The Past, Present and Future of Flow Cytometry in Central Nervous System Malignancies



{kind=link}
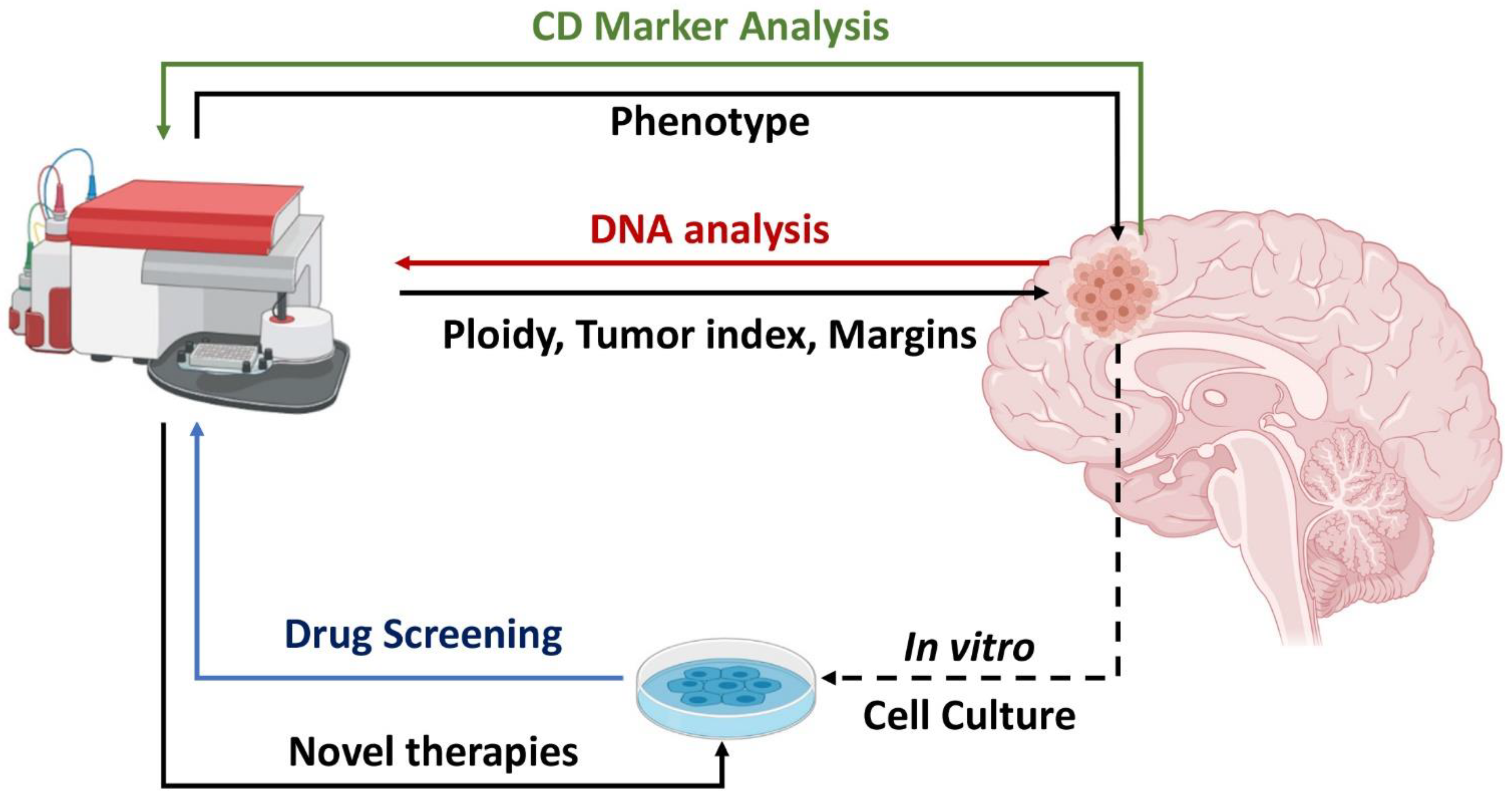
Abstract
:1. Introduction
2. DNA Content Analysis by Flow Cytometry in Brain Malignancies
2.1. History and Early Analysis
2.2. Development of DNA Content Analysis
2.3. Intraoperative Flow Cytometry
3. Phenotypic Analysis
4. Flow Cytometry for Study of Anticancer Agent Efficacy
5. Conclusions
Author Contributions
Funding

Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseau, A.; Mokhtari, K.; Duyckaerts, C. The 2007 WHO classification of tumors of the central nervous system—What has changed? Curr. Opin. Neurol. 2008, 21, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Reifenberger, G.; Von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization classification of tumors of the central nervous system: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Shete, S.; Etzel, C.J.; Scheurer, M.; Alexiou, G.; Armstrong, G.; Tsavachidis, S.; Liang, F.-W.; Gilbert, M.; Aldape, K. Polymorphisms of LIG4, BTBD2, HMGA2, and RTEL1 genes involved in the double-strand break repair pathway predict glioblastoma survival. J. Clin. Oncol. 2010, 28, 2467–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexiou, G.A.; Tsamis, K.I.; Vartholomatos, E.; Peponi, E.; Tzima, E.; Tasiou, I.; Lykoudis, E.; Tsekeris, P.; Kyritsis, A.P. Combination treatment of TRAIL, DFMO and radiation for malignant glioma cells. J. Neuro Oncol. 2015, 123, 217–224. [Google Scholar] [CrossRef]
- Hart, M.G.; Garside, R.; Rogers, G.; Stein, K.; Grant, R. Temozolomide for high grade glioma. Cochrane Libr. 2013. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, Y.; Yu, T.-S.; McKay, R.M.; Burns, D.K.; Kernie, S.G.; Parada, L.F. A restricted cell population propagates glioblastoma growth after chemotherapy. Nature 2012, 488, 522–526. [Google Scholar] [CrossRef] [Green Version]
- Shapiro, H.M. Practical Flow Cytometry; John Wiley & Sons: Hoboken, NJ, USA, 2005. [Google Scholar]
- Spitzer, M.H.; Nolan, G.P. Mass cytometry: Single cells, many features. Cell 2016, 165, 780–791. [Google Scholar] [CrossRef] [Green Version]
- Giesen, C.; Wang, H.A.; Schapiro, D.; Zivanovic, N.; Jacobs, A.; Hattendorf, B.; Schüffler, P.J.; Grolimund, D.; Buhmann, J.M.; Brandt, S. Highly multiplexed imaging of tumor tissues with subcellular resolution by mass cytometry. Nat. Methods 2014, 11, 417–422. [Google Scholar] [CrossRef]
- Nolan, J.P.; Condello, D. Spectral flow cytometry. Curr. Protoc. Cytom. 2013, 63, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Erdbrügger, U.; Rudy, C.K.; Etter, M.E.; Dryden, K.A.; Yeager, M.; Klibanov, A.L.; Lannigan, J. Imaging flow cytometry elucidates limitations of microparticle analysis by conventional flow cytometry. Cytom. Part A 2014, 85, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Barteneva, N.S.; Vorobjev, I.A. Imaging Flow Cytometry; Springer: Berlin, Germany, 2016. [Google Scholar]
- Chang, Q.; Ornatsky, O.I.; Siddiqui, I.; Loboda, A.; Baranov, V.I.; Hedley, D.W. Imaging mass cytometry. Cytom. Part A 2017, 91, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Horan, P.K.; Wheeless, L.L. Quantitative single cell analysis and sorting. Science 1977, 198, 149–157. [Google Scholar] [CrossRef]
- Frederiksen, P.; Reske-Nielsen, E.; Bichel, P. Flow cytometry in tumours of the brain. Acta Neuropathol. 1978, 41, 179–183. [Google Scholar] [CrossRef]
- Kawamoto, K.; Herz, F.; Wolley, R.; Hirano, A.; Kajikawa, H.; Koss, L. Flow cytometric analysis of the DNA distribution in human brain tumors. Acta Neuropathol. 1979, 46, 39–44. [Google Scholar] [CrossRef]
- Hoshino, T.; Nomura, K.; Wilson, C.B.; Knebel, K.D.; Gray, J.W. The distribution of nuclear DNA from human brain-tumor cells: Flow cytometric studies. J. Neurosurg. 1978, 49, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Petersen, S.E.; Frederiksen, P.; Friedrich, U. Cytogenetic analysis and flow cytometric DNA measurement of a human tumor with pronounced hypodiploidy. Cancer Genet. Cytogenet. 1981, 4, 1–9. [Google Scholar] [CrossRef]
- Helson, L.; Traganos, F.; Allen, J.C. Brain tumor cells; flow cytofluorometric analyses in cerebrospinal fluid. N. Y. State J. Med. 1982, 82, 1255–1259. [Google Scholar] [PubMed]
- Nishizaki, T.; Orita, T.; Furutani, Y.; Ikeyama, Y.; Aoki, H.; Sasaki, K. Flow-cytometric DNA analysis and immunohistochemical measurement of Ki-67 and BUdR labeling indices in human brain tumors. J. Neurosurg. 1989, 70, 379–384. [Google Scholar] [CrossRef]
- Danova, M.; Riccardi, A.; Gaetani, P.; Wilson, G.D.; Mazzini, G.; Brugnatelli, S.; Buttini, R.; Butti, G.; Ucci, G.; Paoletti, P. Cell kinetics of human brain tumors: In vivo study with bromodeoxyuridine and flow cytometry. Eur. J. Cancer Clin. Oncol. 1988, 24, 873–880. [Google Scholar] [CrossRef]
- Crone, K.R.; Challa, V.R.; Kute, T.E.; Moody, D.M.; Kelly, D.L., Jr. Relationship between flow cytometric features and clinical behavior of meningiomas. Neurosurgery 1988, 23, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Jay, V.; Squire, J.; Becker, L.E.; Humphreys, R. Malignant transformation in a ganglioglioma with anaplastic neuronal and astrocytic components. Report of a case with flow cytometric and cytogenetic analysis. Cancer 1994, 73, 2862–2868. [Google Scholar] [CrossRef]
- Nishizaki, T.; Ohshita, N.; Nagatsugu, Y.; Orita, T.; Ito, H.; Sasaki, K. Clinical evaluation of DNA index in human brain tumors. J. Neurooncol. 1993, 17, 9–13. [Google Scholar] [CrossRef]
- Kros, J.M.; van Eden, C.G.; Vissers, C.J.; Mulder, A.H.; van der Kwast, T.H. Prognostic relevance of DNA flow cytometry in oligodendroglioma. Cancer 1992, 69, 1791–1798. [Google Scholar] [CrossRef] [Green Version]
- Coons, S.W.; Johnson, P.C.; Haskett, D.; Rider, R. Flow cytometric analysis of deoxyribonucleic acid ploidy and proliferation in choroid plexus tumors. Neurosurgery 1992, 31, 850–856. [Google Scholar] [CrossRef]
- Allam, A.; Taghian, A.; Gioioso, D.; Duffy, M.; Suit, H.D. Intratumoral heterogeneity of malignant gliomas measured in vitro. Int. J. Radiat. Oncol. Biol. Phys. 1993, 27, 303–308. [Google Scholar] [CrossRef]
- Coons, S.W.; Johnson, P.C. Regional heterogeneity in the DNA content of human gliomas. Cancer 1993, 72, 3052–3060. [Google Scholar] [CrossRef]
- Greaves, M.; Maley, C.C. Clonal evolution in cancer. Nature 2012, 481, 306–313. [Google Scholar] [CrossRef]
- Alexiou, G.A.; Vartholomatos, G.; Goussia, A.; Batistatou, A.; Tsamis, K.; Voulgaris, S.; Kyritsis, A.P. Fast cell cycle analysis for intraoperative characterization of brain tumor margins and malignancy. J. Clin. Neurosci. 2015, 22, 129–132. [Google Scholar] [CrossRef]
- Alexiou, G.A.; Vartholomatos, E.; Goussia, A.; Dova, L.; Karamoutsios, A.; Fotakopoulos, G.; Kyritsis, A.P.; Voulgaris, S. DNA content is associated with malignancy of intracranial neoplasms. Clin. Neurol. Neurosurg. 2013, 115, 1784–1787. [Google Scholar] [CrossRef]
- Vartholomatos, G.; Alexiou, G.; Batistatou, A.; Kyritsis, A.P. Intraoperative cell-cycle analysis to guide brain tumor removal. Proc. Natl. Acad. Sci. USA 2014, 111, E3755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santagata, S.; Eberlin, L.S.; Norton, I.; Calligaris, D.; Feldman, D.R.; Ide, J.L.; Liu, X.; Wiley, J.S.; Vestal, M.L.; Ramkissoon, S.H.; et al. Intraoperative mass spectrometry mapping of an onco-metabolite to guide brain tumor surgery. Proc. Natl. Acad. Sci. USA 2014, 111, 11121–11126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexiou, G.A.; Vartholomatos, G.; Voulgaris, S.; Kyritsis, A.P. Letter: Glioblastoma Resection Guided by Flow Cytometry. Neurosurgery 2016, 78, E761. [Google Scholar] [CrossRef] [PubMed]
- Hauser, S.B.; Kockro, R.A.; Actor, B.; Sarnthein, J.; Bernays, R.L. Combining 5-Aminolevulinic Acid Fluorescence and Intraoperative Magnetic Resonance Imaging in Glioblastoma Surgery: A Histology-Based Evaluation. Neurosurgery 2016, 78, 475–483. [Google Scholar] [CrossRef] [Green Version]
- Kockro, R.A.; Hauser, S.B.; Bernays, R.L. In Reply: Glioblastoma Resection Guided by Flow Cytometry. Neurosurgery 2016, 78, E761–E762. [Google Scholar] [CrossRef]
- Alexiou, G.A.; Vartholomatos, G.; Stefanaki, K.; Markopoulos, G.S.; Kyritsis, A.P. Intraoperative Flow Cytometry for Diagnosis of Central Nervous System Lesions. J. Cytol. 2019, 36, 134–135. [Google Scholar] [CrossRef]
- Jindal, A.; Kaur, K.; Mathur, K.; Kumari, V.; Diwan, H. Intraoperative Squash Smear Cytology in CNS Lesions: A Study of 150 Pediatric Cases. J. Cytol. 2017, 34, 217–220. [Google Scholar] [CrossRef]
- Shioyama, T.; Muragaki, Y.; Maruyama, T.; Komori, T.; Iseki, H. Intraoperative flow cytometry analysis of glioma tissue for rapid determination of tumor presence and its histopathological grade. J. Neurosurg. 2013, 118, 1232–1238. [Google Scholar] [CrossRef] [Green Version]
- Alexiou, G.A.; Vartholomatos, G.; Kobayashi, T.; Voulgaris, S.; Kyritsis, A.P. The emerging role of intraoperative flow cytometry in intracranial tumor surgery. Clin. Neurol. Neurosurg. 2020, 192, 105742. [Google Scholar] [CrossRef]
- Saito, T.; Muragaki, Y.; Shioyama, T.; Komori, T.; Maruyama, T.; Nitta, M.; Yasuda, T.; Hosono, J.; Okamoto, S.; Kawamata, T. Malignancy index using intraoperative flow cytometry is a valuable prognostic factor for glioblastoma treated with radiotherapy and concomitant temozolomide. Neurosurgery 2019, 84, 662–672. [Google Scholar] [CrossRef]
- Vartholomatos, G.; Alexiou, G.A.; Batistatou, A.; Lykoudis, E.; Voulgaris, S.; Kyritsis, A.P. GV/GA Sarissa-Lancet: A Proposed Real-Time Flow Cytometer for Intraoperative Identification of Glioma Margins. Surg. Innov. 2016, 23, 104–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vartholomatos, G.; Alexiou, G.A.; Lianos, G.D.; Kyritsis, A.P. From bench to operating theater: Has the time come for a molecular scalpel? Future Oncol. 2017, 13, 121–123. [Google Scholar] [CrossRef] [PubMed]
- Vartholomatos, G.; Alexiou, G.A.; Lianos, G.D.; Harissis, H.; Voulgaris, S.; Kyritsis, A.P. Intraoperative cell cycle analysis for tumor margins evaluation: The future is now? Int. J. Surg. 2018, 53, 380–381. [Google Scholar] [CrossRef] [PubMed]
- Vartholomatos, G.; Basiari, L.; Exarchakos, G.; Kastanioudakis, I.; Komnos, I.; Michali, M.; Markopoulos, G.S.; Batistatou, A.; Papoudou-Bai, A.; Alexiou, G.A. Intraoperative flow cytometry for head and neck lesions. Assessment of malignancy and tumour-free resection margins. Oral Oncol. 2019, 99, 104344. [Google Scholar] [CrossRef] [PubMed]
- Vartholomatos, G.; Basiari, L.; Kastanioudakis, I.; Psichogios, G.; Alexiou, G.A. The Role of Intraoperative Flow Cytometry in Surgical Margins of Head and Neck Malignancies. Ear Nose Throat J. 2020. [Google Scholar] [CrossRef]
- Andreou, M.; Vartholomatos, E.; Harissis, H.; Markopoulos, G.S.; Alexiou, G.A. Past, Present and Future of Flow Cytometry in Breast Cancer—A Systematic Review. EJIFCC 2019, 30, 423–437. [Google Scholar]
- Koriyama, S.; Nitta, M.; Shioyama, T.; Komori, T.; Maruyama, T.; Kawamata, T.; Muragaki, Y. Intraoperative Flow Cytometry Enables the Differentiation of Primary Central Nervous System Lymphoma from Glioblastoma. World Neurosurg. 2018, 112, e261–e268. [Google Scholar] [CrossRef]
- Vartholomatos, G.; Alexiou, G.A.; Voulgaris, S.; Kyritsis, A.P. Intraoperative Immunophenotypic Analysis for Diagnosis and Classification of Primary Central Nervous System Lymphomas. World Neurosurg. 2018, 117, 464–465. [Google Scholar] [CrossRef]
- Vartholomatos, G.; Stefanaki, K.; Alexiou, G.A.; Batistatou, A.; Markopoulos, G.S.; Tzoufi, M.; Sfakianos, G.; Prodromou, N. Pediatric Brain Tumor Grading Based on CD56 Quantification. J. Pediatr. Neurosci. 2018, 13, 524–527. [Google Scholar] [CrossRef]
- Vartholomatos, G.; Alexiou, G.A.; Stefanaki, K.; Lykoudis, E.G.; Tseka, G.; Tzoufi, M.; Sfakianos, G.; Prodromou, N. The value of cell cycle analysis by propidium-iodine staining of CD56+ cells in pediatric brain tumors. Clin. Neurol. Neurosurg. 2015, 133, 70–74. [Google Scholar] [CrossRef]
- Craig, F.E.; Foon, K.A. Flow cytometric immunophenotyping for hematologic neoplasms. Blood 2008, 111, 3941–3967. [Google Scholar] [CrossRef] [PubMed]
- Moors, I.; Vandepoele, K.; Philippé, J.; Deeren, D.; Selleslag, D.; Breems, D.; Straetmans, N.; Kerre, T.; Denys, B. Clinical implications of measurable residual disease in AML: Review of current evidence. Crit. Rev. Oncol. Hematol. 2019, 133, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Fuda, F.; Chen, W. Minimal/Measurable Residual Disease Detection in Acute Leukemias by Multiparameter Flow Cytometry. Curr. Hematol. Malig. Rep. 2018, 13, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Schuurhuis, G.J.; Heuser, M.; Freeman, S.; Béné, M.-C.; Buccisano, F.; Cloos, J.; Grimwade, D.; Haferlach, T.; Hills, R.K.; Hourigan, C.S. Minimal/measurable residual disease in AML: A consensus document from the European LeukemiaNet MRD Working Party. Blood 2018, 131, 1275–1291. [Google Scholar] [CrossRef] [Green Version]
- Engel, P.; Boumsell, L.; Balderas, R.; Bensussan, A.; Gattei, V.; Horejsi, V.; Jin, B.-Q.; Malavasi, F.; Mortari, F.; Schwartz-Albiez, R. CD nomenclature 2015: Human leukocyte differentiation antigen workshops as a driving force in immunology. J. Immunol. 2015, 195, 4555–4563. [Google Scholar] [CrossRef] [Green Version]
- Sheridan, C.; Kishimoto, H.; Fuchs, R.K.; Mehrotra, S.; Bhat-Nakshatri, P.; Turner, C.H.; Goulet, R.; Badve, S.; Nakshatri, H. CD44+/CD24-breast cancer cells exhibit enhanced invasive properties: An early step necessary for metastasis. Breast Cancer Res. 2006, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Mylona, E.; Giannopoulou, I.; Fasomytakis, E.; Nomikos, A.; Magkou, C.; Bakarakos, P.; Nakopoulou, L. The clinicopathologic and prognostic significance of CD44+/CD24−/low and CD44−/CD24+ tumor cells in invasive breast carcinomas. Hum. Pathol. 2008, 39, 1096–1102. [Google Scholar] [CrossRef]
- Beier, D.; Hau, P.; Proescholdt, M.; Lohmeier, A.; Wischhusen, J.; Oefner, P.J.; Aigner, L.; Brawanski, A.; Bogdahn, U.; Beier, C.P. CD133+ and CD133− glioblastoma-derived cancer stem cells show differential growth characteristics and molecular profiles. Cancer Res. 2007, 67, 4010–4015. [Google Scholar] [CrossRef] [Green Version]
- Greve, B.; Kelsch, R.; Spaniol, K.; Eich, H.T.; Götte, M. Flow cytometry in cancer stem cell analysis and separation. Cytom. Part A 2012, 81, 284–293. [Google Scholar] [CrossRef]
- Mizrak, D.; Brittan, M.; Alison, M. CD133: Molecule of the moment. J. Pathol. A J. Pathol. Soc. Great Br. Irel. 2008, 214, 3–9. [Google Scholar] [CrossRef]
- Singh, S.K.; Clarke, I.D.; Terasaki, M.; Bonn, V.E.; Hawkins, C.; Squire, J.; Dirks, P.B. Identification of a cancer stem cell in human brain tumors. Cancer Res. 2003, 63, 5821–5828. [Google Scholar] [PubMed]
- Hemmati, H.D.; Nakano, I.; Lazareff, J.A.; Masterman-Smith, M.; Geschwind, D.H.; Bronner-Fraser, M.; Kornblum, H.I. Cancerous stem cells can arise from pediatric brain tumors. Proc. Natl. Acad. Sci. USA 2003, 100, 15178–15183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lottaz, C.; Beier, D.; Meyer, K.; Kumar, P.; Hermann, A.; Schwarz, J.; Junker, M.; Oefner, P.J.; Bogdahn, U.; Wischhusen, J. Transcriptional profiles of CD133+ and CD133− glioblastoma-derived cancer stem cell lines suggest different cells of origin. Cancer Res. 2010, 70, 2030–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, K.M.; Kim, S.Y.; Jin, X.; Song, S.Y.; Kong, D.-S.; Lee, J.-I.; Jeon, J.W.; Kim, M.H.; Kang, B.G.; Jung, Y. Clinical and biological implications of CD133-positive and CD133-negative cells in glioblastomas. Lab. Investig. 2008, 88, 808–815. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Khaki, L.; Zhu, T.S.; Soules, M.E.; Talsma, C.E.; Gul, N.; Koh, C.; Zhang, J.; Li, Y.M.; Maciaczyk, J. NOTCH pathway blockade depletes CD133-positive glioblastoma cells and inhibits growth of tumor neurospheres and xenografts. Stem Cells 2010, 28, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Arber, D.; Weiss, L. CD15: A review. Appl. Immunohistochem. 1993, 1, 17–30. [Google Scholar]
- Reifenberger, G.; Sieth, P.; Niederhaus, M.; Wechsler, W. Expression of CD15 in tumours of the nervous system. Histochem. J. 1992, 24, 890–901. [Google Scholar] [CrossRef]
- Son, M.J.; Woolard, K.; Nam, D.H.; Lee, J.; Fine, H.A. SSEA-1 is an enrichment marker for tumor-initiating cells in human glioblastoma. Cell Stem Cell 2009, 4, 440–452. [Google Scholar] [CrossRef] [Green Version]
- Pruszak, J.; Ludwig, W.; Blak, A.; Alavian, K.; Isacson, O. CD15, CD24, and CD29 define a surface biomarker code for neural lineage differentiation of stem cells. Stem Cells 2009, 27, 2928–2940. [Google Scholar] [CrossRef] [Green Version]
- Panchision, D.M.; Chen, H.L.; Pistollato, F.; Papini, D.; Ni, H.T.; Hawley, T.S. Optimized flow cytometric analysis of central nervous system tissue reveals novel functional relationships among cells expressing CD133, CD15, and CD24. Stem Cells 2007, 25, 1560–1570. [Google Scholar] [CrossRef] [Green Version]
- Mechtersheimer, G.; Staudter, M.; Möller, P. Expression of the natural killer cell-associated antigens CD56 and CD57 in human neural and striated muscle cells and in their tumors. Cancer Res. 1991, 51, 1300–1307. [Google Scholar]
- Leelatian, N.; Doxie, D.B.; Greenplate, A.R.; Mobley, B.C.; Lehman, J.M.; Sinnaeve, J.; Kauffmann, R.M.; Werkhaven, J.A.; Mistry, A.M.; Weaver, K.D.; et al. Single cell analysis of human tissues and solid tumors with mass cytometry. Cytom. Part B Clin. Cytom. 2017, 92, 68–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasquez, J.C.; Huttner, A.; Zhang, L.; Marks, A.; Chan, A.; Baehring, J.M.; Kahle, K.T.; Dhodapkar, K.M. SOX2 immunity and tissue resident memory in children and young adults with glioma. J. Neurooncol. 2017, 134, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Hu, A.X.; Adams, J.J.; Vora, P.; Qazi, M.; Singh, S.K.; Moffat, J.; Sidhu, S.S. EPH Profiling of BTIC Populations in Glioblastoma Multiforme Using CyTOF. Methods Mol. Biol. 2019, 1869, 155–168. [Google Scholar] [CrossRef]
- Sankowski, R.; Bottcher, C.; Masuda, T.; Geirsdottir, L.; Sindram, E.; Seredenina, T.; Muhs, A.; Scheiwe, C.; Shah, M.J.; Heiland, D.H.; et al. Mapping microglia states in the human brain through the integration of high-dimensional techniques. Nat. Neurosci. 2019, 22, 2098–2110. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Wang, W.; Li, H.; Jiao, Y.; Weng, J.; Huo, R.; Yan, Z.; Wang, J.; Xu, H.; Wang, S.; et al. High Dimensional Mass Cytometry Analysis Reveals Characteristics of the Immunosuppressive Microenvironment in Diffuse Astrocytomas. Front. Oncol. 2020, 10, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alban, T.J.; Alvarado, A.G.; Sorensen, M.D.; Bayik, D.; Volovetz, J.; Serbinowski, E.; Mulkearns-Hubert, E.E.; Sinyuk, M.; Hale, J.S.; Onzi, G.R.; et al. Global immune fingerprinting in glioblastoma patient peripheral blood reveals immune-suppression signatures associated with prognosis. JCI Insight 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dusoswa, S.A.; Verhoeff, J.; Garcia-Vallejo, J.J. OMIP-054: Broad Immune Phenotyping of Innate and Adaptive Leukocytes in the Brain, Spleen, and Bone Marrow of an Orthotopic Murine Glioblastoma Model by Mass Cytometry. Cytom. A 2019, 95, 422–426. [Google Scholar] [CrossRef]
- Friebel, E.; Kapolou, K.; Unger, S.; Nunez, N.G.; Utz, S.; Rushing, E.J.; Regli, L.; Weller, M.; Greter, M.; Tugues, S.; et al. Single-Cell Mapping of Human Brain Cancer Reveals Tumor-Specific Instruction of Tissue-Invading Leukocytes. Cell 2020, 181, 1626–1642. [Google Scholar] [CrossRef]
- Robinson, M.H.; Vasquez, J.; Kaushal, A.; MacDonald, T.J.; Velazquez Vega, J.E.; Schniederjan, M.; Dhodapkar, K. Subtype and grade-dependent spatial heterogeneity of T-cell infiltration in pediatric glioma. J. Immunother. Cancer 2020, 8. [Google Scholar] [CrossRef]
- Tsamis, K.I.; Alexiou, G.A.; Vartholomatos, E.; Kyritsis, A.P. Combination treatment for glioblastoma cells with tumor necrosis factor-related apoptosis-inducing ligand and oncolytic adenovirus delta-24. Cancer Investig. 2013, 31, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, M.; Barthel, F.P.; Malta, T.M.; Sabedot, T.S.; Salama, S.R.; Murray, B.A.; Morozova, O.; Newton, Y.; Radenbaugh, A.; Pagnotta, S.M. Molecular profiling reveals biologically discrete subsets and pathways of progression in diffuse glioma. Cell 2016, 164, 550–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisapia, D.J. The updated World Health Organization glioma classification: Cellular and molecular origins of adult infiltrating gliomas. Arch. Pathol. Lab. Med. 2017, 141, 1633–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zacher, A.; Kaulich, K.; Stepanow, S.; Wolter, M.; Köhrer, K.; Felsberg, J.; Malzkorn, B.; Reifenberger, G. Molecular diagnostics of Gliomas using next generation sequencing of a Glioma-tailored gene panel. Brain Pathol. 2017, 27, 146–159. [Google Scholar] [CrossRef] [PubMed]
- Karsy, M.; Gelbman, M.; Shah, P.; Balumbu, O.; Moy, F.; Arslan, E. Established and emerging variants of glioblastoma multiforme: Review of morphological and molecular features. Folia Neuropathol. 2012, 50, 301–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondy, M.; Wiencke, J.; Wrensch, M.; Kyritsis, A.P. Genetics of primary brain tumors: A review. J. Neuro Oncol. 1994, 18, 69–81. [Google Scholar] [CrossRef]
- Arita, H.; Yamasaki, K.; Matsushita, Y.; Nakamura, T.; Shimokawa, A.; Takami, H.; Tanaka, S.; Mukasa, A.; Shirahata, M.; Shimizu, S. A combination of TERT promoter mutation and MGMT methylation status predicts clinically relevant subgroups of newly diagnosed glioblastomas. Acta Neuropathol. Commun. 2016, 4, 79. [Google Scholar] [CrossRef] [Green Version]
- Neftel, C.; Laffy, J.; Filbin, M.G.; Hara, T.; Shore, M.E.; Rahme, G.J.; Richman, A.R.; Silverbush, D.; Shaw, M.L.; Hebert, C.M. An integrative model of cellular states, plasticity, and genetics for glioblastoma. Cell 2019, 178, 835–849. [Google Scholar] [CrossRef]
- Stupp, R.; Mason, W.P.; Van Den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef]
- Hegi, M.E.; Diserens, A.-C.; Gorlia, T.; Hamou, M.-F.; De Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L. MGMT gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar] [CrossRef] [Green Version]
- Van Nifterik, K.A.; Van den Berg, J.; Slotman, B.J.; Lafleur, M.V.M.; Sminia, P.; Stalpers, L.J. Valproic acid sensitizes human glioma cells for temozolomide and γ-radiation. J. Neuro Oncol. 2012, 107, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.; Hassiotou, F.; Nowak, A. Glioblastoma stem-like cells: At the root of tumor recurrence and a therapeutic target. Carcinogenesis 2015, 36, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Gao, G.; Wang, L.; Wang, T.; Yu, J.; Zhao, Z. CD24 expression as a marker for predicting clinical outcome in human gliomas. Biomed. Res. Int. 2012. [Google Scholar] [CrossRef] [PubMed]
- Alexiou, G.A.; Vartholomatos, E.; Tsamis, K.I.; Peponi, E.; Markopoulos, G.; Papathanasopoulou, V.A.; Tasiou, I.; Ragos, V.; Tsekeris, P.; Kyritsis, A.P. Combination treatment for glioblastoma with temozolomide, DFMO and radiation. J. BUON 2019, 24, 397–404. [Google Scholar]
- Alexiou, G.A.; Lazari, D.; Markopoulos, G.; Vartholomatos, E.; Hodaj, E.; Galani, V.; Kyritsis, A.P. Moschamine inhibits proliferation of glioblastoma cells via cell cycle arrest and apoptosis. Tumor Biol. 2017, 39, 1010428317705744. [Google Scholar] [CrossRef] [Green Version]
- Lazari, D.; Alexiou, G.A.; Markopoulos, G.S.; Vartholomatos, E.; Hodaj, E.; Chousidis, I.; Leonardos, I.; Galani, V.; Kyritsis, A.P. N-(p-coumaroyl) serotonin inhibits glioblastoma cells growth through triggering S-phase arrest and apoptosis. J. Neuro Oncol. 2017, 132, 373–381. [Google Scholar] [CrossRef]
- Vartholomatos, E.; Alexiou, G.; Markopoulos, G.; Lazari, D.; Tsiftsoglou, O.; Chousidis, I.; Leonardos, I.; Kyritsis, A. Deglucohellebrin. A Potent Agent for Glioblastoma Treatment. Anti Cancer Agents Med. Chem. 2019. [Google Scholar] [CrossRef]
- Su, C.-C.; Wang, M.-J.; Chiu, T.-L. The anti-cancer efficacy of curcumin scrutinized through core signaling pathways in glioblastoma. Int. J. Mol. Med. 2010, 26, 217–224. [Google Scholar]
- Pozsgai, E.; Bellyei, S.; Cseh, A.; Boronkai, A.; Racz, B.; Szabo, A.; Sumegi, B.; Hocsak, E. Quercetin increases the efficacy of glioblastoma treatment compared to standard chemoradiotherapy by the suppression of PI-3-kinase-Akt pathway. Nutr. Cancer 2013, 65, 1059–1066. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhu, J.-H.; Cao, L.-P.; Sun, Q.; Liu, H.-D.; Li, J.-S.; Hang, C.-H. Growth inhibitory in vitro effects of glycyrrhizic acid in U251 glioblastoma cell line. Neurol. Sci. 2014, 35, 1115–1120. [Google Scholar] [CrossRef]
- Whittaker, S.; Madani, D.; Joshi, S.; Chung, S.A.; Johns, T.; Day, B.; Khasraw, M.; McDonald, K.L. Combination of palbociclib and radiotherapy for glioblastoma. Cell Death Discov. 2017, 3, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, F.; Isihou, R.; Alexiou, G.A.; Tsalios, T.; Vartholomatos, E.; Markopoulos, G.S.; Sioka, C.; Tsekeris, P.; Kyritsis, A.P.; Galani, V. Haloperidol Induced Cell Cycle Arrest and Apoptosis in Glioblastoma Cells. Biomedicines 2020, 8, 595. [Google Scholar] [CrossRef] [PubMed]
- Omoruyi, S.; Ekpo, O.; Semenya, D.; Jardine, A.; Prince, S. Exploitation of a novel phenothiazine derivative for its anti-cancer activities in malignant glioblastoma. Apoptosis 2020, 25, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kondo, N.; Hikida, M.; Nakada, M.; Sakurai, Y.; Hirata, E.; Takeno, S.; Suzuki, M. Glioma Stem-Like Cells Can Be Targeted in Boron Neutron Capture Therapy with Boronophenylalanine. Cancers 2020, 12, 3040. [Google Scholar] [CrossRef]
- Gudbergsson, J.M.; Christensen, E.; Kostrikov, S.; Moos, T.; Duroux, M.; Kjær, A.; Johnsen, K.B.; Andresen, T.L. Conventional treatment of glioblastoma reveals persistent CD44+ subpopulations. Mol. Neurobiol. 2020, 57, 3943–3955. [Google Scholar] [CrossRef]
- Erhart, F.; Blauensteiner, B.; Zirkovits, G.; Printz, D.; Soukup, K.; Klingenbrunner, S.; Fischhuber, K.; Reitermaier, R.; Halfmann, A.; Lötsch, D. Gliomasphere marker combinatorics: Multidimensional flow cytometry detects CD 44+/CD 133+/ITGA 6+/CD 36+ signature. J. Cell. Mol. Med. 2019, 23, 281–292. [Google Scholar] [CrossRef]
- Mostafa, H.; Pala, A.; Högel, J.; Hlavac, M.; Dietrich, E.; Westhoff, M.A.; Nonnenmacher, L.; Burster, T.; Georgieff, M.; Wirtz, C.R. Immune phenotypes predict survival in patients with glioblastoma multiforme. J. Hematol. Oncol. 2016, 9, 77. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vartholomatos, E.; Vartholomatos, G.; Alexiou, G.A.; Markopoulos, G.S. The Past, Present and Future of Flow Cytometry in Central Nervous System Malignancies. Methods Protoc. 2021, 4, 11. https://0-doi-org.brum.beds.ac.uk/10.3390/mps4010011
Vartholomatos E, Vartholomatos G, Alexiou GA, Markopoulos GS. The Past, Present and Future of Flow Cytometry in Central Nervous System Malignancies. Methods and Protocols. 2021; 4(1):11. https://0-doi-org.brum.beds.ac.uk/10.3390/mps4010011
Chicago/Turabian StyleVartholomatos, Evrysthenis, George Vartholomatos, George A. Alexiou, and Georgios S. Markopoulos. 2021. "The Past, Present and Future of Flow Cytometry in Central Nervous System Malignancies" Methods and Protocols 4, no. 1: 11. https://0-doi-org.brum.beds.ac.uk/10.3390/mps4010011