteins in plants. J. Exp. Bot. 1996, 47, 325–338. [Google Scholar] [CrossRef]
- Boston, R.S.; Viitanen, P.V.; Vierling, E. Molecular chaperones and protein folding in plants. Plant Mol. Biol. 1996, 32, 191–222. [Google Scholar] [CrossRef]
- Hu, W.; Hu, G.; Han, B. Genome-wide survey and expression profiling of heat shock proteins and heat shock factors revealed overlapped and stress specific response under abiotic stresses in rice. Plant Sci. 2009, 176, 583–590. [Google Scholar] [CrossRef]
- Li, J.; Zhang, J.; Jia, H.; Li, Y.; Xu, X.; Wang, L.; Lu, M. The Populus trichocarpa PtHSP17.8 involved in heat and salt stress tolerances. Plant Cell Rep. 2016, 35, 1587–1599. [Google Scholar] [CrossRef]
- Eyles, S.J.; Gierasch, L.M. Nature’s molecular sponges: Small heat shock proteins grow into their chaperone roles. Proc. Natl. Acad. Sci. USA 2010, 107, 2727–2728. [Google Scholar] [CrossRef] [Green Version]
- Martin, H. A first line of stress defense: Small heat shock proteins and their function in protein homeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar]
- Tyedmers, J.; Mogk, A.; Bukau, B. Cellular strategies for controlling protein aggregation. Nat. Rev. Mol. Cell Biol. 2010, 11, 777–788. [Google Scholar] [CrossRef]
- Waters, E.R. The evolution, function, structure, and expression of the plant sHSPs. J. Exp. Bot. 2013, 64, 391–403. [Google Scholar] [CrossRef] [Green Version]
- Lambert, W.; Koeck, P.J.B.; Ahrman, E.; Purhonen, P.; Cheng, K.; Elmlund, D.; Hebert, H.; Emanuelsson, C. Subunit arrangement in the dodecameric chloroplast small heat shock protein HSP21. Protein Sci. 2011, 20, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Korotaeva, N.E.; Antipina, A.I.; Grabelnykh, O.I.; Varakina, N.N.; Borovskii, G.B.; Voinikov, V.K. Mitochondrial Low-Molecular-Weight Heat-Shock Proteins and the Tolerance of Cereal Mitochondria to Hyperthermia. Russ. J. Plant Physiol. 2001, 48, 798–803. [Google Scholar] [CrossRef]
- Nieto-Sotelo, J.; Martínez, L.M.; Ponce, G.; Cassab, G.I.; Alagón, A.; Meeley, R.B.; Ribaut, J.-M.; Yang, R. Maize HSP101 Plays Important Roles in Both Induced and Basal Thermotolerance and Primary Root Growth. Plant Cell Online 2002, 14, 1621–1633. [Google Scholar] [CrossRef]
- Sun, W.; Bernard, C.; Cotte, B.V.D.; Montagu, M.V.; Verbruggen, N. At-HSP17.6A, encoding a small heat-shock protein in Arabidopsis, can enhance osmotolerance upon overexpression. Plant J. 2010, 27, 407–415. [Google Scholar] [CrossRef]
- Sedaghatmehr, M.; MuellerRoeber, B.; Balazadeh, S. The plastid metalloprotease FtsH6 and small heat shock protein HSP21 jointly regulate thermomemory in Arabidopsis. Nat. Commun. 2016, 7, 12439. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Xu, Z.; Hwang, I. AtHSP17.8 overexpression in transgenic lettuce gives rise to dehydration and salt stress resistance phenotypes through modulation of ABA-mediated signaling. Plant Cell Rep. 2013, 32, 1953–1963. [Google Scholar] [CrossRef]
- Nicky, D.; Xu, J.; Peters, J.L.; Sunghun, P.; Ivo, R. Multi-Level Interactions Between Heat Shock Factors, Heat Shock Proteins, and the Redox System Regulate Acclimation to Heat. Front. Plant Sci. 2015, 6, 999. [Google Scholar]
- Guo, M.; Liu, J.H.; Ma, X.; Zhai, Y.F.; Gong, Z.H.; Lu, M.H. Genome-wide analysis of the HSP70 family genes in pepper (Capsicum annuum L.) and functional identification of CaHSP70-2 involvement in heat stress. Plant Sci. 2016, 252, 246–256. [Google Scholar] [CrossRef]
- Zou, X. Cultivation Seasons and Cropping patterns of Capsicum in China. J. China Capsicum 2002, 3, 1672–4542. [Google Scholar]
- Guo, M.; Zhai, Y.-F.; Lu, J.-P.; Chai, L.; Chai, W.-G.; Gong, Z.-H.; Lu, M.-H. Characterization of CaHSP70-1, a Pepper Heat-Shock Protein Gene in Response to Heat Stress and Some Regulation Exogenous Substances in Capsicum annuum L. Int. J. Mol. Sci. 2014, 15, 19741–19759. [Google Scholar] [CrossRef] [Green Version]
- Pagamas, P.; Nawata, E. Sensitive stages of fruit and seed development of chili pepper (Capsicum annuum L. var. Shishito) exposed to high-temperature stress. Sci. Hortic. 2008, 117, 21–25. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.H.; Lu, J.P.; Zhai, Y.F.; Wang, H.; Gong, Z.H.; Wang, S.B.; Lu, M.H. Genome-wide analysis of the CaHSP20 gene family in pepper: Comprehensive sequence and expression profile analysis under heat stress. Front. Plant Sci. 2015, 6, 806. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Cheng, G.; Khan, A.; Wei, A.; Gong, Z. CaHSP16.4, a small heat shock protein gene in pepper, is involved in heat and drought tolerance. Protoplasma 2019, 256, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhang, H.; Ali, M.; Gai, W.; Gong, Z. A small heat shock protein CaHSP25.9 positively regulates heat, salt, and drought stress tolerance in pepper (Capsicum annuum L.). Plant Physiol. Biochem. 2019, 142, 151–162. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Gao, N.; Shan, Z.; Guo, S. Pepper Chloroplast Small Heat Shock Proteins CaHSP26 Enhanced the Ability of Anti-high Temperature Stress in Arabidopsis. North. Hortic. 2011, 6, 140–143. [Google Scholar]
- Sun, J.; Cheng, G.; Huang, L.; Liu, S.; Ali, M.; Khan, A.; Yu, Q.; Yang, S.; Luo, D.; Gong, Z. Modified expression of a heat shock protein gene, CaHSP22.0, results in high sensitivity to heat and salt stress in pepper (Capsicum annuum L.). Sci. Hortic. 2019, 249, 364–373. [Google Scholar] [CrossRef]
- Yu, J.; Cheng, Y.; Feng, K.; Ruan, M.; Ye, Q.; Wang, R.; Li, Z.; Zhou, G.; Yao, Z.; Yang, Y.; et al. Genome-Wide Identification and Expression Profiling of Tomato HSP20 Gene Family in Response to Biotic and Abiotic Stresses. Front. Plant Sci. 2016, 7, 1215. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zou, Z.; Li, Q.; Sun, K.; Chen, X.; Li, X. The CsHSP17.2 molecular chaperone is essential for thermotolerance in Camellia sinensis. Sci. Rep. 2017, 7, 1237. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Y.; Wang, H.; Liang, M.; Lu, M. Both silencing- and over-expression of pepper CaATG8c gene compromise plant tolerance to heat and salt stress. Environ. Exp. Bot. 2017, 141, 10–18. [Google Scholar] [CrossRef]
- Wang, J.E.; Liu, K.K.; Li, D.W.; Zhang, Y.L.; Zhao, Q.; He, Y.M.; Gong, Z.H. A novel peroxidase CaPOD gene of pepper is involved in defense responses to Phytophthora capsicum infection as well as abiotic stress toleranc. Int. J. Mol. Sci. 2013, 14, 3158–3177. [Google Scholar] [CrossRef]
- Wan, H.; Yuan, W.; Ruan, M.; Ye, Q.; Wang, R.; Li, Z.; Zhou, G.; Yao, Z.; Zhao, J.; Liu, S.; et al. Identification of reference genes for reverse transcription quantitative real-time PCR normalization in pepper (Capsicum annuum L.). Biochem. Biophys. Res. Commun. 2011, 416, 24–30. [Google Scholar] [CrossRef]
- Livaka, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. J. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Yu, C.; Zhan, Y.; Feng, X.; Huang, Z.A.; Sun, C. Identification and Expression Profiling of the Auxin Response Factors in Capsicum annuum L. under Abiotic Stress and Hormone Treatments. Int. J. Mol. Sci. 2017, 18, 2719. [Google Scholar] [CrossRef] [Green Version]
- Clough, S.J.; Bent, A. Floral dip: A simplified method for Agrobacterium -mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.X.; Guo, W.L.; Zhang, Y.L.; Ji, J.J.; Xiao, H.J.; Yan, F.; Zhao, Y.Y.; Zhu, W.C.; Chen, R.G.; Chai, W.G. Cloning and characterisation of a pepper aquaporin, CaAQP, which reduces chilling stress in transgenic tobacco plants. Plant Cell Tissue Organ Cult. 2014, 118, 431–444. [Google Scholar] [CrossRef]
- Arkus, K.A.J.; Cahoon, E.B.; Jez, J.M. Mechanistic analysis of wheat chlorophyllase. Arch. Biochem. Biophys. 2005, 438, 146–155. [Google Scholar] [CrossRef]
- Campos, P.S.; nia Quartin, V.; chicho Ramalho, J.; Nunes, M.A. Electrolyte leakage and lipid degradation account for cold sensitivity in leaves ofCoffea sp. plants. J. Plant Physiol. 2003, 160, 283–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.L.; Chen, R.G.; Gong, Z.H.; Yin, Y.X.; Ahmed, S.S.; He, Y.M. Exogenous abscisic acid increases antioxidant enzymes and related gene expression in pepper (Capsicum annuum) leaves subjected to chilling stress. Genet. Mol. Res. GMR 2012, 11, 4063–4080. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Aebi, H. [13] Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Jiang, C.; Xu, J.; Zhang, H.A.; Zhang, X.; Shi, J.; Li, M.I.; Ming, F. A cytosolic class I small heat shock protein, RcHSP17.8, of Rosa chinensis confers resistance to a variety of stresses to Escherichia coli, yeast and Arabidopsis thaliana. Plant Cell Environ. 2009, 32, 1046–1059. [Google Scholar] [CrossRef]
- Lopes-Caitar, V.S.; de Carvalho, M.C.; Darben, L.M.; Kuwahara, M.K.; Nepomuceno, A.L.; Dias, W.P.; Abdelnoor, R.V.; Marcelino-Guimarães, F.C. Genome-wide analysis of the HSP20 gene family in soybean: Comprehensive sequence, genomic organization and expression profile analysis under abiotic and biotic stresses. BMC Genom. 2013, 14, 1471–2164. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Huang, W.; Yang, Z.; Liu, J.; Huang, B. Transcriptional regulation of heat shock proteins and ascorbate peroxidase by CtHsfA2b from African bermudagrass conferring heat tolerance in Arabidopsis. Sci. Rep. 2016, 6, 28021. [Google Scholar] [CrossRef]
- Schramm, F.; Ganguli, A.; Kiehlmann, E.; Englich, G.; Walch, D.; von Koskull-Döring, P. The Heat Stress Transcription Factor HsfA2 Serves as a Regulatory Amplifier of a Subset of Genes in the Heat Stress Response in Arabidopsis. Plant Mol. Biol. 2006, 60, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gao, X.; Dong, J.; Tian, X.; Wang, J.; Palta, J.A.; Xu, S.; Fang, Y.; Wang, Z. Over-Expression of the Heat-Responsive Wheat Gene TaHSP23.9 in Transgenic Arabidopsis Conferred Tolerance to Heat and Salt Stress. Front Plant Sci. 2020, 11, 243. [Google Scholar] [CrossRef]
- He, Y.; Yao, Y.; Li, L.; Li, Y.; Gao, J.; Fan, M. A heat-shock 20 protein isolated from watermelon (ClHSP22.8) negatively regulates the response of Arabidopsis to salt stress via multiple signaling pathways. PeerJ. 2021, 9, e10524. [Google Scholar] [CrossRef] [PubMed]
- Yao, B.; Yue, S.; Sun, L.; Ma, Z.; Su, J.; Liu, X. Development of Virus Induced Gene Silencing and Application of VIGS in Plant Abiotic Stresses. Chin. Agric. Sci. Bull. 2016, 32, 131–136. [Google Scholar]
- Ammar, J.B.; Lanoiselle, J.-L.; Lebovka, N.I.; Hecke, E.V.; Vorobiev, E.J.J. Impact of a Pulsed Electric Field on Damage of Plant Tissues: Effects of Cell Size and Tissue Electrical Conductivity. J. Food Sci. 2011, 76, E90–E97. [Google Scholar] [CrossRef]
- Ma, B.; Lu, M.; Gong, Z.H. Responses of growth and physiology of pepper (Capsicum annuum L.) seedlings to high temperature stress. J. Northwest A F Univ. 2013, 41, 112–118. [Google Scholar]
- Shang, H.; Cao, S.; Yang, Z.; Cai, Y.; Zheng, Y. Effect of Exogenous γ-Aminobutyric Acid Treatment on Proline Accumulation and Chilling Injury in Peach Fruit after Long-Term Cold Storage. J. Agric. Food Chem. 2011, 59, 1264–1268. [Google Scholar] [CrossRef]
- Rossel, J.B.; Wilson, I.W.; Pogson, B.J. Global Changes in Gene Expression in Response to High Light in Arabidopsis. Plant Physiol. 2002, 130, 1109–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghdam, M.S.; Sevillano, L.; Flores, F.B.; Bodbodak, S. Heat shock proteins as biochemical markers for postharvest chilling stress in fruits and vegetables. Sci. Hortic. 2013, 160, 54–64. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Li, J.; Hao, R.; Guo, Y. Activation of catalase activity by a peroxisome-localized small heat shock protein HSP17.6CII. J. Genet. Genom. 2017, 44, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Liu, Y.; Kong, X.; Zhang, D.; Pan, J.; Zhou, Y.; Wang, L.; Li, D.; Yang, X. ZmHSP16.9, a cytosolic class I small heat shock protein in maize (Zea mays), confers heat tolerance in transgenic tobacco. Plant Cell Rep. 2012, 31, 1473–1484. [Google Scholar] [CrossRef] [PubMed]
- Meifang, L.; Xiujian, W.; Shangjing, G. Obtain of Transgenic Sweet Pepper Plants with Gene from Endoplasmic Reticulum Small Heat Shock Protein CaHSP22.5. Mol. Plant Breed. 2018, 7, 2212–2218. [Google Scholar]
- Zhao, J.; Liang, J.; Wu, X.; Liu, X.; Li, H.; Zhu, S. Expression Profiling of Rice ROS Scavenging System Related Genes under Salt or Low Temperature Stress. Acta Bot. Boreali-Occident. Sin. 2015, 35, 872–883. [Google Scholar]
- Kim, J.M.; Woo, D.H.; Kim, S.H.; Lee, S.Y.; Park, H.Y.; Seok, H.Y.; Chung, W.S.; Moon, Y.H. Arabidopsis MKKK20 is involved in osmotic stress response via regulation of MPK6 activity. Plant Cell Rep. 2012, 31, 217–224. [Google Scholar] [CrossRef]
- Tuteja, N.; Banu, M.S.A.; Huda, K.M.K.; Gill, S.S.; Jain, P.; Xuan, H.P.; Tuteja, R. Pea p68, a DEAD-Box Helicase, Provides Salinity Stress Tolerance in Transgenic Tobacco by Reducing Oxidative Stress and Improving Photosynthesis Machinery. PLoS ONE 2014, 9, e98287. [Google Scholar] [CrossRef] [Green Version]
- Ron, M.; Sandy, V.; Martin, G.; Frank, V.B. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar]
- Cao, S.; Yang, Z.; Cai, Y.; Zheng, Y. Fatty acid composition and antioxidant system in relation to susceptibility of loquat fruit to chilling injury. Food Chem. 2011, 127, 1777–1783. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell Online 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Moller, I.M. Plant Mitochondria and Oxidative Stress: Electron Transport, Nadph Turnover, and Metabolism of Reactive Oxygen Species. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 561–591. [Google Scholar] [CrossRef] [Green Version]
- Charng, Y.; Liu, H.; Liu, N.; Chi, W.; Wang, C.; Chang, S.; Wang, T. A Heat-Inducible Transcription Factor, HsfA2, Is Required for Extension of Acquired Thermotolerance in Arabidopsis. Plant Physiol. 2007, 143, 251–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heerklotz, D.; Döring, P.; Bonzelius, F.; Winkelhaus, S.; Nover, L. The balance of nuclear import and export determines the intracellular distribution and function of tomato heat stress transcription factor HsfA2. Mol. Cell. Biol. 2001, 21, 1759–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, D.; Kazuo, Y.; Nishiuchi, T. High-level overexpression of the Arabidopsis HsfA2 gene confers not only increased themotolerance but also salt/osmotic stress tolerance and enhanced callus growth. J. Exp. Bot. 2007, 58, 3373–3383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pv, K.D.; Scharf, K.D.; Nover, L. The diversity of plant heat stress transcription factors. Trends Plant Sci. 2007, 12, 452–457. [Google Scholar]
- Burke, J.J.; Chen, J. Enhancement of reproductive heat tolerance in plants. PLoS ONE 2015, 10, e0122933. [Google Scholar] [CrossRef] [Green Version]
- Charng, Y.Y.; Liu, H.C.; Liu, N.Y.; Hsu, F.C.; Ko, S.S. Arabidopsis Hsa32, a novel heat shock protein, is essential for acquired thermotolerance during long recovery after acclimation. Plant Physiol. 2006, 4, 1297–1305. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, N.H.; Nguyen, C.T.T.; Jung, C.; Cheong, J.-J. AtMYB44 suppresses transcription of the late embryogenesis abundant protein gene AtLEA4-5. Biochem. Biophys. Res. Commun. 2019, 511, 931–934. [Google Scholar] [CrossRef]
- Nakashima, K.; Shinwari, Z.K.; Sakuma, Y.; Seki, M.; Miura, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Organization and expression of two Arabidopsis DREB2 genes encoding DRE-binding proteins involved in dehydration- and high-salinity-responsive gene expression. Plant. Mol. Biol. 2000, 42, 657–665. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C. Proline accumulation in plants: A review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Rymaszewski, W.; Vile, D.; Bediee, A.; Dauzat, M.; Luchaire, N.; Kamrowska, D.; Granier, C.; Hennig, J. Stress-Related Gene Expression Reflects Morphophysiological Responses to Water Deficit. Plant. Physiol. 2017, 174, 1913–1930. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Huang, L.; Zhang, X.; Wang, M.; Xu, G.; Xia, X. Expression of rice geneOsMSR4confers decreased ABA sensitivity and improved drought tolerance inArabidopsis thaliana. Plant. Growth Regul. 2015, 75, 549–556. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.-L.; Liu, S.; Xiao, J.-J.; Cheng, G.-X.; ul, H.S.; Gong, Z.-H. CaHSP18.1a, a Small Heat Shock Protein from Pepper (Capsicum annuum L.), Positively Responds to Heat, Drought, and Salt Tolerance. Horticulturae 2021, 7, 117. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7050117
Liu Y-L, Liu S, Xiao J-J, Cheng G-X, ul HS, Gong Z-H. CaHSP18.1a, a Small Heat Shock Protein from Pepper (Capsicum annuum L.), Positively Responds to Heat, Drought, and Salt Tolerance. Horticulturae. 2021; 7(5):117. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7050117
Chicago/Turabian StyleLiu, Yan-Li, Shuai Liu, Jing-Jing Xiao, Guo-Xin Cheng, Haq Saeed ul, and Zhen-Hui Gong. 2021. "CaHSP18.1a, a Small Heat Shock Protein from Pepper (Capsicum annuum L.), Positively Responds to Heat, Drought, and Salt Tolerance" Horticulturae 7, no. 5: 117. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7050117