
Genome-Wide Identification and Expression Analysis of AMT Gene Family in Apple (Malus domestica Borkh.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of AMT Gene Family Members in Apple
2.2. Phylogenetic Analysis, Gene Structures and Genomic Locations
2.3. Analysis on Phosphorylation of Residues within the C-Terminal Cytoplasmic Tail
2.4. Analysis on Putative Cis-Elements of AMT Genes in Apple
2.5. Transcription Modes of AMT Genes in Multifarious Tissues and Diurnal Rhythms
2.6. Plant Growth and Treatments
2.7. RNA Segregation and Quantitative RT–PCR Analysis
2.8. Functional Analysis in Yeast
3. Results
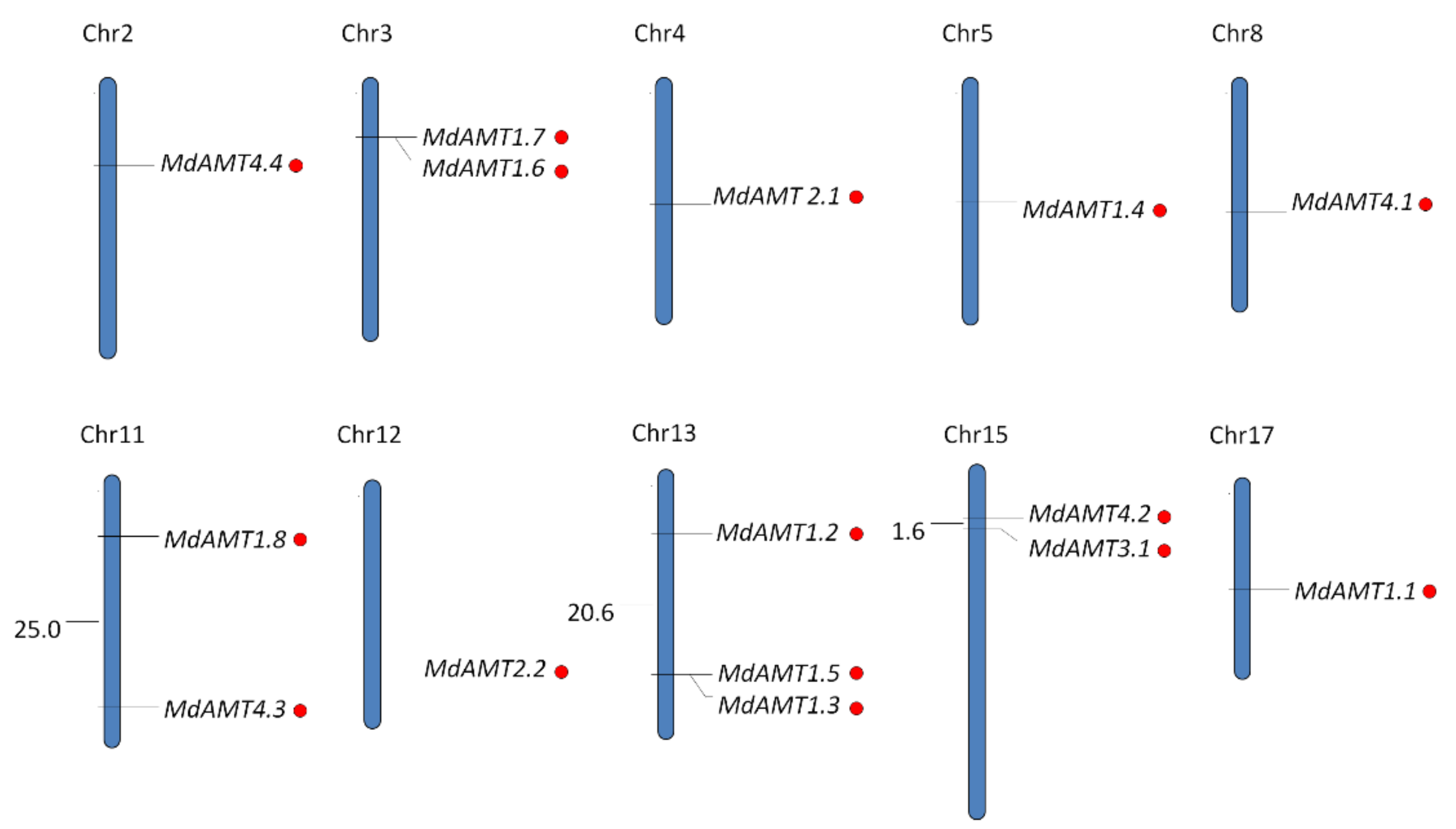
3.1. Identification and Chromosomal Location of AMT Genes in Apple

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Locus | Genomic Position | CDS Length | Protein Size | pI | MW (kDa) | TMHs |
|---|---|---|---|---|---|---|---|
| MdAMT1.1 | MDP0000427102 | chr17:14131094-14132698 | 1605 | 534 | 6.34 | 57.15 | 11 |
| MdAMT1.2 | MDP0000572721 | chr13:7142043-7143557 | 1515 | 504 | 6.55 | 54.12 | 11 |
| MdAMT1.3 | MDP0000266406 | chr13:27756622-27758139 | 1518 | 505 | 7.62 | 53.74 | 11 |
| MdAMT1.4 | MDP0000909614 | chr5:15840835-15842328 | 1494 | 497 | 6.82 | 52.88 | 9 |
| MdAMT1.5 | MDP0000182546 | chr13:27756243-27757760 | 1518 | 505 | 7.19 | 53.72 | 11 |
| MdAMT1.6 | MDP0000518257 | chr3:6372147-6373547 | 1401 | 466 | 5.10 | 50.13 | 10 |
| MdAMT1.7 | MDP0000240418 | chr3:6371890-6373290 | 1401 | 466 | 5.10 | 50.13 | 10 |
| MdAMT1.8 | MDP0000289954 | chr11:6610329-6611729 | 1401 | 466 | 5.18 | 50.06 | 9 |
| MdAMT2.1 | MDP0000650358 | chr4:16260628-16264571 | 1473 | 490 | 8.46 | 52.82 | 11 |
| MdAMT2.2 | MDP0000137929 | chr12:24732899-24735197 | 1473 | 490 | 8.66 | 52.69 | 11 |
| MdAMT3.1 | MDP0000245173 | chr15:7176000-7180690 | 1467 | 488 | 6.78 | 52.68 | 11 |
| MdAMT4.1 | MDP0000268493 | chr8:17433482-17435888 | 1503 | 500 | 8.77 | 54.52 | 9 |
| MdAMT4.2 | MDP0000177077 | chr15:5549862-5551942 | 1455 | 484 | 8.17 | 52.72 | 11 |
| MdAMT4.3 | MDP0000186795 | chr11:31604485-31607250 | 1416 | 471 | 6.24 | 51.58 | 10 |
| MdAMT4.4 | MDP0000211452 | chr2:10641333-10643931 | 1296 | 431 | 6.70 | 47.77 | 9 |
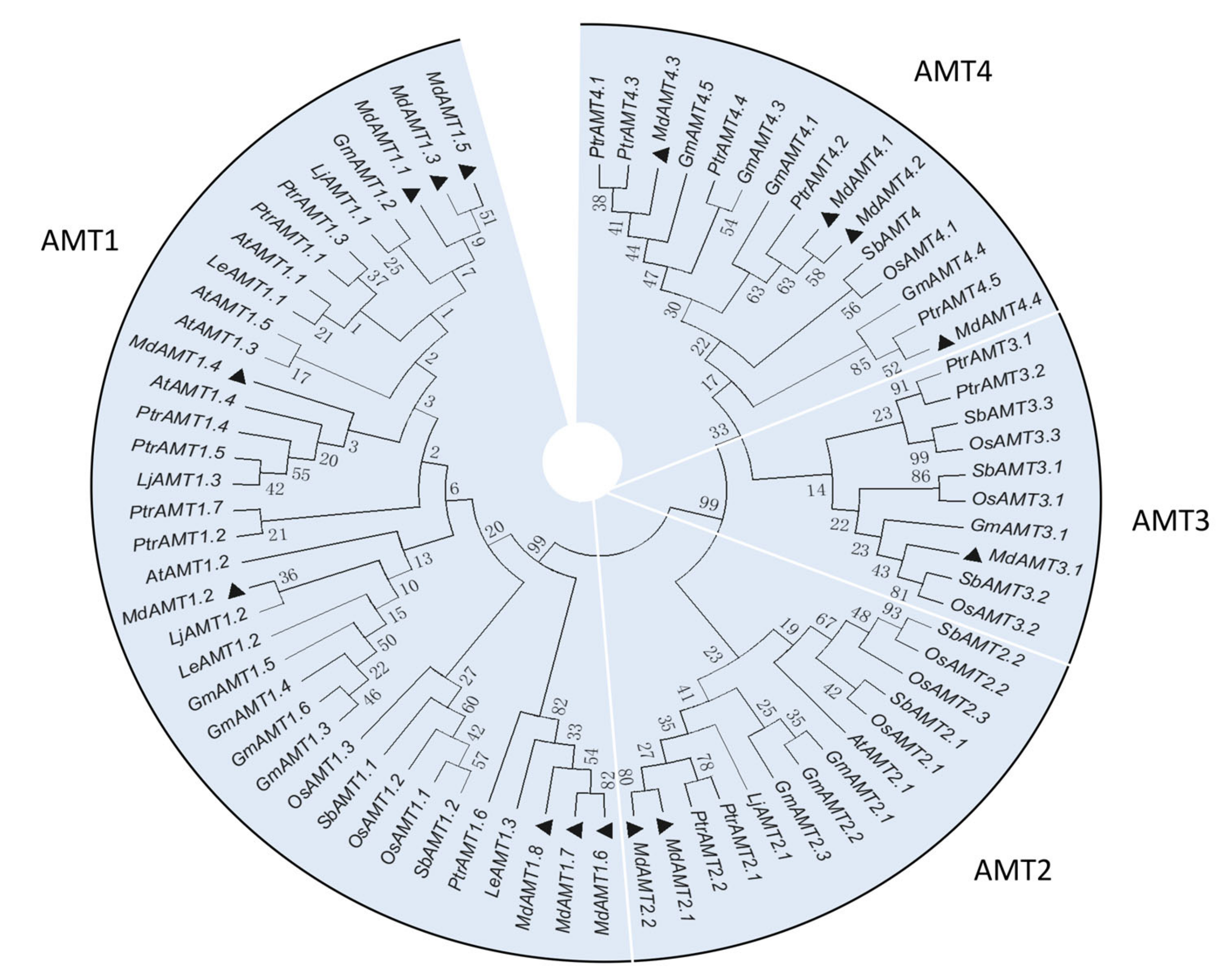
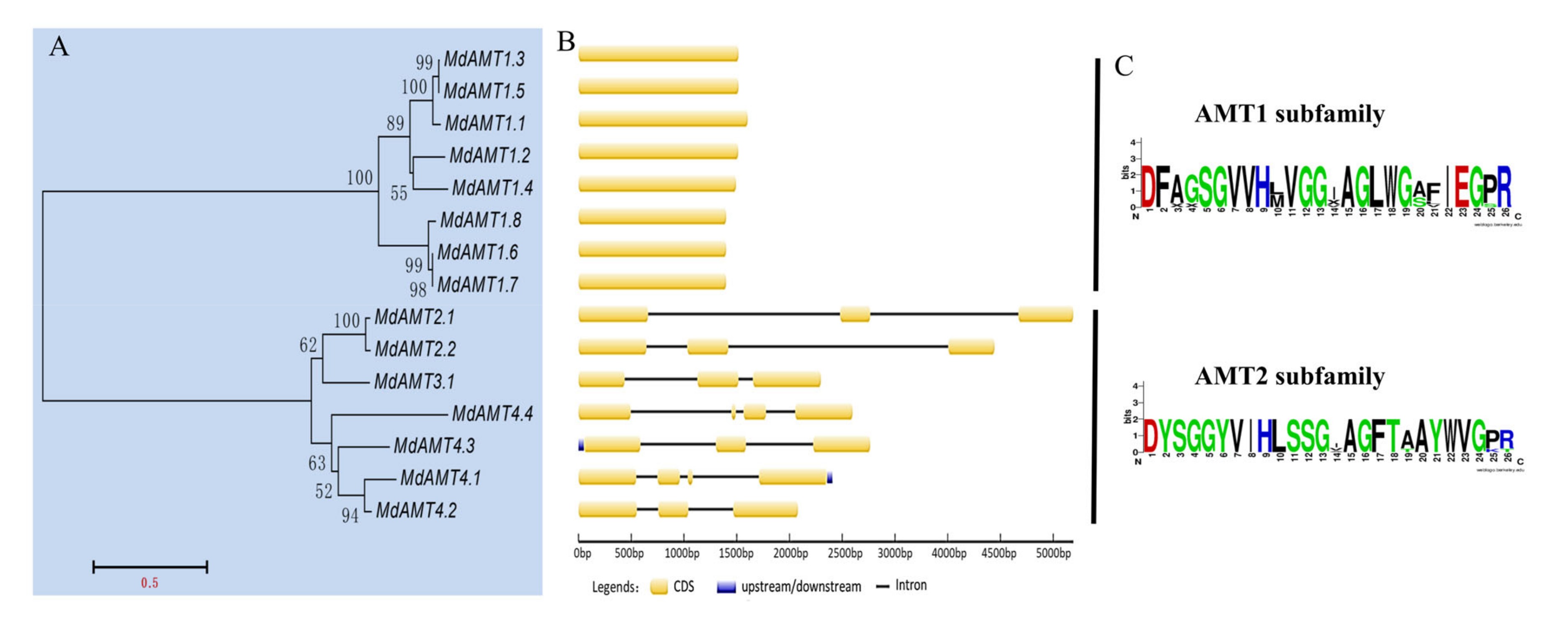
3.2. Phylogenetic Analysis and Gene Structure of AMT Genes
3.3. Analysis on Phosphorylation of Residues within the C-Terminal Cytoplasmic Tail
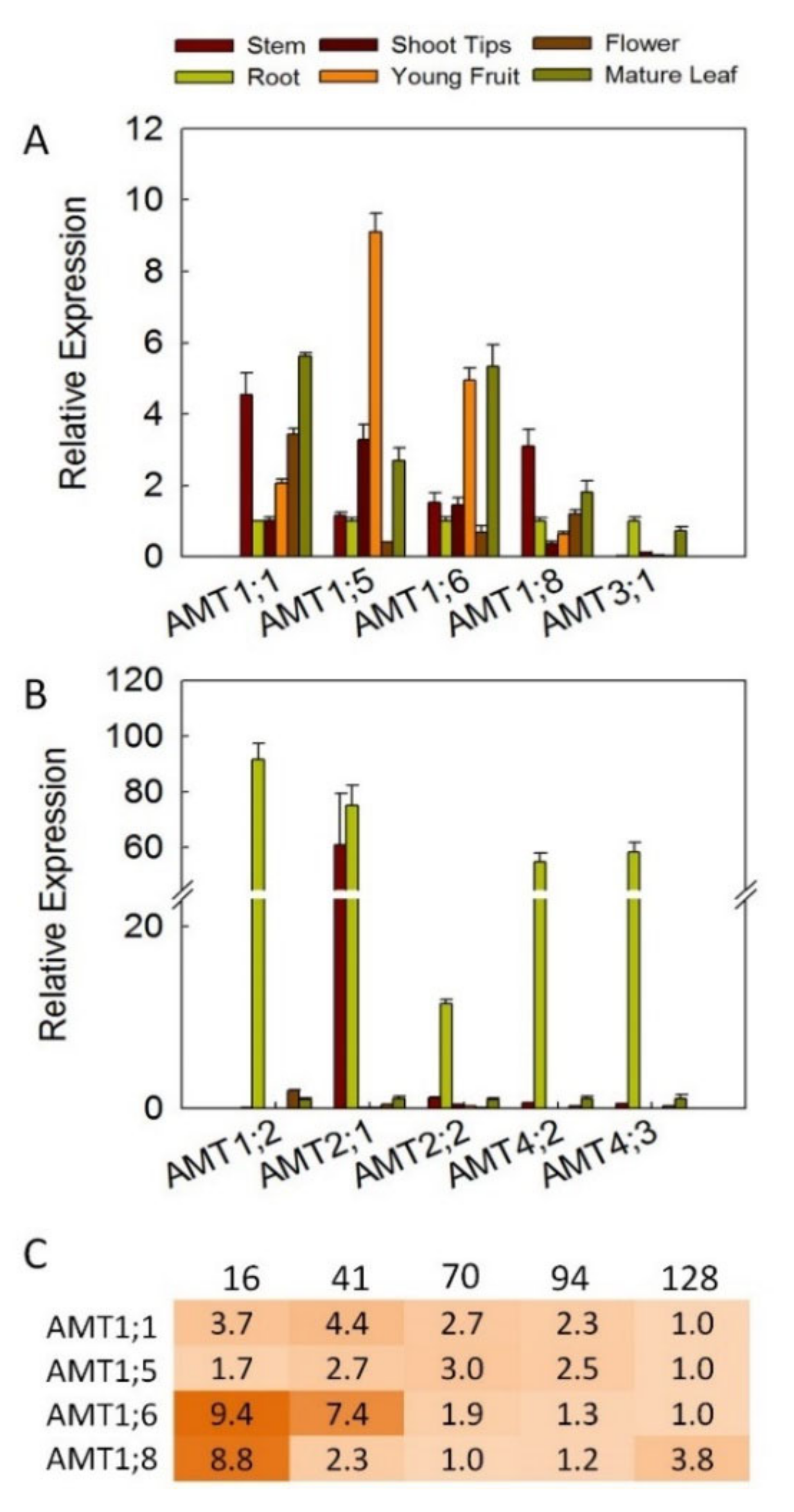
3.4. Expression Profiles for AMT Genes in Different Tissues
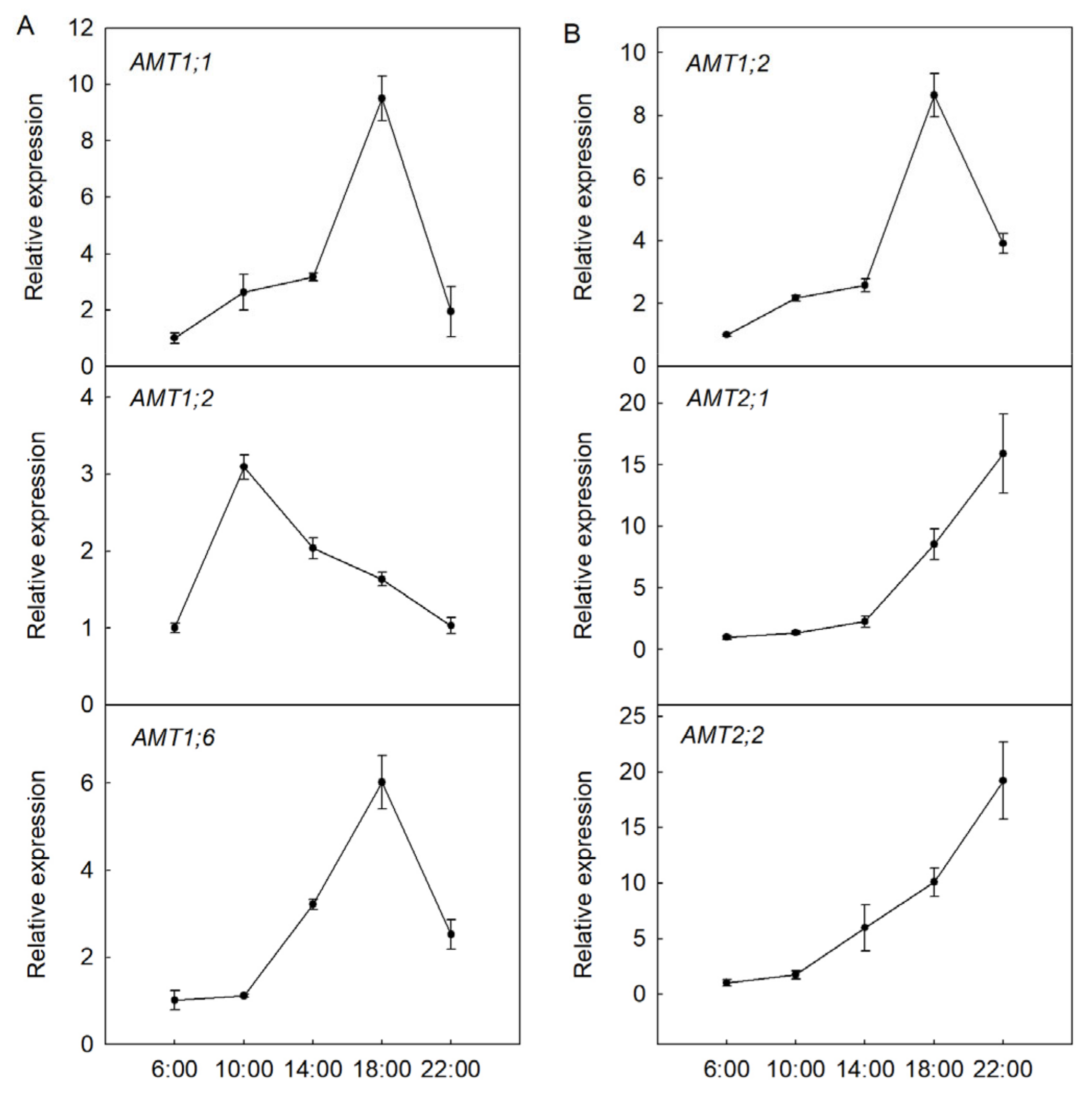
3.5. Transcription Modes of AMT Genes during Diurnal Rhythms
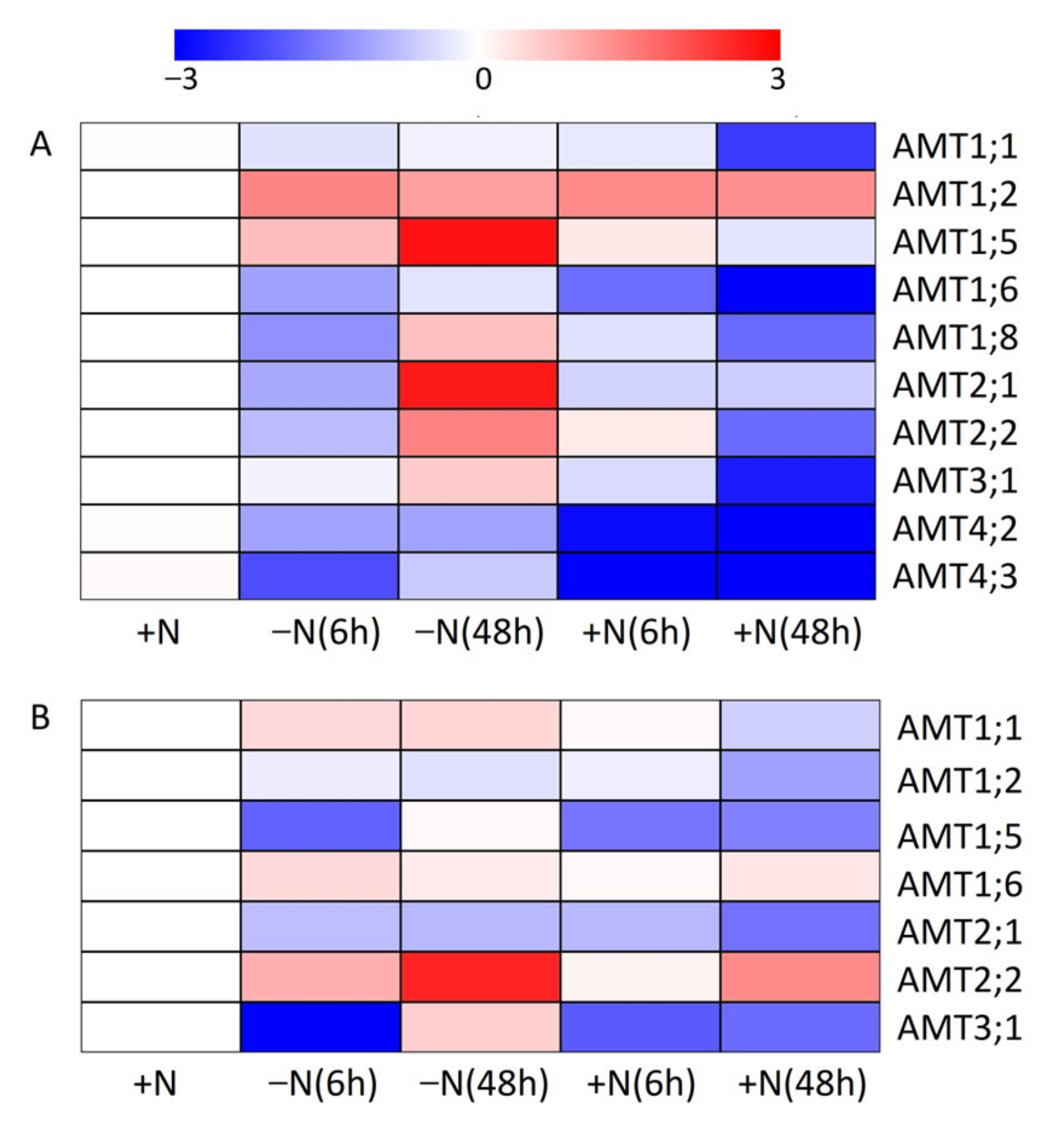
3.6. Impact of N-Starvation and Ammonium Resupply on Expression Profiles of AMT Genes
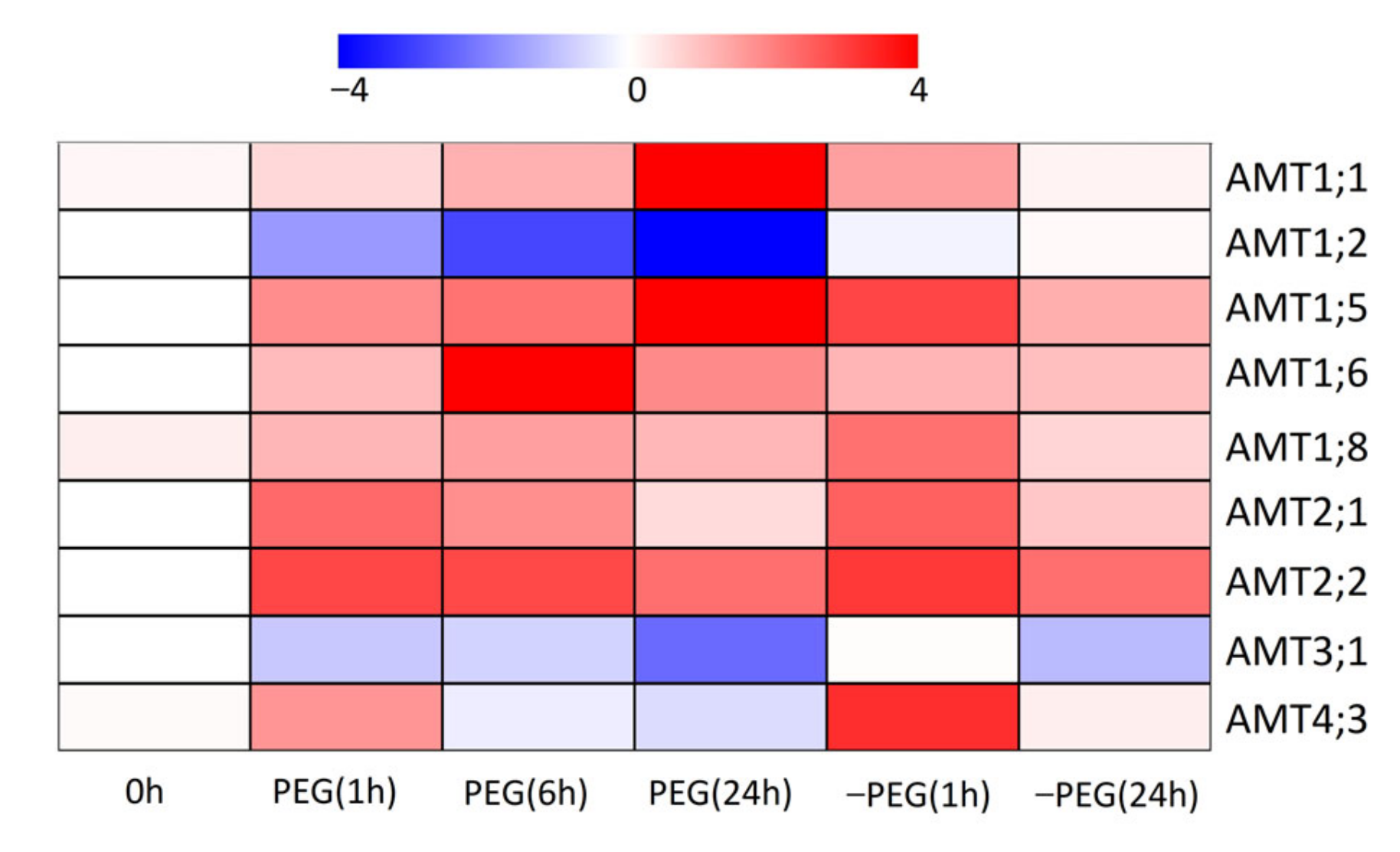
3.7. Effects of Osmotic Stress on Expression Patterns of AMT Genes
3.8. Functional Analysis of AMT1;2 in an Ammonium Uptake-Deficient Yeast Mutant
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Khasawneh, A.; Alsmairat, N.; Othman, Y.A.; Ayad, J.Y.; Al-Qudah, T.; Leskovar, D.I. Influence of nitrogen source on physiology, yield and fruit quality of young apricot trees. J. Plant Nutr. 2021, 44, 2597–2608. [Google Scholar] [CrossRef]
- Nava, G.; Dechen, A.R.; Nachtigall, R.G. Nitrogen and potassium fertilization affect apple fruit quality in Southern Brazil. Commun. Soil Sci. Plant Anal. 2007, 39, 96–107. [Google Scholar] [CrossRef]
- Yamasaki, A.; Yano, T. Effect of supplemental application of fertilizers on flower bud initiation and development of strawberry-possible role of nitrogen. Acta Hortic. 2009, 842, 765–768. [Google Scholar] [CrossRef]
- Bloom, A.J.; Sukrapanna, S.S.; Warner, R.L. Root respiration associated with ammonium and nitrate absorption and assimilation in barley. Plant Physiol. 1992, 99, 1294–1301. [Google Scholar] [CrossRef] [Green Version]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef] [Green Version]
- Couturier, J.; Montanini, B.; Martin, F.; Brun, A.; Blaudez, D.; Chalot, M. The expanded family of ammonium transporters in the perennial poplar plant. New Phytolo. 2007, 174, 137–150. [Google Scholar] [CrossRef]
- Yuan, L.X.; Loqué, D.; Kojima, S.; Rauch, S.; Ishiyama, K.; Takahashi, H.; Von Wirén, N. The organization of high-affinity ammonium uptake in Arabidopsis roots depends on the spatial arrangement and biochemical properties of AMT1-type transporters. Plant Cell 2007, 19, 2636–2652. [Google Scholar] [CrossRef] [Green Version]
- Tang, M.; Li, Y.; Chen, Y.; Han, L.; Zhang, H.; Song, Z. Characterization and Expression of Ammonium Transporter in Peach (Prunus persica) and Regulation Analysis in Response to External Ammonium Supply. Phyton 2020, 89, 4. [Google Scholar] [CrossRef]
- Ludewig, U.; Wilken, S.; Wu, B.; Jost, W.H.; Obrdlik, P.; Bakkoury, M.E.; Marini, A.; André, B.; Hamacher, T.; Boles, E.; et al. Homo- and hetero-oligomerization of ammonium transporter-1 NH4+ uniporters. J. Biol. Chem. 2003, 278, 45603–45610. [Google Scholar] [CrossRef] [Green Version]
- Loqué, D.; Yuan, L.; Kojima, S.; Gojon, A.; Wirth, J.; Gazzarrini, S.; Ishiyama, K.; Takahashi, H.; Von Wirén, N. Additive contribution of AMT1;1 and AMT1;3 to high-affinity ammonium uptake across the plasma membrane of nitrogen-deficient Arabidopsis roots. Plant J. 2006, 48, 522–534. [Google Scholar] [CrossRef]
- Konishi, N.; Ma, J. Three polarly localized ammonium transporter 1 members are cooperatively responsible for ammonium uptake in rice under low ammonium condition. New Phytolo. 2021, 232, 1778–1792. [Google Scholar] [CrossRef]
- Loqué, D.; Lalonde, S.; Looger, L.L.; Von Wirén, N.; Frommer, W.B. A cytosolic trans-activation domain essential for ammonium uptake. Nature 2007, 446, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Straub, T.; Ludewig, U.; Neuhäuser, B. The kinase CIPK23 inhibits ammonium transport in Arabidopsis thaliana. Plant Cell 2017, 29, 409–422. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.X.; Li, Y.; Yang, X.X.; Li, H.J.; Shen, Q.R.; Guo, S.W. Ammonium nutrition increases water absorption in rice seedlings (Oryza sativa L.) under water stress. Plant Soil 2010, 331, 193–201. [Google Scholar] [CrossRef]
- Ding, L.; Gao, C.; Li, Y.; Zhu, Y.; Xu, G.; Guo, S.W. The enhanced drought tolerance of rice plants under ammonium is related to aquaporin (AQP). Plant Sci. 2015, 234, 14–21. [Google Scholar] [CrossRef]
- Kant, S.; Kant, P.; Lips, H.; Barak, S. Partial substitution of NO3− by NH4+ fertilization increases ammonium assimilating enzyme activities and reduces the deleterious effects of salinity on the growth of barley. J. Plant Physiol. 2007, 164, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Crespo, E.; Camanes, G.; García-Agustín, P. Ammonium enhances resistance to salinity stress in citrus plants. J. Plant Physiol. 2012, 169, 1183–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.L.; Li, M.J.; Shao, Y.; Sun, T.T.; Li, C.Y.; Ma, F.W. Ammonium uptake increases in response to PEG-induced drought stress in Malus hupehensis Rehd. Environ. Exp. Bot. 2018, 151, 32–42. [Google Scholar] [CrossRef]
- Meng, S.; Zhang, C.X.; Su, L.; Li, Y.M.; Zhao, Z. Nitrogen uptake and metabolism of Populus simonii in response to PEG-induced drought stress. Environ. Exp. Bot. 2016, 123, 78–87. [Google Scholar] [CrossRef]
- Bu, Y.; Takano, T.; Liu, S. The role of ammonium transporter (AMT) against salt stress in plants. Plant Signaling Behav. 2019, 14, 8. [Google Scholar] [CrossRef]
- Shelden, M.C.; Dong, B.; de Bruxelles, G.L.; Trevaskis, B.; Whelan, J.; Ryan, P.R.; Howitt, S.M.; Udvardi, M.K. Arabidopsis ammonium transporters, AtAMT1;1 and AtAMT1;2, have different biochemical properties and functional roles. Plant Soil 2001, 231, 151–160. [Google Scholar] [CrossRef]
- Gu, R.; Duan, F.; An, X.; Zhang, F.; Von Wirén, N.; Yuan, L. Characterization of AMT-mediated high-affinity ammonium uptake in roots of maize (Zea mays L.). Plant Cell Physiol. 2013, 54, 1515–1524. [Google Scholar] [CrossRef]
- Liu, R.H.; Meng, J.L. MapDraw: A Microsoft Excel macro for drawing genetic linkage maps based on given genetic linkage data. Yi Chuan 2003, 25, 317–321. [Google Scholar]
- Chang, S.; Puryear, J.; Cairney, J. Simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Sun, T.; Jia, D.; Huang, L.L.; Shao, Y.; Ma, F.W. Comprehensive genomic identification and expression analysis of the nucleobase-ascorbate transporter (NAT) gene family in apple. Sci. Hortic. 2016, 198, 473–481. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Marini, A.M.; Soussi-Boudekou, S.; Vissers, S.; Andre, B. A family of ammonium transporters in Saccharomyces cerevisiae. Mol. Cell Biol. 1997, 17, 4282–4293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.Y.; Hao, D.L.; Cong, Y.; Jin, M.; Su, Y.H. The rice OsAMT1;1 is a proton-independent feedback regulated ammonium transporter. Plant Cell Rep. 2015, 34, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Lanquar, V.; Loqué, D.; Hörmann, F.; Yuan, L.; Bohner, A.; Engelsberger, W.R.; Lalonde, S.; Schulze, W.X.; Von Wirén, N.; Frommer, W.B. Feedback inhibition of ammonium uptake by a phospho-dependent allosteric mechanism in Arabidopsis. Plant Cell 2009, 21, 3610–3622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, D.L.; Zhou, J.Y.; Yang, S.Y.; Qi, W.; Yang, K.J.; Su, Y.H. Function and Regulation of Ammonium Transporters in Plants. Int. J. Mol. Sci. 2020, 21, 3557. [Google Scholar] [CrossRef] [PubMed]
- Ninnemann, O.; Jauniaux, J.C.; Frommer, W.B. Identification of a high affinity NH4+ transporter from plants. EMBO J. 1994, 13, 3464–3471. [Google Scholar] [CrossRef]
- Sonoda, Y.; Ikeda, A.; Saiki, S.; Yamaya, T.; Yamaguchi, J. Feedback regulation of the ammonium transporter gene family AMT1 by glutamine in rice. Plant Cell Physiol. 2003, 44, 1396–1402. [Google Scholar] [CrossRef] [Green Version]
- Giehl, R.F.; Laginha, A.M.; Duan, F.; Rentsch, D.; Yuan, L.; Von Wirén, N. A Critical Role of AMT2;1 in Root-To-Shoot Translocation of Ammonium in Arabidopsis. Mol. Plant 2017, 10, 1449–1460. [Google Scholar] [CrossRef]
- Von Wirén, N.; Lauter, F.R.; Ninnemann, O.; Gillissen, B.; Walch-Liu, P.; Engels, C.; Jost, W.H.; Frommer, W.B. Differential regulation of three functional ammonium transporter genes by nitrogen in root hairs and by light in leaves of tomato. Plant J. Cell Mol. Biol. 2000, 21, 167–175. [Google Scholar] [CrossRef]
- Lee, Y.H.; Tegeder, M. Selective expression of a novel high-affinity transport system for acidic and neutral amino acids in the tapetum cells of Arabidopsis flowers. Plant J. 2004, 40, 60–74. [Google Scholar] [CrossRef]
- Yuan, L.; Graff, L.; Loqué, D.; Kojima, S.; Tsuchiya, Y.N.; Takahashi, H.; Von Wirén, N. AtAMT1;4, a Pollen-Specific High-Affinity Ammonium Transporter of the Plasma Membrane in Arabidopsis. Plant Cell Physiol. 2009, 50, 13–25. [Google Scholar] [CrossRef]
- D’Apuzzo, E.; Rogato, A.; Simon-Rosin, U.; Alaoui, H.E.; Barbulova, A.; Betti, M.; Dimou, M.; Katinakis, P.; Márquez, A.J.; Marini, A.; et al. Characterization of three functional high-affinity ammonium transporters in Lotus japonicus with differential transcriptional regulation and spatial expression. Plant Physiol. 2004, 134, 1763–1774. [Google Scholar] [CrossRef] [Green Version]
- Ranathunge, K.; El-Kereamy, A.; Gidda, S.; Bi, Y.M.; Rothstein, S.J. AMT1;1 transgenic rice plants with enhanced NH4+ permeability show superior growth and higher yield under optimal and suboptimal NH4+ conditions. J. Exp. Bot. 2014, 65, 965–979. [Google Scholar] [CrossRef]
- Zhong, L.; Huang, X.; Zhu, Y.; Kou, E.; Liu, H.; Sun, G.; Chen, R.; Songm, S. Characterization and expression analysis of BcAMT1;4, an ammonium transporter gene in flowering Chinese cabbage. Hortic. Environ. Biotechnol. 2019, 60, 563–572. [Google Scholar] [CrossRef]
- Gansel, X.; Munos, S.; Tillard, P.; Gojon, A. Differential regulation of the NO3- and NH4+ transporter genes AtNrt2.1 and AtAmt1.1 in Arabidopsis: Relation with long-distance and local controls by N status of the plant. Plant J. 2001, 26, 143–155. [Google Scholar] [CrossRef]
- Li, H.; Han, J.; Chang, Y.; Lin, J.; Yang, Q. Gene characterization and transcription analysis of two new ammonium transporters in pear rootstock (Pyrus betulaefolia). J. Plant Res. 2016, 129, 737–748. [Google Scholar] [CrossRef]
- Huang, L.L.; Li, M.J.; Zhou, K.; Sun, T.T.; Hu, L.Y.; Li, C.Y.; Ma, F.W. Uptake and metabolism of ammonium and nitrate in response to drought stress in Malus prunifolia. Plant Physiol. Biochem. 2018, 127, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Hessini, K.; Ben, H.K.; Gandour, M.; Mejri, M.; Abdelly, C.; Cruz, C. Ammonium nutrition in the halophyte Spartina alterniflora under salt stress, evidence for a priming effect of ammonium? Plant Soil 2013, 370, 163–173. [Google Scholar] [CrossRef]
- Yuan, L.; Gu, R.; Xuan, Y.; Smith-Valle, E.; Loqué, D.; Frommer, W.B.; Von Wirén, N. Allosteric regulation of transport activity by heterotrimerization of Arabidopsis ammonium transporter complexes in vivo. Plant Cell 2013, 25, 974–984. [Google Scholar] [CrossRef] [Green Version]
- Ludewig, U.; Neuhäuser, B.; Dynowski, M. Molecular mechanisms of ammonium transport and accumulation in plants. FEBS Lett. 2007, 581, 2301–2308. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.; Li, J.; Zhang, B.; Hao, Y.; Ma, F. Genome-Wide Identification and Expression Analysis of AMT Gene Family in Apple (Malus domestica Borkh.). Horticulturae 2022, 8, 457. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8050457
Huang L, Li J, Zhang B, Hao Y, Ma F. Genome-Wide Identification and Expression Analysis of AMT Gene Family in Apple (Malus domestica Borkh.). Horticulturae. 2022; 8(5):457. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8050457
Chicago/Turabian StyleHuang, Linlin, Jiazhen Li, Bin Zhang, Yanyan Hao, and Fengwang Ma. 2022. "Genome-Wide Identification and Expression Analysis of AMT Gene Family in Apple (Malus domestica Borkh.)" Horticulturae 8, no. 5: 457. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8050457