In Vitro Effects of Bisphenol A and Tetrabromobisphenol A on Cell Viability and Reproduction-Related Gene Expression in Pituitaries from Sexually Maturing Atlantic Cod (Gadus morhua L.)

 ,
,  and
and
Abstract
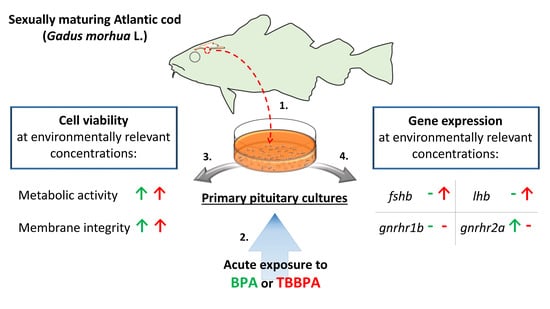
:
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Dispersed Pituitary Primary Cell Cultures
2.3. BPA and TBBPA Exposure
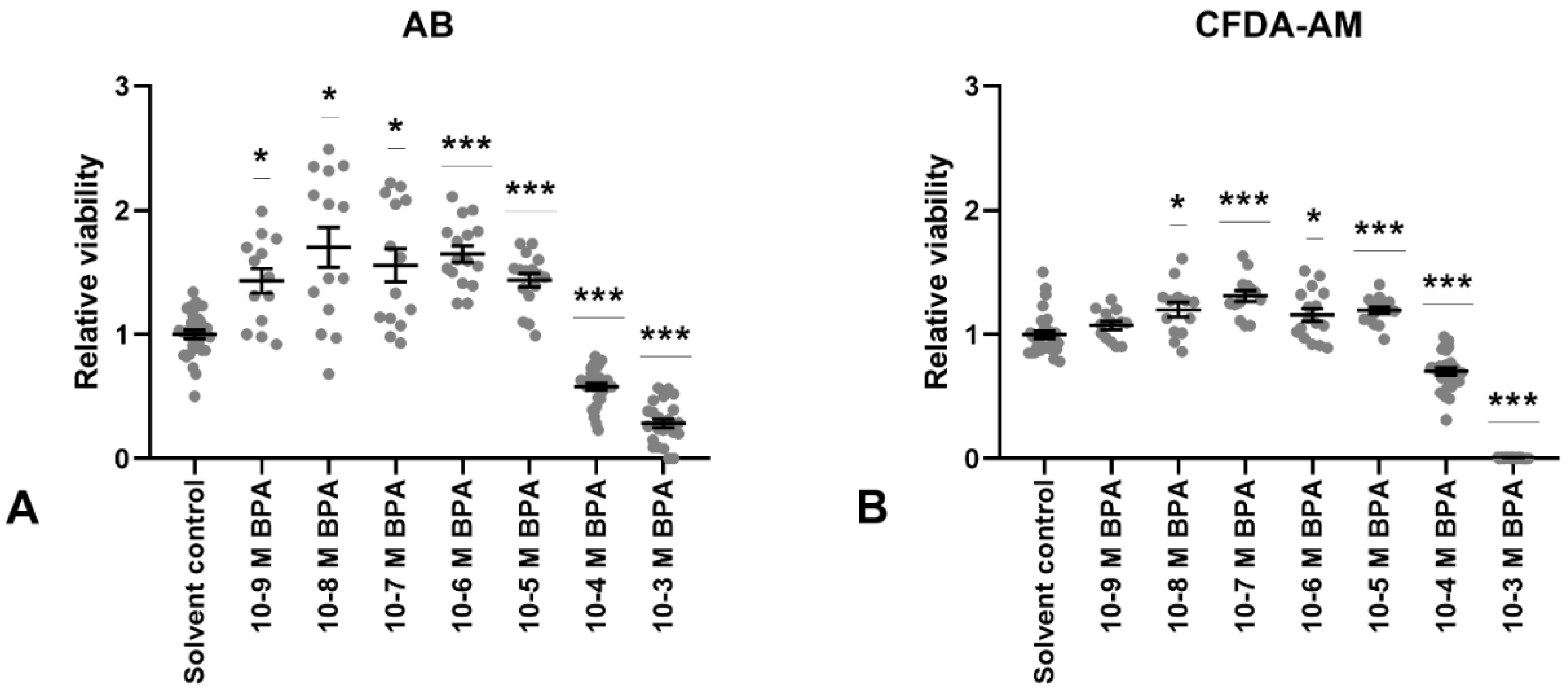
2.4. Viability Assays
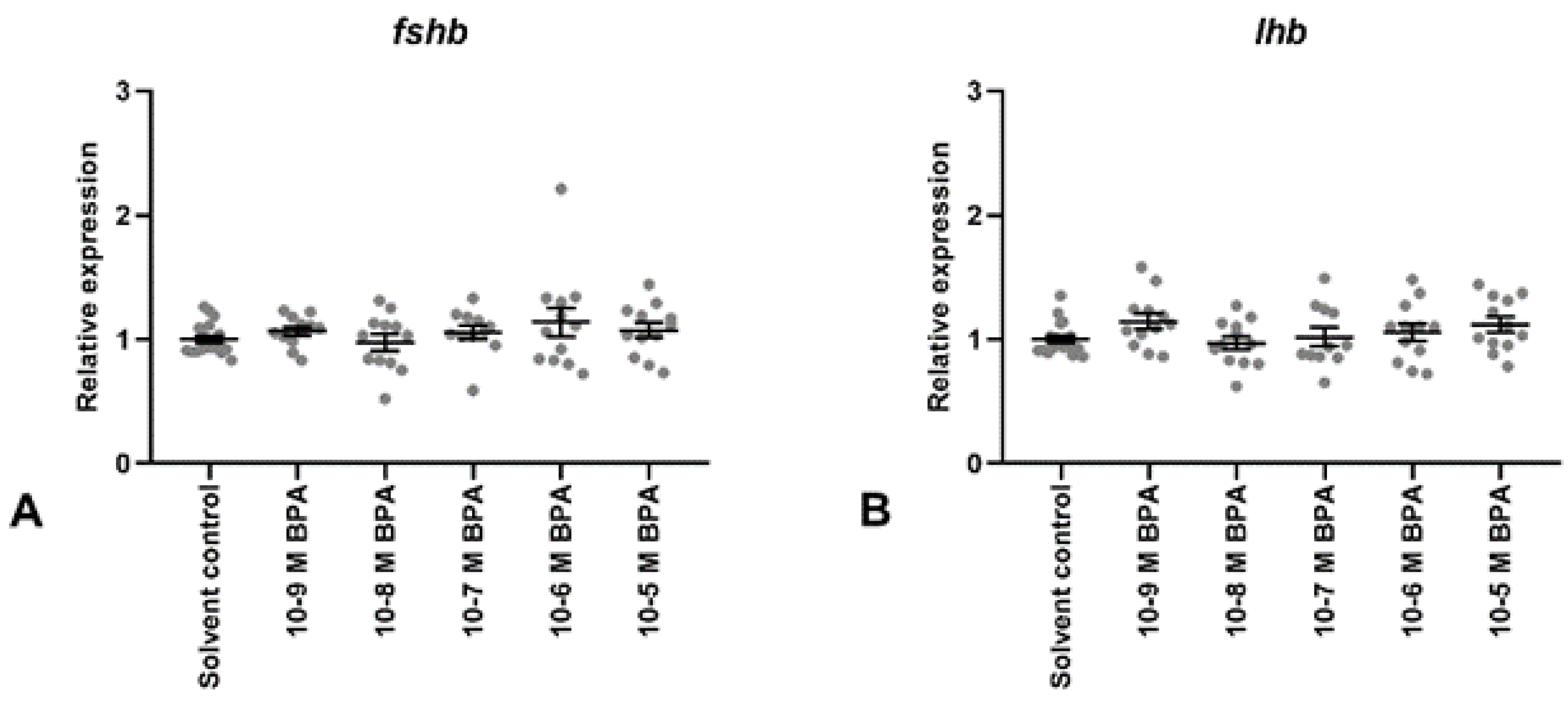
2.5. Quantification of Gene Expression
2.5.1. RNA Extraction and cDNA Synthesis
2.5.2. Primers and Reference Genes
2.5.3. qPCR Analysis
2.6. Statistical Analysis and Data Presentation
3. Results
3.1. Solvent Control vs Control Blank
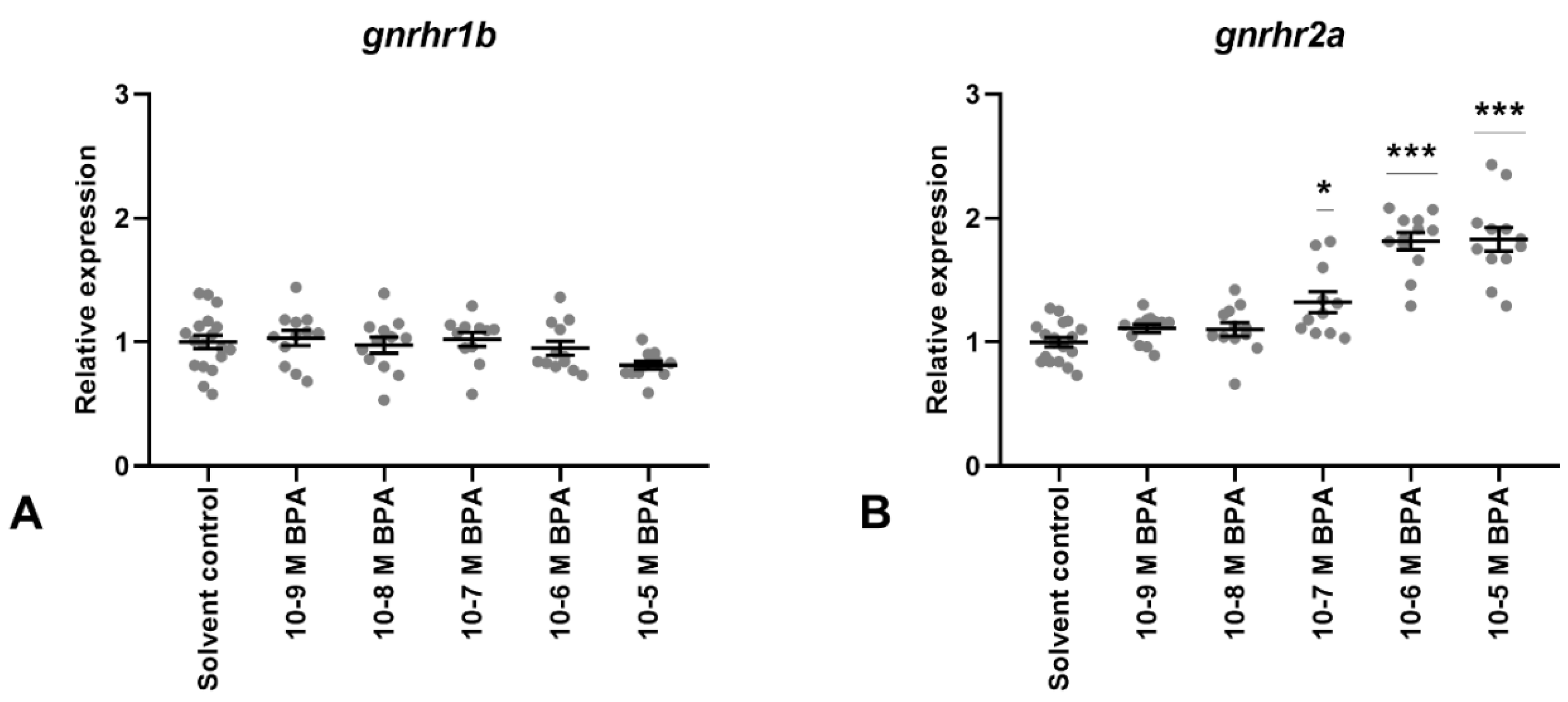
3.2. BPA Exposure
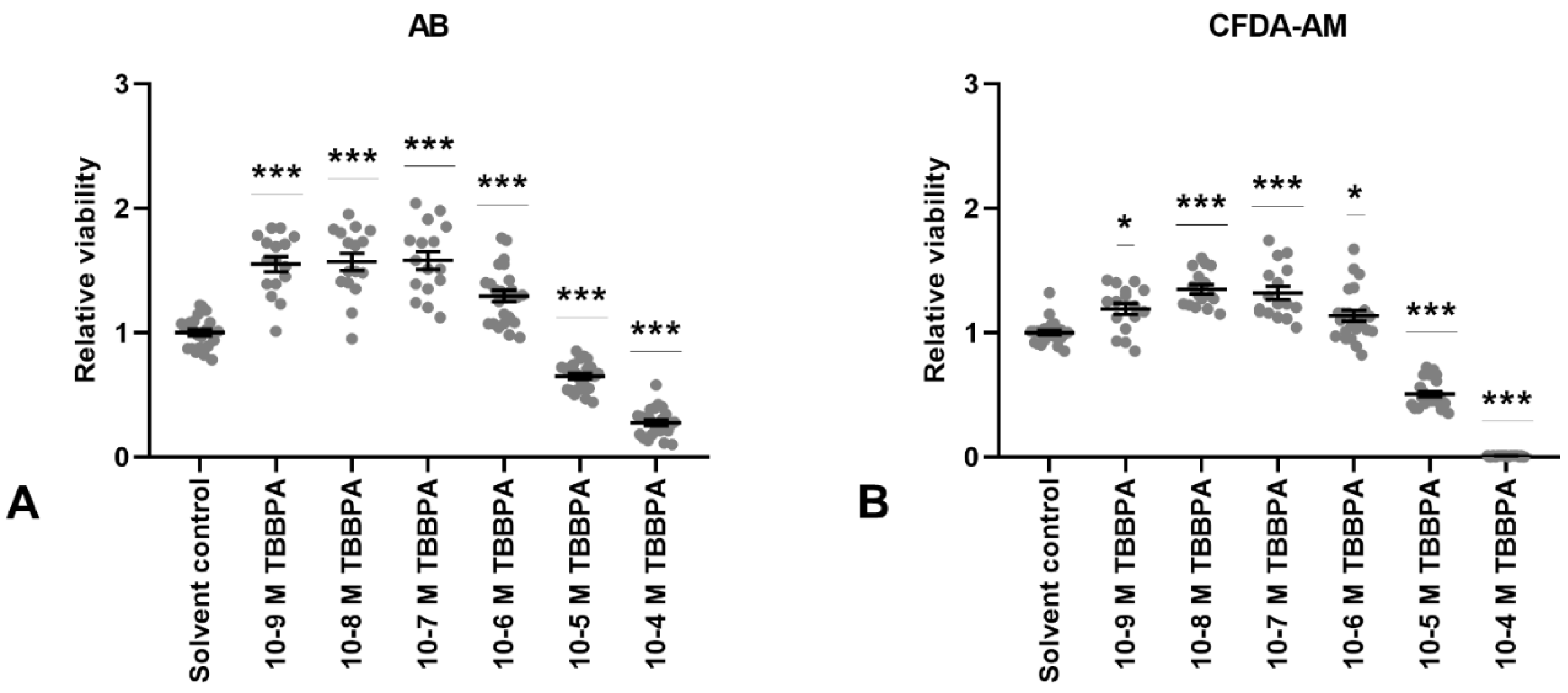
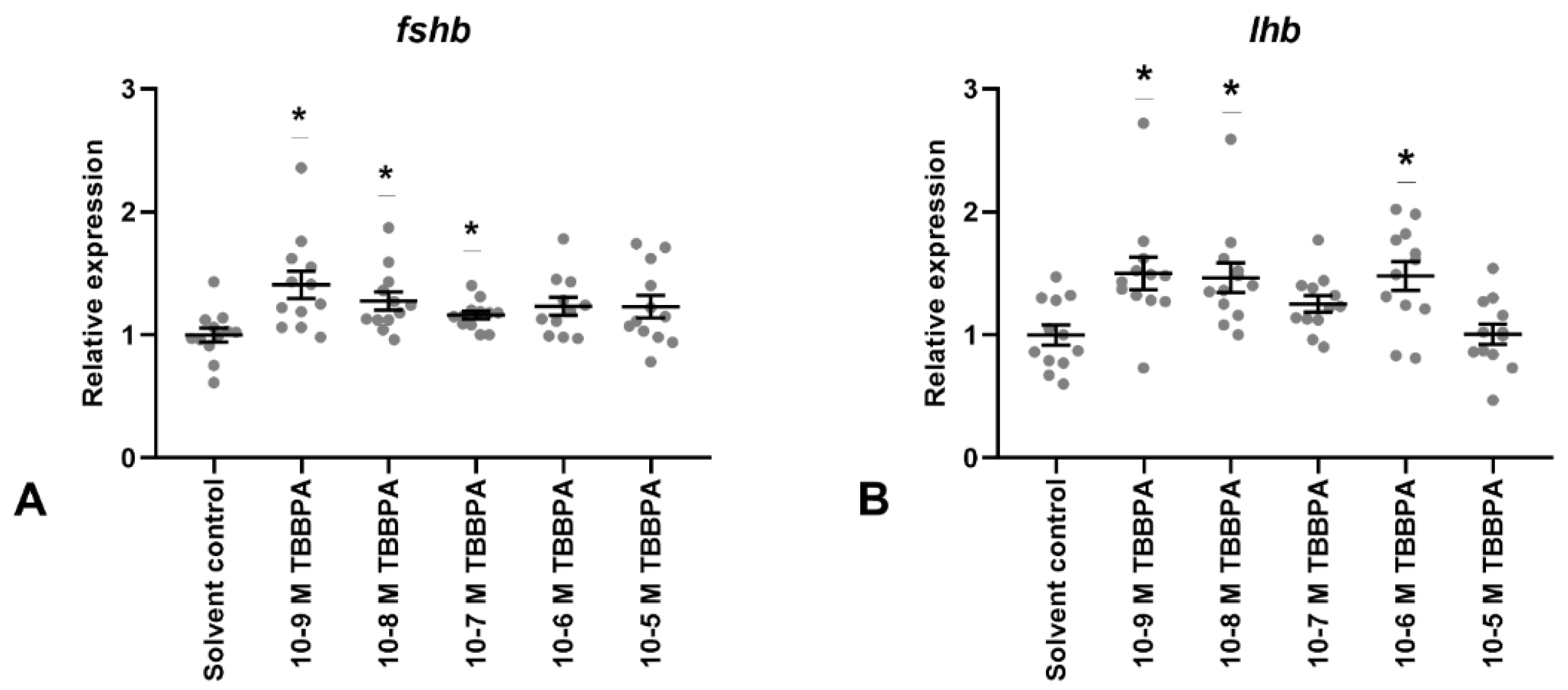
3.3. TBBPA Exposure
4. Discussion
4.1. Methodological Considerations
4.2. BPA
4.3. TBBPA
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yaron, Z.; Gur, G.; Melamed, P.; Rosenfeld, H.; Elizur, A.; Levavi-Sivan, B. Regulation of fish gonadotropins. In International Review or Cytology—A Survey of Cell Biology; Jeon, K.W., Ed.; Elsevier: Amsterdam, The Netherlands, 2003; Volume 225, pp. 131–185. [Google Scholar]
- Levavi-Sivan, B.; Bogerd, J.; Mananos, E.L.; Gomez, A.; Lareyre, J.J. Perspectives on fish gonadotropins and their receptors. Gen. Comp. Endocrinol. 2010, 165, 412–437. [Google Scholar] [CrossRef] [PubMed]
- von Krogh, K.; Bjørndal, G.T.; Nourizadeh-Lillabadi, R.; Hodne, K.; Ropstad, E.; Haug, T.M.; Weltzien, F.A. Sex steroids differentially regulate FSHb, LHb and GnRHr expression in Atlantic cod (Gadus morhua) pituitary. Reproduction 2017, 154, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Weltzien, F.A.; Andersson, E.; Andersen, Ø.; Shalchian-Tabrizi, K.; Norberg, B. The brain-pituitary-gonad axis in male teleosts, with special emphasis on flatfish (pleuronectiformes). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2004, 137, 447–477. [Google Scholar] [CrossRef] [PubMed]
- WHO/ICPS. Environmental Health Criteria 172: Tetrabromobisphenol A and Derivatives; World Health Organization: Geneva, Switzerland, 1995. [Google Scholar]
- EU. European Union Risk Assessment Report for 4,40-isopropylidenediphenol (bisphenol-A); Office for Official Publications of the European Communities Communities: Luxembourg, 2008. [Google Scholar]
- Flint, S.; Markle, T.; Thompson, S.; Wallace, E. Bisphenol A exposure, effects, and policy: A wildlife perspective. J. Environ. Manag. 2012, 104, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Q.; Wong, C.K.C.; Zheng, J.S.; Bouwman, H.; Barra, R.; Wahlstrom, B.; Neretin, L.; Wong, M.H. Bisphenol A (BPA) in China: A review of sources, environmental levels, and potential human health impacts. Environ. Int. 2012, 42, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Crain, D.A.; Eriksen, M.; Iguchi, T.; Jobling, S.; Laufer, H.; LeBlanc, G.A.; Guillette, L.J. An ecological assessment of bisphenol-A: Evidence from comparative biology. Reprod. Toxicol. 2007, 24, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.; Allchin, C.R.; Zegers, B.N.; Haftka, J.J.; Boon, J.P.; Belpaire, C.; Leonards, P.E.; Van Leeuwen, S.P.; De Boer, J. Distribution and fate of HBCD and TBBPA brominated flame retardants in North Sea estuaries and aquatic food webs. Environ. Sci. Technol. 2004, 38, 5497–5504. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wang, S.; Liu, H.; Yan, Z. Tetrabromobisphenol A: Tissue distribution in fish, and seasonal variation in water and sediment of Lake Chaohu, China. Environ. Sci. Pollut. Res. Int. 2012, 19, 4090–4096. [Google Scholar] [CrossRef]
- Gong, W.-J.; Zhu, L.-Y.; Jiang, T.-T.; Han, C. The occurrence and spatial-temporal distribution of tetrabromobisphenol A in the coastal intertidal zone of Qingdao in China, with a focus on toxicity assessment by biological monitoring. Chemosphere 2017, 185, 462–467. [Google Scholar] [CrossRef]
- Sajiki, J.; Yonekubo, J. Leaching of bisphenol A (BPA) to seawater from polycarbonate plastic and its degradation by reactive oxygen species. Chemosphere 2003, 51, 55–62. [Google Scholar] [CrossRef]
- Kang, J.H.; Kondo, F. Bisphenol A degradation in seawater is different from that in river water. Chemosphere 2005, 60, 1288–1292. [Google Scholar] [CrossRef] [PubMed]
- Corrales, J.; Kristofco, L.A.; Steele, W.B.; Yates, B.S.; Breed, C.S.; Williams, E.S.; Brooks, B.W. Global Assessment of Bisphenol A in the Environment: Review and Analysis of Its Occurrence and Bioaccumulation. Dose Response 2015, 13, 1559325815598308. [Google Scholar] [CrossRef] [PubMed]
- Staniszewska, M.; Koniecko, I.; Falkowska, L.; Krzymyk, E. Occurrence and distribution of bisphenol A and alkylphenols in the water of the gulf of Gdansk (Southern Baltic). Mar. Pollut. Bull. 2015, 91, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Wang, L.H.; Wang, X.Q.; Liu, S.Z.; Xu, P.; Wang, H.P.; Wu, T.T.; Zhang, Y.Y.; Zheng, Y.; Li, M.; et al. Bisphenol A affects gene expression of gonadotropin-releasing hormones and type I GnRH receptors in brains of adult rare minnow Gobiocypris rarus. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2013, 157, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Faheem, M.; Jahan, N.; Khaliq, S.; Lone, K.P. Modulation of brain kisspeptin expression after bisphenol-A exposure in a teleost fish, Catla catla. Fish. Physiol. Biochem. 2019, 45, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.; Genco, M.C.; Megrelis, L.; Ruderman, J.V. Effects of bisphenol A and triclocarban on brain-specific expression of aromatase in early zebrafish embryos. Proc. Natl. Acad. Sci. USA 2011, 108, 17732–17737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molina, A.; Abril, N.; Morales-Prieto, N.; Monterde, J.; Ayala, N.; Lora, A.; Moyano, R. Hypothalamic-pituitary-ovarian axis perturbation in the basis of bisphenol A (BPA) reproductive toxicity in female zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2018, 156, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Cano-Nicolau, J.; Vaillant, C.; Pellegrini, E.; Charlier, T.D.; Kah, O.; Coumailleau, P. Estrogenic Effects of Several BPA Analogs in the Developing Zebrafish Brain. Front. Neurosci. 2016, 10, 112. [Google Scholar] [CrossRef] [Green Version]
- Rhee, J.-S.; Kim, R.-O.; Seo, J.S.; Kang, H.S.; Park, C.-B.; Soyano, K.; Lee, J.; Lee, Y.-M.; Lee, J.-S. Bisphenol A modulates expression of gonadotropin subunit genes in the hermaphroditic fish, Kryptolebias marmoratus. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 152, 456–466. [Google Scholar] [CrossRef]
- Qiu, W.H.; Zhao, Y.L.; Yang, M.; Farajzadeh, M.; Pan, C.Y.; Wayne, N.L. Actions of Bisphenol A and Bisphenol S on the Reproductive Neuroendocrine System During Early Development in Zebrafish. Endocrinology 2016, 157, 636–647. [Google Scholar] [CrossRef]
- Chen, W.T.; Lau, S.W.; Fan, Y.Q.; Wu, R.S.S.; Ge, W. Juvenile exposure to bisphenol A promotes ovarian differentiation but suppresses its growth—Potential involvement of pituitary follicle stimulating hormone. Aquat. Toxicol. 2017, 193, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, H.; Yang, M.; Yu, Y.; Yan, M.; Zhou, L.; Liu, X.; Xiao, S.; Yang, Y.; Wang, Y.; et al. Toxic effects of bisphenol A on goldfish gonad development and the possible pathway of BPA disturbance in female and male fish reproduction. Chemosphere 2019, 221, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Mandich, A.; Bottero, S.; Benfenati, E.; Cevasco, A.; Erratico, C.; Maggioni, S.; Massari, A.; Pedemonte, F.; Vigano, L. In vivo exposure of carp to graded concentrations of bisphenol A. Gen. Comp. Endocrinol. 2007, 153, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Faheem, M.; Khaliq, S.; Lone, K.P. Disruption of the Reproductive Axis in Freshwater Fish, Catla catla, After Bisphenol-A Exposure. Zool. Sci. 2017, 34, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Chen, Y.; Lin, L.; Liu, Y.; Chi, Y.; Lin, Y.; Ye, G.; Zhu, H.; Dong, S. Different effects of bisphenol a and its halogenated derivatives on the reproduction and development of Oryzias melastigma under environmentally relevant doses. Sci. Total Environ. 2017, 595, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Hatef, A.; Alavi, S.M.H.; Abdulfatah, A.; Fontaine, P.; Rodina, M.; Linhart, O. Adverse effects of bisphenol A on reproductive physiology in male goldfish at environmentally relevant concentrations. Ecotoxicol. Environ. Saf. 2012, 76, 56–62. [Google Scholar] [CrossRef]
- Kuiper, R.V.; Canton, R.F.; Leonards, P.E.G.; Jenssen, B.M.; Dubbeldam, M.; Wester, P.W.; Van den Berg, M.; Vos, J.G.; Vethaak, A.D. Long-term exposure of European flounder (Platichthys flesus) to the flame-retardants tetrabromobisphenol A (TBBPA) and hexabromocyclododecane (HBCD). Ecotoxicol. Environ. Saf. 2007, 67, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, R.V.; van den Brandhof, E.J.; Leonards, P.E.G.; Van der Ven, L.T.M.; Wester, P.W.; Vos, J.G. Toxicity of tetrabromobisphenol A (TBBPA) in zebrafish (Danio rerio) in a partial life-cycle test. Arch. Toxicol. 2007, 81, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Suzuki, T.; Sanoh, S.; Kohta, R.; Jinno, N.; Sugihara, K.; Yoshihara, S.; Fujimoto, N.; Watanabe, H.; Ohta, S. Comparative study of the endocrine-disrupting activity of bisphenol A and 19 related compounds. Toxicol. Sci. 2005, 84, 249–259. [Google Scholar] [CrossRef]
- Hamers, T.; Kamstra, J.H.; Sonneveld, E.; Murk, A.J.; Kester, M.H.A.; Andersson, P.L.; Legler, J.; Brouwer, A. In Vitro Profiling of the Endocrine-Disrupting Potency of Brominated Flame Retardants. Toxicol. Sci. 2006, 92, 157–173. [Google Scholar] [CrossRef] [Green Version]
- Krivoshiev, B.V.; Dardenne, F.; Covaci, A.; Blust, R.; Husson, S.J. Assessing in-vitro estrogenic effects of currently-used flame retardants. Toxicol. In Vitro 2016, 33, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, L.B.; Pedersen, K.L.; Pedersen, S.N.; Korsgaard, B.; Bjerregaard, P. In vivo comparison of xenoestrogens using rainbow trout vitellogenin induction as a screening system. Environ. Toxicol. Chem. 2000, 19, 1867–1874. [Google Scholar] [CrossRef]
- Song, M.; Liang, D.; Liang, Y.; Chen, M.; Wang, F.; Wang, H.; Jiang, G. Assessing developmental toxicity and estrogenic activity of halogenated bisphenol A on zebrafish (Danio rerio). Chemosphere 2014, 112, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Chow, W.S.; Chan, W.K.-L.; Chan, K.M. Toxicity assessment and vitellogenin expression in zebrafish (Danio rerio) embryos and larvae acutely exposed to bisphenol A, endosulfan, heptachlor, methoxychlor and tetrabromobisphenol A. J. Appl. Toxicol. 2013, 33, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.-Y.; Ying, G.-G.; Liang, Y.-Q.; Zhao, J.-L.; Yang, B.; Liu, S.; Liu, Y.-S. Hormonal effects of tetrabromobisphenol A using a combination of in vitro and in vivo assays. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2013, 157, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Ronisz, D.; Farmen Finne, E.; Karlsson, H.; Förlin, L. Effects of the brominated flame retardants hexabromocyclododecane (HBCDD), and tetrabromobisphenol A (TBBPA), on hepatic enzymes and other biomarkers in juvenile rainbow trout and feral eelpout. Aquat. Toxicol. 2004, 69, 229–245. [Google Scholar] [CrossRef] [PubMed]
- Meerts, I.; van Zanden, J.J.; Luijks, E.A.C.; van Leeuwen-Bol, I.; Marsh, G.; Jakobsson, E.; Bergman, A.; Brouwer, A. Potent competitive interactions of some brominated flame retardants and related compounds with human transthyretin in vitro. Toxicol. Sci. 2000, 56, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, S.; Jinno, N.; Ohta, S.; Kuroki, H.; Fujimoto, N. Thyroid hormonal activity of the flame retardants tetrabromobisphenol A and tetrachlorobisphenol A. Biochem. Biophys. Res. Commun. 2002, 293, 554–559. [Google Scholar] [CrossRef]
- Chan, W.K.; Chan, K.M. Disruption of the hypothalamic-pituitary-thyroid axis in zebrafish embryo-larvae following waterborne exposure to BDE-47, TBBPA and BPA. Aquat. Toxicol. 2012, 108, 106–111. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, W.; Chen, B.; He, J.; Chen, F.; Shan, X.; Du, Q.; Li, N.; Jia, X.; Tang, J. Differences in reproductive toxicity of TBBPA and TCBPA exposure in male Rana nigromaculata. Environ. Pollut. 2018, 243, 394–403. [Google Scholar] [CrossRef]
- van der Ven, L.T.M.; van de Kuil, T.V.; Verhoef, A.; Verwer, C.M.; Lilienthal, H.; Leonards, P.E.G.; Schauer, U.M.D.; Canton, R.F.; Litens, S.; De Jong, F.H.; et al. Endocrine effects of tetrabromobisphenol-A (TBBPA) in Wistar rats as tested in a one-generation reproduction study and a subacute toxicity study. Toxicology 2008, 245, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Hodne, K.; von Krogh, K.; Weltzien, F.A.; Sand, O.; Haug, T.M. Optimized conditions for primary culture of pituitary cells from the Atlantic cod (Gadus morhua). The importance of osmolality, pCO(2), and pH. Gen. Comp. Endocrinol. 2012, 178, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Bopp, S.K.; Lettieri, T. Comparison of four different colorimetric and fluorometric cytotoxicity assays in a zebrafish liver cell line. BMC Pharmacol. 2008, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- von Krogh, K.; Bjørndal, G.T.; Nourizadeh-Lillabadi, R.; Ropstad, E.; Haug, T.M.; Weltzien, F.-A. Cortisol differentially affects cell viability and reproduction-related gene expression in Atlantic cod pituitary cultures dependent on stage of sexual maturation. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 236, 110517. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Hodne, K.; Haug, T.M.; Weltzien, F.A. Single-cell qPCR on dispersed primary pituitary cells—An optimized protocol. BMC Mol. Biol. 2010, 11, 82. [Google Scholar] [CrossRef]
- Weltzien, F.A.; Pasqualini, C.; Vernier, P.; Dufour, S. A quantitative real-time RT-PCR assay for European eel tyrosine hydroxylase. Gen. Comp. Endocrinol. 2005, 142, 134–142. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Roche. Roche Applied Science; Technical note No. LC 13; Roche Diagnostics GmbH: Mannheim, Germany, 2001. [Google Scholar]
- Ager-Wick, E.; Hodne, K.; Fontaine, R.; von Krogh, K.; Haug, T.M.; Weltzien, F.A. Preparation of a High-quality Primary Cell Culture from Fish Pituitaries. J. Vis. Exp. 2018, e58159. [Google Scholar] [CrossRef] [Green Version]
- Baker, R.C.; Kramer, R.E. Cytotoxicity of short-chain alcohols. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 127–150. [Google Scholar] [CrossRef]
- Pal, R.; Mamidi, M.K.; Das, A.K.; Bhonde, R. Diverse effects of dimethyl sulfoxide (DMSO) on the differentiation potential of human embryonic stem cells. Arch. Toxicol. 2012, 86, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Fjeld, E.; Schlabach, M.; Berge, J.A.; Eggen, T.; Snilsberg, P.; Källberg, G.; Rognerud, S.; Enge, E.K.; Borgen, A.; Gundersen, H. Kartlegging av Utvalgte nye Organiske Miljøgifter—Bromerte Flammehemmere, Klorerte Parafiner, Bisfenol A og Triclosan; NIVA Rapport 4809-2004; Norsk institutt for vannforskning (NIVA): Oslo, Norway, 2004. (In Norwegian) [Google Scholar]
- Lv, X.; Zhou, Q.; Song, M.; Jiang, G.; Shao, J. Vitellogenic responses of 17β-estradiol and bisphenol A in male Chinese loach (Misgurnus anguillicaudatus). Environ. Toxicol. Pharmacol. 2007, 24, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Larsen, B.K.; Bjornstad, A.; Sundt, R.C.; Taban, I.C.; Pampanin, D.M.; Andersen, O.K. Comparison of protein expression in plasma from nonylphenol and bisphenol A-exposed Atlantic cod (Gadus morhua) and turbot (Scophthalmus maximus) by use of SELDI-TOF. Aquat. Toxicol. 2006, 78, S25–S33. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.J.; Chen, D.W.; Liu, J.L.; Zhang, J.H.; Luo, H.S.; Cui, S. Estradiol promotes pituitary cell proliferation and gonadotroph differentiation at different doses and with different mechanisms in chick embryo. Steroids 2009, 74, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Brannick, K.E.; Craig, Z.R.; Himes, A.D.; Peretz, J.R.; Wang, W.; Flaws, J.A.; Raetzman, L.T. Prenatal Exposure to Low Doses of Bisphenol A Increases Pituitary Proliferation and Gonadotroph Number in Female Mice Offspring at Birth. Biol. Reprod. 2012, 87, 10. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Chan, K.M. Evaluation of the toxic effects of brominated compounds (BDE-47, 99, 209, TBBPA) and bisphenol A (BPA) using a zebrafish liver cell line, ZFL. Aquat. Toxicol. 2015, 159, 138–147. [Google Scholar] [CrossRef]
- Ma, M.; Crump, D.; Farmahin, R.; Kennedy, S.W. Comparing the effects of tetrabromobisphenol-A, bisphenol A, and their potential replacement alternatives, TBBPA-bis(2,3-dibromopropyl ether) and bisphenol S, on cell viability and messenger ribonucleic acid expression in chicken embryonic hepatocytes. Environ. Toxicol. Chem. 2015, 34, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Akingbemi, B.T.; Sottas, C.M.; Koulova, A.I.; Klinefelter, G.R.; Hardy, M.P. Inhibition of Testicular Steroidogenesis by the Xenoestrogen Bisphenol A Is Associated with Reduced Pituitary Luteinizing Hormone Secretion and Decreased Steroidogenic Enzyme Gene Expression in Rat Leydig Cells. Endocrinology 2004, 145, 592–603. [Google Scholar] [CrossRef]
- Wisniewski, P.; Romano, R.M.; Kizys, M.M.L.; Oliveira, K.C.; Kasamatsu, T.; Giannocco, G.; Chiamolera, M.I.; Dias-Da-Silva, M.R.; Romano, M.A. Adult exposure to bisphenol A (BPA) in Wistar rats reduces sperm quality with disruption of the hypothalamic-pituitary-testicular axis. Toxicology 2015, 329, 1–9. [Google Scholar] [CrossRef]
- Hildahl, J.; Sandvik, G.K.; Edvardsen, R.B.; Norberg, B.; Haug, T.M.; Weltzien, F.-A. Four gonadotropin releasing hormone receptor genes in Atlantic cod are differentially expressed in the brain and pituitary during puberty. Gen. Comp. Endocrinol. 2011, 173, 333–345. [Google Scholar] [CrossRef]
- Rhee, J.-S.; Seo, J.S.; Raisuddin, S.; Ki, J.-S.; Lee, K.-W.; Kim, I.-C.; Yoon, Y.-D.; Lee, J.-S. Gonadotropin-releasing hormone receptor (GnRHR) gene expression is differently modulated in gender types of the hermaphroditic fish Kryptolebias marmoratus by endocrine disrupting chemicals. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2008, 147, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, Z.; Wang, Q.; Zhai, Y. Effects of BPA and E2 on expression profiles of genes related to hypothalamic-pituitary-gonadal axis of half-smooth tongue sole Cynoglossus semilaevis. Chin. J. Oceanol. Limnol. 2013, 31, 598–608. [Google Scholar] [CrossRef]
- Hu, J.; Liang, Y.; Chen, M.; Wang, X. Assessing the toxicity of TBBPA and HBCD by zebrafish embryo toxicity assay and biomarker analysis. Environ. Toxicol. 2009, 24, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Reistad, T.; Mariussen, E.; Ring, A.; Fonnum, F. In vitro toxicity of tetrabromobisphenol-A on cerebellar granule cells: Cell death, free radical formation, calcium influx and extracellular glutamate. Toxicol. Sci. 2007, 96, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Dittrich, R.; Beckmann, M.W.; Oppelt, P.G.; Hoffmann, I.; Lotz, L.; Kuwert, T.; Mueller, A. Thyroid hormone receptors and reproduction. J. Reprod. Immunol. 2011, 90, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Dardente, H.; Hazlerigg, D.G.; Ebling, F.J.P. Thyroid hormone and seasonal rhythmicity. Front. Endocrinol. 2014, 5, 19. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Reference | Primer Sequence | Amplicon Size (nt) | Efficiency | |
|---|---|---|---|---|---|
| lhb | [49] | Forward | 5′-GTGGAGAAGAAGGGCTGTCC-3′ | 81 | 1.93 |
| Reverse | 5′-GGACGGGTCCATGGTG-3′ | ||||
| fshb | [49] | Forward | 5′-GAACCGAGTCCATCAACACC-3′ | 63 | 1.84 |
| Reverse | 5′-GGTCCATCGGGTCCTCCT-3′ | ||||
| gnrhr1b | [3] | Forward | 5′-GCTACTCCCGAATCCTCCTC-3′ | 73 | 1.96 |
| Reverse | 5′-CGCCTCAGGTATGACTCTCC-3′ | ||||
| gnrhr2a | [3] | Forward | 5′-TTCACCTTCTGCTGCCTCTT-3′ | 113 | 1.99 |
| Reverse | 5′-TCCGTGGAGGAAAGATTGTC-3′ | ||||
| bactin | [45] | Forward | 5′-TTCTACAACGAGCTGAGAGTGG-3′ | 102 | 1.84 |
| Reverse | 5′-CATGATCTGGGTCATCTTCTCC-3′ | ||||
| arp2 | [45] | Forward | 5′-GGAGGTTAGAAGTAGCAAGGAGC-3′ | 107 | 1.94 |
| Reverse | 5′-TGCTGACTCTCACGGAGTTG-3′ | ||||
| ef1a | [49] | Forward | 5′-CCTTCAACGCCCAGGTCAT-3′ | 100 | 1.92 |
| Reverse | 5′-AACTTGCAGGCGATGTGA-3′ | ||||
| ubiquitin | [45] | Forward | 5′-TGTCAAAGCCAAGATTCAGG-3′ | 111 | 1.86 |
| Reverse | 5′-TGGATGTTGTAATCCGAGAGG-3′ | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
von Krogh, K.; Ropstad, E.; Nourizadeh-Lillabadi, R.; Haug, T.M.; Weltzien, F.-A. In Vitro Effects of Bisphenol A and Tetrabromobisphenol A on Cell Viability and Reproduction-Related Gene Expression in Pituitaries from Sexually Maturing Atlantic Cod (Gadus morhua L.). Fishes 2019, 4, 48. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4030048
von Krogh K, Ropstad E, Nourizadeh-Lillabadi R, Haug TM, Weltzien F-A. In Vitro Effects of Bisphenol A and Tetrabromobisphenol A on Cell Viability and Reproduction-Related Gene Expression in Pituitaries from Sexually Maturing Atlantic Cod (Gadus morhua L.). Fishes. 2019; 4(3):48. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4030048
Chicago/Turabian Stylevon Krogh, Kristine, Erik Ropstad, Rasoul Nourizadeh-Lillabadi, Trude Marie Haug, and Finn-Arne Weltzien. 2019. "In Vitro Effects of Bisphenol A and Tetrabromobisphenol A on Cell Viability and Reproduction-Related Gene Expression in Pituitaries from Sexually Maturing Atlantic Cod (Gadus morhua L.)" Fishes 4, no. 3: 48. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes4030048