1. Introduction
The intimate interaction of fish with the aquatic environment, where challenges are persistent, places them under immense biological pressure [
1]. Nonetheless, fish have developed a complex repertoire of immune defense to combat these challenging biotic and abiotic factors. The mucosal surfaces serve as the primary barrier between the internal and external environments, and they interact with the water matrix more than any other organs in fish. Considered to be the first line of defense, mucosal surfaces have several layers of physical and biological barriers [
2]. The mucus layer is an emblematic feature of mucosal surfaces. These viscoelastic glycopolymeric secretions from goblet or mucous producing cells are dynamic matrices, and their composition varies amongst fish species [
3]. Water is the major component of mucus, comprising approximately 90%–95%, and serves as the solvent and diffusion medium for other chemical factors present in the secretions [
4]. One distinctive characteristic of this aqueous substance is the presence of a myriad of antimicrobial peptides (AMPs), lysozymes, lectins, proteases and other lytic enzymes that play essential roles in the protective function of mucus. The activities of these molecules are affected by various factors (e.g., stress, water temperature, pH, and infections); hence, the biochemical and physical characteristics of mucus have significant implications for the overall health of fish [
5].
The daily cycle of light and darkness has a striking influence on many organisms, including fish, as exhibited by roughly 24-h cycle in the physiological processes which can be collectively referred to as circadian rhythms [
6]. The ability of organisms to attune their physiology and behavior to these cyclic changes provides them with an adaptive advantage to ensure health and fitness. Immune responses have been shown in mammals to exhibit circadian rhythms [
7,
8]. The circadian control of the immune system is crucial for the homeostatic integrity of an organism. This two-way connection is manifested in inflammatory response, autoimmunity, septic shock, or chronic infection, amongst others [
8]. Even though there is limited evidence from fish of this interplay [
9,
10,
11,
12,
13], all earlier reports have revealed that many of the circulating immune molecules exhibit daily/circadian rhythms. We have previously shown that this is not only present in systemic immunity but also in mucosal immunity [
14]. Despite the considerable evidence from mammals, that mucus secretion and its associated biological functions have rhythmicity [
15,
16], this is not well-documented in fish.
One of the significant functions of immunological rhythms is in the temporal sensitivity and functionality of immunity. The ability of an organism to mount a response to a stimulus has been shown to be gated by the time of day. For example, responses to bacteria have been shown to vary depending on time of infection, and this has been attributed to the partitioning of the immune system into two states: (1) a state of anticipation and enhanced immune activity and (2) an opposing period that may relate to repair and regeneration of the immune system [
17]. We have previously shown that systemic immune response to a bacterial endotoxin challenge in Nile tilapia (
Oreochromis niloticus) exhibited a robust temporal sensitivity [
13]. It remains to be investigated whether such a distinct temporal gating is similarly present in the mucus.
In this study, we employed Nile tilapia (
O. niloticus) as a model to understand how secretory proteins previously identified to play vital functions in mucosal skin immunity in fish [
3,
5,
11,
13] respond to environmental photic changes such as rearing two groups under different photoperiod conditions. Moreover, we explored the dynamics of mucosal response following a time-dependent bacterial endotoxin challenge to evaluate whether the diel feature in these proteins has implications on the temporal sensitivity of mucosal immunity.
2. Materials and Methods
2.1. Ethics Statement
All fish handling procedures employed in the study were in accordance with national and EU legislation (2010/63/EU) on animal experimentation. All experiments were performed at the Technical University of Denmark (DTU Aqua) facilities in Hirtshals, Denmark.
2.2. Experimental Fish
All-male, sexually immature tilapia (O. niloticus) procured from a commercial supplier (Til-Aqua International, Someren, The Netherlands) were reared in a recirculating aquarium system with water temperature maintained at 25–27 °C. Fish were fed once daily to visual satiation with a commercial diet (EFICO Alpha 830F, BioMar, France) and approximately 20% of the water volume was replaced every day. Fish were maintained under a photoperiod of 24L:0D until onset of experiments.
2.3. Light Manipulation Experiment
The first experiment investigated the impacts of light conditions on innate immune factors in the skin mucus. Fish (average weight 82 ± 4 g) from the holding tank were transferred to six 189-L cylindrical–conical thermoplastic tank (10 fish per tank), which were connected to a different recirculation system. Water temperature was maintained at 28 °C, and dissolved oxygen saturation was above 90%. The photoperiod was set at an equal length of day and night (12L:12D, hereafter LD group). The fish were allowed to acclimate to the experimental unit for at least two weeks. Thereafter, the photoperiod of one group was changed to total darkness (0L:24D, hereafter DD group) while the other group was continued to be reared under the same photoperiod as in the acclimation stage. Each treatment group was represented with three replicate tanks. The average water surface intensity during the light phase in LD was approximately 350 lux (RS 180-7133 Light Meter, Northants, UK). A commercially formulated diet (EFICO Alpha 830F, BioMar, France) was delivered randomly through feeders connected to a timer at a daily ration corresponding to 3% of the body weight. The fish were maintained under these conditions for two weeks, similarly as in previous studies [
13,
14]. Feeding was synchronized 48 h before sample collection so that all fish did not have feed access 24 h prior to sampling. Mucus samples were collected at
Zeitgeber time 2 (ZT2, 2 h after lights on) and 14 (ZT14, 2 h after lights off). On the other hand, the same samples were collected at corresponding times in the group under DD (circadian time—CT, 2 and 14). Mucus was collected from a sedated (ethylene glycol; Merck, Darmstadt, Germany) fish as previously described [
14]. The collected mucus was immediately stored at −80 °C until extraction (
Section 2.5). Sampled fish were then transferred to a separate tank for recovery.
2.4. Time-Dependent Lipopolysaccharide Stimulation
It was hypothesized that the day–night variations in the skin mucosal defenses would affect the ability to respond to an immunological stimulus delivered in a time-dependent manner. Samples used in this part of the study were collected from a previously published trial [
3]. Briefly, tilapia that were entrained under a 12L:12D photoperiod were challenged intraperitoneally with lipopolysaccharide (LPS) from
Escherichia coli 0111:B4 (Sigma, Steinheim, Germany) suspended in phosphate-buffered saline (PBS). There were two challenge strategies employed: one group of fish was injected with LPS at ZT3 to represent the response during daytime, while the other group was challenged during nighttime at ZT15. For each challenge occasion, an additional group of fish was injected with PBS to serve as a sham-control. Skin mucus was collected as described earlier [
14] from six fish (two from each tank) at 2, 4 and 8 h post-injection (hpi).
2.5. Secretory Proteins in the Skin Mucus
Skin mucus extract was prepared by re-suspending the collected sample in one volume of chilled sterile PBS (pH = 7.4) followed by vigorous shaking. After centrifugation at 10,000× g for 10 min at 4 °C (SIGMA 3-18K, Osterode, Germany), the supernatant was collected, aliquoted and kept at −80 °C until analysis.
Alkaline phosphatase activity was determined in a 20 min kinetic reaction assay using
p-nitrophenyl phosphate as a substrate [
18]. The enzyme activity was measured by the initial rate of reaction and one unit of activity was defined as the amount of enzyme required to release 1 mmol of
p-nitrophenol product in 1 min. The anti-proteolytic activity was measured by quantifying the inhibition potential against trypsin activity [
19]. After stopping the 2 h reaction between the mucus extract and the substrate azocasein, the optical density was read at 450 nm, and the percentage of inhibition was calculated by comparing the values to 100% control. Mucus ceruloplasmin activity was measured in an enzyme reaction mixture containing
para-phenylenediamine-sodium azide in acetate buffer as a substrate. The kinetic increase of absorbance was followed for 15 min and one unit was defined as an increase of OD of 0.001 min
−1 at 550 nm [
20]. Turbidimetric method using
Micrococcus luteus as a substrate was used to measure the activity of skin mucus lysozyme [
21], following a protocol adapted for a 96-well microplate [
22]. Lysozyme activity was defined as the amount of enzyme that caused a decrease in absorbance of 0.001 per minute at 450 nm. Peroxidase was measured following previously described protocol [
23], with modifications [
14], using 3,30,5,50-tetramethyl benzidine hydrochloride as a reaction substrate. The activity was expressed as absorbance unit at optical density (OD) 450 nm. All OD measurements were conducted using a microplate reader (TECAN GENios, Salzburg, Austria).
2.6. Statistics
All statistical analyses were carried out in SigmaStat (SystatSoftware, London, UK). The difference in the levels of defense molecules between the two sampling points in the photoperiod manipulation experiment was determined by Student’s t-test for independent samples. The same statistical test was used to identify differences in the activity/level of a defense molecule at a specific period between CT and ZT, and to evaluate differences in the response between LPS-injected and PBS-injected groups at a particular time point. The activities of defense molecules post-challenge were subjected to a one-way ANOVA. For data sets that complied the tests of normality and equal variance, and Tukey’s multiple comparison test was employed to identify differences between time points. For non-parametric data sets, Kruskal–Wallis one-way ANOVA on ranks followed by Dunn’s multiple comparison test was used instead. In all statistical calculations, the level of significance was set at P < 0.05.
3. Results and Discussion
Environmental light conditions have significant impacts on fish physiology. For example, photic signals mediate several critical behavioral and physiological processes in fish, including swimming, feeding, maturation, and spawning [
24,
25,
26,
27]. Although the influence of photic environmental cues on immunity is explored to some extent in fish [
9], the current understanding is incomplete. Also, most of the reported studies were on the impact of photoperiodic manipulations on systemic immunity, and research on the mucosa is scant. Here we describe how mucosal defenses responded to two different photic environments and how these secretory defense proteins were mobilised when prompted with a stimulus at different time of day.
Fish skin mucus contains a plethora of bioactive molecules and many of which play an essential function in ensuring a robust barrier between the epithelial layer and the water matrix. The presence of extracellular plasma proteins in the skin mucus of fish may be described in several mechanistic scenarios [
5]. First, mucosal epidermal cells secrete these proteins. Secondly, their presence may be a result of a leakage from circulating plasma to the mucus. Another mechanism is a phenomenon called “shuttling effect” wherein they are delivered to the mucus via the secondary circulatory system that connects the vascular system, blood and mucus [
28,
29]. In an earlier study in the permit (
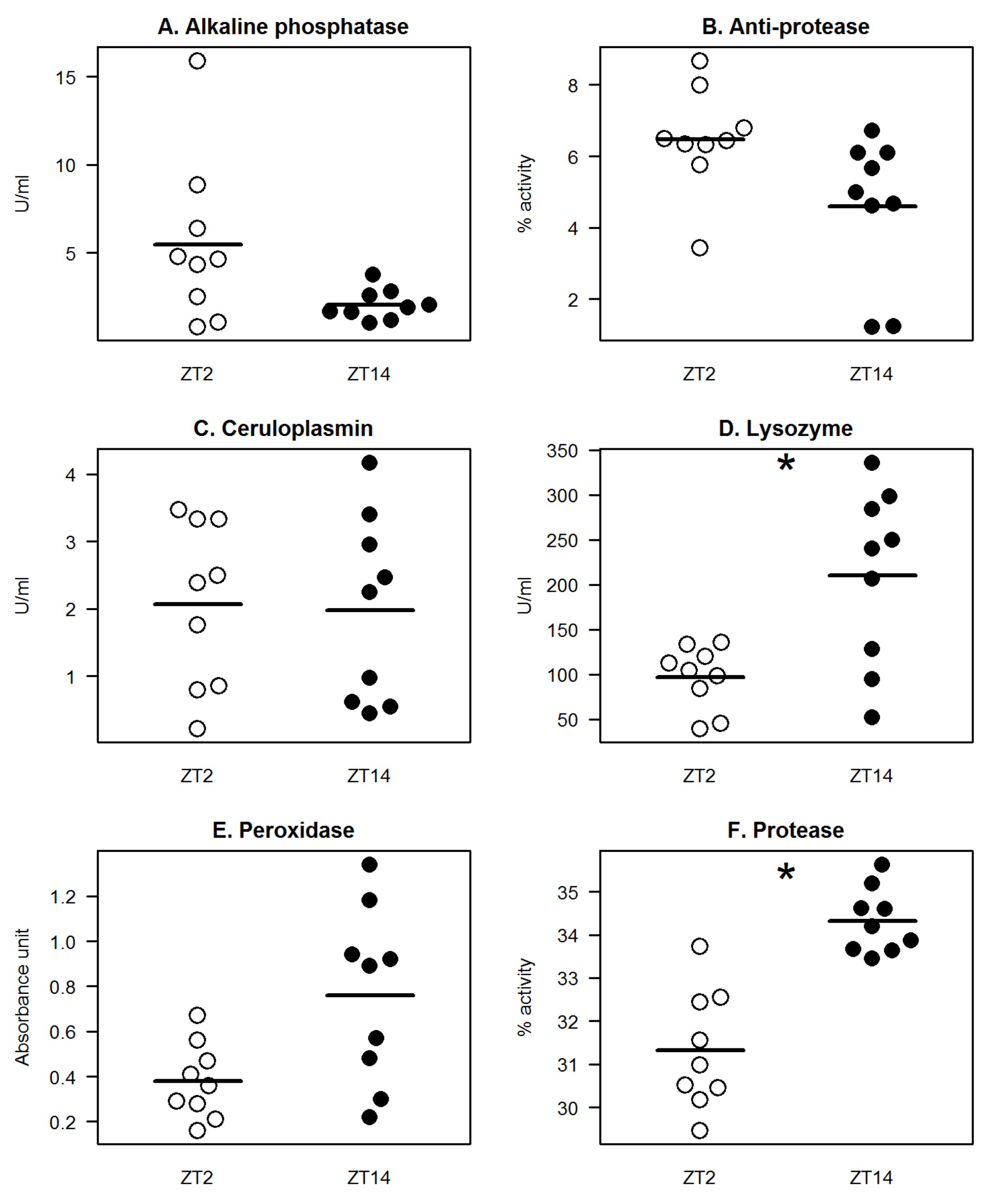
Trachinotus falcatus), it has been shown that immune molecules in the skin mucus including anti-protease, lysozyme, alkaline phosphatase, esterase, glutathione peroxidase, catalase, and superoxide dismutase demonstrated day–night profiles; many of which exhibited elevated activity/level during daytime. In the current study, we have identified that mucus lysozyme and protease exhibited significant diel variations, where the levels/activities of these proteins at night were higher compared to during the day (
Figure 1). Lysozyme activity at night (ZT14) was one-fold higher than during the day (ZT2;
Figure 1D). On the other hand, protease activity was higher by at least 8% at ZT14 compared with the recorded value at ZT2 (
Figure 1F). Serum lysozyme in tilapia has been shown earlier to be more active during daytime [
13]. Mucosal and circulating lysozyme molecules might be regulated differently in response to light-dark changes, although we cannot ascertain the origin of the quantified mucus lysozyme. Another possible scenario was that a delay occurs in the relaying response between mucus (night-biased activity) and serum (day-biased activity), or vice versa, hence the occurrence of contrasting profiles. Interestingly, both serum and mucosal protease in tilapia displayed a similar night-biased activity, indicating a potential stringent regulation of both the systemic and mucosal protease, which is worth exploring in future studies. Peroxidase showed an apparent nocturnal activity pattern; however, the difference between the daytime and night-time level was not statistically significant. Alkaline phosphatase and anti-protease activities seemed to exhibit a more day-biased pattern, though the differences between the two time-points were not statistically significant. Mucus ceruloplasmin level was almost identical between daytime and nighttime.
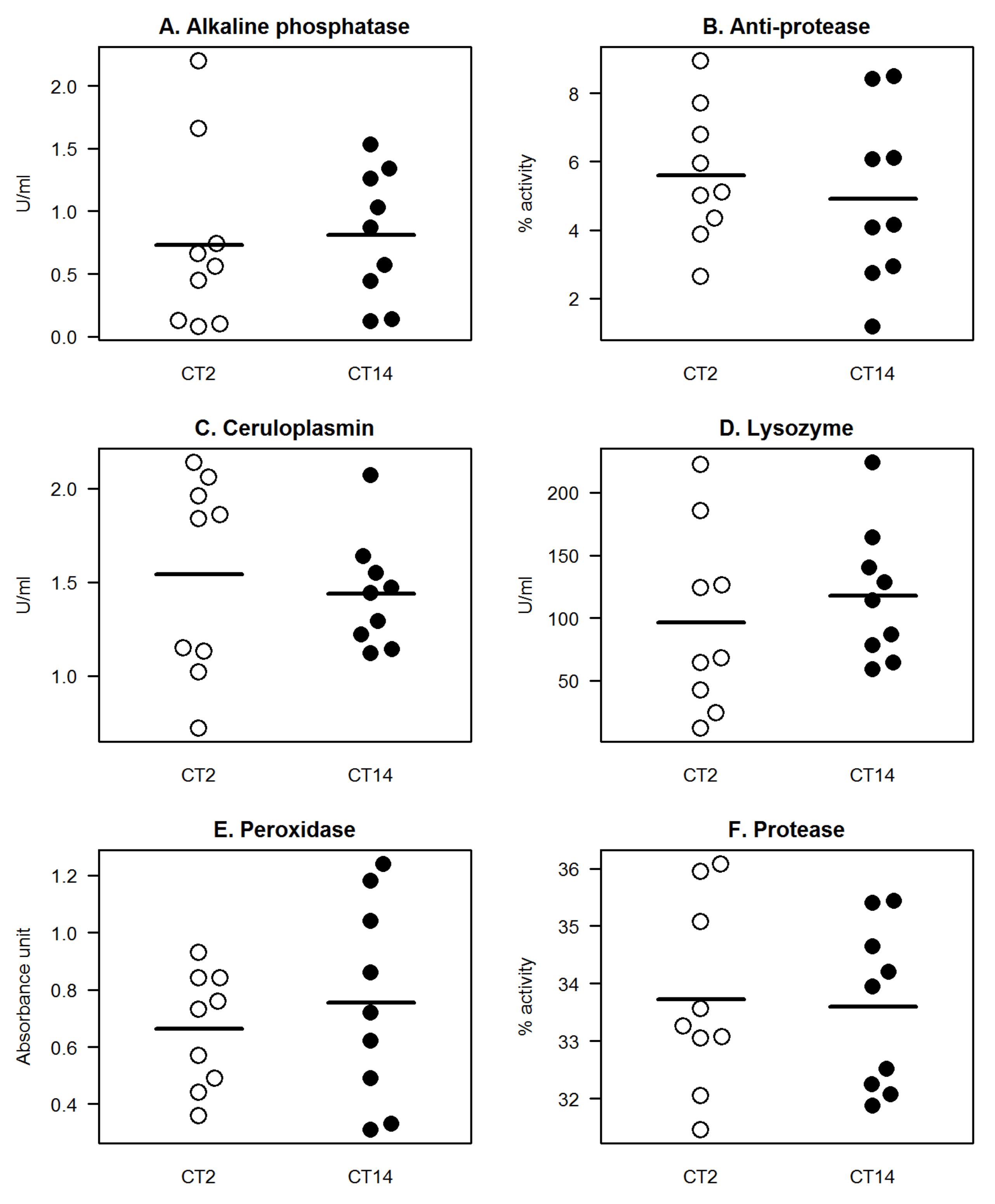
We then asked whether the diel variability could persist even under constant darkness or the absence of an environmental LD signal. Therefore, we reared tilapia under total darkness with all other environmental parameters unchanged. None of the studied parameters exhibited significant differences in their levels/activities between CT2 (time equivalent of ZT2) and CT14 (time equivalent of ZT14), with almost identical patterns between the two time-points (
Figure 2). Notably, for lysozyme and protease, that showed distinct day-night differences under LD, such a temporal pattern was absent under DD condition. In a companion study, a similar tendency of loss of diel variability was observed in the serum defense parameters in tilapia when cultured under DD and when the photoperiod was changed from LD to DD [
13]. These corroborating results reveal that environmental photic conditions have a substantial influence on both systemic and mucosal defenses in tilapia and further indicates that the absence of an LD signal interferes with the daily variability of immune defenses [
9,
11,
12,
13,
14]. It has been suggested earlier that LD signal ensures the fidelity of how defenses are mobilised during the daily cycle and the diel variations exhibited by these molecules allow organisms to mount a concerted response that is temporally gated. It is still not clear how mucosal defenses are influenced by the internal time-keeping system. Further, it is yet to be investigated whether there is a local or a systemic control in the temporal order of mucosal defenses in fish, specifically in tilapia.
We compared the level of a defense molecule between the same ZT and CT and found some striking differences (
Figure 3). For alkaline phosphatase, the level was significantly higher at ZT2 than at CT2 whereas for lysozyme, the activity was significantly higher at ZT14 than at CT14. For peroxidase and protease, the activities were significantly higher at CT2 than at ZT2 (
Figure 3E,F). Though there was no apparent overall pattern in the comparative response between ZT and CT, it is worth noting that photic cues could influence not only the level of these defense molecules between day and night but also at a particular period in a day. We can speculate that the differences may be a form of an adaptive response to photoperiodic changes, and different defense proteins exhibit varying photic plasticity profiles.
One of the important implications of the daily variations in immunity is the ability of an organism to mount different responses to a stimulus, depending on the time of day. Temporal sensitivity of immunity has a significant function to host health and survival, and such a relationship has been documented in mammals [
7,
17] and in fish [
12,
13,
30]. We have previously shown that the time of day had a marked influence on how tilapia mobilised their circulating immune molecules in response to a bacterial endotoxin. Tilapia were able to mount stronger systemic immune responses when they were exposed to the endotoxin at daytime [
13]. In the present study, we investigated whether such a temporal sensitivity could likewise be manifested as a secondary peripheral response in the skin mucus. The post-stimulation profiles did not reveal a similar striking response to endotoxin, as in the previous study [
13]. Nonetheless, a number of significant responses to bacterial endotoxin were observed during the night (
Figure 4D,F). The levels of mucus lysozyme and protease were significantly elevated 4 h after endotoxin challenge at night. Interestingly, both proteins displayed significantly elevated levels at this period of the LD cycle (
Figure 1). The distinctive nocturnal profile substantiates, at least in part, as to why significant responses from these molecules were triggered more actively at night. It could be speculated further that these two molecules play an essential role in the nocturnal mucosal defense of tilapia. Ceruloplasmin, an acute phase ferroxidase protein, is activated when an organism encounters stressful events [
31,
32]. Ceruloplasmin was the sole protein in the panel to display an elevated level when the endotoxin was administered at daytime (
Figure 4C). The level increased twice in LPS-exposed fish compared with the PBS-control group at 8 h post-injection. Ceruloplasmin has been shown as well to be involved during photic injury [
33]. We can speculate that the increased in ceruloplasmin level may be attributed to a potential compounding effect of LPS and light, hence the stronger response at daytime, even though mucus ceruloplasmin did not exhibit diel differences (
Figure 1).



{kind=link}
{kind=link}
{kind=link}
{kind=link}



