
The Role of Canary Island Date Palms in Physical Amenity Provisioning for Urban Landscape Settings


Institute for Land, Water and Society, Charles Sturt University, P.O. Box 789, Albury, NSW 2640, Australia
Horticulturae 2021, 7(7), 201; https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7070201
Submission received: 3 June 2021
/
Revised: 15 July 2021
/
Accepted: 15 July 2021
/
Published: 19 July 2021
(This article belongs to the Special Issue Urban Horticulture and Nature-Based Solutions for Better Cities )
Abstract
:Canary Island date palms (Phoenix canariensis) have been planted as a landscaping feature plant throughout warm, temperate, and subtropical climates. The physical amenity provisioning of this species (shade effects, microclimate amelioration, water usage, etc.) has so far not been systematically assessed. This paper reports on temperature and humidity measurements in both a suburban and a rural location in SE Australia. The study demonstrates the effects of the palm canopy as regulator of humidity and provider of shade and, thus, amenity values in urban landscape settings. Drawing on published energy savings and growth requirements of the plant, the paper argues that Canary Island date palms are landscaping plants suitable to ameliorate the microclimate in urban neighborhoods with varied socio-economic conditions.
1. Introduction
Urban design has recently become concerned with thermal comfort, an issue that is gaining significance in the face of actual and projected climate change [1,2,3,4]. Thermal stress (‘heat stress’) has been implicated in increased mortality among infants as well as the elderly [5,6,7,8,9], with lower socio-economic strata disproportionality affected [10,11].
Trees have been advocated as a means to ameliorate thermal stress, both through the provision of shade and through cooling provided by evapotranspiration [12,13,14,15]. With exceptions [16], palms have been shown to contribute to urban cooling [17,18,19,20] due to leaf albedo [21], shade foliage, evapotranspiration, low heat capacity of vegetation, and high aerodynamic roughness [22]. However, there have been few systematic studies examining individual palm species [23,24].
1.1. Palms in Landscaping
In the past, motivations to plant palms over other species included the desire to invoke the sense of the exotic [25,26,27] and, on a public level, to demonstrate colonial connections and aspirations [28,29]. Today, many of these early plantings form an integral part of several communities’ heritages [30,31,32]. In the 1990s and early 2000s, palms saw a resurgence in popularity, especially in private gardens, primarily driven by the fact that mature plants can be readily transported and planted [33,34] and, therefore, lend themselves to instant landscaping or gardens [35,36,37] as well as streetscapes and promenades [38,39,40]. The large-scale use of palms in urban areas has come under criticism from ecologists for their perceived low contribution to biodiversity [41]; from town planners and landscape architects [42]; and from researchers modelling the carbon sink potential of urban forests [43,44,45].
This criticism is, by and large, undifferentiated as it treats ‘palms’ as a collective uniform class of plants. Dissenting voices, providing a more nuanced approach, are few [23]. In reality, however, the palm species planted as ornamentals exhibit a wide range of morphological characteristics such as crown patterns circumscribed by leaf shapes (pinnate vs. palmate), leaf numbers (and thus crown density) and leaf dimensions (and thus crown diameters), stem heights, as well as the quantity, dimensions, flesh content and ripening duration of their fruits (drupes).
These morphological characteristics have direct implications on the suitability of a palm species as a provider of shade for people (the topic of this paper); as a habitat for native and exotic birds [46]; and as a provider of food for native and exotic species [47]. As Canary Island date palms constitute the majority of formal plantings in private gardens, streets, and in public parks in most parts of the warm temperate climates, this species will be the focus of this paper.
1.2. Canary Islands Date Palms
Canary Island date palms (Phoenix canariensis Wildpret ex Chabaud) are endemic to the Canary Islands off of the west coast of Africa [48]. In its natural setting, P. canariensis will grow to about 18–20 m in height, with a stem usually of 0.6 to 0.8 m, but can grow up to 1.2 m diameter. The crown has a diameter of 6–8 m (max 10 m), made up of in excess of 200 arching, curved pinnate fronds of 4–6 m (max 7 m) in length. Unless affected by disease or pests, the plant can live for 200–300 years. Fully mature palms can weigh up to 10 tons in mass [49].
Introduced into the European nursery trade in the 1860s [50], they found ready acceptance as feature trees in private and urban parks and were employed for effect as linear plantings in streets and on promenades (e.g., at Nice or Cannes) during the late nineteenth and early twentieth century [51,52], so much so that they are now distributed throughout many warm temperate climates [53].
One hundred and fifty years after its introduction by the horticultural industry, the Canary Island date palm has become one of the most ubiquitous ornamental palm species. They serve as single or grouped feature trees in public and private gardens, not only in the urban and sub-urban built environment, but also in landscaped homestead blocks in agricultural settings. Moreover, in many communities, Canary Island date palms line drive-ways, public streets, and sea-side promenades.
1.3. Shade Effects of Canary Islands Date Palms
Some studies have classified the leaf index of palms as being equal to a shrub despite palms being larger structures [54]. This, however, was based on Syagrus romanzoffiana, a palm with a much more open canopy than P. canariensis. The only study that examined the shade and evapotranspiration effects of Canary Island date palms was carried out in southern Greece during a single month (June). Georgi and Dimitriou found that the air temperature underneath Canary Island date palms was on average 1.95 ± 0.87 °C cooler than a shaded pavement, and 2.92 ± 0.81 °C cooler than a sunlit pavement (all measured at 1.5 m above ground) (both differences significant [t-test] at p > 0.001) [55]. In their study, Canary Island date palms had better shade values than bitter orange (Citrus aurantium) and olive trees (Olea europea) (the latter significantly better; p = 0.049), but lesser shade values than stone pines (Pinus brutia) and Indian laurel figs (Ficus retusa ssp nitida) (the latter being significantly worse; p = 0.002). While the humidity was significantly higher underneath the Canary Island date palm compared to the pavement readings (shaded: 5.63 ± 3.83% RH; p = 0.010; sunlit: 5.30 ± 3.44% RH; p = 0.018), there was no significant difference in the humidity readings under any of the plant canopies. Georgi’s and Dimitriou’s findings have limitations, however, as they are limited to single measurements close to mid-day [55].
Shashua-Bar et al. compared the shade effects of the commercial date palm (Phoenix dactylifera) with those of Indian laurel figs and Rosewood (Tipuana tipu) [56]. The Date Palm tree characteristics are a sparse and narrow canopy, with a considerable percentage of sunny spots (30–40%) and high solar transmissivity (an average of 15%). Assessing the temperatures in Tel Aviv (Israel) at four times of the day (at 6:00, 12:00, 15:00 and 24:00), Shashua-Bar et al., found a net cooling efficient of 0.4 which increased to 0.7 when planted over grass [56]. Date palms had better shade effects on temperature during the day (12:00 and 15:00) and better heat retention during the night (24:00 and 06:00) than the ambient environment. Their shade effects were only marginally less effective than those of Indian laurel figs but exhibited a higher heat retention during the night. Rosewood was found to be less effective overall.
Given the paucity of studies specifically dealing with Canary Island date palms, there was a need to empirically examine the diurnal microclimatic variations of temperature and humidity under the canopy of Canary Island date palms in an urban and a rural setting in order to provide data that (a) examine the suitability of Canary Island date palms as shade trees in urban settings and (b) illuminate the environmental conditions the Canary Island date palm provides as habitat for animal species in both rural and urban settings.
The experimental study was to test whether there were significant differences in the microclimate between the crown canopy and trunk of a Canary Island date palm compared with a control and a tree with medium-sided leaves. It was further posited that there would be differences in the microclimate amelioration provided by palms in urban settings compared to exposed settings in farmland.
This paper will report on the findings of the microclimate study and will then discuss the contribution made by Canary Islands date palms as regulators of humidity and providers of shade and thus amenity values in urban landscape settings. Drawing on the energy savings and growth requirements of the plant, the paper will conclude with some commentary on the suitability of the plant for various urban neighborhoods.
2. Materials and Methods
2.1. Locations
Temperature sensors (see below) were inserted in a Canary Island date palm as well as two controls at two locations: a 50-year old, 9.2 m tall Canary Island date palm (trunk height 4.4 m) in an urban setting in Albury (Figure S1; co-ordinates −36.074156; 146.907122) and a 70-year old, 7.8 m tall Canary Island date palm (trunk height 2.2 m) in an open, rural setting at Alma Park (Figure S2 co-ordinates −35.583805, 146.784574) [57]. The specific locations were chosen as P. canariensis drupes and seeds collected from these locations had been used for a range of research [58,59,60]. The Canary Island date palm in the urban setting is surrounded by low density housing in an established residential neighborhood with a low density of street and feature trees and was thus representative of common ornamental plantings. The Canary Island date palm in the rural setting has been planted as part of a single row lining a driveway in a rural production landscape with isolated paddock trees (spaced 150 to 300 m apart). This location can be considered to be representative of an unprotected palm, unencumbered by urban or suburban development.
2.2. Experimental Set Up
Temperatures and relative humidity data were logged with a TinyTag Plus 2 TGP-4500 temperature and humidity logger mounted on a 100 × 50 × 25 mm section of dressed pine which had been mounted to the measuring location with fencing wire. Each set up comprised of one sensor fastened to the stem just below the crown, at absolute heights of 2 m (Alma Park) to 3.5 m (Albury) above ground. One sensor was suspended from a petiole in the crown; one sensor was fastened to a fence post (to collect ambient temperature); and one sensor was fastened to a branch in a close-by eucalypt tree (Figure S3). In the Albury case the locations for placing the control sensor were limited (to avoid vandalism or theft). In consequence, during the course of the day, the chosen location was unfortunately temporarily shaded by an adjacent building (see below).
In the Albury case it was not possible to use a eucalypt as all were located at an elevation that differed too much from that of the source palm. The closest eucalypt (a Eucalyptus melliodora) was located 95 m to the northeast, with its base 8 m above that of the palm, and its lowest leaf-bearing branch 8 m above ground. The total height difference of 10 m equates to 1–2 °C in temperature difference. Thus a silver birch (Betula pendula) was chosen, which grew 30 m to the southwest and on the same elevation as the Canary Island date palm (Figure S1 n° 4; Figure S3c). In the Alma Park scenario, a suitable eucalypt (E. melliodora) was located 150 m to the northwest, with an elevation difference of less than 1 m (Figure S2 n° 8; Figure S3d). In both cases the sensor was installed on the eastern side of the tree.
2.3. Duration
The experiment was conducted from 25 November 2018 to 11 April 2019. The readings were obtained at 15 min intervals until 6 February 2019 (8245 readings) and at 6 min intervals thereafter (15,396 readings). Two sets of four data series were obtained.
2.4. Data Cleaning and Analysis
The data series for each location were synchronized during data cleaning (deviation ± 4 min at 15 min interval readings, ±1 min at 6 min interval). The humidity recording of one device at Alma Park provided inverted humidity percentages (100-% RH instead of % RH), which was easily corrected during data cleaning.
The humidity data recordings were, on occasion, erratic, which may be attributed to sensors that were unshielded and may have been exposed to rain. This was resolved during the data cleaning process, when all four data sets (per location) were compared and excessive outliers identified. In these cases, the affected readings/times were deleted in all four data sets.
The vapor pressure deficit (ed) was calculated as the difference between actual vapor pressure (ea) and saturation vapor pressure (es):
where T is temperature (°C) and RH is relative humidity (%) [61].
ed = ea − es
es = 0.6108 * exp(17.27 * T/(T + 237.3))
ea = RH/100 * es
2.5. Shade Modelling
2.6. Tree-Benefits Analysis
The economic benefits of Canary Islands date palms in an Australian setting were calculated using the U.S. National Tree Benefit Calculator (NTBC) [66] with the comparison of Australian climate zones [67] with US climate zones based on Horne, et al. [68]. To avoid fluctuating exchange rates from influencing the discussion of absolute monetary cost comparisons, all values were calculated relative to the most common street tree, the London plane tree (Platanus × acerifolia). The costs are based on the palms’ contribution to storm water, electricity, gas, air quality and CO2.
In the NTBC, electricity value is measured by energy conserved by cooling caused by the presence of the tree due to shading, evapotranspiration, and wind interception. Natural gas is based on the same calculations but refers to savings in oil or natural gas consumption normally required for heating. Air quality quantifies the amount of pollutants that are avoided, absorbed, or intercepted by the tree. These benefits can be broken down by ozone, nitrogen dioxide, sulfur dioxide, PM10, and other volatile organic compounds. CO2 refers to the reduction of atmospheric carbon dioxide by carbon sequestration (the amount of CO2 processed by the plant) and the amount of CO2 emissions avoided by the reduced heating and air conditioning demands.
The NTBC also includes an estimation of the contribution to property values, which have not been included here since this valuation is far too subjective. The NTBC shows that palms make a very low contribution to property values, which, at least in the Australian setting, is contradicted by the fact that mature palms have been transplanted in large numbers as instant landscaping in metropolitan cities to value add to new estates.
3. Results
3.1. Temperature
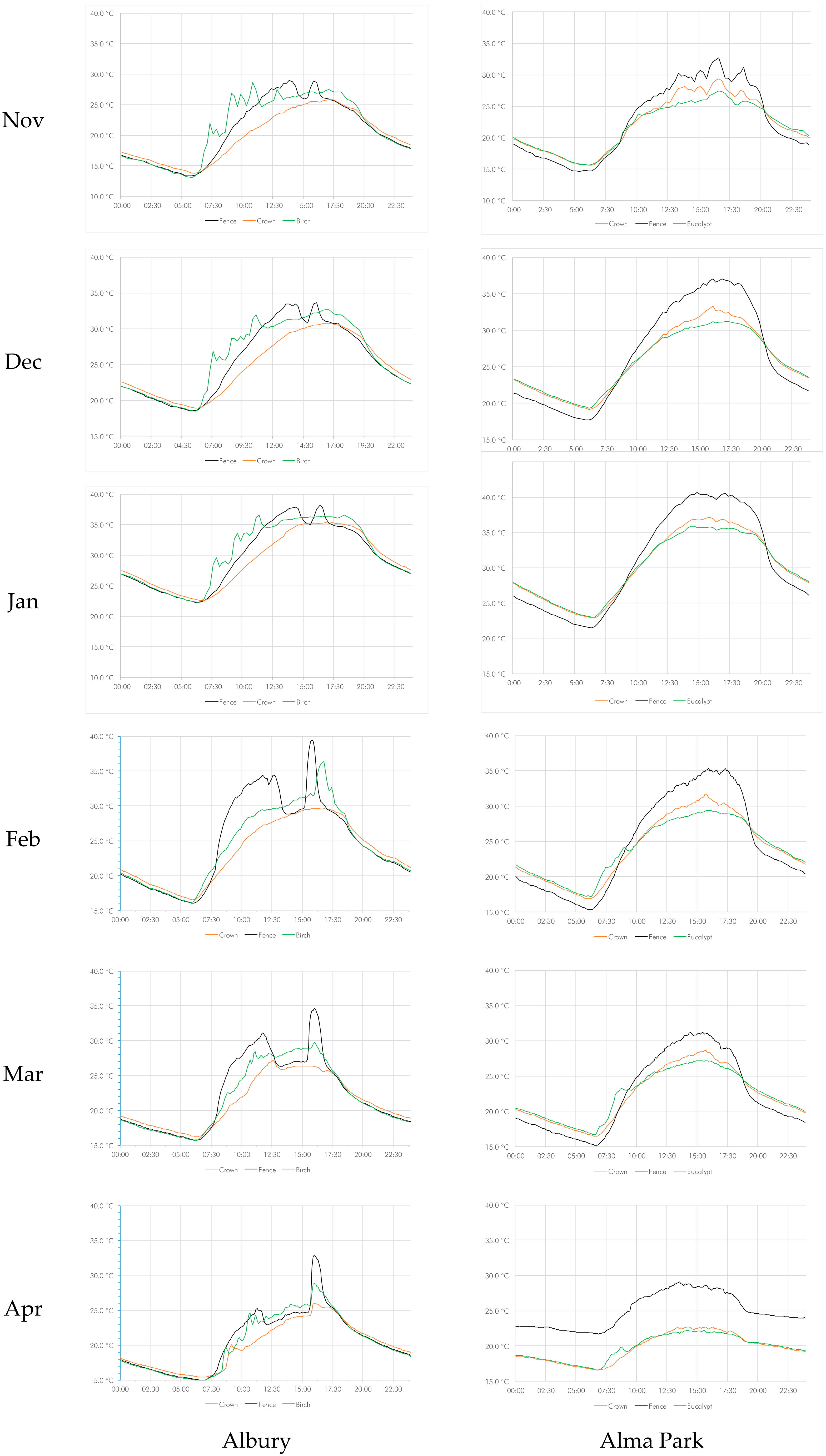
The average hourly values for temperature have been graphed at monthly intervals in Figure 1. The data show that temperatures in the crown of the palm are significantly cooler during the day than the ambient temperature and warmer during the night. This is consistent both in the urban and the rural setting (Tables S1 and S2).
The temperature graphs for the Albury control sample (a fence post) exhibit a distinct and well-defined drop in temperature during the middle of the day, which becomes more pronounced from month to month (Figure 1). This was caused by the shade effect of the adjacent building, which only affected the control sensor.
The linear model showed that the temperature differences at the measured time intervals for the full time series were significant at p < 0.001 for the three pairs tested: between the palm crown and the control (fence); between palm trunk and control; and between palm crown and tree. As a caveat, it should be noted that the datasets are very large and that, therefore, by default, the confidence intervals are narrow. Collapsing the time series to months, the differences between all sample pairs were significant at p < 0.001, with one palm crown vs. tree marginally less significant at p = 0.023 in November (Table S1).
When considering the hour of day, only two of the sample pairs were not significant at p < 0.05 or better. One was the pair crown vs. control of the Albury sample at noon (p = 0.867) and the other was the pair trunk vs. control of the Alma Park sample at 07:00 (p = 0.906) (Table S2). When collapsing the individual readings into time of day (dawn, day, dusk, and night), using the nautical twilight for the determination of ‘dawn’ and ‘dusk’ [69], the temperature differences were significant at p < 0.001 for both day and night in both the urban and the rural setting (Table 1). At the urban location the temperature differences during the transition phases (dawn and dusk) were significantly different as well (p < 0.001, except palm trunk vs. control which were identical at p = 0.011). At the rural location, however, the temperature differences during the transition phases were not significant (with the exception of palm trunk vs. control, p = 0.007) (Table 1).
3.2. Humidity
The average hourly values for humidity have been graphed in Figure 2. The curves for both November and April are more erratic, in particular during the morning hours, indicating variability. It should be noted, however, that the datasets for these two months are incomplete (only six and eleven days, respectively). The data show that in the urban setting, the humidity in the crown was lower than the humidity at both the control location and in the canopy of the deciduous trees.
This discrepancy was particularly pronounced in the middle of the southern summer (between January and February). The same applies to the rural setting, where the difference between the palm’s crown and the eucalypt tree was less pronounced.
The linear model showed that the humidity differences at the measured time intervals for the full time series were significant at p < 0.001 for the three pairs tested: between the palm crown and the control (fence); between palm trunk and control; and between palm crown and tree. As before, as a caveat it should be noted that the datasets are very large and that, therefore, by default the confidence intervals are narrow. Collapsing the time series to months, the differences between all sample pairs in the rural setting were significant at p < 0.001, with the differences of two sample pairs marginally less significant at p = 0.002 and p = 0.006 in December (Table S7). The urban setting provides a very different picture however. The humidity differences between the crown and the control/tree were significant at p < 0.001, with the exception of the crown vs. tree pair in November (which is almost statistically identical p = 0.946). The differences between the palm trunk and the control were insignificant with the exception of the months of November and March (both p < 0.001) (Table S7).
When considering the hour of day, the picture is less clear as well. While in the rural setting the differences between the palm (both crown and trunk) and control were significant at p < 0.001 (with the exception of 19:00; p = 0.325), the differences between the crown of the palm and the eucalypt were insignificant in the entire period between 17:00 and midnight, as well as at various hours during the morning (Table S8). At the urban location the humidity difference of the vast majority of hourly sample pairs was significant at p < 0.001, with a small number of pairs showing insignificant differences (but without an obvious pattern).
When collapsing the individual readings into time of day (dawn, day, dusk, and night), again using the nautical twilight for the determination of ‘dawn’ and ‘dusk’ [69], the temperature differences were significant at p < 0.001 for both day and night in both the urban and the rural setting (Table 2). At the urban location the temperature differences during the transition phases (dawn and dusk) were significantly different as well (p < 0.001, except palm trunk vs. control at dusk). At the rural location, however, the temperature differences during the transition phases were not significant with the exception of crown vs. tree (Table 2).
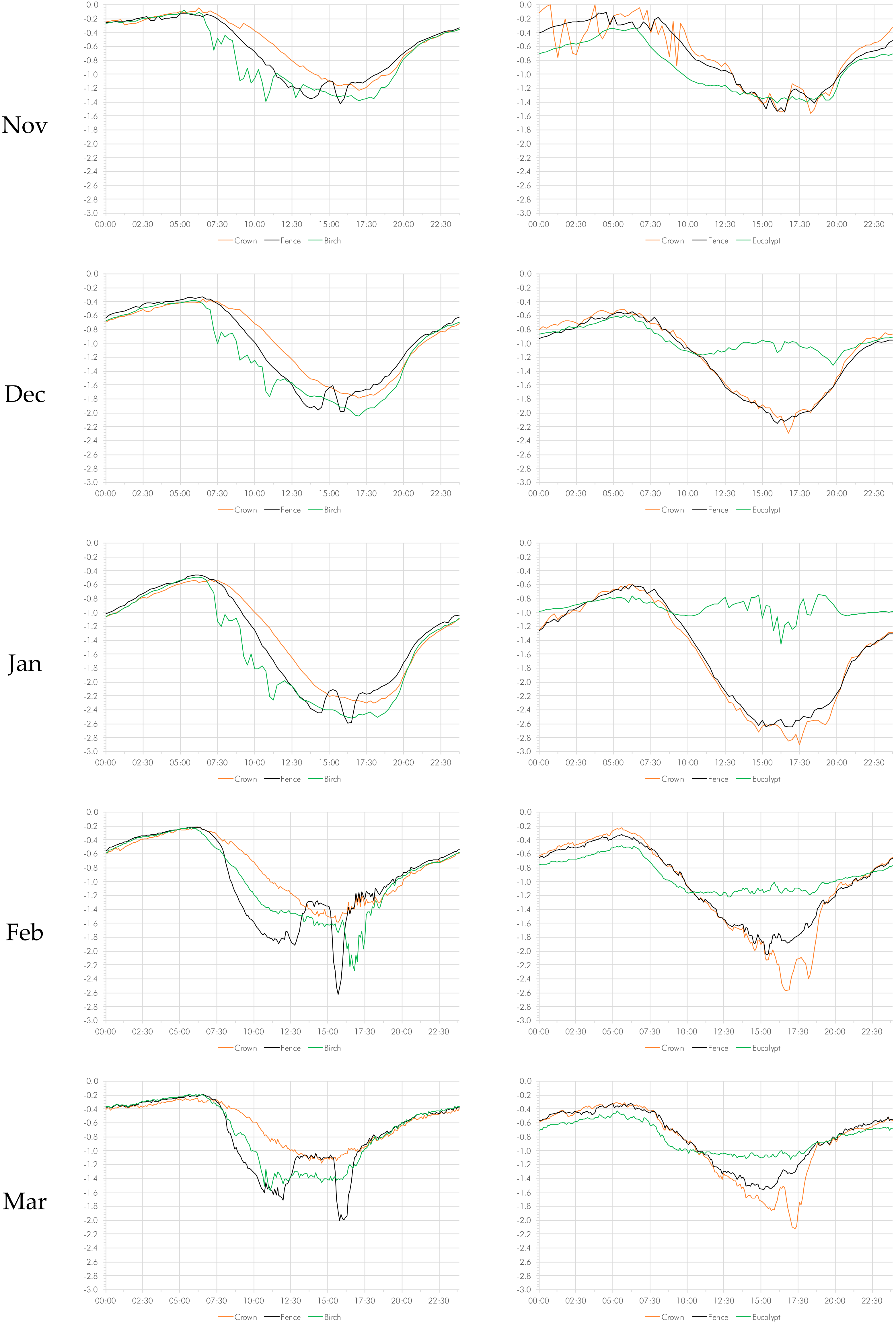
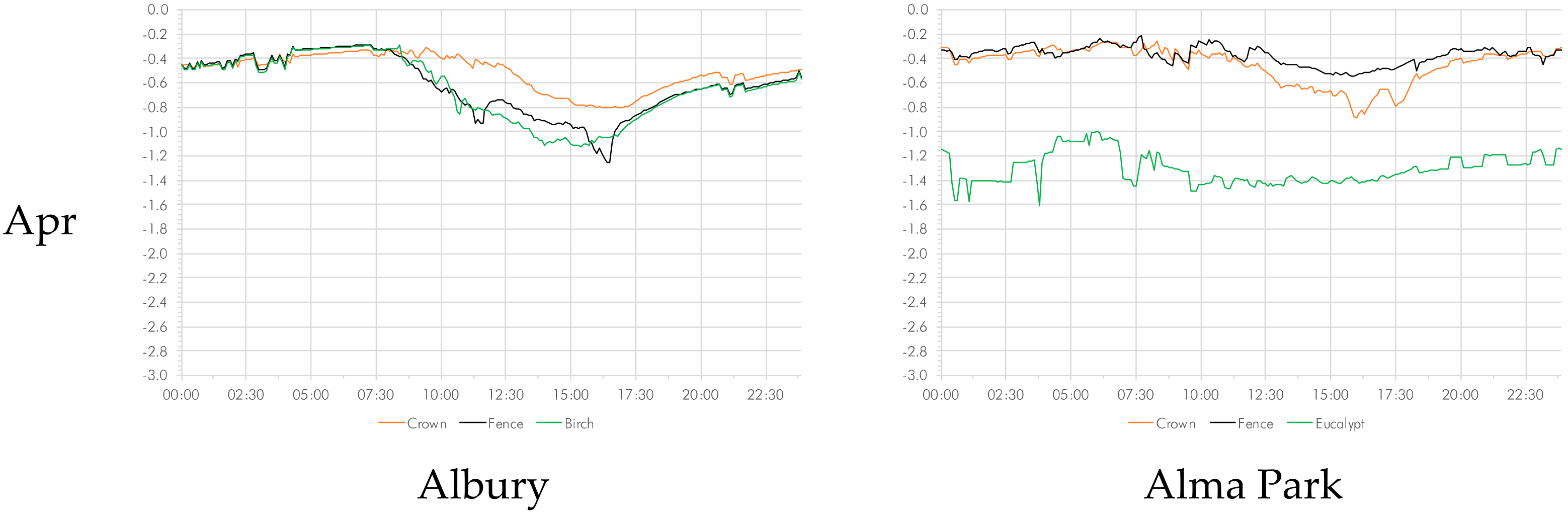
3.3. Vapor Pressure Deficit
As was the case for relative humidity, the curves for both November and April were more erratic, in particular during the morning hours, indicating variability (Figure 3). The data show that in the urban setting, the vapor pressure deficit in the crown was consistently lower than the vapor pressure deficit at both the control location and in the canopy of the deciduous tree. This discrepancy was particularly pronounced in the middle of the southern summer (January and February). In the rural setting, however, the difference between the palm’s crown and the eucalypt tree was inverted, with the vapor pressure deficit consistently lower in the eucalypt tree.
Temperature differences at the measured time intervals for the full time series were significant at p < 0.001 for the three pairs tested: between the palm crown and the control (fence); between palm trunk and control; and between palm crown and tree. Collapsing the time series to months, the differences between all sample pairs in the urban setting were significant at p < 0.001 (Table S9). The rural setting provides a different picture. The humidity differences between the crown and the tree were significant at p < 0.001 throughout, whereas there were differences between the crown vs. control pair (p < 0.001) except for November and December. The differences between the palm trunk and the control were significant (p < 0.001) with the exception of the months of Marchand April (Table S9).
When collapsing the individual readings into time of day (dawn, day, dusk, and night), no clear picture emerges (Table S10). In the urban rural setting, the vapor pressure deficit in the crown of the was significantly lower (p < 0.001) than that in measured at the control. The same applies to the comparison to the birch (p < 0.001) (with the exception of 6 a.m. and 8 p.m.) and to the comparison of trunk with control (p < 0.01) (except for 3 and 4 p.m.). In the rural setting, the pattern was less clear. The vapor pressure deficit in the crown was significantly higher (p < 0.01) than that in measured at the eucalypt. The differences between the other two pairs (crown vs. control; trunk vs. control) were by and large insignificant with the exception of the period between 11:00 and 19:00 p.m. p < 0.001).
3.4. Shade Modelling
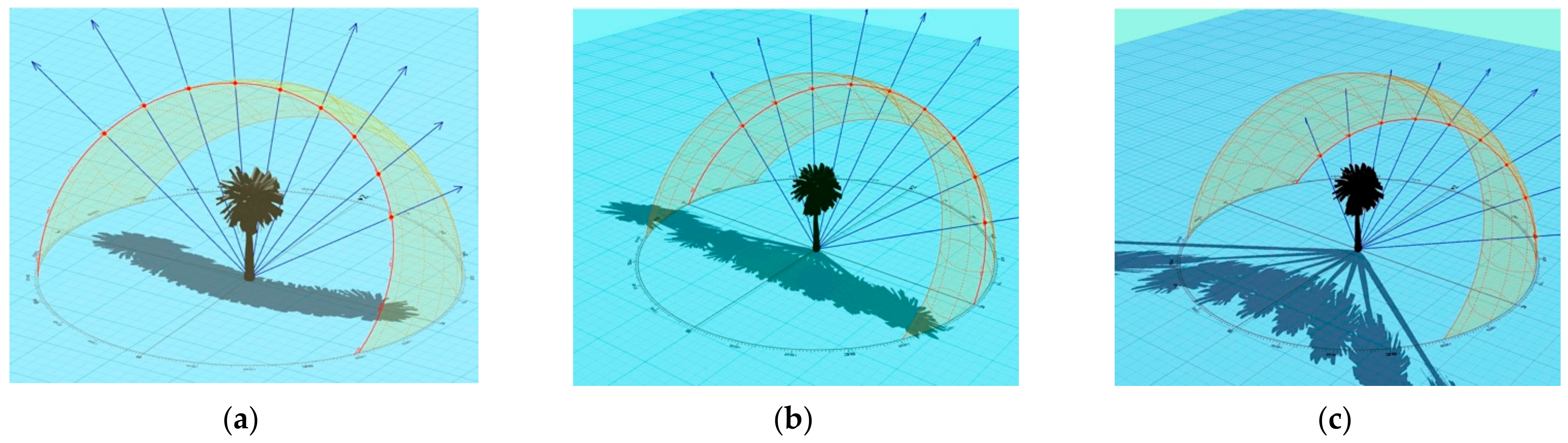
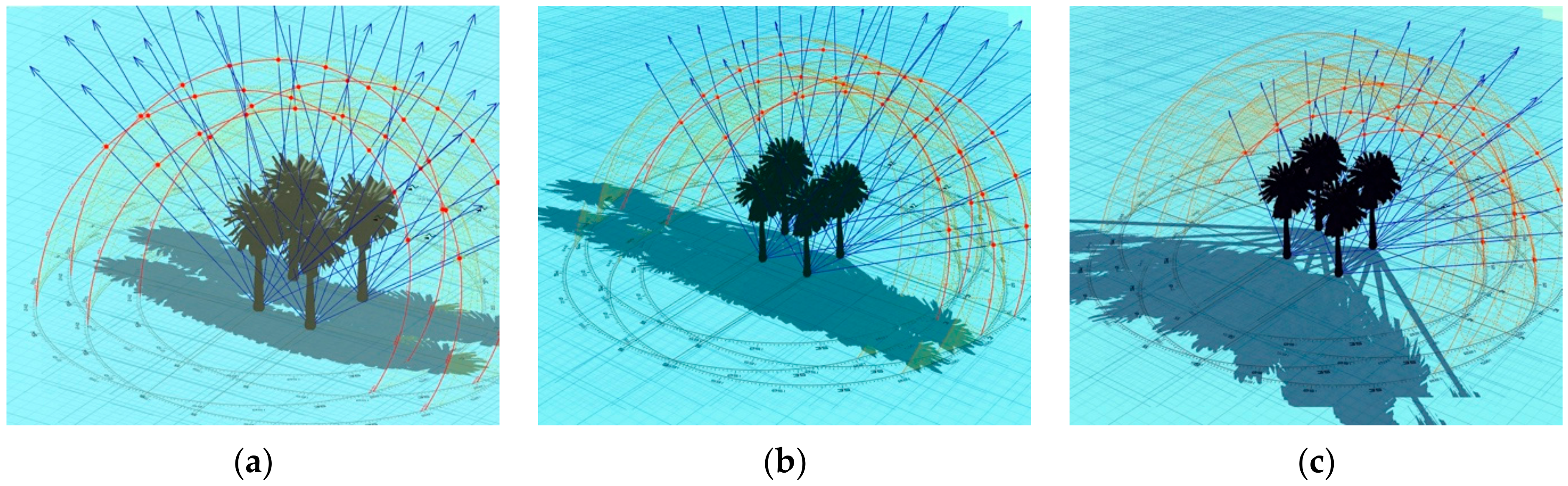
Depending on the angle of the sun, the shadow cast by a Canary Islands date palm has the semblance of a rosette when the sun was overhead (Figures S5 and S6a) or that of a feather duster in the late afternoon (Figures S4 and S6b). The shade modelling shows that, depending on the time of year and the time of day, the shadow will cover a wide radius, with shadows up to 10 m away from the base of the palm (depending on palm height) (Figure 4). The effects of height and of gradual dissipation of shadows with increasing stem height can be ameliorated by dense (trans-)plantings of mature palms, which can be set as close as 6 × 6 m, thereby forming a near continuous canopy (Figure 5 and Figure 6).
3.5. Tree-Benefits Analysis of Canary Islands Date Palms
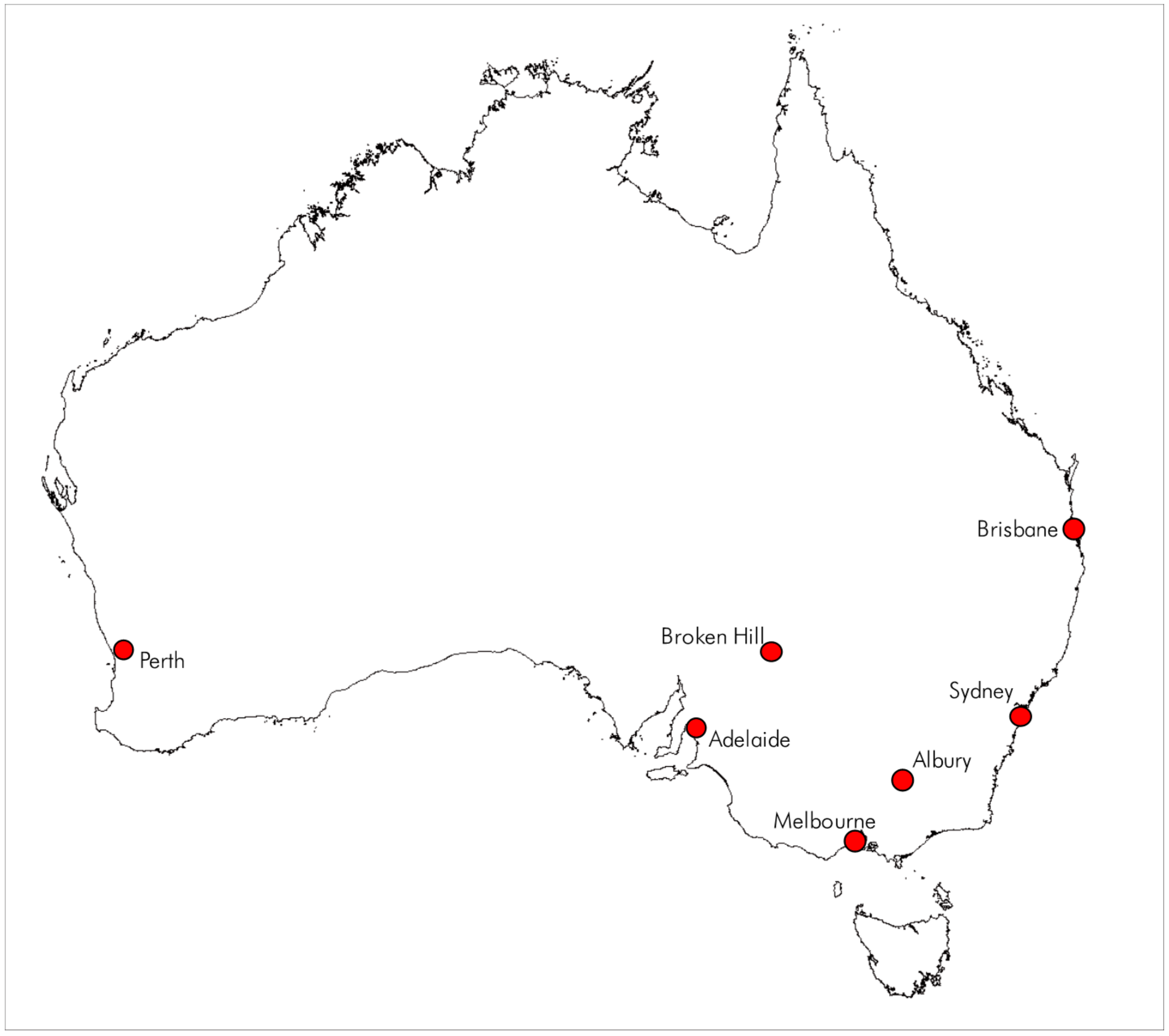
The cost-benefit analysis of Canary Islands date palms has been calculated for five Australian communities which are representative of the eastern Australian seaboard as well as an inland area (Figure 7) The shade effects date palms differ based on the climatic zone, with the highest benefits (relative to London Plane trees) in the warm inland zones (i.e., Albury and Broken Hill) (Table 3).
The analysis compared the performance of P. canariensis with five other tree species, one native (Eucalyptus sp.), three exotic trees with small (Betula pendula) and large leaves (Liquidambar styraciflua, Platanus × acerifolia), and one palm species (Washingtonia sp.). In terms of overall benefits, Canary Islands date palms performed better than all other species in the northern and inland areas, but were less beneficial in Sydney/Adelaide and Melbourne (Table 3) where eucalypts as well as the large-leaved Liquidambar styraciflua and Platanus × acerifolia provided the best returns. Californian and Mexican fan palms (Washingtonia sp.) had the lowest values throughout. Canary Islands date palms performed consistently better than birch trees, with the exception of CO2 sequestration. CO2 sequestration as well as stormwater amelioration where the weak points of Canary Islands date palms, mainly due to slow leaf production and low water requirements. The permanent canopy of palms also becomes a liability in situations where the shading during winter months reduces the needed solar radiation and thus increases heating costs (e.g., Melbourne).
Most palm genera are regarded as low maintenance plants [70]. The costs of maintaining Canary Islands date palms depend on the extent of pruning of old (or undesirable) leaves and the stem height of the palm. If leaves are freely allowed to age, autogenously abscise, and drop to the ground, then costs are minimal and limited to the removal of the fallen leaves. If pruning is desired, then the maintenance costs depend on the extent of the pruning (i.e., pruning of old leaves only or pruning of the crown into a ‘pineapple’ shape), the resulting pruning intervals, and the height of the palm (i.e., whether the use of cherry picker is required). Actual data are difficult to come by. Calegari notes an average cost of US $60.68 per plant for Canary Islands date palm maintenance at the University of Arizona campus. [71] Other sources suggest pruning costs between US $75 and $400 for palms of 10 m height or less and between US $200 and $1200 for palms of 18 m height and above [72]. The pruning frequency decreases as the severity of pruning increases. A severe pruning (but not a pineapple cut) has a triennial recurrence interval, while moderate pruning may require attention every second year. Mere removal of fallen leaves can occur as part of regular lawn and garden care and will cost (pro rata) less than US $20 p.a. This suggests that the benefits of palms well exceed the costs.
4. Discussion
This section will address the contribution made by Canary Islands date palms as providers of shade and as regulators of humidity and, therefore, servicing amenity values in urban landscape settings. The discussion of the resulting energy savings will then be expanded to address the suitability of the plant for various neighborhoods.
Not covered in the discussion will be the exoticism of the plant, which gave rise to its rapid introduction into the horticultural market during the late nineteenth century [50,51], and which is still a factor in the minds of many property owners who continue to plant Canary Islands date palms and other palm species to surround the suburban homes [35,37]. Also not discussed in detail, but important to be mentioned, are the broader benefits of amenity green spaces in urban areas for mental health [77,78,79], a sense of safety [80], and a general sense of place [81].
4.1. Provision of Shade by Canary Islands Date Palms
As the leaves of Canary Islands date palms are feather-like, they do not provide a full, light excluding shadow, but a shaded area or varied density where some sections are covered by solid linear shadows and other sections are in full or partial sunlight (Figures S4 and S6). The cumulative overlap of the leaves intensifies and consolidates this shadow. The closer to the stem, the more overlap and therefore the more shadow exists. As the palm leaves will readily move even in light wind, they do not provide constant but variable shadow coverage, depending on air movements, with the greatest movement, and thus shadow variability, close to the tip of the leaf. Compared to other street and amenity trees, deciduous or eucalypts, Canary Islands date palms with their denser crown cast a smaller, denser and more consistent shadow.
The density of the canopy of a Canary Islands date palm will vary between male and female plants as during the fruiting period the full infructescences will weigh down the lower leaves, thereby opening up the crown. This has an impact on the nature of the shadow at low light angles. As Canary Islands date palms only have a single growth point, its crown diameter remains the same irrespective of stem height once the plant is mature. In consequence, the shade provision is gradually dissipated and reduced as the palm gains height. This is, however a prolonged process as Canary Island date palms are comparatively slow growing, with reported growth rates of 0.3–0.5 m/year for under 20 year old plants, and slower rates (0.1–0.2 m/year) for older plants [82]. The availability of moisture, as well as winter temperatures, will also regulate the palm growth.
The experimental data demonstrate the thermoregulation effects of the canopy of Canary Islands date palms, with shading effects during the day and heat retention during the night. The dense and homogenous canopy results in a uniform temperature distribution that is less susceptible to wind effects (which move the leaves and branches on other three species). This can be illustrated by the temperature curve of the control tree in the urban setting which is significantly higher in during the mornings from November to January.
The analysis of the temperature time series showed less significant results during the dawn and dusk periods. This is not surprising as this is the period when the incoming solar energy ramps up (dawn) and declines (dusk), both points where the temperature curve for the Canary Islands date palm transitions through that of the control/tree from a temperate deficit (shade) during the day to a temperature surplus (heat retention) during the day.
The shading benefits of amenity trees in public spaces clearly depends on the nature and spatial layout of the space. The effects of trees on the urban microclimate is small in urban streets with deep open spaces which are shaded by multi-story buildings during mornings and later afternoons [83]. In such cases, trees with small canopy size such as Canary Islands date palm are suitable, as they are mostly needed for the shading and thermal comfort of pedestrians in the noon hours [56].
4.2. Conservation of Day-Time Heat by Canary Islands Date Palms
Canary Islands date palms have a singular growth point, with all leaves (sometimes erroneously called ‘fronds’) emerging from the top of the stem. The leaves of Canary Islands date palms are feather-like (pinnate) with a lenticular outline and emerge sequentially in the fashion of an upwards spiral. There are about the same number of developing leaves as there are mature leaves in the crown. [84] Because all leaves from a mid-crown point bend downwards, and individual leaves of different ‘generations’ overlap, an umbrella-like canopy is created. The lowest, and oldest, leaves bend almost parallel with the stem until such a time when they abscise.
The umbrella-like lower section of the crown, combined with the dense overlapping leaf matter, traps and retains some the day-time heat throughout the night, which is clearly evident in the temperature curves (Figure 1). This effect is more pronounced in an exposed rural setting. While the night-time heat retention effect is nowhere near as marked as the shade effect, it is still significant. The heat retention is created by the dense canopy of multilayered leaves that both traps the heat and slows down vertical escape. The effect of this on street-level amenity, however, will be significantly lessened with increasing palm height.
4.3. Regulation of Humidity by Canary Islands Date Palms
The dense overlapping leaf matter of the canopy creates an umbrella-like cover. The induplicate leaflets of the Canary Islands date palm have V-shaped cross-section opening to the top, which ensures that all precipitation is caught by one of the overlapping leaves and either directed via the petiole to the stem (in case of the leaves above the crown-midpoint) or directed to the crown’s perimeter. Very little (if any) rainfall percolates to the bottom of the lower leaves.
The dense overlapping leaf matter of the canopy also creates a trap for humidity, both relative humidity of the ambient air, evapotranspiration from the ground surface, and as evapotranspiration by the leaves of the palm. The effects of changes in temperature to relative humidity are incorporated when considering vapor pressure deficit which have a direct bearing on both the functioning of organisms [85] and the perceived amenity values of a street tree. The experimental data are ambiguous. In an open, unprotected rural setting, the canopy of a Canary Islands date palm has only a very minor effect on regulating the changes in temperature and humidity compare to the control sample, whereas the eucalypt tree causes a major reduction of the vapor pressure deficit during the hot summer months (Figure 3). In the urban setting, however, the vapor pressure deficit was consistently lower than that of both the control and the tree, in particular during daytime. It can be speculated that the more open, and the whole unprotected nature of the Canary Islands date palms in the rural setting, leads to an increased air exchange, exacerbated by light winds, and thus reduces the palm’s ability to trap humidity and create a microclimate. In the urban setting, any winds are buffered by the surrounding buildings as well as by street and feature trees. As ornamental Canary Islands date palms are predominantly planted in urban and suburban settings, these observations have greater relevance.
4.4. Tree Benefits Analysis
The Tree Benefits Analysis showed that Canary Islands date palm did not rate well in terms of CO2 sequestration as well as stormwater amelioration, mainly due to slow leaf production and low water requirements. The slow leaf production compared to deciduous large-leaved species is offset by reduced landscaping and leaf litter removal costs. The low water requirements can become an asset, as such trees are more resilient to low ware and even drought conditions [20]. This holds particularly true in the foreseeable future, as climate change scenarios for southern Australia not only predict continued reduction of rainfall [86] and increased temperatures, [87] but also forecast altered soil conditions which will impact ecosystems [88].
4.5. Suitability for Low-Income Neighbourhoods
While the presence of trees on private properties may not be correlated with property values [89], neighborhood studies have shown a correlation between developed green space and the socio-economic status of the residents. The poorer the neighborhoods, the fewer trees [10,90,91], with the cost of water and arboricultural maintenance identified as the main causes [92]. Given their hardiness, Canary Island date palms are well suited for such situations as they can thrive in many soil and climatic conditions (i.e., those which do not fall below −6 °C for extended periods or below −9 °C for shorter periods) [93,94,95,96,97]. Critically, once established, the water demands of Canary Island date palms are low compared to other ornamentals and the plants are relatively drought tolerant [20,35,98,99,100].
Moreover, the Canary Island date palms are comparatively low maintenance [70], as only 10–12 leaves are abscised annually, compared to the plethora of leaves shed by deciduous trees or eucalypts. Leaf removal is time consuming, however, as the palm leaves have very stiff, short leaflets near the base of the petiole which form sharp spines that can break off easily and embed themselves in body tissue [101,102,103]. Unlike the leaf matter produced by deciduous trees, which can only be composted, the size of the leaves and the fibrous nature of the petiole and rachis of Canary Islands date palms allows for the valorization of abscised leaves through manufacture of particle boards [104], biosorbents [105,106], and fuel [107,108].
4.6. Services to Urban Ecology
While this is not the place to discuss in depth the role that Canary Island date palms can play in novel ecosystems, it should be noted that they possess habitat value to a range of native and exotic species [109]. Due to the abundance of drupes produced by a single female plant (in excess of 30,000), Canary Island date palms contribute to the diet of frugivores [47,57], in particular during the winter months [110].
4.7. Structural Benefits for Landscaping in Urban Areas
From a landscaping perspective, Canary Islands date palms have a number of additional benefits that frequently tend to be overlooked in the discussion. By virtue of the structure of the stem, the Canary Islands date palms are resistant to damage by high-speed winds, such as hurricanes [111,112].
The allelopathic effects of these palms tend to be autoallelopathic and thus negligible in inhibiting seedling growth of other species [113], compared, for example, to eucalypts [114,115,116]. This ensures that Canary Islands date palms can be planted on grass, in ornamental flower beds, or in combination with low-level shade-tolerant shrubs. Moreover, the root pattern of palms is quite narrow in that they can tolerate being surrounded by hardened surfaces [33]. The narrow pattern also implies that palms can be planted much closer to foundations than other trees without imperiling structural integrity.
The growth pattern of Canary Islands date palms is uniform, spatially confined, and predictable. This ensures that, if planted correctly, the palms will not interfere with suburban powerlines as they age—unlike other shade trees that require continual pruning. Significantly, given the spatially confined growth form with an umbrella-like canopy, Canary Islands date palms provide shade without forcing landscape architects to forgo the usage of space underneath the trees. As the root development of palms is well confined, with no near major surface roots extending from the stem [33], the interstitial spaces between palms remain open and available for varied uses.
If planted in a regular, dense pattern, Canary Islands date palms can be used to create a continuous shade canopy. As this palm species is eminently suitable for transplantation in its mature stage, uniform shade coverage can be achieved with only marginal loss of useable ground surface. If marginal shade is acceptable at some points, then the palms can be spaced in a 10 × 10 m grid. If better coverage is required, a spacing of 7.5 × 7.5 m will result in a near contiguous coverage. The shade effects and other impacts will not vary substantially over time (unless the palm is very old and tall).
From a landscaping design point of view, the uniform growth pattern of the species, combined with the slow growth rate once the stem has reached 6–8 m, provides several advantages. The designed landscape will not change rapidly, and therefore provides long-term visual continuity of the original design and visual stability allowing to people to develop a sense of attachment to place.
5. Conclusions
The experimental study has shown that the crowns of Canary Islands date palms provide very significant cooling during day-time and a small, but significant, elevation of temperature at night. The uniform, spatially confined, and predictable growth form of these palms, if planted closely, allows one to provide contiguous shade cover while also allowing for usage of space underneath the trees. Canary Island date palms have proven to be hardy with low water requirements and comparatively low maintenance costs, making them well suited for low-income neighborhoods. Additionally, they provide habitats and contribute to the diet of frugivores. In view of the wide range of benefits, which outweigh the costs, Canary Islands date palms should be reconsidered in the repertoire of urban landscaping, particularly in areas where climate change predictions signal reduced availability of soil moisture.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/horticulturae7070201/s1: Table S1, Observed differences in average temperature by times of day (Albury); Table S2, Observed differences in average temperature by times of day (Alma Park); Table S3, Observed differences in average humidity by times of day (Albury); Table S4, Observed differences in average humidity by times of day (Alma Park); Table S5, Significance of temperature differences between measurement locations: months; Table S6, Significance of temperature differences between measurement locations: hours of day; Table S7, Significance of humidity differences between measurement locations: months; Table S8, Significance of humidity differences between measurement locations: hours of day; Table S9, Significance of humidity differences between measurement locations: months; Table S10, Significance of humidity differences between measurement locations: hours of day; Figure S1, Temperature and humidity sensors as deployed in Albury (NSW). Sensors 1–2 in palm; 3 on fence; 4 in birch tree. Base image: Land and Property Information; Figure S2, Temperature and humidity sensors as deployed at Alma Park (NSW). Sensors 5–6 in palm; 7 on fence; 8 in eucalypt tree. Base image: Land and Property Information; Figure S3, Examples of temperature recorder locations (photos by the author); Figure S4, Aerial views of shadows cast by palms in the Albury Botanic Gardens 2011–a, e; 2014–b; 2015–c, f, g; 2017–d, h, i. Image sources: 2011, 2015 and 2017 aerial image Albury City; 2014 photo; Figure S5, Examples of canopies seen from below. a–e Canary Island Date Palms; f–Mexican Fan Palm (The Corso, Manly, NSW, photos by the author 8 June 2019); Figure S6, Schematic shadow pattern of a Canary Islands date palm. Top: vertical midday sun; right: low-angle late afternoon sun.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
I am indebted to Wayne Robinson (Institute for Land, Water, and Socie ty; Charles Sturt University) for statistical support.
Conflicts of Interest
The author declares no conflict of interest.
References
- Wang, Y.; Chan, A.; Lau, G.N.C.; Li, Q.; Yang, Y.; Yim, S.H.L. Effects of urbanization and global climate change on regional climate in the Pearl River Delta and thermal comfort implications. Int. J. Climatol. 2019, 39, 2984–2997. [Google Scholar] [CrossRef]
- Aminipouri, M.; Rayner, D.; Lindberg, F.; Thorsson, S.; Knudby, A.J.; Zickfeld, K.; Middel, A.; Krayenhoff, E.S. Urban tree planting to maintain outdoor thermal comfort under climate change: The case of Vancouver’s local climate zones. Build. Environ. 2019, 158, 226–236. [Google Scholar] [CrossRef]
- Müller, N.; Kuttler, W.; Barlag, A.-B. Counteracting urban climate change: Adaptation measures and their effect on thermal comfort. Theor. Appl. Clim. 2014, 115, 243–257. [Google Scholar] [CrossRef] [Green Version]
- Thorsson, S.; Lindberg, F.; Björklund, J.; Holmer, B.; Rayner, D. Potential changes in outdoor thermal comfort conditions in Gothenburg, Sweden due to climate change: The influence of urban geometry. Int. J. Climatol. 2011, 31, 324–335. [Google Scholar] [CrossRef]
- Hatvani-Kovacs, G.; Belusko, M.; Skinner, N.; Pockett, J.; Boland, J. Heat stress risk and resilience in the urban environment. Sustain. Cities Soc. 2016, 26, 278–288. [Google Scholar] [CrossRef]
- Kovats, R.S.; Hajat, S. Heat stress and public health: A critical review. Annu. Rev. Public Health 2008, 29, 41–55. [Google Scholar] [CrossRef]
- Knowlton, K.; Rotkin-Ellman, M.; King, G.; Margolis, H.G.; Smith, D.; Solomon, G.; Trent, R.; English, P. The 2006 California heat wave: Impacts on hospitalizations and emergency department visits. Environ. Health Perspect. 2008, 117, 61–67. [Google Scholar] [CrossRef] [PubMed]
- De Barros, F.S.; Gonçalves, F.L.T.; Gobo, J.P.A.; Chiquetto, J.B. Analysis of the association between meteorological variables and mortality in the elderly applied to different climatic characteristics of the State of São Paulo, Brazil. Theor. Appl. Clim. 2021, 144, 327–338. [Google Scholar] [CrossRef]
- Kuchcik, M. Mortality and thermal environment (UTCI) in Poland—Long-term, multi-city study. Int. J. Biometeorol. 2020, 1–13. [Google Scholar] [CrossRef]
- Harlan, S.L.; Brazel, A.J.; Darrel Jenerette, G.; Jones, N.S.; Larsen, L.; Prashad, L.; Stefanov, W.L. In the shade of affluence: The inequitable distribution of the urban heat island. In Equity and the Environment; Emerald Group Publishing Limited: Bingley, UK, 2007; pp. 173–202. [Google Scholar]
- Zemtsov, S.; Shartova, N.; Varentsov, M.; Konstantinov, P.; Kidyaeva, V.; Shchur, A.; Timonin, S.; Grischchenko, M. Intraurban social risk and mortality patterns during extreme heat events: A case study of Moscow, 2010–2017. Health Place 2020, 66, 102429. [Google Scholar] [CrossRef]
- Brown, R.D.; Vanos, J.; Kenny, N.; Lenzholzer, S. Designing urban parks that ameliorate the effects of climate change. Landsc. Urban Plann. 2015, 138, 118–131. [Google Scholar] [CrossRef]
- Shashua-Bar, L.; Tsiros, I.X.; Hoffman, M. Passive cooling design options to ameliorate thermal comfort in urban streets of a Mediterranean climate (Athens) under hot summer conditions. Build. Environ. 2012, 57, 110–119. [Google Scholar] [CrossRef]
- Lee, H.; Mayer, H.; Chen, L. Contribution of trees and grasslands to the mitigation of human heat stress in a residential district of Freiburg, Southwest Germany. Landsc. Urban Plann. 2016, 148, 37–50. [Google Scholar] [CrossRef]
- Zaki, S.A.; Toh, H.J.; Yakub, F.; Mohd Saudi, A.S.; Ardila-Rey, J.A.; Muhammad-Sukki, F. Effects of roadside trees and road orientation on thermal environment in a tropical city. Sustainability 2020, 12, 1053. [Google Scholar] [CrossRef] [Green Version]
- Hiemstra, J.A.; Saaroni, H.; Amorim, J.H. The urban heat Island: Thermal comfort and the role of urban greening. In The Urban Forest; Springer: Berlin/Heidelberg, Germany, 2017; pp. 7–19. [Google Scholar]
- Zhang, Z.; Lv, Y.; Pan, H. Cooling and humidifying effect of plant communities in subtropical urban parks. Urban For. Urban Green. 2013, 12, 323–329. [Google Scholar] [CrossRef]
- Boudjellal, L.; Bourbia, F. An evaluation of the cooling effect efficiency of the oasis structure in a Saharan town through remotely sensed data. Int. J. Environ. Stud. 2018, 75, 309–320. [Google Scholar] [CrossRef]
- Abaas, Z.R. Impact of development on Baghdad’s urban microclimate and human thermal comfort. Alex. Eng. J. 2020, 59, 275–290. [Google Scholar] [CrossRef]
- Bouzaher, L.B.; Alkama, D. Palm trees reuses as sustainable element in the Sahara. The case of Ziban, as self-sustainable urban units. Energ. Procedia 2012, 18, 1076–1085. [Google Scholar] [CrossRef] [Green Version]
- Fahed, J.; Kinab, E.; Ginestet, S.; Adolphe, L. Impact of urban heat island mitigation measures on microclimate and pedestrian comfort in a dense urban district of Lebanon. Sustain. Cities Soc. 2020, 61, 102375. [Google Scholar] [CrossRef]
- Bencheikh, H.; Rchid, A. The effects of green spaces (Palme trees) on the microclimate in arides zones, case study: Ghardaia, Algeria. Energ. Procedia 2012, 18, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Hosek, L.-K.; Roloff, A. Species site matching: Selecting palms (Arecaceae) for urban growing spaces. Urban For. Urban Green. 2016, 20, 113–119. [Google Scholar] [CrossRef]
- McPherson, E.G.; Xiao, Q.; Johnson, N.; van Doorn, N.S.; Peper, P.J.; Albers, S. Data for: Shade factors for 149 taxa of in-leaf urban trees in the U.S.A. Mendeley Data V1 2018, 31. [Google Scholar] [CrossRef]
- Hodel, D.R. Biology of Palms and Implications for Management in the Landscape. Hortechnology 2009, 19, 676–681. [Google Scholar] [CrossRef] [Green Version]
- Watts, J.A. Picture taking in paradise: Los Angeles and the creation of regional identity, 1880–1920. Hist. Photogr. 2000, 24, 243–250. [Google Scholar] [CrossRef]
- Piana, P.; Watkins, C.; Balzaretti, R. The Palm Landscapes of the Italian Riviera. Landscapes 2019, 19, 43–65. [Google Scholar] [CrossRef]
- Casid, J.H. Sowing Empire: Landscape and Colonization; U of Minnesota Press: Minneapolis, MN, USA, 2015. [Google Scholar]
- Griggs, P. For shade, colour and in memory of sacrifice: Amenity and memorial tree planting in Queensland’s towns and cities, 1915–1955. Qld. Rev. 2015, 22, 30–48. [Google Scholar] [CrossRef]
- Emanuele, S.; Valeria, B.; Giuseppe, I.; Ezio, P.; Vittorio, F.; Salvatore, G. Urban Landscape Evolution as a Consequence of an Invasive Pest: The Case of a Small Sicilian Town. Landsc. Online 2017, 52, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Manachini, B.; Billeci, N.; Palla, F. Exotic insect pests: The impact of the Red Palm Weevil on natural and cultural heritage in Palermo (Italy). J. Cult. Herit. 2013, 14, e177–e182. [Google Scholar] [CrossRef] [Green Version]
- Noto, G.; Romano, D. Palms in the urban environment in the southern latitudes of Italy. Sci. Manag. Veg. Urban Environ. 1985, 195, 91–97. [Google Scholar] [CrossRef]
- Hodel, D.R.; Pittenger, D.R.; Downer, A.J. Palm Root Growth and Implications for Transplanting. J. Arboric. 2005, 31, 171–181. [Google Scholar]
- Hodel, D.R.; Downer, A.J.; Pittenger, D.R. Transplanting palms. HortTechnology 2009, 19, 686–689. [Google Scholar] [CrossRef] [Green Version]
- Hernández, M.; Morales, A.; Saurí, D. Ornamental plants and the production of nature (s) in the Spanish real estate boom and bust: The case of Alicante. Urban Geogr. 2014, 35, 71–85. [Google Scholar] [CrossRef] [Green Version]
- McDonald, G.V.; Schnelle, M.A.; Arnold, M.A. Palms on the Edge: Species and Strategies for Landscape Utilization. HortTechnology 2009, 19, 705–709. [Google Scholar] [CrossRef] [Green Version]
- Cismasu, C.; Crasnopolschi, A.; Culescu, M.; Tudora, I. Why we [don’t] love palm-trees? Landscape design between local identity and exoticism. Lucr. Științifice-Univ. Științe Agron. Med. Vet. Bucur. Ser. B Hortic. 2011, 55, 276–283. [Google Scholar]
- AlMohannadi, M.; Zaina, S.; Zaina, S.; Furlan, R. Integrated Approach for the Improvement of Human Comfort in the Public Realm: The Case of the Corniche, the Linear Urban Link of Doha. Am. J. Sociol. Res. 2015, 5, 89–100. [Google Scholar]
- Bull, C.J. New Conversations with an Old Landscape: Landscape Architecture in Contemporary Australia; Images Publishing: Melbourne, Australia, 2002. [Google Scholar]
- Calhoun, S. How Melbourne became the most liveable city in the world. Plan. News 2016, 42, 12. [Google Scholar]
- Jim, C.Y.; Liu, H. Species diversity of three major urban forest types in Guangzhou City, China. For. Ecol. Manag. 2001, 146, 99–114. [Google Scholar] [CrossRef]
- Crewe, K. Arizona native plants and the urban challenge. Landsc. J. 2013, 32, 215–229. [Google Scholar] [CrossRef]
- Byrne, J.; Jinjun, Y. Can urban greenspace combat climate change? Towards a subtropical cities research agenda. Aust. Plan. 2009, 46, 36–43. [Google Scholar] [CrossRef]
- Aguaron, E.; McPherson, E.G. Comparison of methods for estimating carbon dioxide storage by Sacramento’s urban forest. In Carbon Sequestration in Urban Ecosystems; Springer: Berlin/Heidelberg, Germany, 2012; pp. 43–71. [Google Scholar]
- Velasco, E.; Roth, M.; Norford, L.; Molina, L.T. Does urban vegetation enhance carbon sequestration? Landsc. Urban Plann. 2016, 148, 99–107. [Google Scholar] [CrossRef]
- Gochfeld, M. Starling roost-site selection: Preference for palm trees. Emu 1978, 78, 39–40. [Google Scholar] [CrossRef]
- Spennemann, D.H.R. Observations on the consumption and dispersal of Phoenix canariensis drupes by the Grey-headed flying fox (Pteropus poliocephalus). Europ. J. Ecol. 2018, 4, 41–49. [Google Scholar] [CrossRef]
- Rivera, D.; Obón, C.; Alcaraz, F.; Egea, T.; Carreño, E.; Laguna, E.; Santos, A.; Wildpret, W. A review of the nomenclature and typification of the Canary Islands endemic palm, Phoenix canariensis (Arecaceae). Taxon 2013, 62, 1275–1282. [Google Scholar] [CrossRef] [Green Version]
- Barrow, S.C. A monograph of Phoenix L. (Palmae: Coryphoideae). Kew Bull. 1998, 53, 513–575. [Google Scholar] [CrossRef]
- Spennemann, D.H.R. Canary Islands Palms (Phoenix canariensis) as ornamental plants. The first thirty years of the horticultural trade. Huntia 2019, 17, 79–102. [Google Scholar]
- Zona, S. The horticultural history of the Canary Island Date Palm (Phoenix canariensis). Gard. Hist. 2008, 36, 301–308. [Google Scholar] [CrossRef]
- Spennemann, D.H.R. Canary Islands Palms (Phoenix canariensis) in Australia: Introduction and early dispersal. Palms 2018, 62, 185–201. [Google Scholar]
- Spennemann, D.H.R. Geographical Distribution of Four Key Ornamental and Production Palm Species Phoenix canariensis, P. dactylifera, Washingtonia filifera and W. robusta; Institute for Land, Water and Society, Charles Sturt University: Albury, NSW, Australia, 2018. [Google Scholar]
- Pomeroy, J.; Director, B.M. Greening the urban habitat: Singapore. CTBUH J. 2012, 2012, 30–35. [Google Scholar]
- Georgi, J.N.; Dimitriou, D. The contribution of urban green spaces to the improvement of environment in cities: Case study of Chania, Greece. Build. Environ. 2010, 45, 1401–1414. [Google Scholar] [CrossRef] [Green Version]
- Shashua-Bar, L.; Potchter, O.; Bitan, A.; Boltansky, D.; Yaakov, Y. Microclimate modelling of street tree species effects within the varied urban morphology in the Mediterranean city of Tel Aviv, Israel. Int. J. Climatol. 2010, 30, 44–57. [Google Scholar] [CrossRef]
- Spennemann, D.H.R. Consumption of Canary Island Date Palm Phoenix canariensis drupes by Pied Currawongs Strepera graculina. Aust. Field Ornithol. 2020, 37, 201–211. [Google Scholar] [CrossRef]
- Spennemann, D.H.R. Phoenix Canariensis Seed Encountered in Scats and Ejecta Collected at Alma Park; Institute for Land, Water and Society, Charles Sturt University: Albury, NSW, Australia, 2018; p. 34. [Google Scholar]
- Spennemann, D.H.R. Background to the Palms at Alma Park (NSW) III: Glenalvon Homestead; Institute for Land, Water and Society, Charles Sturt University: Albury, NSW, Australia, 2020; p. 4. [Google Scholar]
- Spennemann, D.H.R.; Pike, M. Rites of Passage: Germination of regurgitated and defecated Phoenix canariensis seeds. Proc. Linn. Soc. NSW 2019, 141, 49–59. [Google Scholar]
- Allen, R.G.; Walter, I.A.; Elliott, R.L.; Howell, T.A.; Itenfisu, D.; Jensen, M.E.; Snyder, R.L. The ASCE Standardized Reference Evapotranspiration Equation; Technical Committee on Standardization of Reference Evapotranspiration, American Society of Civil Engineers: Reston, BA, USA, 2000. [Google Scholar]
- SPSS. IBM SPSS Statistics. Statistical Package for the Social Sciences, 25; International Business Machines Inc.: Armonk, NY, USA, 2017. [Google Scholar]
- IBM. GLM Repeated Measures. Available online: https://www.ibm.com/support/knowledgecenter/en/SSLVMB_24.0.0/spss/advanced/idh_glmr.html (accessed on 1 May 2020).
- Marsh, A. 3D SunPath; Andrew Marsh. 2014. Available online: http://andrewmarsh.com/apps/staging/sunpath3d.html (accessed on 1 May 2020).
- Larsen, K. Palm Tree 3D Model. 2012. Available online: https://free3d.com/3d-model/date-palm-2286.html (accessed on 1 May 2020).
- Casey Trees; Davey Tree Expert Co. National Tree Benefit Calculator. Available online: http://www.treebenefits.com/calculator/ (accessed on 1 May 2021).
- Australian Building Codes Board. Climate Zone Map Australia Wide. Available online: https://www.abcb.gov.au/Resources/Tools-Calculators/Climate-Zone-Map-Australia-Wide (accessed on 1 May 2021).
- Horne, R.; Hayles, C.; Hes, D.; Jensen, C.; Opray, L.; Wakefield, R.; Wasiluk, K. International comparison of building energy performance standards. In Report to Australian Greenhouse Office; Department of Environment and Heritage: Canberra, Australia, 2005. [Google Scholar]
- USNO. Rise, Set, and Twilight Definitions. Available online: https://aa.usno.navy.mil/faq/docs/RST_defs.php (accessed on 1 May 2020).
- Haynes, J.; McLaughlin, J.; Vasquez, L.; Hunsberger, A.G.B. Low-Maintenance Landscape Plants for South Florida; The Environmental Horticulture Department, Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, University of Florida: Gainesville, FL, USA, 2019; Volume ENH 854. [Google Scholar]
- Calegari, J. Assessing Suitability of Landscape Palm Trees in the Urban Environments of Southern Arizona; University of Arizona: Phoenix, AZ, USA, 2017. [Google Scholar]
- TreeRemoval. Palm Tree Removal & Trimming/How Much Does it Cost? Available online: http://www.treeremoval.com/costs/palm-tree-removal-and-trimming-prices/#.XPjnui3FLdQ (accessed on 1 May 2020).
- Rosenfeld, E. Effects of pruning on the health of palms. J. Arboric. 2009, 35, 294. [Google Scholar]
- Broschat, T.K. Pruning Palms; Institute of Food and Agricultural Sciences, University of Florida: Gainesville, FL, USA, 2011; Volume ENH 1182. [Google Scholar]
- Downer, A.J.; Hodel, D.R.; Mochizuki, M.J. Pruning landscape palms. HortTechnology 2009, 19, 695–699. [Google Scholar]
- Hosek, L.-K.; Roloff, A. Relations between cityscape-related and palm-inherent variables and the pruning state of urban Arecaceae suggest three reasons for overpruning. Urban For. Urban Green. 2015, 14, 975–981. [Google Scholar] [CrossRef] [Green Version]
- Callaghan, A.; McCombe, G.; Harrold, A.; McMeel, C.; Mills, G.; Moore-Cherry, N.; Cullen, W. The impact of green spaces on mental health in urban settings: A scoping review. J. Ment. Health 2020, 33, 212–222. [Google Scholar]
- Wolf, K.L.; Lam, S.T.; McKeen, J.K.; Richardson, G.R.; van den Bosch, M.; Bardekjian, A.C. Urban trees and human health: A scoping review. Int. J. Environ. Res. Public Health 2020, 17, 4371. [Google Scholar]
- Nutsford, D.; Pearson, A.; Kingham, S. An ecological study investigating the association between access to urban green space and mental health. Public Health 2013, 127, 1005–1011. [Google Scholar] [PubMed]
- Kuo, F.E.; Sullivan, W.C. Environment and crime in the inner city: Does vegetation reduce crime? Environ. Behav. 2001, 33, 343–367. [Google Scholar]
- Elmendorf, W. The importance of trees and nature in community: A review of the relative literature. Arboric. Urban For. 2008, 34, 152. [Google Scholar]
- Spennemann, D.H.R. The Ornamental Planting of Palms at the Western End of Dean Street, Albury (NSW) 1929–2019; Institute for Land, Water and Society Charles Sturt University: Albury, NSW, Australia, 2019. [Google Scholar]
- Coutts, A.M.; White, E.C.; Tapper, N.J.; Beringer, J.; Livesley, S.J. Temperature and human thermal comfort effects of street trees across three contrasting street canyon environments. Theor. Appl. Clim. 2016, 124, 55–68. [Google Scholar] [CrossRef]
- Dalrymple, N.K.; Fisher, J.B. The relationship between the number of expanded and developing leaves in shoot apices of palms. Am. J. Bot. 1994, 81, 1576–1581. [Google Scholar] [CrossRef]
- Anderson, D.B. Relative humidity or vapor pressure deficit. Ecology 1936, 17, 277–282. [Google Scholar] [CrossRef]
- Dey, R.; Lewis, S.C.; Arblaster, J.M.; Abram, N.J. A review of past and projected changes in Australia’s rainfall. Wiley Interdiscip. Rev. Clim. Chang. 2019, 10, e577. [Google Scholar] [CrossRef]
- Wang, B.; Li Liu, D.; Macadam, I.; Alexander, L.V.; Abramowitz, G.; Yu, Q. Multi-model ensemble projections of future extreme temperature change using a statistical downscaling method in south eastern Australia. Clim. Chang. 2016, 138, 85–98. [Google Scholar] [CrossRef]
- Gerzabek, M.H.; Bajraktarevic, A.; Keiblinger, K.; Mentler, A.; Rechberger, M.; Tintner, J.; Wriessnig, K.; Gartner, M.; Valenzuela, X.S.; Troya, A. Online Early. Soil Res. 2019, 57, 467–481. [Google Scholar]
- Pandit, R.; Polyakov, M.; Tapsuwan, S.; Moran, T. The effect of street trees on property value in Perth, Western Australia. Landsc. Urban Plann. 2012, 110. [Google Scholar] [CrossRef]
- Jenerette, G.D.; Harlan, S.L.; Brazel, A.; Jones, N.; Larsen, L.; Stefanov, W.L. Regional relationships between surface temperature, vegetation, and human settlement in a rapidly urbanizing ecosystem. Landsc. Ecol. 2007, 22, 353–365. [Google Scholar] [CrossRef]
- Schwarz, K.; Fragkias, M.; Boone, C.G.; Zhou, W.; McHale, M.; Grove, J.M.; O’Neil-Dunne, J.; McFadden, J.P.; Buckley, G.L.; Childers, D. Trees grow on money: Urban tree canopy cover and environmental justice. PLoS ONE 2015, 10, e0122051. [Google Scholar] [CrossRef] [Green Version]
- Cariñanos, P.; Calaza-Martínez, P.; O’Brien, L.; Calfapietra, C. The cost of greening: Disservices of urban trees. In The Urban Forest; Springer: Berlin/Heidelberg, Germany, 2017; pp. 79–87. [Google Scholar]
- Sakai, A.; Larcher, W. Frost Survival of Plants: Responses and Adaptation to Freezing Stress; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Larcher, W.; Winter, A. Frost susceptibility of palms: Experimental data and their interpretation. Principes 1981, 25, 143–152. [Google Scholar]
- Plugatar, Y.V.; Maksimov, A.P.; Trikoz, N.N.; Kovalev, M.; Khromov, A. PEЗУЛЬTATЫ ИHTPOДУKЦИИ ΦИHИKA KAHAPCKOГO (PHOENIX CANARIENSIS CHAB.) HA ЮЖHOM БEPEГУ KPЫMA. [Results of the introduction of the Canary Islands date Palm (Phoenix Canariensis Chab.) on the southern coast of Crimea]. BECTHИK BГУ CEPИЯ XИMИЯ БИOЛOГИЯ ΦAPMAЦИЯ [Proc. Appl. Bot. Genet. Breed.] 2018, 179, 221–229. [Google Scholar]
- Ruan, Z.; Liao, Q.; Ding, Y. Cold Resistance of introduced Phoenix canariensis in Xiamen. Chin. J. Trop. Agric. 2006, 26, 7–9. [Google Scholar]
- Jones, W.D. Effects of the 1978 freeze on native plants of Sonora, Mexico. Desert Plants 1979, 1, 33–36. [Google Scholar]
- Pittenger, D.R.; Downer, A.J.; Hodel, D.R.; Mochizuki, M. Estimating water needs of landscape palms in Mediterranean climates. HortTechnology 2009, 19, 700–704. [Google Scholar] [CrossRef] [Green Version]
- Cubino, J.P.; Subirós, J.V.; Lozano, C.B. Maintenance, modifications, and water use in private gardens of Alt Empordà, Spain. HortTechnology 2014, 24, 374–383. [Google Scholar] [CrossRef]
- May, P.B.; Livesley, S.J.; Shears, I. Managing and monitoring tree health and soil water status during extreme drought in Melbourne, Victoria. Arboric. Urban 2013, 39, 136–145. [Google Scholar]
- Adams, C.D.; Timms, F.J.; Hanlon, M. Phoenix date palm injuries: A review of injuries from the Phoenix date palm treated at the Starship Children’s Hospital. Aust. N. Z. J. Surg. 2000, 70, 355–357. [Google Scholar] [CrossRef]
- Cahill, N.; King, J.D. Palm thorn synovitis. J. Pediatr. Orthop. 1984, 4, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Cozen, L.; Fonda, M. Palm thorn injuries: Difficulty in diagnosis of late sequelae. Calif. Med. 1953, 79, 40. [Google Scholar] [PubMed]
- Ferrández-García, A.; Ferrández-Villena, M.; Ferrández-García, C.E.; García-Ortuño, T.; Ferrández-García, M.T. Potential use of Phoenix canariensis biomass in binderless particleboards at low temperature and pressure. BioResources 2017, 12, 6698–6712. [Google Scholar] [CrossRef] [Green Version]
- Martín-González, M.; González-Díaz, O.; Susial, P.; Araña, J.; Herrera-Melián, J.; Doña-Rodríguez, J.; Pérez-Peña, J. Reuse of Phoenix canariensis palm frond mulch as biosorbent and as precursor of activated carbons for the adsorption of Imazalil in aqueous phase. Chem. Eng. J. 2014, 245, 348–358. [Google Scholar] [CrossRef]
- Correia, R.; Gonçalves, M.; Nobre, C.; Mendes, B. Material valorization of arundo donax l. and Phoenix canariensis biomass as adsorbents of methylene blue dye. In Proceedings of the Wates: Solutions, Treatments and Opportunities 3rd International Conference, Viana Do Castelo, Portugal, 14–16 September 2015; pp. 112–114. [Google Scholar]
- Sajdak, M.; Velázquez-Martí, B.; López-Cortés, I. Quantitative and qualitative characteristics of biomass derived from pruning Phoenix canariensis hort. ex Chabaud. and Phoenix dactilifera L. Renew. Energy 2014, 71, 545–552. [Google Scholar] [CrossRef]
- Correia, R.; Gonçalves, M.; Nobre, C.; Mendes, B. Impact of torrefaction and low-temperature carbonization on the properties of biomass wastes from Arundo donax L. and Phoenix canariensis. Bioresour. Technol. 2017, 223, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Spennemann, D.H.R. The connective potential of vertebrate vectors responsible for the dispersal of the Canary Island date palm (Phoenix canariensis). Flora 2019, 259, 151468. [Google Scholar] [CrossRef]
- Spennemann, D.H.R. The contribution of the Canary Island date palm (Phoenix canariensis) to the winter diet of frugivores in novel ecosystems. Europ. J. Ecol. 2019, 5, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Duryea, M.L.; Kampf, E.; Littell, R.C.; Rodriguez-Pedraza, C.D. Hurricanes and the urban forest: II. Effects on tropical and subtropical tree species. Arboric. Urban For. 2007, 33, 98. [Google Scholar]
- Duryea, M.L.; Kampf, E.; Littell, R.C. Hurricanes and the urban forest: I. Effects on southeastern United States coastal plain tree species. Arboric. Urban For. 2007, 33, 83. [Google Scholar]
- Warrag, M.; AI-Wasel, A. Autoallelopathic Potential of Leaflets and Seeds on Seedling Growth of Date Palm (Phoenix dactylifera L.). J. Agric. Mar. Sci. 2000, 5, 7–10. [Google Scholar] [CrossRef] [Green Version]
- Del Moral, R.; Muller, C.H. The allelopathic effects of Eucalyptus camaldulensis. Am. Midl. Nat. 1970, 83, 254–282. [Google Scholar] [CrossRef]
- May, F.; Ash, J. An assessment of the allelopathic potential of Eucalyptus. Aust. J. Bot. 1990, 38, 245–254. [Google Scholar] [CrossRef]
- Ahmed, R.; Hoque, A.R.; Hossain, M.K. Allelopathic effects of leaf litters of Eucalyptus camaldulensis on some forest and agricultural crops. J. For. Res. 2008, 19, 19–24. [Google Scholar] [CrossRef]
Figure 1.
Diurnal variations in temperature on a monthly basis from November to April.

Figure 2.
Diurnal variations in relative humidity on a monthly basis from November to April.

Figure 3.
Diurnal variations in vapor pressure deficit on a monthly basis from November to April.


Figure 4.
Sun shadows of a 10 m tall palm at the Alma Park location.

Figure 5.
Cumulative shadows cast by a single 10 m tall palm between 8:00 and 16:00 (Alma Park location). (a) (southern) summer solstice; (b) (southern) autumn equinox; and (c) (southern) winter solstice.
Figure 5.
Cumulative shadows cast by a single 10 m tall palm between 8:00 and 16:00 (Alma Park location). (a) (southern) summer solstice; (b) (southern) autumn equinox; and (c) (southern) winter solstice.

Figure 6.
Cumulative shadows cast by a group of four 10 m tall palms between 8:00 and 16:00 (Alma Park location). (a) (southern) summer solstice; (b) (southern) autumn equinox; and (c) (southern) winter solstice.
Figure 6.
Cumulative shadows cast by a group of four 10 m tall palms between 8:00 and 16:00 (Alma Park location). (a) (southern) summer solstice; (b) (southern) autumn equinox; and (c) (southern) winter solstice.

Figure 7.
Map of Australia showing the locations mentioned in Table 3.
Figure 7.
Map of Australia showing the locations mentioned in Table 3.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Significance (p) of temperature differences between measurement locations: time of day.
| Albury | Alma Park | |||||
|---|---|---|---|---|---|---|
| Crown vs. Control | Trunk vs. Control | Crown vs. Birch | Crown vs. Control | Trunk vs. Control | Crown vs. Eucalypt | |
| Dawn | 0.000 | 0.000 | 0.000 | 0.111 | 0.084 | 0.338 |
| Day | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Dusk | 0.000 | 0.989 | 0.000 | 0.226 | 0.007 | 0.283 |
| Night | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
Table 2.
Significance (p) of humidity differences between measurement locations: time of day.
| Albury | Alma Park | |||||
|---|---|---|---|---|---|---|
| Crown vs. Control | Trunk vs. Control | Crown vs. Birch | Crown vs. Control | Trunk vs. Control | Crown vs. Eucalypt | |
| Dawn | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.120 |
| Day | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Dusk | 0.000 | 0.584 | 0.000 | 0.000 | 0.000 | 0.421 |
| Night | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.001 |
Table 3.
Tree Benefits (annual $ value) provided by Phoenix canariensis in % of London plane trees (45 cm stem Ø) at six Australian locations (see map Figure 7).
Table 3.
Tree Benefits (annual $ value) provided by Phoenix canariensis in % of London plane trees (45 cm stem Ø) at six Australian locations (see map Figure 7).
| Storm Water | Electricity | Gas | Air Quality | CO2 | Total | |
|---|---|---|---|---|---|---|
| Broken Hill | ||||||
| Betula pendula | 59.8 | 70.6 | 76.2 | 98.8 | 83.5 | 76.3 |
| Eucalyptus sp. | 124.4 | 108.0 | 101.0 | 107.9 | 142.6 | 112.5 |
| Phoenix canariensis | 82.1 | 103.2 | 123.8 | 166.3 | 54.6 | 111.6 |
| Platanus × acerifolia | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Liquidambar styraciflua | — | — | — | — | — | — |
| Washingtonia sp. | 0.2 | 72.3 | 77.2 | 48.3 | 35.3 | 53.8 |
| Albury | ||||||
| Betula pendula | 42.0 | 61.9 | 74.8 | 47.4 | 49.2 | 54.0 |
| Eucalyptus sp. | 117.6 | 84.8 | 64.5 | 90.6 | 164.6 | 94.0 |
| Phoenix canariensis | 100.8 | 99.1 | 88.4 | 126.4 | 72.4 | 107.2 |
| Platanus × acerifolia | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Liquidambar styraciflua | 53.6 | 121.9 | 43.9 | 57.3 | 100.6 | 86.7 |
| Washingtonia sp. | 30.1 | 12.0 | 20.0 | 12.8 | 16.6 | 15.4 |
| Brisbane | ||||||
| Betula pendula | 69.3 | 80.9 | 87.7 | 79.1 | 107.8 | 77.7 |
| Eucalyptus sp. | 65.4 | 55.8 | 46.7 | 60.3 | 62.9 | 60.2 |
| Phoenix canariensis | 92.4 | 88.1 | 104.0 | 100.9 | 52.3 | 90.0 |
| Platanus × acerifolia | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Liquidambar styraciflua | 62.3 | 62.4 | 65.9 | 62.0 | 82.2 | 63.9 |
| Washingtonia sp. | 17.5 | 16.1 | 17.1 | 17.7 | 9.0 | 16.5 |
| Adelaide/Sydney | ||||||
| Betula pendula | 79.1 | 75.5 | 54.0 | 77.6 | 49.3 | 75.2 |
| Eucalyptus sp. | 102.2 | 231.7 | 109.4 | 364.3 | 236.8 | 154.0 |
| Phoenix canariensis | 101.0 | 48.9 | 14.7 | 87.8 | 20.7 | 80.8 |
| Platanus × acerifolia | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Liquidambar styraciflua | 72.5 | 295.6 | 87.3 | 246.4 | 144.3 | 131.3 |
| Washingtonia sp. | 16.2 | 30.2 | 18.5 | 54.4 | 18.9 | 22.2 |
| Melbourne | ||||||
| Betula pendula | 67.7 | 97.1 | 43.0 | 67.4 | 59.2 | 79.9 |
| Eucalyptus sp. | 102.1 | 93.3 | 126.3 | 55.4 | 214.7 | 99.4 |
| Phoenix canariensis | 83.0 | 43.3 | 19.5 | 268.3 | 22.3 | 78.5 |
| Platanus × acerifolia | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Liquidambar styraciflua | 89.6 | 129.1 | 115.3 | 37.0 | 310.4 | 117.3 |
| Washingtonia sp. | 11.0 | 12.4 | 17.4 | 15.8 | 13.3 | 13.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Spennemann, D.H.R. The Role of Canary Island Date Palms in Physical Amenity Provisioning for Urban Landscape Settings. Horticulturae 2021, 7, 201. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7070201
AMA Style
Spennemann DHR. The Role of Canary Island Date Palms in Physical Amenity Provisioning for Urban Landscape Settings. Horticulturae. 2021; 7(7):201. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7070201
Chicago/Turabian StyleSpennemann, Dirk H. R. 2021. "The Role of Canary Island Date Palms in Physical Amenity Provisioning for Urban Landscape Settings" Horticulturae 7, no. 7: 201. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7070201
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.