
Assessment of the in Vitro Antithrombotic Properties of Sardine (Sardina pilchardus) Fillet Lipids and Cod Liver Oil

 ,
,
Abstract
:
1. Introduction
2. Results and Discussion
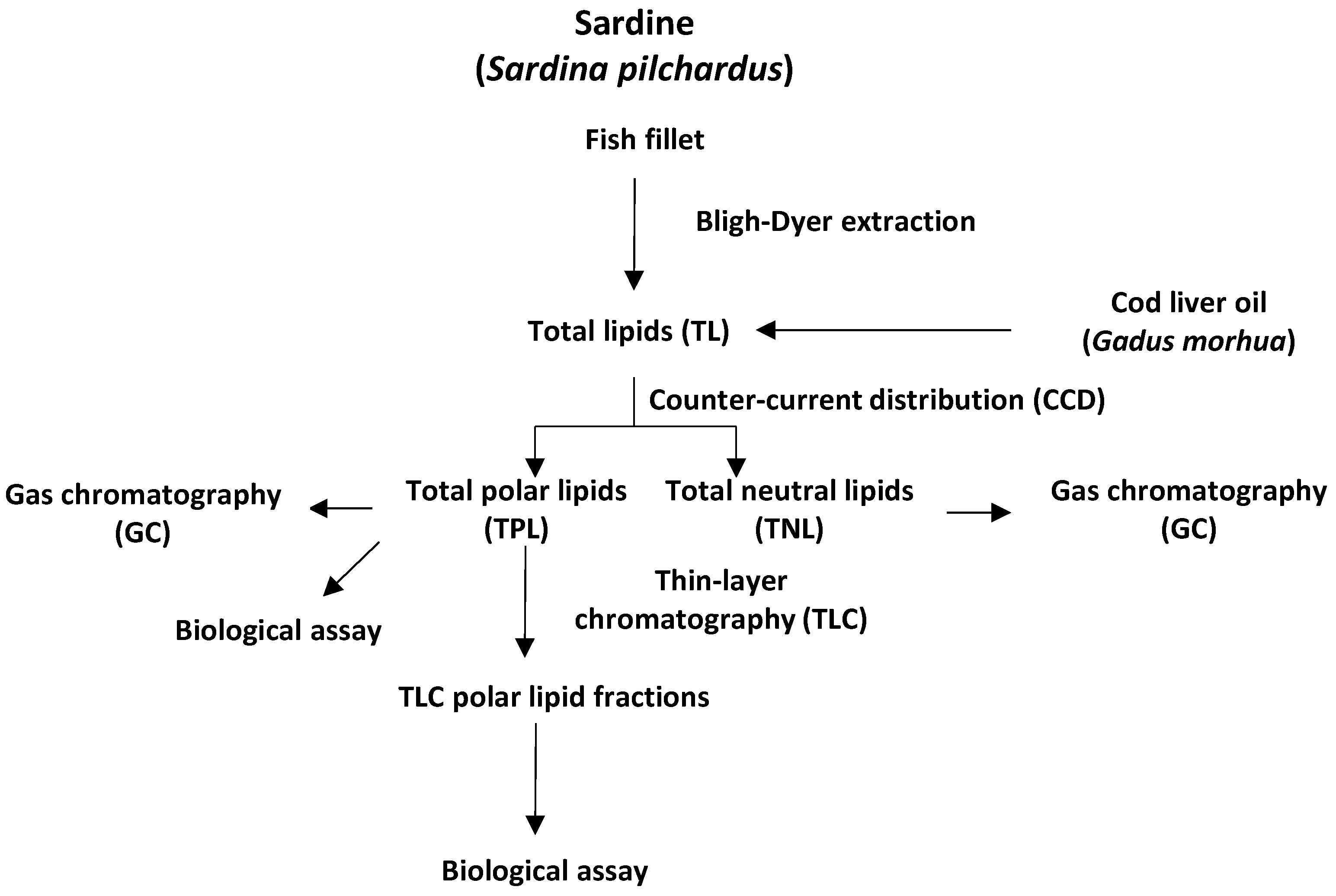
2.1. Total Lipids (TL), Total Polar Lipids (TPL) and Total Neutral Lipids (TNL) of Sardine (Sardina pilchardus) Fillet Lipids and Cod Liver Oil


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | TPL (g/100 g Fish Lipids/Oil) | TNL (g/100 g Fish Lipids/Oil) |
|---|---|---|
| Sardine fillet lipids | 57.7 ± 1.27 a,* | 31.2 ± 0.65 a,** |
| Cod liver oil | 1.00 ± 0.03 b,* | 93.6 ± 2.81 b,** |
2.2. Fatty Acid Profile of TPL and TNL of Sardine Fillet and Cod Liver
| Fatty Acid | Sardine Fillet Lipid (mg/kg of Fish Lipid) | Cod Liver Oil (mg/kg of Fish Oil) |
|---|---|---|
| 14:0 | n.d. | 33.3 ± 0.99 |
| 16:0 | 6011 ± 1421 a | 69.0 ± 0.80 b |
| 16:1 (ω-7) | n.d. | 53.3 ± 0.87 |
| 18:0 | 927 ± 4.72 a | 12.8 ± 0.07 b |
| 18:1 cis (ω-9) | 778 ± 223 a | 69.9 ± 0.25 b |
| 18:1 trans (ω-9) | 60.3 ± 84.7 a | 8.48 ± 1.37 b |
| 18:2 (ω-6) | n.d. | 11.5 ± 0.61 |
| 18:3 (ω-3) | n.d. | 7.16 ± 0.72 |
| 20:4 (ω-6) | 352 ± 6.30 a | 1.46 ± 0.05 b |
| 20:5 (ω-3) | 1527 ± 19.9 a | 71.3 ± 0.44 b |
| 22:5 (ω-3) | 75.8 ± 0.74 a | 5.03 ± 0.23 b |
| 22:6 (ω-3) | 14,077 ± 120 a | 64.7 ± 1.94 b |
| Total ω-3 | 15,680 ± 141 a | 148 ± 3.32 b |
| Total ω-6 | 352 ± 6.30 a | 13.0 ± 0.66 b |
| Total ω-7 | n.d. | 53.3 ± 0.87 |
| Total ω-9 | 838 ± 308 a | 78.4 ± 1.62 b |
| Total SFA | 6938 ± 6.14 a | 115 ± 1.86 b |
| Total MUFA | 838 ± 308 a | 132 ± 2.49 b |
| ω-6/ω-3 | 0.022 | 0.087 |
| Fatty Acid | Sardine Fillet Lipid (mg/kg of Fish Lipid) | Cod Liver Oil (mg/kg of Fish Oil) |
|---|---|---|
| 14:0 | 16.8 ± 5.43 a | 123 ± 24.7 b |
| 16:0 | 843 ± 12.3 a | 635 ± 16.5 b |
| 16:1 (ω-7) | 64.0 ± 3.09 a | 439 ± 94.7 b |
| 18:0 | 338 ± 9.72 | n.d. |
| 18:1 cis (ω-9) | 640 ± 8.74 a | 1000 ± 33.1 b |
| 18:1 trans (ω-9) | 94.9 ± 10.7 | n.d. |
| 18:2 (ω-6) | 62.8 ± 45.2 | n.d. |
| 18:3 (ω-3) | n.d. | n.d. |
| 20:4 (ω-6) | n.d. | n.d. |
| 20:5 (ω-3) | 161 ± 17.0 a | 577 ± 26.8 b |
| 22:5 (ω-3) | n.d. | n.d. |
| 22:6 (ω-3) | 1484 ± 224 a | 728 ± 40.9 b |
| Total ω-3 | 1645 ± 241 a | 1304 ± 67.7 b |
| Total ω-6 | 62.8 ± 45.2 | n.d. |
| Total ω-7 | 64.0 ± 3.09 a | 439 ± 94.7 b |
| Total ω-9 | 735 ± 19.5 a | 1000 ± 33.1 b |
| Total SFA | 1198 ± 27.43 a | 7578 ± 41.2 b |
| Total MUFA | 799 ± 22.55 a | 1439 ± 128 b |
| ω-6/ω-3 | 0.04 | 0.00 |


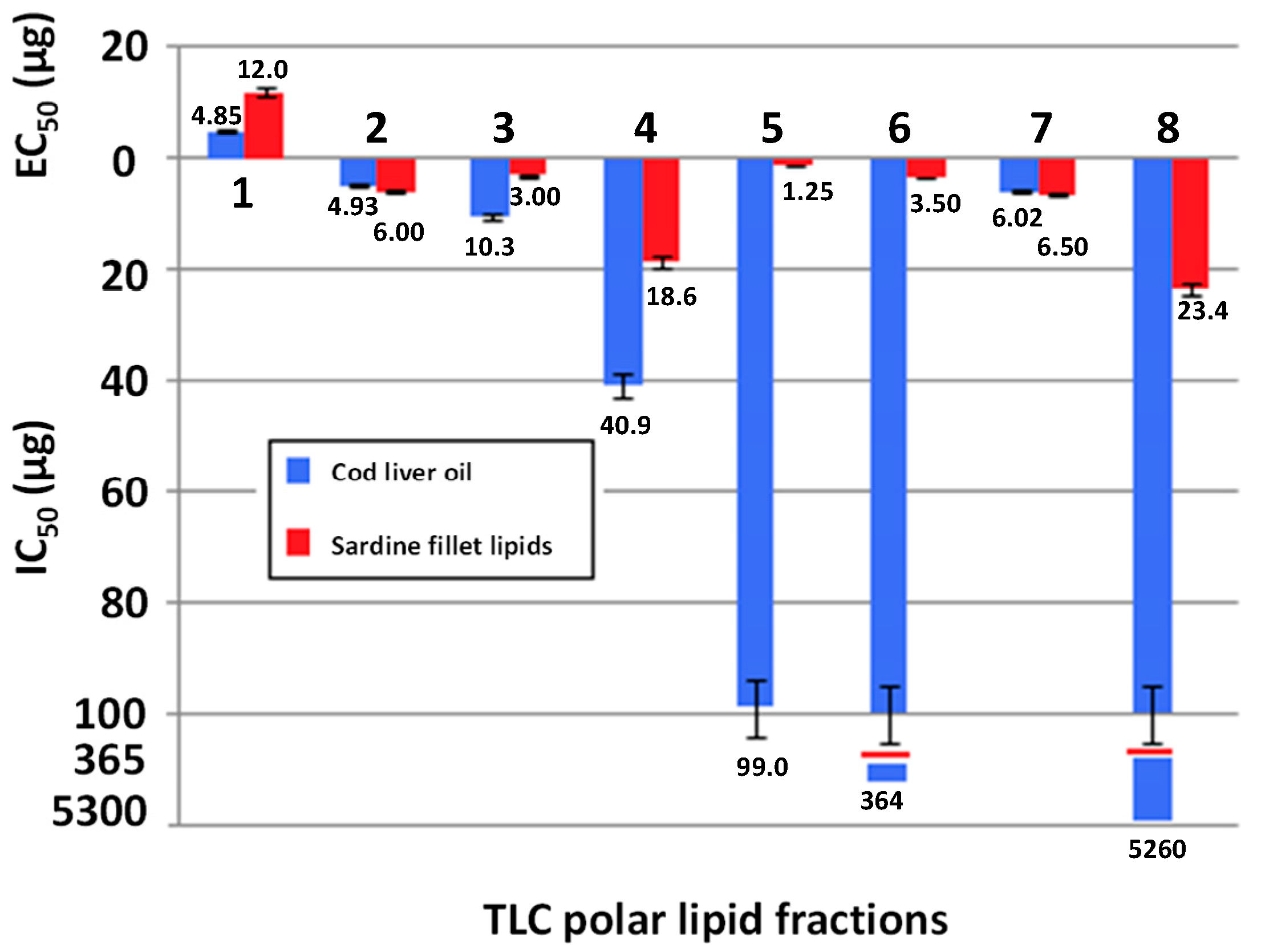
2.3. Biological Activity of Sardine Fillet and Cod Liver TPL
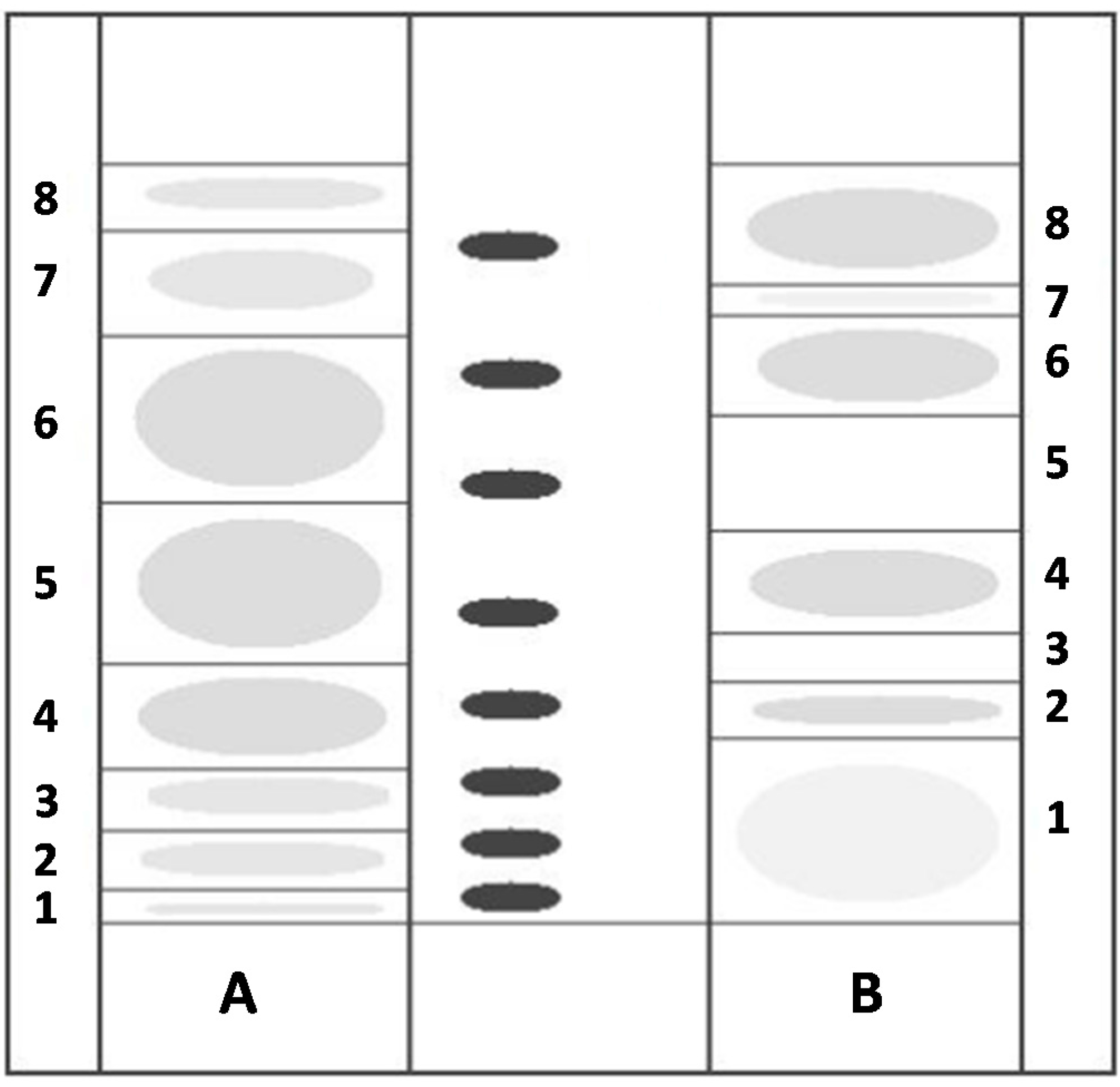
2.4. TLC Separation of Polar Lipids and Biological Activity of Sardine Fillet and Cod Liver TLC Polar Lipid Fractions


3. Experimental Section
3.1. Reagents
3.2. Sardine (Sardina pilchardus) and Cod Liver Oil Sampling
3.3. Isolation of Fish Total Lipids of Sardine (Sardina pilchardus) and Cod Liver Oil
3.4. Gas Chromatography Analysis
3.5. Fractionation of TPL by TLC
3.6. Biological Assay of the in Vitro Antithrombotic Properties
3.7. Desensitization Experiment
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Conflicts of Interest
References
- Kris-Etherton, P.M.; Harris, W.S.; Appel, L.J. Fish consumption, fish oil, ω-3 fatty acids and cardiovascular disease. Arterioscler. Thromb. Vasc. Boil. 2003, 23, 20–31. [Google Scholar] [CrossRef]
- Din, J.N.; Newby, D.E.; Flapan, A.D. ω-3 Fatty acids and cardiovascular disease-fishing for a natural treatment. Br. Med. J. 2004, 328, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, S.D.; Iversen, A.M.; Schmidt, E.B. n-3 Polyunsaturated fatty acids and coronary thrombosis. Lipids 2001, 36, S79–S82. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.A.; Beilin, L.J.; Burke, V.; Morris, J.; Ritchie, J. Interactions between dietary fat, fish, and fish oil and their effects on platelet function in men at risk of cardiovascular disease. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Rementzis, J.; Antonopoulou, S.; Demopoulos, C.A. Identification and study of gangliosides from Scomber scombrus muscles. J. Agric. Food Chem. 1997, 45, 611–615. [Google Scholar] [CrossRef]
- Panayiotou, A.; Samartzis, D.; Nomikos, T.; Fragopoulou, E.; Karantonis, H.C.; Demopoulos, C.A.; Demopoulos, C.A.; Zabetakis, I. Lipid fractions with aggregatory and antiaggregatory activity toward platelets in fresh and fried cod (Gadus morhua): Correlation with platelet-activating factor and atherogenesis. J. Agric. Food Chem. 2000, 48, 6372–6379. [Google Scholar] [CrossRef] [PubMed]
- Nomikos, T.; Karantonis, H.C.; Skarvelis, C.; Demopoulos, C.A.; Zabetakis, I. Antiatherogenic properties of lipid fractions of raw and fried fish. Food Chem. 2006, 96, 29–35. [Google Scholar] [CrossRef]
- Nasopoulou, C.; Nomikos, T.; Demopoulos, C.A.; Zabetakis, I. Comparison of antiatherogenic properties of lipids obtained from wild and cultured sea bass (Dicentrarchus labrax) and gilthead sea bream (Sparus aurata). Food Chem. 2007, 100, 560–567. [Google Scholar] [CrossRef]
- Nasopoulou, C.; Karantonis, H.C.; Perrea, D.N.; Theocharis, S.E.; Iliopoulos, D.G.; Demopoulos, C.A.; Zabetakis, I. In vivo anti-atherogenic properties of cultured gilthead sea bream (Sparus aurata) polar lipid extracts in hypercholesterolaemic rabbits. Food Chem. 2010, 120, 831–836. [Google Scholar] [CrossRef]
- Roger, V.L.; Go, A.S.; Lloyd-Jones, D.M.; Benjamin, E.J.; Berry, J.D.; Borden, W.B.; Bravata, D.M.; Dai, S.; Ford, E.S.; Fox, C.S.; et al. Executive Summary: Heart disease and stroke statistics—2012 update. Circulation 2012, 125, 188–197. [Google Scholar] [PubMed]
- Demopoulos, C.A.; Pinckard, R.N.; Hanahan, D.J. Platelet-activating factor. Evidence for 1-O-alkyl-2-acetyl-sn-glyceryl-3-phosphoryl-choline as the active component (a new class of lipid chemical mediators). J. Biol. Chem. 1979, 254, 9355–9358. [Google Scholar] [PubMed]
- Demopoulos, C.A.; Karantonis, H.C.; Antonopoulou, S. Platelet activating factor—A molecular link between atherosclerosis theories. Eur. J. Lipid Sci. Technol. 2003, 105, 705–716. [Google Scholar] [CrossRef]
- Nasopoulou, C.; Smith, T.; Detopoulou, M.; Tsikrika, C.; Papaharisis, L.; Barkas, D.; Zabetakis, I. Structural elucidation of olive pomace fed sea bass (Dicentrarchus labrax) polar lipids with cardioprotective activities. Food Chem. 2014, 145, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Nasopoulou, C.; Gogaki, V.; Stamatakis, G.; Papaharisis, L.; Demopoulos, C.A.; Zabetakis, I. Evaluation of the in vitro anti-atherogenic properties of lipid fractions of olive pomace, olive pomace enriched fish feed and gilthead sea bream (Sparus aurata) fed with olive pomace enriched fish feed. Mar. Drugs 2013, 11, 3676–3688. [Google Scholar] [CrossRef] [PubMed]
- De Leonardis, A.; Macciola, V. A study on the lipid fraction of Adriatic sardine filets (Sardina pilchardus). Food/Nahrung 2004, 48, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Kangsen, M.; Qinghui, A.; Milley, E.J.; Lall, P.S. Lipid and fatty acid compositions of cod (Gadus morhua), haddock (Melanogrammus aeglefinus) and halibut (Hippoglossus hippoglossus). J. Ocean Univ. China 2010, 9, 381–388. [Google Scholar]
- Kurlansky, M. Cod: A Biography of the Fish That Changed the World; Penguin Group: London, UK, 1999. [Google Scholar]
- Lie, O.E.; Lied, E.; Lambertsen, G. Liver retention of fat and of fatty acids in cod (Gadus morhua) fed different oils. Aquaculture 1986, 59, 187–196. [Google Scholar] [CrossRef]
- Calvano, C.D.; Zambonin, C.G.; Foti, C.; Cassano, N.; Vena, G.A. A matrix assisted laser desorption ionization time-of-flight mass spectrometry investigation to assess the composition of cod liver oil based products which displayed a different in vivo allergenic power. Food Chem. Toxicol. 2008, 46, 3580–3585. [Google Scholar] [CrossRef] [PubMed]
- Rice, R. Fish Oils/Dietary Importance. In Encyclopedia of Food Sciences and Nutrition; The Fish Foundation: Tiverton, UK, 2003. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Galanos, D.S.; Kapoulas, V.M. Isolation of polar lipids from triglyceride mixtures. J. Lipid Res. 1962, 3, 134–137. [Google Scholar]
- Swapna, H.C.; Rai, A.K.; Bhaskar, N.; Sachindra, N.M. Lipid classes and fatty acid profile of selected Indian fresh water fishes. J. Food Sci. Technol. 2010, 47, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.G.; Han, B.W.; Kim, H.S.; Park, C.H.; Chung, I.K.; Choi, Y.J.; Kim, J.S.; Heu, M.S. Lipid characteristics of fish frame as a functional lipid resource. J. Korean Soc. Food Sci. Nutr. 2005, 34, 380–388. [Google Scholar]
- Ackman, R.G. Nutritional composition of fats in seafoods. Prog. Food Nutr. Sci. 1989, 13, 161–241. [Google Scholar] [PubMed]
- Bandarra, N.M.; Batista, I.; Nunes, M.L.; Empis, J.M.; Christie, W.W. Seasonal changes in lipid composition of sardine (Sardina pilchardus). J. Food Sci. 1997, 62, 40–42. [Google Scholar] [CrossRef]
- Feliste, R.; Perret, B.; Braqueta, P.; Chap, H. Protective effect of BN 52021, a specific antagonist of platelet-activating factor (PAF acether) against diet-induced cholesteryl ester deposition in rabbit arta. Atherosclerosis 1989, 78, 151–158. [Google Scholar] [CrossRef]
- Karantonis, H.C.; Antonopoulou, S.; Perrea, D.N.; Sokolis, D.P.; Theocharis, S.E.; Kavantzas, N.; Iliopoulos, D.G.; Demopoulos, C.A. In vivo antiatherogenic properties of olive oil and its constituent lipid classes in hyperlipidemic rabbits. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Wierzbicki, A.S.; Poston, R.; Ferro, A. The lipid and non-lipid effects of statins. Pharmacol. Ther. 2003, 99, 95–112. [Google Scholar] [CrossRef]
- Stafforini, D.M.; Zimmerman, G.A. Unraveling the PAF-AH/Lp-PLA 2 controversy. J. Lipid Res. 2014, 55, 1811–1814. [Google Scholar] [CrossRef] [PubMed]
- Tsantila, N.; Karantonis, H.C.; Perrea, D.N.; Theocharis, S.E.; Iliopoulos, D.G.; Antonopoulou, S.; Demopoulos, C.A. Antithrombotic and antiatherosclerotic properties of olive oil and pomace polar extracts in rabbits. Med. Inflamm. 2007, 2007, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Nasopoulou, C.; Stamatakis, G.; Demopoulos, C.A.; Zabetakis, I. Effects of olive pomace and olive pomace oil on growth performance, fatty acid composition and cardioprotective properties of gilthead sea bream (Sparus aurata) and sea bass (Dicentrarchus labrax). Food Chem. 2011, 129, 1108–1113. [Google Scholar] [CrossRef] [PubMed]
- Lazana, L.K.; Lazanas, M.; Koussissis, S.; Tournis, S.; Demopoulos, C.A. PAF of Biological fluids in desease: Blood levels in allergic rhinitis. Haematologica 1988, 73, 379–382. [Google Scholar]
- Antonopoulou, S.; Demopoulos, C.A.; Andrikopoulos, N.K. Lipid separation from Urtica dioica: Existence of platelet-activating factor. J. Agric. Food Chem. 1996, 44, 3052–3056. [Google Scholar] [CrossRef]
- Bimbo, A.P. Sources of marine oils. In Marine Oils, from Sea to Pharmaceuticals, 1st ed.; Nova Science Publishers: Hauppauge, NY, USA, 2015; p. 1. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morphis, G.; Kyriazopoulou, A.; Nasopoulou, C.; Sioriki, E.; Demopoulos, C.A.; Zabetakis, I. Assessment of the in Vitro Antithrombotic Properties of Sardine (Sardina pilchardus) Fillet Lipids and Cod Liver Oil. Fishes 2016, 1, 1-15. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes1010001
Morphis G, Kyriazopoulou A, Nasopoulou C, Sioriki E, Demopoulos CA, Zabetakis I. Assessment of the in Vitro Antithrombotic Properties of Sardine (Sardina pilchardus) Fillet Lipids and Cod Liver Oil. Fishes. 2016; 1(1):1-15. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes1010001
Chicago/Turabian StyleMorphis, Gregory, Aggeliki Kyriazopoulou, Constantina Nasopoulou, Eleni Sioriki, Constantinos A. Demopoulos, and Ioannis Zabetakis. 2016. "Assessment of the in Vitro Antithrombotic Properties of Sardine (Sardina pilchardus) Fillet Lipids and Cod Liver Oil" Fishes 1, no. 1: 1-15. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes1010001