Life History and Population Dynamics of Green Crabs (Carcinus maenas)


1
Biology Department, Salem State University, Salem, MA 01970, USA
2
New England Biolabs, Ipswich, MA 01938, USA
*
Author to whom correspondence should be addressed.
Fishes 2020, 5(1), 4; https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5010004
Submission received: 25 November 2019
/
Revised: 20 December 2019
/
Accepted: 23 December 2019
/
Published: 31 December 2019
Abstract
:Carcinus maenas (the “shore crab” or “European green crab”) is a very proficient invader (considered to be one of the world’s 100 worst invaders by the IUCN) due to its phenotypic plasticity, wide temperature and salinity tolerance, and an extensive omnivorous diet. Native to Atlantic Europe, it has established two well-studied nonindigenous populations in the northwestern Atlantic and northeastern Pacific and less-studied populations in Australia, Argentina and South Africa. Green crabs are eurythermal and euryhaline as adults, but they are limited to temperate coastlines due to more restrictive temperature requirements for breeding and larval development. They cannot tolerate wave-swept open shores so are found in wave-protected sheltered bays, estuaries and harbors. Carcinus maenas has been the subject of numerous papers, with over 1000 published in the past decade. This review provides an up-to-date account of the current published information on the life history and population dynamics of this very important species, including genetic differentiation, habitat preferences, physical parameter tolerances, reproduction and larval development, sizes of crabs, densities of populations, sex ratios, ecosystem dynamics and ecological impacts in the various established global populations of green crabs.
1. Introduction
1.1. Overview
The first extensive examination of C. maenas was J.H. Crothers’ “The Biology of the Shore Crab Carcinus maenas” Parts 1 and 2 published in 1967 and 1968 respectively [1,2]. At that time very few studies had been conducted on C. maenas [3,4,5,6,7] so much of Crothers’ paper presents original information. Many research studies followed but it was several decades before Sylvia Behrens Yamada published her review book “Global Invader: The European Green Crab” in 2001 [8], followed soon after by “A Biological Synopsis of the European Green Crab, Carcinus maenas” by Greg Klassen and Andrea Locke in 2007 [9]. The present review is an attempt to assimilate previous data with studies that have been published more recently, most notably involving the nonindigenous populations and the variable methods and conclusions relating to population genetics as a result of advancements in phylogenetic methods.
Due to the ease with which it can be collected, identified, sexed and measured, there have been many studies of C. maenas, but most have concentrated on laboratory-based experiments and observations of physiological and biological processes. There remains considerable lack of consensus about many of the species’ ecological attributes, especially among populations from different geographical regions. This is due at least in part, if not mostly, to the extensive phenotypic plasticity of the species. Carcinus maenas is able to alter many aspects of its biology to fit particular environments, such flexibility and adaptability making it an ideal invader. In addition, there are evolutionary differences in physiology among the various source populations such as from northern Europe or south-central European regions. It has not yet been determined if physiological differences in crabs from different areas is correlated with the three different genetic lineages in the native range (northern Europe, western Europe, and off-shelf of the Faroe Islands and Iceland) [10,11]. Genome-wide studies are exposing the complexity of the genetic background of this species [12,13,14,15,16,17,18]. Whenever possible this review will provide the consensus view on a particular topic, but in most cases there is so much variability in published data that no consensus is possible and we will instead present a variety of results from different studies.
Of the numerous studies on C. maenas (a literature search conducted by Darling 2011 revealed 1393 papers published in the first decade of the new millennium) the vast majority have involved three main populations—the native population in Europe and the two prominent nonindigenous populations in the northwestern Atlantic and northeastern Pacific. Studies of C. maenas populations in the native European population span most of the range but we are not aware of any published reports from Iceland aside from inclusion of some crabs from this area in genetics studies or from the northwestern Africa region of the native range other than a sample of 40 crabs from Morocco analyzed for DNA [13]. Most of the northwestern Atlantic studies have been conducted on Atlantic Canada and New England populations, with data from south of those regions included in only a few publications. In both the native and northwestern Atlantic populations, the numbers of crabs in the southern part of the ranges are greatly reduced, making collecting sufficient specimens for study difficult. Research in the northeastern Pacific has involved most of the range, including studies from British Columbia, Washington, Oregon and California. There are relatively few studies on the established nonindigenous populations in South Africa, Argentina or Australia. This review examines published papers by various researchers studying C. maenas in the native European and northwestern Atlantic and northeastern Pacific nonindigenous populations, where relevant studies from each major population will be discussed in appropriate sections to follow, with occasional mention of data from other populations when available and appropriate.
1.2. Native European Population
The native range of Carcinus maenas (where it is commonly known as the “shore crab”) is the northeast Atlantic coastline, from Mauritania in northern Africa to Norway, the British Isles and Iceland [2,10,11,19,20]. It is the most common intertidal decapod crustacean in Europe. In its native region, C. maenas is considered to be an important and profitable species, commercially fished for use as bait or food [21,22]. A decade ago the biology and population dynamics of C. maenas in the UK was reviewed in response to concerns that its abundance was declining due to overfishing and that a maintenance program might be needed to insure sustainability [22]. In contrast, in other parts of the globe C. maenas is thought of as a very destructive invasive species and is considered to be one of the world’s 100 worst invaders by the International Union for the Conservation of Nature (IUCN) [9,23,24,25,26,27,28]. It was the first marine organism to be designated as an aquatic nuisance species by the Aquatic Nuisance Species Task Force (ANSTF) [29]. The Latin name “Carcinus maenas” translates as “raving mad crab” and is known as “le crab enragé” in France. The common translation of maenas from Latin to English as “sprats” is not accurate. In Greek mythology, ‘maenads’ were the female followers of Dionysus and their name literally translates to “the raving ones” because of their frenzied state associated with rituals involving wild dancing and copious amounts of wine. Later, in Roman mythology the Latin feminine noun ‘maenas’ was used, meaning ‘frenzied or raging woman.’ Perhaps it is this meaning that Linnaeus had in mind when he named the crab Carcinus maenas (P. Sean McDonald, U. Washington, personal communication). There are invasive populations in various locations throughout the world, where the species is referred to as the “European green crab,” just “green crab” or sometimes “European shore crab” [20,30,31,32].
1.3. Northwestern Atlantic Nonindigenous Population
Carcinus maenas was first discovered along the mid-Atlantic United States (location not given but assumed to be New Jersey or New York) in 1817, presumably carried from Europe in solid ballast or on the outside of wooden ship hulls, [10,20,33,34,35,36,37]. This part of the range now extends over approximately 1000–1200 kilometers of coastline [28,38,39]. The species spread southward to Chincoteague Bay, Virginia, with occasional sightings of individuals further south, but further southward expansion of the population apparently is limited by both temperature and the predatory blue crab Callinectes sapidus [40]. However, C. maenas has expanded mostly northward, arriving in northeastern Massachusetts, New Hampshire, and southern Maine waters in the 1890s or early 1900s, and eventually reaching southern Nova Scotia, Canada by the early 1950s, Prince Edward Island by 1997, and Newfoundland by 2007 [8,9,20,31,32,41,42,43,44]. The species’ expansion northward from southern Nova Scotia was found to be associated with a second introduction, from northern Europe, probably Norway, around 1980, rather than being a northward expansion of the original introduction in the early 1800’s that was derived from a source in south-central Europe [10,17,18,27,45,46,47]. A model of the invasion dynamics predicted that the two lineages would overlap only in the neighborhood of the invasion fronts, a region approximately 100–200 km wide [48] but in fact the original and later introductions have extensively interbred, leading to genetic admixture throughout much of the species’ northwestern Atlantic range [10,14,17,45,46,49].
1.4. Northeastern Pacific Nonindigenous Population
In 1989–90 a well-established breeding population of green crabs was discovered on the northeastern Pacific coast, in San Francisco Bay in California [36]. Populations have become successfully established only 125 km to the south in Elkhorn Slough in the Monterey Bay area, but have spread over 1500 km northward in just 12 years, reaching the west coast of Vancouver Island, British Columbia, Canada by 1999 [31,50,51,52]. In Oregon and Washington estuaries, C. maenas continues to be found in low densities but the crabs are abundant in the inlets of the west coast of Vancouver Island [53]. More recently, crabs have been found in the Salish Sea (Sooke Basin in 2012, Westcott Bay and Padilla Bay in 2016), and Puget Sound (Dungeness Spit, Sequim and Whidbey Island in 2017) [53,54]. There have been occasional sightings of individual crabs elsewhere in the Puget Sound but they are rare and probably do not represent breeding populations [53,54]. The northeastern Pacific population is genetically depauperate and represents a secondary introduction from the initial northwestern Atlantic introduced population [10,12,55]. The likely transmission route was juvenile or adult crabs transported in seaweed packed around baitworm shipments and live lobsters shipped from New England to California restaurants [36,56,57], with subsequent range expansion by larval transport via ocean currents, especially after warm El Nino winters [50,51,53,58,59]. Movement into the Salish Sea has been slow due to two physical barriers: estuarine circulation normally moves water out of the Sea and water temperatures are typically less than the 10 °C lower thermal tolerance for larval development. Only during periods of El Nino does the current flow reverse and the temperature rise above 10 °C [53]. Several sites are areas of oyster aquaculture so it is possible that shellfish transplants may play a role in some range expansion [51,53]. By whatever means, further expansion northward along the northeastern Pacific coast is thought to be inevitable [59,60,61,62].
1.5. Other Nonindigenous Populations
The northwestern Atlantic and northeastern Pacific populations are the most-studied invasive populations. Additional secondary and tertiary introduced populations are now located in South Africa, Australia and Argentina [31,63]. Green crab populations are established in South Africa between Saldana Bay to the north and Camps Bay to the south after being introduced in 1983 to the Table Bay Docks area in Cape Town, likely from northern Europe via ballast water [64,65,66,67,68]. In Australia the population extends from near Sydney in New South Wales south to Port Phillip Bay, Victoria; Bruny Island, Tasmania and the Coorong, South Australia, likely introduced into Port Phillip Bay, Victoria, from Atlantic Europe in the late 1800’s via solid ballast and hull fouling [69,70,71,72,73,74,75,76,77,78,79,80]. Occurrence outside its normal range in Tasmania is often in association with aquaculture activities [76]. There is a population in Atlantic Patagonia, Argentina, from Camarones Bay north to San Jorge Gulf, introduced from Australia or Tasmania in 1999 via ballast water [81,82,83]. Although there have been reports of an established population of C. maenas in Japan, the Japanese population apparently represents hybridization between C. maenas and its congener in the Mediterranean, C. aestuarii (=C. mediterraneus) [8,10,27,63,76,84] and will not be included in this review.
1.6. Additional Introductions and Potential Invasion Sites
There are established populations of green crabs on every continent except Antarctica but introductions into tropical regions including Brazil, Panama, Hawaii, the Red Sea, Sri Lanka, Madagascar, Union of Myanmar, Pakistan and India have not resulted in established populations [9,20] because females require temperatures at or below 18 °C to successfully incubate their eggs [1,85]. Based on temperature and salinity information from both native and established invasive ranges of C. maenas, one study identified several new sites at risk of invasion: coastal areas of Chile, China, Russia, Northern Japan, the Yellow Sea, Alaska, Namibia, Uruguay, southern Brazil, and New Zealand [86] (see Figure 1). Perhaps all that is needed is a means of introduction into these new environments, especially of a lineage, possibly one from northern Europe, that is adapted for the conditions, such as colder waters, in these new regions [86] (Carlton [87] discusses the many possible vectors).
2. Population Genetics
One of the newest research frontiers is the advancement in the use of genetic markers to elucidate population affinities and differences. An early goal of using molecular methods in the study of green crabs was to provide insight into the differences within the native range between C. maenas along the Atlantic coast and C. aestuari (formerly C. mediterraneus) in the Mediterranean Sea, differences that had been well characterized morphologically [8], and genetically [27]. Over the years, Carcinus populations have been studied with an evolving suite of genetic markers including allozymes [88], mitochondrial 16S [89] and cytochrome c oxidase I (COI) genes [11,46,90], microsatellite markers [10,12,14,63,91,92], and SNPs [15,16,18,49]. Increasingly high resolution studies of population genetics have led to a greater understanding of intraspecific and interspecies diversity. For example, an initial analysis of 19 allozyme loci led to the conclusion that the genetic differentiation between C. maenas and C. aestuarii was insufficient to warrant independent species status and a designation of two subspecies of C. maenas was recommended [88]. However, later work identified species-diagnostic differences in partial sequences of the 16S small subunit rRNA gene [89], supporting the now accepted idea that C. maenas and C. aestuarii indeed are separate species, although they can hybridize.
A more comprehensive study based on the mitochondrial cytochrome c oxidase subunit I (COI) gene revealed considerable divergence between northwestern Atlantic populations and a single Mediterranean population, confirming a species-level taxonomic distinction, as well as significant intraspecific differentiation between populations from Iceland and the Faeroe Islands and those from continental Europe [11]. Within continental Europe, the COI data supported weak but significant differentiation between crabs from northern Europe (Norway, Sweden, Finland, Denmark, Germany, and Poland) and those from southwestern Europe (the Netherlands, Belgium, France, Spain and Portugal [11]. This divide is not clear-cut, and the United Kingdom population contains a mix of haplotypes from both the northern Europe and southwestern Europe groups [93]. Using microsatellite markers in the native European population, C. maenas from Faeroe Islands and Iceland were found to be genetically differentiated from continental populations when using microsatellite markers, and like earlier mitochondrial markers, also showed a significant partitioning of genetic variance between northern Europe (including Germany and Scandinavian populations) and western Europe (including the Netherlands, England, and Atlantic Iberian populations) [10].
Along the northeastern Pacific coast, microsatellite markers were used to assess the genetic diversity in 1040 C. maenas samples from 21 sites representing the major episodes of population establishment and expansion in that region [12]. It was concluded that C. maenas in this population likely derived from a single introduction of a small number of founders to San Francisco Bay, California, from the western Atlantic, supporting an early conjecture based on observations of crabs in live seafood shipments arriving in the western US from the northeastern US [36]. The use of both mitochondrial and nuclear markers in reconstructing invasion pathways for Carcinus serves as a valuable illustration of the utility of multiple genetic markers and multiple analytical methods in genetic studies of biological invasions [27].
An updated genetic analysis of seven populations from the native European, northwestern Atlantic, and northeastern Pacific ranges, using a panel of 10,809 transcriptome-derived nuclear SNPs, identified significant population structure that was previously undifferentiated with earlier markers [15]. This SNP panel represented a considerably larger portion of the genome than previous approaches, covering at least 1673 regions, and identified both neutral and non-neutral mutations in the SNP panel in an attempt to predict what genes may support a successful invasion. By all measures, genetic diversity was highest in the European native range and lowest on the northeastern Pacific coast [15], consistent with serial bottlenecking caused by introduction from Europe to the US east coast, and from the east coast to the US west coast. The data revealed significant genetic structure between all locations, even between invasive populations that appeared undifferentiated when using mitochondrial DNA and microsatellites [10].
There have been several attempts to reconstruct introduction histories for invasive Carcinus populations using population genetics [27] (see “Introduction” section). The first introduction of green crabs outside of Europe was to the northwestern Atlantic in the early nineteenth century [36], where they spread northwards along the coastline, eventually reaching a stable northern range limit near Halifax, Nova Scotia by the 1970s [32]. In the late 1980s, a second introduction of green crabs from northern Europe to northern Nova Scotia occurred in concert with a rapid expansion of the species up the coast, where it achieved high densities by the 2000s [45]. This second invasion in the northwestern Atlantic was derived from a native northern European source that was genetically distinct from the earlier invasion [45].
The introgression cline of east coast US has been most extensively studied with a number of markers being used. The first such investigation used only the COI marker, and reported a strong genetic cline along the Scotian Shelf with a prominent Mid-Atlantic haplotype that diminished in frequency from south to north [45]. The structure of the cline was attributed to differentiation through selection in the northern part of the eastern US range and/or contact of two invasion fronts through a more recent invasion of new genetic lineages in Nova Scotia [45]. The aforementioned microsatellite markers [92] yielded a slightly different result. Although analyses of both mitochondrial and nuclear genetic loci indicated admixture between the two invasion fronts, two marker systems disagreed in the geographic extent of the admixture zones [46]. While COI data suggested that genetic introgression from the more recently introduced northern C. maenas population has extended well into the western Bay of Fundy [45], microsatellite data show very little evidence of admixture at northern Bay of Fundy sites, and introgression of northeastern Nova Scotian alleles is low even at sites along the southern Bay of Fundy [46]. Still, vessel traffic and shipping were considered the most likely vector candidates for the Newfoundland introduction [46], agreeing with previous conclusions [45].
Over time, it was predicted that continued patterns of dispersal and connectivity would result in a persistent southward shift [90], however this would later be found to be incorrect with an advance in genetic markers [49]. The original hypothesis was that larval dispersal alone could not explain the displacement of C. maenas clines, where introgression occurred more rapidly for mitochondrial DNA relative to nuclear markers, suggesting demographics (e.g., gene surfing) or natural selection (e.g., thermal tolerance) may be influencing invasion dynamics [14,45]. Although an analysis of SNPs revealed that there was not a southward shift in genetic cline as previously predicted [90] both neutral and selective processes were attributed as drivers of introgression and all marker types resolved a common spatial genetic pattern, where current clines for COI, microsatellites, and SNP allele frequencies overlapped in cline center and width [16,18,49]. The application of multiple genetic markers to elucidate invasion history is well supported from the study of the introgression cline of the eastern US coast, as the understanding of green crab invasion dynamics has changed with increasingly high-resolution markers.
Molecular genetics methods offer new insight into the population dynamics of C. maenas, and have proved to be a very useful tool in understanding genetic lineages in many marine invasive species. With advancements and improvements in the extent of the genome that can be characterized using high-throughput sequencing, further genomic research may help to elucidate the species’ invasion history and, most importantly, shed light on the species’ potential future expansion.
3. Habitat Preferences
Carcinus maenas is able to occupy a variety of sheltered habitats such as estuaries, harbors, and other wave-protected areas but are not able to survive on open shore wave swept coastlines because they lack sufficient vertical tenacity and ability to grip onto rocky substrates [8,83,94]. Megalopae settle out of the plankton in protected areas of the intertidal zone. Juvenile crabs prefer gravel or cobble areas or mussel beds where they can shelter under rocks and seaweed to avoid predation and cannibalism [8,95,96,97,98,99,100,101,102,103,104,105,106]. Juvenile crabs frequently display striking colors and mottling patterns that may help to camouflage them in geometrically complex habitats with variable substrates [107,108]. As crabs age they move into lower intertidal and subtidal zones, where they are able to shelter under rocks and boulders and macroalgae in rocky areas and amidst eelgrass and fouling communities or in burrows along Spartina banks in estuaries. Both juveniles and adults prefer areas with high structural complexity [109,110,111] and avoid open sandy areas that do not offer any refuge, although adults are common in muddy sands where they can burrow for concealment [112,113,114]. Mussel beds on mudflats offer optimal refuge from predators such as large crabs, fishes and birds [109]. In an experiment conducted on the Swedish west coast predation mortality of tethered crabs was 80–90% in open sand habitats and much lower in mussel beds and vegetated areas that provided shelter [101]. Adult C. maenas are very tolerant of low oxygen levels as well as temperature and salinity variations (see “Tolerances” below) so are able to remain in moist burrows above the neap high tide line for up to 10 days until water returns with the next spring high tide [8]. Adult crabs forage over extensive areas of their habitat, travelling as much as 2 km in 6 h [115], but do not return to a home shelter after excursions [116].
Adult C. maenas can be found to depths of 10 m or greater [117,118,119] but they are most commonly found at depths not exceeding 6–7 m and their numbers diminish greatly below this depth [2,5,9,75,113,120,121,122,123,124]. In some locations they migrate into deeper water during the cold winter months but in other areas many appear to remain in the shallow subtidal throughout the year, although numbers are reduced [113,125]. Following a colder than normal winter the density of the population is reduced [113,126,127] and many crabs can be found dead inside their Spartina bank burrows (M. Fregeau, Salem State U., pers com), evidence that they did not migrate. During the summer months females tend to be found in higher salinity subtidal areas except when they congregate in shallow water to mate.
4. Tolerances
4.1. Temperature
Carcinus maenas can tolerate a wide range of environmental conditions, due at least in part to its phenotypic plasticity, whereby it can alter its physiology and other responses to match local conditions [128]. For example, C. maenas is eurythermal, able to survive short-term exposure to temperatures between 0 and 33–35 °C [3,8,9,96,126,129]. Adult C. maenas exposed to air on mud flats at low tide are able to evaporatively cool their bodies by as much as 2 °C, making them more resistant to high temperature than a subtidal relative, Portunus marmoreus [130]. A critical thermal maximum (CTMax) of 35.1 °C was estimated for crabs collected in Portugal [131] and of 35.8 °C for North Sea crabs caught in the fall and acclimated to 22 °C [132]. Crabs from California near the southern part of the northeastern Pacific range had higher thermal tolerance (36.2 °C) than those near the northern limit in British Columbia (34.7 °C) [39]. Green crabs, like many species that inhabit variable hot environments such as estuaries and intertidal zones, have high levels of heat shock proteins (e.g., HSP70) that are important components in the cellular defense against proteotoxic stress [131]. The abundance of green crabs has declined after harsh winters in their native range [98,133,134] as well as in nonindigenous populations [113,126,135,136]. Adult C. maenas from British Columbia survived an 18-week duration period in the lab at temperatures at or below 5 °C [137]. The maximum temperature tolerated for breeding is 26 °C [138] and for brooding eggs, 18 °C [8], restricting sustainable populations to sub-polar and temperate coasts in both the northern and southern hemispheres where temperatures range from −1 °C to 22 °C [20,36,39]. There are no populations of C. maenas in tropical or sub-tropical environments, despite introductions in many such locations [20]. This likely explains why nonindigenous populations have expanded only slightly southward but much more extensively northward from introduction sites.
The minimum temperature for growth is 10 °C [8,139]. The lower temperature limit for feeding is generally considered to be 7 °C [8], although Ropes [112] concludes that at temperatures as low as 7 °C crabs from Massachusetts do not suppress feeding but activity and presumably feeding probably cease at some temperature below 7 °C. In most populations, adult crabs migrate offshore when the water temperature drops below 8 °C in winter and then return as the water warms in the spring [2,4,5,125,140,141,142,143,144,145,146]. Audet et al. [147] report that C. maenas in Prince Edward Island waters may feed at temperatures below 6 °C but only in the fall, not in the spring, and Eriksson et al. [96] indicates that feeding may occur below 7 °C but almost ceases below 3–4 °C for crabs in Sweden [96]. The lower temperature limits reported for the Sweden population may represent the more cold-tolerant Scandinavian lineage identified by Roman [45] and introduced to northern Nova Scotia. It is quite possible that it is this lineage that is represented in the PEI population studied by Audet et al. [147] and is presumed to be more cold-tolerant than the southern European lineage found in the nonindigenous populations in the northwestern Atlantic south of Nova Scotia and in the northeastern Pacific region. To date no tolerance studies have been conducted with C. maenas from northern Nova Scotia and Newfoundland to determine if they are indeed more cold-tolerant than their southern relatives.
Larvae exhibit higher physiological tolerance to temperature fluctuations than adults [85,148] and juveniles are more tolerant of temperature variations than older individuals [28,107,149,150]. deRivera et al. [85] found that mortality of cultured larvae at temperatures below 12.5 °C was very high due to extended exposure to cold. Development time is temperature dependent in that it takes 62 days at 12 °C and only 32 days at 18 °C for a crab to complete its early development to where it is considered an adult [151].
4.2. Salinity
Carcinus maenas is also euryhaline, with the long-term adult survival minimum salinity variously reported as 11‰ [8], 9‰ [152], 5‰ [96,140] or 4‰ [3]. At high salinity the crab is an osmotic conformer but becomes an osmotic regulator at a critical low salinity [153]. It is important to consider the synergistic effect of temperature and salinity in combination because stress due to sub-optimal values of one variable can result in diminished survival at sub-optimal values of other variables. Broekhuysen [3] found that in water of about 15‰ C. maenas died at a temperature of 0.6 °C in about thirty days but at a temperature of 9.7 °C no deaths occurred. Perkins et al. [154] reported that C. maenas were commonly found in salinities of 1.4–3.2‰ at 8.0–11.0 °C for periods of at least 8 h and, while crabs maintained at a temperature of 6.6–10.0 °C died rapidly in salinities of 1–2‰, they survived for at least 34 days at salinities at or above 3‰. Carcinus maenas does not acclimate rapidly to reduce salinity and still exhibits an increase in oxygen consumption after 3–4 days of immersion [155]. Newly molted crabs are green in color but as an intermolt period progresses the color transitions through yellow and orange to red. Green phase crabs are known to be more tolerant of low salinity than red phase crabs and males are generally considered to be more tolerant than females [140,146,156,157,158,159] although Himes et al. [125] found that females in a Maine population were more tolerant than males. These differences in tolerance to low salinity as well as to oscillating salinities result from greater expression of the osmoregulatory genes behind ion transport (Na+/K+-ATPase, cytoplasmic carbonic anhydrase) in green phase C. maenas, making the green morphotype more tolerant than red [125]. In the native range, green crabs are more common in estuarine and intertidal habitats while red crabs are more common in the more saline waters of open shore and subtidal environments [2,150,160]. Bessa et al. [111] found that the green color morph comprised 93% of all crabs caught in an estuarine study site in Portugal. Furthermore, red females are more abundant than red males at estuarine sites [2,146,150,157]. These different distributions were not seen, however, in the nonindigenous population in southern Maine, where there was an even distribution of green phase crabs across sex in the intertidal and red phase crabs of both sexes were present in the intertidal throughout the year and at nearly the same abundances as green phase crabs during the summer months [125]. Adult C. maenas can tolerate short-term exposure to high salinities of up to 54‰ [1,3,8,96] but the preferred salinity range based on laboratory choice experiments performed with Isle of Man crabs is reported to be 27–40‰ [115,161], and Broekhuysen [3] indicated that crabs are rarely found in salinities > 31‰.
In contrast to the pattern for temperature, larvae are less tolerant of salinity fluctuations than juveniles but juveniles are still more tolerant than adults [28,149,150,158,159,162]. Anger et al. [163] found that C. maenas early larvae from the North Sea showed reduced respiration at low salinities, suggesting a very low or lacking osmoregulatory capacity, but salinity tolerance increases with successive larval stages to megalopae and euryhalinity is finally achieved in juveniles [152,164].
4.3. Oxygen
As long as they remain moist and at a moderate temperature, C. maenas adults are able to survive out of water for at least 10 days [2,165]. In response to lower oxygen tension they are able to breath air by reversing the direction of the scaphognathite beat to cause air to enter the branchial chamber via the normally exhalent openings and exit through the normally inhalant Milne-Edwards openings, a process referred to as “bubbling” [2,9,166,167,168]. Perhaps this ability allowed green crabs to be introduced from New England into California by transport in shipments of live bait worms and seafood packed in wet seaweed [20,27]. Red phase crabs leave the water and begin bubbling at much higher oxygen tension, are less able to compensate for hypoxia and die more rapidly in anoxic conditions than green phase crabs [167,169] and therefore avoid the intertidal zone. Adult C. maenas are less able to compensate for low levels of oxygen when their oxygen consumption rate is high, especially during ebb tide [169]. Under conditions of declining oxygen tension, the degree of respiratory independence shown by C. maenas is dependent on the animal’s level of activity and inactive animals can maintain respiratory independence down to a PO2 of 60–80 mmHg [170]. Both male and female adult crabs from Helgoland, Germany, increased their respiration with increases in temperature as expected for an ectothermic animal, but had relatively low Q10 values, suggesting that C. maenas has a capacity to stabilize its metabolic rate under physically diverse habitats [171]. Large numbers of crabs move into the intertidal to forage during high tide [2,118,167] and some remain in the intertidal during low tide [2] so can experience hypoxia. As is the case with other animals living in tidepool environments, C. maenas can continue to function even in extreme hypoxia by a variety of mechanisms (see McMahon [172]).
5. Reproduction and Development
5.1. Molting
During its life cycle, C. maenas molts about 18 times, four during the zoea and megelopa larval stages and 14 after the larval stages [173]. The molting cycle consists of an intermolt period (termed diecdysis if short, anecdysis if longer) during which C. maenas feeds actively, followed by a relatively shorter molting period (ecdysis) when the old exoskeleton is shed to increase the body size. The molting process itself takes approximately 2–3 h [3], followed by a hardening of the carapace that varies with temperature from 3–4 days at 16 °C to 16 days at 10–11 °C [3]. As stated previously, newly molted crabs are green in color but as an intermolt period continues the color transitions through yellow and orange to red. It is accepted that green phase crabs are devoting energy to growth (so molt more frequently) whereas red phase crabs are devoting energy to reproduction (so are molting and growing much less) [2,113,125,158,174,175,176,177]. Many red crabs are in a terminal anecdysis and will not molt again. Those that do molt usually become pale green or yellow in color but some females have been reported occasionally to remain red immediately after ecdysis [178,179]. Red morphotypes generally present a heavier and thicker carapace while green morphotypes are lighter and thinner [173]. It is likely that a crab’s energy allocation shifts once it molts but that has not been examined. It is believed that molting occurs throughout the year for C. maenas in all populations, although Queiroga [180] suggests that the actual molting period in the native European range is only between February and October, rather than all year. In any case molting is age and temperature dependent, and the process peaks at certain intervals [150]. Several studies describe peak molting times for populations in the Netherlands, South Wales and France as between May and June [3,5,7], but in Mondego estuary, Portugal, the peak is April to June for males, and July to October for females [150].
For the western Atlantic population molting in male C. maenas in Prince Edward Island, Canada, occurs in a synchronized molting window during July [181]. In New Hampshire and Massachusetts C. maenas populations, molting in males is most common during the summer whereas molting in females is most common during the fall months [113,182]. In British Columbia within the northeastern Pacific range mature and older males with new shells were found from January through November, being most common from April through July and again in September and least common in August; numbers of new shell mature and older female crabs peaked in June and July, decreased in August and peaked again in September [59]. A recent molt of a large male crab (74 mm CW) was collected in August 2016 in Roche Harbor, Wescott Bay, Washington [8].
In comparison, male C. maenas in the Southern Hemisphere (e.g., Argentina) molt in November (early summer) and females molt in January–March (summer and fall) [82]. It is assumed that green crabs molt significantly less in the coldest temperatures of the year (below about 10 °C) due to observed low activity [9].
Whatever the true peak time of molting is for each regional population, age is always inversely proportional with molting frequency [9]. Green crabs molt much more frequently when young, with short intermolt periods (diecdysis) and intermolt intervals become much longer (anecdysis) as crabs age, until the crab enters a terminal anecdysis. Limb autotomy is essential for survival in juvenile C. maenas but is a rare event in sexually mature crabs [183]. Regeneration of autotomized limbs in first year crabs may be completed in one molt [1] but takes at least two or three molts after the first year [183,184].
5.2. Mating
Mating in C. maenas can take place only after the female has molted, so a peak in mating behavior is synchronized with sexually mature female ecdysis [3]. Males can mate repeatedly over a long period of time [185] without molting, and therefore typically molt earlier in the year in comparison to females [2,186]. They wait in shallow water closer to the shore for females to arrive [22]. Males locate receptive females by sensing pheromones that females release just prior to molting [9]. A male green crab then embraces a pre-molt female before, during, and after copulation to both claim the female and ensure her safety after copulation. Large red phase males have a competitive advantage in acquiring and mating with females over smaller male rivals [187]. The seasonality of mating varies among populations. In the native European population, C. maenas mating behavior is reported to occur between June and October [188], with a peak in August [3]. Studies in both Prince Edward Island [147] and Maine [139] suggest that the northwestern Atlantic population shares July–October mating behavior seasonality with the native European population. No studies of mating period specifically in the northeastern Pacific population have been published, but Klassen & Locke [9] indicate that C. maenas in British Columbia, Canada are seen in mating pairs around July. In Argentina, female C. maenas mate beginning in January (mid-summer) prior to moving to lower littoral levels where they remain through the fall and winter [82].
5.3. Ovigerous Females and Egg Release
There is no predictable link between time of mating and time of egg bearing because female C. maenas have the capacity to store male spermatophores for 4 1/2 months or longer [3]. As noted previously by Baeta, et al. [150] seasonality of ovigerous females differs considerably with study and region due to latitude (temperature) and other factors (see Table 1).
Studies from the Netherlands and Wales independently observed the appearance of ovigerous females between March and April [3,5], and observations from the Netherlands found that there are two windows in which ovigerous females are prevalent—during November and December, and during the spring and early summer [3]. At a site on the southwest coast of Ireland, there were two reproductive events annually, a primary winter cycle in which the larger crabs reproduced and a secondary summer cycle when smaller crabs reproduced [200]. It is possible for the same female to be ovigerous in more than one season, both because one fertilization event can yield two clutches for a female C. maenas [3] and because females carry their eggs for several months, depending on the water temperature [8]. In Massachusetts, 79% of ovigerous females were caught between May and August, and all others were caught between November and January [113], similar to the seasonality of the native European population. In British Columbia, female C. maenas were reported to be ovigerous during April and May [9,147] but more recently ovigerous females have been found in every month sampled from January through October [59]. It is suggested that C. maenas may have one breeding season in adverse conditions but may have a continuous breeding season in favorable conditions such as in newly invaded areas [59]. Ovigerous females are most abundant from May to September (winter months) along the Argentina coast [82]. Using baited traps to determine peak ovigerous periods could be misleading because ovigerous females tend to burrow into sediment or other shelter and remain inactive to avoid predation so do not feed and will not be caught in traps [9,112,146,147,150,182]. Perhaps the use of Fyke nets or other passive collection methods in addition to traps might catch a broader range of C. maenas to mitigate the error inherent in using only baited traps [147].
Depending on the persistence of colder temperatures (<10°), ovigerous females produce one or two clutches per year in all populations [9,139]. The number of clutches and eggs per gravid female varies with the size of the female, resources availability, and temperature [106]. Larger females, in warmer water with high food availability, are the optimal variables for greatest number of eggs per clutch [147,201,202]. In the native European population, the earliest estimation of C. maenas clutch size is 200,000 eggs [203] and Broekhuysen [3] calculated the number of eggs on a C. maenas of moderate size (46 mm CW) to be 185,000. Estimates of clutch size in the northwestern Atlantic population vary considerably, from 4781 to 165,940 eggs, with a mean of 75,577 ± 37,808 eggs [204]. However, the clutch size when the female is first ovigerous is not the same as when the eggs are released because eggs are lost during the brooding period. Early and late developmental stages of C. maenas in Prince Edward Island had average estimated clutch sizes of 195,833 ± 83,673 and 140,374 ± 60,717 eggs, respectively [147]. Therefore, fecundity likely varies for the same ovigerous female during different stages of embryonic development.
Carcinus maenas employs diverse strategies for egg release [196], and the timing of egg extrusion can vary by region because females have the capacity to release eggs when conditions are optimal [3]. After reaching sexual maturity, C. maenas females migrate to downstream areas where eggs can be shed near the open ocean [150]. In the native European population, eggs are generally released between February and July [195]. In the Wadden Sea off the coast of the Netherlands, the megalopal stage of C. maenas settles in large numbers from the end of June or early July onwards [205], most likely from the clutch carried in the winter [3]. In the Canal de Mira, Portugal, eggs generally were released between February and June, usually soon after a nighttime high tide [195].
In the northwestern Atlantic, the larval release window in Newfoundland is from late May or early June until August, peaking in June and July [196]. In Prince Edward Island the release occurs between the middle of July and the end of September [147], and the peak is in September in Maine [139]. Within the northeastern Pacific population, larval release peaks in British Columbia earlier, in May [9,198]. Coastal water temperatures in the northeastern Pacific are comparable to those in the northwestern Atlantic in summer months, but the northeastern Pacific waters are warmer in the winter, which offers ovigerous females a longer spawning window [3,9,38]. Therefore, it is possible that longer periods of warmer waters could positivelty influence the success of invasions in the northeastern Pacific, especially during El Niño events [51]. However, Best et al. [196] suggests that temperature is not likely to be the sole cause of difference in larval release patterns, but rather a combination of temperature, salinity [152], photoperiod [195], and food abundance [1] may all influence the seasonality of various reproductive events in temperate latitudes [206].
5.4. Larval Development
The planktonic component of the green crab life cycle consists of four zoeal stages and one megalopal larval stage [86,151,207,208]. Survival of C. maenas larvae is highly dependent on water chemistry parameters and laboratory experiments have shown that they respond negatively to exposure to reduced salinities. Larvae can tolerate salinities between 20 and 40‰ but apparently cannot develop below 20‰ [163], although the lower limit is reported as 19‰ by Rasmussen [209], 17‰ by Behrens Yamada [8] and 10‰ by Broekhuysen [3]. Broekhuysen [3] found that larvae at 10 °C require salinity >26‰ whereas those at 16–17 °C can develop at salinity as low as 20‰. Nagaraj [152] found that larvae developed to megalopae in all tested combinations of temperature (10, 15, 20, 25 °C) and salinity (20, 25, 30, 35‰) but highest survival was in the combinations of lowest temperature (10 °C) and highest salinities (30 and 35‰). Ovigerous females tend to congregate near the mouth of an estuary [111], allowing the larvae to exit the estuarine habitat via tidal cycles and complete their development in high salinity coastal waters before returning to the estuary after four to six weeks as megalopae or juveniles [76]. It is not surprising, then, that zoeae migrate out of the estuary into higher salinity coastal waters to complete development during the developmental period. Early zoeal C. maenas vertically migrate nocturnally, utilizing the ebb tide [210] to migrate offshore where they develop [195,208]. Benthic recruitment of early life stages primarily occurs in the upstream area of estuaries so the megalopal stage reinvades the estuary on a spring flood tide to restart the cycle [195,211,212,213]. Megalopae choose complex substrates to settle in and habitat selection continues through early instars [1,102,107,214].
Interestingly, larvae exhibit higher physiological tolerance to temperature fluctuations than adults [85,148] but larvae are more susceptible to cold temperatures (below 12.5 °C), thereby limiting range expansion into higher latitudes [85]. Temperature is inversely proportional to the time needed to complete development of the five larval stages (Zoea 1 through Megalopa) of C. maenas [151]. In controlled conditions, all five stages can be completed in as little as 18 days at a constant temperature of 25 °C, or in as long as 66 days at a constant temperature of 12 °C [151]. In nature, this early development can last for upward of 50 days or up to 82 days in laboratory experiments [85,215]. This long larval development means that the planktonic larvae remain in the water column for a long time and may be carried by currents and tides considerable distances to replenish or extend the range of existing populations. At study sites in England it was determined that megalopae that settle in April reached sexual maturity in the same autumn whereas megalopae that settled in August reached maturity the following spring at a slightly smaller carapace width [7,183,216]. In contrast, it took crabs in western Sweden more than two years to reach sexual maturity [97].
Larval behavior has a significant influence on invasion success [13] but establishment of C. maenas populations outside of the native range is contingent on a number of factors in addition to a successful and timely larval development, including invasions environmental variables such as water temperature [151] and day length [208,212] and biological factors such as food availability [1], competition and predation [40].
5.5. Life Span
The life span of C. maenas in the native European and nonindigenous western Atlantic populations is 5–7 years [9,139,217] and of crabs from Oregon and Washington within the northeastern Pacific nonindigenous population, is reported to be around 4–6 years [8,218]. No data are available for life spans in the other nonindigenous populations.
6. Size
6.1. Adult Maximum Size
The maximum sizes (CW) reported for crabs from many European, northwestern Atlantic and northeastern Pacific sites are compiled in Table 2.
Within the native range in Europe, crabs generally do not exceed 86 mm but a single male with a carapace width of 100 mm was collected in western Sweden (P.-V. Mosnes, unpublished, cited in Behrens Yamada et al. [218]) and a 92 mm male crab from Denmark was reported by Rasmussen [209]. The largest reported female in the native range measured 75 mm from England [226]. The smallest maximum CW reported for males within the native European population is 62 mm from England [231] and more recently 63 mm from the Netherlands [232]; the smallest maximum female reported is a 56 mm crab from Portugal [173]. Based on an analysis of carapace width in 28 native European sites gleamed from a literature review, Kelly et al. [241] concluded that C. maenas appears to follow the temperature-size rule for ectotherms, whereby the largest individuals are found in the coldest parts of the range and a linear regression analyzing male maximum carapace width and the latitude of each sample site in the native range found that latitude explained 61% of the variation in CW [39]. A later study concluded that adult body size showed negative correlation with environmental temperature in both the native and invaded northeastern Pacific ranges [241]. However, several size data values for the native population included in Table S1 in [241] and attributed to various authors actually are not found in the papers therein cited (e.g., sites Ria de Aveiro and Mondego estuary, Portugal; Thurlestone, UK; Dutch Wadden Sea, Netherlands; and the 100 mm value for Gullmarsfjorden, Sweden). Furthermore, a large number of reported values were not included in that analysis, including reports of some of the largest (86 mm) crabs collected near the middle of the range for both latitude and temperature. Elimination of the unsubstantiated data and inclusion of all of the data in our Table 2 suggests that the temperature-size rule actually may not apply to the native population.
The largest size reported for a male crab in the northwestern Atlantic population is 91 mm from both the northernmost part of the range in Nova Scotia [235] and much farther south in New Hampshire [182]; the largest female crab measured 88 mm from New Hampshire [182]. The smallest maximum size for males (69 mm) is also found in Nova Scotia [235] and for females (70 mm) from Maine [139]. Unfortunately, no size data have been published for crabs between Massachusetts and Maryland so any examination of the temperature-size rule would be limited to a small portion of the northwestern Atlantic population at the current time.
In the northeastern Pacific population, crabs from near the northern range limit in British Columbia tend to be larger than those from near the southern limit in California [39,245,246]. In this population the temperature-size rule appears to apply, based on collections from ten sites [241]. The largest C. maenas on record is a 114 mm red male from Barkley Sound, British Columbia and a red female with CW 93 mm was found in the same location [241]; however, the second largest size reported for a female is 89 mm for crabs found in both Oregon and near the southern limit of the range in California [241].
Although the temperature-size relationship is unclear, it does appear that C. maenas in both the northwestern Atlantic and northeastern Pacific nonindigenous populations tend to be larger overall than those in the native European population [198,241,247]. Furthermore, with only rare exceptions (see Young et al. [113]), males are larger than females. Grosholz and Ruiz [31] suggest that crabs of size 110–115 mm CW may be attainable; to date the largest C. maenas on record is the 114 mm male from British Columbia mentioned above [241].
6.2. Size at Sexual Maturity
The minimum size of sexually mature C. maenas varies considerably with no specific trend evident. It also should be noted that physiological maturity precedes morphological maturity [200]. In the native range the greatest minimum size of sexually mature C. maenas males is given as 44 mm in Norway [189], South Wales [5], UK [192] and Belgium [188] whereas the smallest sexually mature males (27 mm) have been reported from Portugal, many kilometers to the south [193,195]. For females the data are more variable, ranging from 38.6 mm in Ireland [200] to 21.5 mm in Portugal [193]. In the northwestern Atlantic, there are only three reports for male minimum size at maturity, 34 mm in Maine [139], 32 mm in Newfoundland [196], and 21.3 mm in PEI [147]. For females the minimum size at sexual maturity ranges from 40 mm in Nova Scotia [235] to 22 mm in Massachusetts (AM Young, personal observation). For the northeastern Pacific population the only value reported is an estimated 34 mm for females in California [8]. In Argentina, Vinuesa [82] concluded that female sexual maturity seems to be attained at about 40 to 50 mm CW based on a small sample size of 25 females collected, where the smallest mature crab measured 45.8 mm. See Table 2 for additional data for the native European and nonindigenous northwestern Atlantic populations.
7. Population Characteristics
7.1. Population Density
There is a significant knowledge gap in green crab population dynamics concerning estimates of population densities. Estimates of numbers of individuals in populations are rare for both juveniles and adults, owing to several factors, not the least of which is variability in time and space. The number of crabs in a particular area varies considerably with environmental factors such as temperature and salinity changes due to water depth and seasonal, diel and tidal movements, as well as internal variations attributed to sex, spawning, molting and life stage (juveniles versus adults) and biotic interactions with other species [93]. There are a variety of possible methods to estimate numbers within a population, each of which presents difficulties. Four of the most commonly used methods are discussed here; a review of additional methods can be found in Pihl & Rosenberg [98]. Individuals can be counted directly along a transect or within quadrats (see for example [52,105,125,145,248]) but this method is practical only for the intertidal zone at low tide (where mostly juveniles will be found) or in very shallow water. Extremely high densities of small juvenile crabs ranging from 200–2000 m−2 have been estimated using this method [95,109,249,250]. Divers can count crabs in subtidal areas but the type of vegetated or rocky bottom that offers suitable refuge also makes crabs very difficult to see. The most common method of collecting C. maenas is through the use of baited traps of various designs. Surveys by trapping do not determine the abundance of crabs in a population but rather the number of crabs captured based on foraging activity [113]. With this method, only feeding crabs will be caught and crabs below a certain size can escape through the trap mesh. Additional issues that affect catch (and therefore an accurate estimate of population size) include trap design, suitable bait, time of day and tide (crabs tend to be more active at night and during high tide, presumably because they are less visible to predators) [2,113,124,143,251,252], crab state (newly molted and ovigerous females tend to burrow and do not feed) [2,112], and mobility (the population as a whole is very active, migrating many meters, even kilometers, in a matter of a few hours) [21,115]. Furthermore, there are concerns that the presence of large crabs or large numbers of crabs in a trap dissuades additional crabs from entering [251,253]. It is estimated that only 80% of a green crab population can be caught at any one time using traps [2]. Underwater video used to evaluate the effectiveness of Fukui traps to catch green crabs revealed only a 16% success rate out of 1226 entry attempts [254]. It has been shown recently that certain modifications to the Fukui traps can significantly increase productivity up to 59% [255]. Passive nets are sometimes used but this method trusts to the currents and wandering crabs to enter the net so may be equally unreliable as a population density estimate. Trawling may be used if the bottom conditions are suitable [256] but often they are not due to the crab’s preference for bivalve beds or other geometrically complex areas. Lastly, a common method for estimating population numbers of a mobile species is tag and recapture. This method requires that a tag not be lost during the study period so care must be taken to ensure that a tag or mark stays intact if a tagged crab molts. Although snipped spines will be replaced when a crab molts, this method of tagging was used successfully to estimate the density of a C. maenas population in areas of the Kattegat in Denmark [220]. There also must be a sufficient number of tagged crabs recaptured to make calculations meaningful. In an attempt to determine growth rate of green crabs in the Ria de Aveiro Lagoon in Portugal, Gomes [21] used a suture-tag method described by Edwards [257] whereby suture filaments with numbered plastic buttons were inserted into the suture line between the gills and the digestive tube, just over the last leg, where there is some flesh. The tags remained in place through a molt, but, despite offering a bounty to crab fisherman for tagged crabs, the recapture of tagged individuals was extremely small (490 out of 6800 or 0.7%), making any accurate determination of growth rate or population size impossible. Nearly 3000 crabs tagged with Floy tags inserted into the right gill chamber along the suture line yielded approximately a 15% recovery [258]. Crabs marked by branding a code on the dorsal carapace with a battery charged soldering bolt retained the marks after molting, although the marks changed color from red to white [187]. Due to the difficulty in obtaining accurate numbers, in most cases the number of green crabs in a population is merely described subjectively, based on various collections and observations, using such terms as “uncommon,” common,” “abundant,” “extremely abundant” or “very large concentration.” In well-established populations, green crabs are almost always abundant or extremely abundant except perhaps at the range limits where they might be uncommon.
7.2. Sex Ratios
Another aspect of C. maenas population dynamics is the male to female sex ratio determined by trapping or other survey methods. Sex ratios can vary depending on the sub-range characteristics (water depth, temperature, salinity) as well as when sampling is conducted because migration patterns and habitat preferences of male and female green crabs differ. Some migration behaviors can be attributed to sex-specific differences in physiological tolerances [113,150]. It is generally stated that female C. maenas are less tolerant of low salinity than males, which keeps the majority of females in the subtidal zone while males migrate in and out of the intertidal zone [143,144,146,176]. Multiple studies in Europe [3,145,177,259] and the western Atlantic [113,149,182,198,260] have shown that in waters with a higher salinity sex ratios favor females. However, a recent study in Maine, US, found that females were more tolerant of low salinity than males and there were many females in the intertidal zone [125]. Skewed sex ratios have also been attributed to migration behavior during the mating periods of mature female C. maenas because the females likely migrate to downstream areas where the salinity is higher when they are ready to mate, leaving a greater proportion of males in upstream areas. In addition, more males than females may be caught in traps because males are more likely to enter traps because they are more active than females [143,144,145,261] and ovigerous females tend to be inactive and burrow into soft sediment to avoid predation so they do not feed and also will not be caught [112,113,182,262].
Most of the sex ratios reported for the native European population show male bias (see Table 3) but in the Mondego estuary in Portugal (latitude 41° N) females outnumbered males caught (SR = 0.89) whereas the sex ratio of juvenile C. maenas in upstream areas favored males in the winter months [150], suggesting that females migrate to downstream areas when they are approaching maturity and will be ready to mate. In the northeastern Pacific nonindigenous population male biased ratios are more common in the higher latitude portion and female biased ratios are more common in the lower latitude portion of the range. At ten trapping sites between Barkley Sound, British Columbia, Canada and Elkhorn Slough, California, US, the sex ratios were biased toward males at the colder sites and became more female biased at the warmer sites [241] (See Table 3 for details).
In reality, there are just as many published studies that report female biased M:F sex ratios (<1) as there are that report male dominated populations (see Table 3). Male biased ratios appear to be more common in the higher latitude (colder) portion of a range whereas female biased ratios appear to be more common in the lower latitude (warmer) region. For example, within the nonindigenous northwestern Atlantic range, the M:F sex ratio is reported to be male-biased in the Prince Edward Island, Canada, region [147], but female-biased in the more southerly area of southern Maine [125], New Hampshire [182] and Massachusetts [113,260]. Although sex ratio appears to be correlated with latitude, it is quite possible that other factors may in fact influence the ratio, such as when (both seasonal and daily variations) and how (type of traps or seining) crabs were collected. It is entirely possible, even likely, that the actual ratio of males to females in the population is very close to 1 (the same number of males and females) and the bias comes from sex differences in activity as well as the survey process itself. In Plum Island Sound, Massachusetts, Ropes [112] found that the sex ratios of downstream and upstream sites were 0.34 and 0.90, respectively. A recent study assessing the distribution and relative abundance of C. maenas in Chignecto Bay, upper Bay of Fundy, Canada, found that the ratio of male to female crabs decreased as the season progressed (but no actual SR values were reported) indicating that season greatly influences sex ratios [126]. Similarly, in Placentia Bay, Newfoundland, the overall proportion of sexes was reported to be 60% male and 40% female (exact SR = 1.48) but the sex ratio varied from a high of 1.76 in the fall to a low of 1.29 in the summer [196].
8. Ecosystem Dynamics
In addition to physical parameter tolerances (e.g., temperature, salinity, oxygen), the other major determinant of whether or not a population can be successfully established and/or expanded involves ecosystem dynamics (e.g., diet, competitors, predators and parasitic infections).
8.1. Diet
Juvenile C. maenas feed primarily on detritus and interstitial meiofauna and then become more carnivorous as they age [97,99,267,268]. Green crab adults are opportunistic omnivores, whose diet varies with season, composition of the community, size of prey, availability of prey and energy expenditure versus energy gained [116,262,269]. Green crabs are known to feed on organisms from at least 158 genera in five plant and protist and fourteen animal phyla [9,28,36,270], with members of all populations showing a strong preference for molluscs, especially bivalves such as clams, mussels and cockles [9,30,44,66,99,136,228,249,271,272,273,274,275,276,277,278]. Additional preferred prey items include gastropod molluscs such as periwinkles [8,37,275,279], other crustaceans such as the common brown shrimp Crangon crangon [262] and recently settled small juvenile lobsters Homarus americanus [280] as well as members of their own species [101,105,106,205,249], polychaetes [281] and amphipods [30]. There appears to be no difference in foraging between males and females, suggesting that the two sexes are probably ecological equivalents [234]. In addition to being predators of a large variety of organisms, green crabs are also scavengers of any dead material along the shore [22,112].
In the native European population, there are regional differences in diet composition. In the temperate Mondego estuary in Portugal, C. crangon, polychaete worms and teleost fish were the most abundant stomach items by occurrence, numbers, and weight [262]. In the Menai Strait, North Wales and in the Rance Basin, France, green crabs were reported to eat mostly other crustaceans, polychaetes and algae [122,282]. Jensen & Jensen [273] found that younger C. maenas in the Danish Wadden Sea favor the common cockle, Cerastoderma edule, over other prey items. Scherer & Reise [249] found that older males continue to feed on molluscs but older females feed primarily on annelid worms, although Baeta et al. [262] found that different age classes and sexes of C. maenas showed no difference in diet when they occurred in the same area of an estuary in Portugal. More research should be done in the topic of differential feeding by different size classes, as it could be utilized for selective removal of certain size classes when trapping.
In the nonindigenous population in South Africa the most important prey items, based on gut contents, were gastropods, isopods and polychaetes (see Le Roux et al. [65] for a list of over 30 organisms known to be consumed by C. maenas), whereas in Australia, crabs preyed predominantly on bivalves such as cockles and mussels and native crab species [78,244,283]. The authors are not aware of any published information on food eaten by C. maenas in Argentina. For a more extensive list of prey organisms, see discussions in Crothers [2]; Le Roux et al. [65]; Behrens Yamada [8]; Klassen & Locke [9].
Herbivory in larger crabs may be by accidental ingestion [262], but there is evidence that some portunid crabs feed selectively on plant material or algae [65,112,122,282]. In the northwestern Atlantic population, C. maenas alters its diet from bivalves, its preferred prey, to consume more algae in the presence of a competitor species, the Asian shore crab Hemigrapsus sanguineus [284]. The wide variety of prey items that C. maenas consumes indicates that they are resilient, and can survive on whatever food is available [8]. Green crabs can even survive three months of starvation [285].
8.2. Competitors
Green crabs experience both intraspecific (within the species) and interspecific (between different species) competition within established populations. In South Wales, C. maenas is by far the most abundant of several crab species, suggesting that it wins most interspecific competition and therefore intraspecific competitions is probably much more important in determining distribution [2]. Intraspecific competition is not well documented nor its overall significance established, but it is assumed that as the density of a population increases, intraspecific competition for available resources also increases. Carcinus maenas is a very aggressive species so it is likely that there are many agonistic encounters with other members of the population. Observed higher rates of leg autotomy within higher density C. maenas populations in British Columbia, Canada, were attributed to intraspecific agonistic behaviors [198].
Interspecific competition is also difficult to document in the field so it generally is assumed that if two somewhat similar species are consuming the same foods and/or occupying similar habitat, they must be competing for those resources. Within the native European population in South Wales, there were 29 other crab species in localities where green crabs were found [286], many of which eat similar food, so C. maenas could be in competition with at least some of them [2], but they all have slightly different habitat preferences that serve to keep them apart (fragmentation of the niche) [287]. Crothers [2] argues that any competition among species would favor C. maenas, restricting the distribution of the other species. The European edible crab Cancer pagurus was able to consume larger bivalves than C. maenas because the species has stronger chelae [288]. The brush-clawed shore crab, Hemigrapsus takanoi, was unintentionally introduced to the Netherlands in the late 1990’s and is now abundant in the Eastern Scheldt, where adults compete with juvenile C. maenas [289]. A sibling species, the Asian shore crab, H. sanguineus, is also found in European waters, having arrived by 2003 in the Mediterranean Sea [290], around 2007 on the coast of France [291] and then Helgoland, Germany, where it is now the second most abundant intertidal crab in the North Sea after C. maenas [171]. It is predicted that the Asian shore crab will assert competitive dominance over the green crab in this region [171].
The native rock crab, Cancer irroratus, and Carcinus maenas coexist in southwestern Nova Scotia so they may compete for resources [122,292,293]. The rock crab is a larger species that is known to outcompete C. maenas in a lab setting [294], and apparently competitively excludes green crabs from their preferred habitat and forces them to occupy the upper intertidal in Chignecto Bay, Bay of Fundy, Canada, resulting in habitat segregation between the two species [136]. On the Isles of Shoals, New Hampshire, green crabs were abundant in the intertidal while rock and Jonah crabs (Cancer borealis) were both more abundant in subtidal waters [295]. In Prince Edward Island green crabs and rock crabs seemed to avoid each other on mussel clumps, suggesting that the two species could coexist [296]. In the Bras d’Or Lakes, Nova Scotia, rock crabs consumed large numbers of juvenile green crabs [297]. In contrast, another study determined that rock crabs were being displaced or preyed upon by C. maenas in Newfoundland [278]. Juvenile American lobster, H. americanus, and two species of crabs native to the northwestern Atlantic, the blue crab, Callinectes sapidus, and the lady crab, Ocellatus ovalipes, all consume similar food (bivalves, gastropods, polychaetes and crustaceans) so are assumed to be competitors with C. maenas [99,112,122,298]. In laboratory experiments adult green crabs were able to physically compete with and, in many cases, dominate larger, heavier lobsters [298]. In addition to consuming small juvenile lobsters green crabs curtail the overall foraging activity and shelter use by lobsters [265,299,300] and may be able to locate and consume a food source before the lobsters can get to it [238]. In Passamaquoddy Bay in the Bay of Fundy, St. George’s Bay in the Northumberland Straight and Chedabucto Bay on the Scotian Shelf the magnitude of the predatory effect of green crabs on lobsters varies and is correlated with the genetic origins of the invasive populations [301].
In the northeastern Pacific population few native species appear to compete with C. maenas for food. In laboratory experiments, juvenile C. maenas were able to out-compete equal-sized Metacarcinus (formerly Cancer) magister (Dungeness crab) whereas adult M. magister could out-compete adult green crabs, but they do not currently share the same habitat so are actually not in direct competition [302]. Limb autonomy was more common in adult C. maenas in areas of Bodega Bay, California that were inhabited by red rock crabs Cancer productus and brown rock crabs Cancer antennarius, likely due to predation by adult rock crabs [9,303]. At Pipestem Inlet, British Columbia, Gillespie et al. [198] noted mixed catches of C. maenas and the smaller graceful rock crab, Cancer gracilis. Both of these species are likely using this low salinity area as a refugium from the larger adult red and brown rock crabs so these two species may be in direct competition. However, green crabs were common and C. gracilis absent in traps set high in the intertidal but the reverse was true for traps set in the low intertidal and subtidal areas, suggesting that these two species are showing niche segregation to avoid interactions [304].
The Asian shore crab, H. sanguineus has recently invaded parts of the northwestern Atlantic where it competes with established C. maenas, and a native northeastern Pacific sibling species, H. oregonensis, that has several common names (yellow shore crab, hairy shore crab, mud-flat crab, Oregon shore crab), is now in direct competition with recently invasive C. maenas [114,276,304,305,306]. Under controlled experimentation H. sanguineus was overwhelmingly dominant over C. maenas whereas C. maenas dominated over H. oregonensis [114]. In the wild, C. maenas in the presence of H. sanguineus shifts its habitat use, abandoning otherwise preferred habitat under rocks in the cobble of intertidal areas [114]. Along the southern New Jersey, Long Island and southern New England coastlines, H. sanguineis has essentially eliminated juvenile C. maenas in the rocky intertidal zone where it formerly was extremely abundant [248,306,307,308,309,310,311,312] and apparently the same thing has happened along the Delaware coastline [313]. In southern New England juvenile C. maenas were the numerically dominant species in the rocky intertidal zone [314] but H. sanguineus is now the numerically dominant (90%) crab species [310,315] while C. maenas densities have declined 40–90% [306]. Hemigrapsus sanguineus can outcompete C. maenas for food and shelter resources, forcing C. maenas to not only occupy a less-desirable habitat but also alter its diet from bivalves, its preferred prey, to more algae [114,284]. In laboratory experiments green crabs were superior competitors to both blue crabs and Asian shore crabs [308], but that does not seem to be the case in the wild, where H. sanguineus adults either prey on or displace C. maenas recruits [306,316].
The authors are not aware of any published information on C. maenas competitors in the other nonindigenous populations in Australia, South Africa or Argentina.
8.3. Predators
The major predators of C. maenas include many species of birds, fish, and other crustaceans, such as the rock crabs and lobsters discussed above (for more in-depth discussion of various predators and lists of predators, see Crothers [2]; Cohen et al. [36]; Klassen & Locke [9]; Wong and Dowd [317]). Cannibalism among green crabs, especially juveniles, is common and often proportional to the population density, so C. maenas must be considered as a predator of itself [101,105,106,112,262,318]. Predators other than in those three groups include a few mammals such as mink, Mustela vison and otters, Lutra lutra [319,320] and harbor seals, Phoca vitulina [321]. The southern limit of the nonindigenous northwestern Atlantic population apparently is determined by the native predator Callinectes sapidus that provides biotic resistance to invasion and prevents the southward spread and establishment of Carcinus maenas within and south of Chesapeake Bay, despite the fact that the temperature regime in those areas falls within the acceptable temperature distribution seen in green crab populations elsewhere in the world [40]. In laboratory studies juvenile green crabs are superior competitors to juvenile blue crabs [304] but adult blue crabs are more than twice the size of green crabs and readily prey on them [40]. As C. maenas has expanded northward into Canadian waters, interactions with native lobsters (Homarus americanus) have increased. Usually adult green crabs retreat from encounters with the larger lobsters but some antagonistic responses result in the crabs being eaten [237,322].
The authors are not aware of any published information on C. maenas predators in the other nonindigenous populations in Australia, South Africa or Argentina, but one could assume that species of gulls and other shorebirds and fish and perhaps other crustacean species would be the major predators, as they are in the other populations.
Perhaps the most significant C. maenas predator is Homo sapiens (humans). In the native population, there are established fisheries for green crabs in several European countries, where they are caught to be fried as a soft-shell crab delicacy or as an ingredient for soups and fish pastes [22]. In Canada, a green crab fishery for food is being explored [323] but the difficulty of removing meat from the carapace by conventional methods and the relatively small size of C. maenas poses challenges for a traditional “hard-shell” fishery [324,325]. A pilot commercial fishery was attempted in 2014, but its long-term practicality was deemed questionable because of the lack of suitable markets for the harvested crabs [181]. There currently is considerable interest and progress in the Atlantic Canada and New England region for development of a variety of green crab based food products [323,326,327,328,329,330,331], with whole books dedicated to the cooking of green crabs [332]. The variable colors (green, yellow, orange, red) seen on the ventral surface of C. maenas are potential indicators for various physiological processes [1,113,139,174,333,334,335]. Color as an indicator of when a green crab is about to molt is of interest to any potential soft-shell crab food business modeled on the lucrative moleche (fried soft-shell crab) industry with Carcinus aestuarii in Venice, Italy. Male C. maenas exhibit three external characteristics that are indicative of an imminent molt: presence of a halo or greying circle on the episternites, darkening of the apex line, and looseness of the ecdysial line (where abdomen meets posterior carapace) [181]. Beyond predation for food consumption, humans also collect green crabs for other uses, such as fish, conch and lobster bait [22,336] and fertilizer [327], as well as utilizing chitin as an intermediate material in the production of self-healing car paints [337], cosmetics [338], antimicrobial textile treatments [339], bioadhesive delivery gel [340], and inexpensive antiviral drugs [341], an affinity ligand for protein purification [342], or just to reduce the density of invasive populations [9,278,343].
8.4. Epibionts
In the native European and nonindigenous northwestern Atlantic populations, C. maenas can become fouled with epibionts, primarily rock barnacles, slipper shells, serpulid tube worms, bryozoans, and occasionally tunicates. During an extended intermolt period, the number of epibionts increases as the crabs age without molting, so red crabs are the most heavily fouled [2,113,344]. In the native European population C. maenas infected with the rhizocephalan parasite Sacculina carcini are much more likely to be fouled than non-infected crabs [344,345]. The authors are not aware of any published information on C. maenas epibionts in any of the other nonindigenous populations.
8.5. Pathogens and Parasites
Green crabs are susceptible to a wide range of pathogens including bacteria, viruses, dinoflagellates, haplosporidians, helminths, nemerteans and trematodes (see review in Leignel et al. [28]). Green crabs have recently been identified as a carrier of novel pathogenic species, from viruses [346] to parasitic Metazoa [347]. While there are a number of parasites that infect green crabs (see lists in Crothers [2], Torchin et al. [348]; Behrens Yamada [8]; Klassen & Locke [9]), the most famous and well-studied is the rhizocephalan barnacle, Sacculina carcini. Rhizocephalans are morphologically modified crustacean parasites such that the main body of the parasite is a root-like system of branches that extends into most of the host’s tissue. Red phase C. maenas are externally parasitized by S. carcini to a higher degree than green phase crabs in the Mondega estuary in Portugal [349], and the Dutch Wadden Sea [350]. The youngest developmental stages of the barnacle predominantly occur when the crabs are green, the intermediate stages when they are orange, and the oldest when they are red [344], and it has been shown most recently that the success of male green crab S. carcini acquisition is susceptible to both seasonal and regional variation in the native range [351]. Infected crabs are castrated by the parasite and cease molting [219,345,352]. As is the case with many species, green crabs in non-native populations are substantially less infected by parasites than in the native population [348]. In Europe, approximately 16% of green crabs are infected with S. carcini but the parasite is not found in any of the nonindigenous populations. Although the global success of C. maenas is attributable to multiple factors, the escape from S. carcini has been hypothesized to be a major reason for the success of green crabs in nonindigenous ranges [348]. Parasites such as S. carcini suppress C. maenas populations in many areas of Europe, and the escape from these natural enemies increases the performance where green crabs have invaded [353]. Non-native crabs in some populations are infected with some of the other parasitic worms, including trematodes, nematodes, acanthocephalans and nemerteans [9,348], but the parasite load in introduced regions generally amounts to only a fraction of that in the native region [353]. Furthermore, the parasites that are most commonly acquired in introduced regions (such as the trematode, Microphallus similis) do not have the same lasting effects on physiology and behavior as S. carcini [354]. Several parasitic species, including S. carcini, have been proposed as potential biological control agents for invasive green crabs [8,28,355,356] but any intentional introduction of another species into an environment is risky and could result in the death of native crab species [357].
9. Ecological Impacts
The green crab is considered an “ecosystem engineer” because it has the ability to modify entire ecosystems [9,358,359], including such habitats as mudflats [252], saltmarshes [112,360], eelgrass beds [361,362,363] and rocky shores [364]. Both native and nonindigenous populations of C. maenas have had impacts, mostly negative, on ecosystem components. For examples, in the Menai Straits it has had a devastating effect on commercial mussel beds [365], and has reduced the density of cockles, Cerstoderma edule, in the Danish Wadden Sea [273] and C. edule and Macoma balthica in Scotland [366]. In the Dutch Wadden Sea, mass mortalities of common eider ducks, Somateria mollissima, were attributed in part to the transmission of the acanthocephalan parasite Polymorphus botulis for which the green crab is an intermediate host [367]. Negative impacts have been far greater in the invaded ranges, however. In the northwestern Atlantic, C. maenas has caused considerable damage to blue mussels [276], American oysters Crassostrea virginica [292], rock crabs Cancer irroratus and lobsters H. americanus [280] along the Atlantic Canada and New England coasts [278]. As of now, there has been no decline in C. irroratus as a result of the emergence of green crabs in the Upper Bay of Fundy, Canada [126], but the situation merits monitoring. In Atlantic Canada green crabs recently have expanded into areas populated by native mud crabs (Dyspanopeus sayi), and C. maenas may threaten the sustainability of this native species [106]. It has been labeled a “moderate to high risk” species, both environmentally and economically, for both coasts of Canada [215].
Green crabs have had a devastating effect on the economically important soft-shell clam Mya arenaria industry in New England [44,112,271,272,274,277,368,369,370]. Some of the poorest clam harvest years in Massachusetts and Maine have followed high population levels of C. maenas and, conversely, some of the biggest clam harvests have followed declines in the C. maenas population, which were probably due to environmental factors such as extremely cold winters [19,44,368]. The early part of this decade has seen a large increase in the C. maenas population in Maine, New Hampshire, and northeastern Massachusetts, followed by subsequent sharp declines in the commercial harvest of soft-shell clams [371] only do adult green crabs feed on juvenile and adult clams but postlarval and juvenile crabs prey heavily on newly settled clam larvae [369,372,373,374,375]. Green crabs also prey on juveniles as well as settling and newly settled postlarvae of another economically important species, American lobsters [280,298,299,300,376]. Green crabs generally do not feed on eelgrass Zostera marina but their foraging activity digging in the soft substrate dislodges the grasses and has contributed to considerable loss of eelgrass beds along the New England coast [359,361,362,363,377,378,379]. Similarly, it has been concluded that green crabs indirectly caused a collapse in a unique strain of giant Irish moss (Chondrus crispus) in Prince Edward Island waters by consuming small blue mussels (Mytilus edulis), thereby depleting the mussel clumps required as attachment surfaces by the Irish moss [236].
In the northeastern Pacific nonindigenous population C. maenas has been present for a much shorter period of time so its ecological effects are not yet apparent but recent models project a potential loss of 2.8–64% in commercial shellfish harvest in Puget Sound as green crab densities increase [61]. Green crabs feed on clams (Nutricula spp.) that are a major food source for wading shore birds so it is feared that if C. maenas becomes abundant, those wading birds could be affected adversely [8,252]. Field experiments have also shown that green crab predation has facilitated the invasion of non-native clams in Bodega Harbor [25]. In laboratory experiments to evaluate competition for food, C. maenas out-competed juvenile Dungeness crabs Metacarcinus magister so increased numbers of green crabs could adversely affect that commercially important species [60,302]. In 1993 C. maenas larvae settled in mesh bags where Manila clams (Venerupis philippinarum) were being aquacultured and the subsequent crabs preyed on a wide size range of clams, reducing the harvest considerably [8,380]. However, changes in outplant timing was found to mitigate these potential impacts [381]. In Tasmania, shellfish growers are not concerned that C. maenas will affect their economically important oyster (Crassostrea gigas) or mussel (Mytilus edulis) industries because the shellfish are gown in racks and on longlines off the bottom and out of reach of green crabs [8,75] but there is concern for the commercial cockle fishery in South Australia [244].
Another potential ecological impact is the transmission of parasites and other symbionts from C. maenas to commercially important native crustacean species. A recent histological survey for symbionts of C. maenas identified 19 separate symbiotic associations in crabs collected from 27 sites. These included a variety of metazoan parasites, microbial eukaryotes, bacteria, and viral pathogens, some of which may pose a risk of transmission to other decapods and native fauna via the use of green crabs as bait [382].
It is interesting to note that the morphological features of some species have been altered due to interactions with C. maenas. The blue mussel Mytilus edulis and the snails Littorina saxatalis, L. obtusata and Nucella lapillus all possess thicker shells in sites where C. maenas is abundant [8,34,383,384,385].
10. Conclusions
Due to its phenotypic plasticity, wide tolerances and omnivorous diet, C. maenas is a very successful invasive species, having established populations in all continents except Antarctica. It is an ecosystem engineer and can have many adverse effects on ecological components within its settled region. Established population ranges are primarily determined by temperature and are restricted to temperate coastlines, where water temperatures do not drop below −1 °C or exceed 22 °C for extended periods. Within this suitable temperature band around the world, suitable habitat is further restricted to wave-protected sheltered bays and estuaries in which salinity does not fall below 4‰ or exceed 31‰ for adults (20‰ for larvae) for extended periods. Geometrically complex habitats such as cobble or mussel beds that offer protection from predators are preferred by both juveniles and adults, but adults also thrive on soft muddy sand substrates where they are able to burrow for shelter.
Despite the long history of this species as the subject of hundreds of studies there still are aspects of the biology that are not known conclusively, at least partly due to the genetic variability in different populations, but also to non-standardized methods for collecting data. For example, there is considerable variability in sex ratios reported for different populations, depending upon geographic as well as within-site location, time of year, and other considerations. One variable that often is not considered is method of collection, whether various nets and traps yield different results in the same study site. Part of a recent study [113] compared several different style traps and baits for their ability to catch crabs but more extensive comparisons are needed to determine if trap or net type affects not just the overall numbers but the size, age and sex data of crabs caught. In some cases the lack of information is simply due to few studies being conducted within certain portions of a population’s range. For example, there are almost no data on crabs found south of Massachusetts in the northwestern Atlantic population. Knowledge of population density in an area is of considerable interest and importance for understanding population dynamics but turns out to be a very difficult value to measure, especially by the traditional tag & recapture method for mobile animals, due to the potential loss of tags during molting as well as the large numbers of crabs that travel extensively over wide areas in short amounts of time. There even is lack of agreement on whether crabs move offshore during the cold winter months or stay near shore, perhaps in tunnels or burrows made by other organisms or the crabs themselves.
The nonindigenous populations in the northeastern Pacific, South Africa and Argentina have been established much more recently than that in the northwestern Atlantic and have been the subject of fewer studies, as has the long-established populations in Australia. Little has been published concerning some important aspects of the biology of C. maenas in these populations (e.g., size at sexual maturity, reproductive periods, longevity) and especially ecosystem functions (e.g., diet, competition, predation, epibionts, parasitism). This information would be very helpful to evaluate the global population dynamics of green crabs and it is hoped that scientists in those areas will provide a better understanding of this important invasive species.
Author Contributions
Both authors contributed significantly to the research and writing of this review. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
We thank Salem State University Interlibrary Loan librarian Rebecca Le Mon for obtaining almost all of the numerous reference papers for us, and we thank Katie Plotner for many helpful edits throughout the manuscript. The clipart green crab illustrations in Figure 1 were drawn by Tanya Rogers. We are grateful for the helpful comments from Ted Grosholz on the sections dealing with the northeastern Pacific population and from Carolyn Tepolt on the population genetics section. Additionally, the valuable comments of anonymous reviewers greatly improved this paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Crothers, J.H. The biology of the shore crab Carcinus maenas (L.) 1. The background—Anatomy, growth and life history. Field Stud. 1967, 1, 407–434. [Google Scholar]
- Crothers, J.H. The biology of the shore crab Carcinus maenas (L.) 2. The life of the adult crab. Field Stud. 1968, 2, 579–614. [Google Scholar]
- Broekhuysen, G.J. On development, growth and distribution of Carcinides maenas (L.). Arch. Neerlandaises de Zoologie 1936, 2, 257–400. [Google Scholar] [CrossRef]
- Edwards, R.L. Movements of individual members in a population of the shore crab Carcinus maenas (L.) in the littoral zone. J. Anim. Ecol. 1958, 27, 37–45. [Google Scholar] [CrossRef]
- Naylor, E. Seasonal changes in a population of Carcinus maenas in the littoral zone. J. Anim. Ecol. 1962, 31, 601–609. [Google Scholar] [CrossRef]
- Bruce, J.R.; Colman, J.S.; Jones, N.S. (Eds.) Marine fauna of the Isle of Man, Edn.2. In Liverpool Marine Biology Committee Memoirs on Typical British Marine Plants and Animals, No. 36; Liverpool University Press: Liverpool, UK, 1963. [Google Scholar]
- Demeusy, N. Biologie-influence des facteurs saisonniers sur la realisation de la puberte au sein dune population de Carcinus maenas L. Des cotes de la manche. Comptes rendus hebdomadaires des seances de l academie des Sci. 1963, 256, 4762–4764. [Google Scholar]
- Behrens Yamada, S. Global Invader: The European Green Crab; Oregon State University: Corvallis, OR, USA, 2001; p. 123. ISBN 1-881826-24-4. [Google Scholar]
- Klassen, G.; Locke, A. A Biological Synopsis of the European Green Crab, Carcinus maenas; Fisheries and Oceans Canada: Moncton, NB, Canada, 2007; pp. 1–75. [Google Scholar]
- Darling, J.A.; Bagley, M.J.; Roman, J.; Tepolt, C.K.; Geller, J.B. Genetic patterns across multiple introductions of the globally invasive crab genus Carcinus. Mol. Ecol. 2008, 17, 4992–5007. [Google Scholar] [CrossRef] [PubMed]
- Roman, J.; Palumbi, S.R. A global invader at home: Population structure of the green crab, Carcinus maenas, in Europe. Mol. Ecol. 2004, 13, 2891–2898. [Google Scholar] [CrossRef]
- Tepolt, C.K.; Darling, J.A.; Bagley, M.J.; Geller, J.B.; Blum, M.J.; Grosholz, E.D. European green crabs (Carcinus maenas) in the northeastern Pacific: Genetic evidence for high population connectivity and current-mediated expansion from a single introduced source population. Divers. Distrib. 2009, 15, 997–1009. [Google Scholar] [CrossRef]
- Domingues, C.P.; Creer, S.; Taylor, M.I.; Queiroga, H.; Carvalho, G.R. Genetic structure of Carcinus maenas within its native range: Larval dispersal and oceanographic variability. Mar. Ecol. Prog. Ser. 2010, 410, 111–123. [Google Scholar] [CrossRef] [Green Version]
- Darling, J.A.; Tsai, Y.H.E.; Blakeslee, A.M.; Roman, J. Are genes faster than crabs? Mitochondrial introgression exceeds larval dispersal during population expansion of the invasive crab Carcinus maenas. R. Soc. Open Sci. 2014, 1, 140202. [Google Scholar] [PubMed] [Green Version]
- Tepolt, C.K.; Palumbi, S. Transcriptome sequencing reveals both neutral and adaptive genome dynamics in a marine invader. Mol. Ecol. 2015, 24, 4145–4158. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, N.W.; DiBacco, C.; Van Wyngaarden, M.; Hamilton, L.C.; Stanley, R.R.E.; Bernier, R.; FitzGerald, J.; Matheson, K.; McKenzie, C.H.; Ravindran, P.N.; et al. RAD sequencing reveals genomewide divergence between independent invasions of the European green crab (Carcinus maenas) in the Northwest Atlantic. Ecol. Evol. 2017, 7, 2513–2524. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, N.; DiBacco, C.; Wringe, B.F.; Stanley, R.R.E.; Hamilton, L.C.; Ravindran, P.N.; Bradbury, I.R. Genomic evidence of hybridization between two independent invasions of European green crab (Carcinus maenas) in the Northwest Atlantic. Heredity 2017, 119, 154. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, N.W.; Bradbury, I.R.; Stanley, R.R.E.; Wringe, B.F.; Van Wyngaarden, M.; Lowen, J.B.; McKenzie, C.H.; Matheson, K.; Sargent, P.S.; DiBacco, C. Genomewide evidence of environmentally mediated secondary contact of European green crab (Carcinus maenas) lineages in eastern North America. Evol. Appl. 2018, 11, 869–882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elner, R.W. The influence of temperature, sex, and chela size in the foraging strategy of the shore crab, Carcinus maenas. Mar. Behav. Physiol. 1980, 7, 15–24. [Google Scholar] [CrossRef]
- Carlton, J.T.; Cohen, A.N. Episodic global dispersal in shallow water marine organisms: The case history of the European shore crabs Carcinus maenas and C. aestuarii. J. Biogeogr. 2003, 30, 1809–1820. [Google Scholar] [CrossRef]
- Gomes, V. First results of tagging experiments on Carcinus maenas (L.) in the Ria de Aveiro Lagoon, Portugal. Cienc. Biol. Ecol. Syst. 1991, 11, 21–30. [Google Scholar]
- Morris-Webb, L.; Goudge, H.; Duce, C. An Introductory Review of the Biology and Population Dynamics of the Green Shore Crab, Carcinus maenas (L.), in the UK, with Specific Reference to Menai Strait; Policy Research Report No. 07/18; Countryside Council for Wales (now Natural Resources Wales): Newtown, UK, 2007. [Google Scholar]
- Lafferty, K.D.; Kuris, A.M. Biological control of marine pests. Ecology 1996, 77, 1989–2000. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; The World Conservation Union (IUCN): Aukland, New Zealand, 2000. [Google Scholar]
- Grosholz, E.D. Recent biological invasions may hasten invasional meltdowns by accelerating historical introductions. Proc. Natl. Acad. Sci. USA 2005, 102, 1088–1091. [Google Scholar] [CrossRef] [Green Version]
- Towle, D.W.; Smith, C.M. Gene discovery in Carcinus maenas and Homarus americanus via expressed sequence tags. Int. Comp. Biol. 2006, 46, 912–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darling, J.A. More than one way to invade: Lessons from genetic studies of Carcinus shore crabs. In In the Wrong Place—Alien Marine Crustaceans: Distribution, Biology and Impacts. Invading Nature—Springer Series in Invasion Ecology, Vol. 6; Galil, B.S., Clark, P.F., Carlton, J.T., Eds.; Springer: New York, NY, USA, 2011; pp. 661–685. [Google Scholar]
- Leignel, V.; Stillman, J.H.; Baringou, S.; Thabet, R.; Metais, I. Overview on the European green crab Carcinus spp. (Portunidae, Decapoda), one of the most famous marine invaders and ecotoxicological models. Environ. Sci. Pollut. Res. 2014, 21, 9129–9144. [Google Scholar] [CrossRef] [PubMed]
- Grosholz, E.D.; Ruiz, G.M. (Eds.) Management Plan for the European Green Crab Submitted to the Aquatic Nuisance Species Task Force; 2002; p. 55. [Google Scholar]
- Grosholz, E.D.; Ruiz, G.M. Spread and potential impact of the recently introduced European green crab, Carcinus maenas, in central California. Mar. Biol. 1995, 122, 239–247. [Google Scholar]
- Grosholz, E.D.; Ruiz, G.M. Predicting the impact of introduced marine species: Lessons from the multiple invasions of the European green crab Carcinus maenas. Biol. Conserv. 1996, 78, 59–66. [Google Scholar] [CrossRef]
- Audet, D.; Dais, D.S.; Miron, G.; Moriyasu, M.; Benhalima, K.; Campbell, R. Geographic expansion of a nonindigenous crab, Carcinus maenas (L.), along the Nova Scotia shore into the southeastern Gulf of St. Lawrence, Canada. J. Shellfish Res. 2003, 22, 255–262. [Google Scholar]
- Almaça, C. Sur le problème de l’origine de Carcinus maenas (L.) du littoral américain. Rev. Fac. Ciènc. Univ. Lisboa. 1963, 11, 121–136. [Google Scholar]
- Vermeij, G.J. Phenotypic evolution in a poorly dispersing snail after arrival of a predator. Nature 1982, 299, 349–350. [Google Scholar] [CrossRef]
- Chilton, C. Note on the dispersal of marine Crustacea by means of ships. Trans. Proc. R. Soc. N. Z. 1910, 43, 131–133. [Google Scholar]
- Cohen, A.N.; Carlton, J.T.; Fountain, M.C. Introduction, dispersal and potential impacts of the green crab Carcinus maenas in San Francisco Bay, California. Mar. Biol. 1995, 122, 225–237. [Google Scholar]
- Eastwood, M.M.; Donahue, M.J.; Fowler, A.E. Reconstructing past biological invasions: Niche shifts in response to invasive predators and competitors. Biol. Invasions 2007, 9, 397–407. [Google Scholar] [CrossRef]
- Jamieson, G.S. European green crab, Carcinus maenas, introductions in North America: Differences between the Atlantic and Pacific experiences. In Proceedings of the 10th International Aquatic Nuisance Species and Zebra Mussel Conference, Toronto, ON, Canada, 13–17 February 2000; pp. 307–312. [Google Scholar]
- Kelley, A.; De Rivera, C.E.; Buckley, B.A. Intraspecific variation in thermotolerance and morphology of the invasive European green crab, Carcinus maenas, on the west coast of North America. J. Exp. Mar. Biol. Ecol. 2011, 409, 70–78. [Google Scholar] [CrossRef]
- deRivera, C.E.; Ruiz, G.M.; Hines, A.H.; Jivoff, P. Biotic resistance to invasion: Native predator limits abundance and distribution of an introduced crab. Ecology 2005, 86, 3364–3376. [Google Scholar] [CrossRef]
- Smith, S.I. The stalk-eyed crustaceans of the Atlantic coast of North America north of Cape Cod. Trans. Conn. Acad. Sci. 1879, 5, 27–138. [Google Scholar]
- Rathbun, M.J. Fauna of New England. Occas. Pap. Boston Soc. Nat. Hist. 1905, 7, 1–117. [Google Scholar]
- Rathbun, M.J. The cancroid crabs of America of the families Euryalidae, Portunidae, Atelecyclidae, Cancridae and Xanthidae. Bull. USA Natl. Mus. 1930, 152, 1–609. [Google Scholar] [CrossRef]
- Glude, J.B. The effects of temperature and predators on the abundance of the soft-shell clam, Mya arenaria, in New England. Trans. Am. Fish. Soc. 1955, 84, 13–26. [Google Scholar] [CrossRef]
- Roman, J. Diluting the founder effect: Cryptic invasions expand a marine invader’s range. Proc. R. Soc. B Biol. Sci. 2006, 273, 2453–2459. [Google Scholar] [CrossRef] [Green Version]
- Blakeslee, A.M.H.; McKenzie, C.H.; Darling, J.A.; Byers, J.E.; Pringle, J.M.; Roman, J. A hitchhiker’s guide to the Maritimes: Anthropogenic transport facilitates long-distance dispersal of a marine crab to Newfoundland. Divers. Distrb. 2010, 16, 879–891. [Google Scholar] [CrossRef]
- Blakeslee, A.M.H. Northwest Atlantic population structure and gene flow in the green crab: Current understanding of a dynamic invasion front, population admixture, continued anthropogenic expansion. In Green Crab Summit Presentation; University of Maine: Orono, ME, USA, 2013; Available online: http://www.seagrant.umaine.edu/green-crab-summit (accessed on 18 July 2018).
- Kanary, L.; Musgrave, J.; Tyson, R.C.; Locke, A.; Lutscher, F. Modelling the dynamics of invasion and control of competing green crab genotypes. Theor. Ecol. 2014, 7, 391–406. [Google Scholar] [CrossRef]
- Lehnert, S.J.; Lehnert, S.J.; DiBacco, C.; Jeffery, N.W.; Blakeslee, A.M.; Isaksson, J.; Roman, J.; Wringe, B.F.; Stanley, R.R.; Matheson, K.; et al. Temporal dynamics of genetic clines of invasive European green crab (Carcinus maenas) in eastern North America. Evol. Appl. 2018, 11, 1656–1670. [Google Scholar] [CrossRef]
- Behrens Yamada, S.; Hunt, C.; Richmond, N. The Arrival of the European Green Crab, Carcinus maenas, in Oregon Estuaries. In Proceedings of the First National Conference on Marine Bioinvasions, Cambridge, MA, USA, 24–27 January 1999; Pederson, J., Ed.; MIT Press: Boston, MA, USA, 2000; pp. 24–34. [Google Scholar]
- Behrens Yamada, S.; Kosro, P.M. Linking ocean conditions to year class strength of the invasive European green crab, Carcinus maenas. Biol. Invasions 2010, 12, 1791–1804. [Google Scholar] [CrossRef]
- McGaw, I.J.; Edgell, T.C.; Kaiser, M.J. Population demographics of native and newly invasive populations of the green crab Carcinus maenas. Mar. Ecol. Prog. Ser. 2011, 430, 235–240. [Google Scholar] [CrossRef]
- Behrens Yamada, S.; Thomson, R.E.; Gillespie, G.E.; Norgard, T.C. Lifting barriers to range expansion: The European green crab Carcinus maenas (Linnaeus, 1758) enters the Salish Sea. J. Shellfish Res. 2017, 36, 201–208. [Google Scholar] [CrossRef]
- Grason, E.; Symer, K.; Martin, K. European Green Crab in Puget Sound. University of Washington Puget Sound Institute and Washington Sea Grant Crab Team. 2018. Available online: https://uw.maps.arcgis.com/apps/Cascade/index.html?appid=807fd68b00ab4f99be1cfc4b87bd5a4f (accessed on 22 July 2019).
- Bagley, M.J. Microsatellite DNA analysis of native and invading populations of European green crabs. In Proceedings of the First National Conference on Marine Bioinvasions, Cambridge, MA, USA, 24–27 January 1999; Pederson, J., Ed.; MIT Press: Boston, MA, USA, 2000; pp. 241–243. [Google Scholar]
- Carlton, J.T. Man’s role in changing the face of the ocean: Biological invasions and implications for conservation of near-shore environments. Conserv. Biol. 1989, 3, 265–273. [Google Scholar] [CrossRef]
- Carlton, J.T. Introduced marine and estuarine mollusks of North America: An end-of-the-20th-century perspective. J. Shellfish Res. 1992, 11, 489–505. [Google Scholar]
- Yamada, S.B.; Peterson, W.T.; Kosro, P.M. Biological and physical ocean indicators predict the success of an invasive crab, Carcinus maenas, in the northern California Current. Mar. Ecol. Prog. Ser. 2015, 537, 175–189. [Google Scholar] [CrossRef]
- Gillespie, G.E.; Norgard, T.C.; Anderson, E.D.; Haggarty, D.R.; Phillips, A.C. Distribution and biological characteristics of European green crab, Carcinus maenas, in British Columbia. 2006–2013. In Canadian Technical Report of Fisheries and Aquatic Science 3120; Fisheries and Oceans Canada, Science Branch, Pacific Region, Pacific Biological Station: Nanaimo, BC, Canada, 2015. [Google Scholar]
- Jamieson, G.S.; Grosholz, E.D.; Armstrong, D.A.; Elner, R.W. Potential ecological implication for the introduction of the European green crab, Carcinus maenas (Linnaeus), to British Columbia, Canada, and Washington, USA. J. Nat. Hist. 1998, 32, 1587–1598. [Google Scholar] [CrossRef]
- Mach, M.E.; Chan, K.M. Trading Green Backs for Green Crabs: Evaluating the Commercial Shellfish Harvest at Risk to European Green Crab Invasion. F1000Res 2013, 2. [Google Scholar] [CrossRef]
- Colnar, A.M.; Landis, W.G. Conceptual model development for invasive species and a regional risk assessment case study: The European green crab, Carcinus maenas, at Cherry Point, Washington, USA. Hum. Ecol. Risk Assess. 2007, 13, 120–155. [Google Scholar] [CrossRef]
- Darling, J.A. Interspecific hybridization and mitochondrial introgression in invasive Carcinus shore crabs. PLoS ONE 2011, 6, e17828. [Google Scholar] [CrossRef] [Green Version]
- Joska, M.A.P.; Branch, G.M. The European shore-crab—Another alien invader? Afr. Wildl. 1986, 40, 63–65. [Google Scholar]
- Le Roux, P.J.; Branch, G.M.; Joska, A.P. On the distribution, diet and possible impact of the invasive European shore crab Carcinus maenas (L.) along the South African coast. S. Afr. J. Mar. Sci. 1990, 9, 85–93. [Google Scholar] [CrossRef]
- Griffiths, C.L.; Hockey, P.A.R.; van Erkom Schuink, C.; Le Roux, P.J. Marine invasive aliens on South African shores: Implications for community structure and trophic functioning. S. Afr. J. Mar. Sci. 1992, 12, 713–722. [Google Scholar] [CrossRef]
- Robinson, T.B.; Griffiths, C.L.; McQuaid, C.D.; Rius, M. Marine alien species of South Africa—Status and impacts. Afr. J. Mar. Sci. 2005, 27, 297–306. [Google Scholar] [CrossRef]
- Griffiths, C.; Robinson, T.; Mead, A. The alien and cryptogenic marine crustaceans of South Africa. In In the Wrong Place—Alien Marine Crustaceans: Distribution, Biology and Impacts. Invading Nature—Springer Series in Invasion Ecology Vol. 6; Galil, B.S., Clark, P.F., Carlton, J.T., Eds.; Springer: New York, NY, USA, 2011; pp. 269–282. [Google Scholar]
- Fulton, S.W.; Grant, F.E. Note on the occurrence of the European crab, Carcinus maenas, Leach, in Port Phillip. Vic. Nat. 1900, 17, 147–148. [Google Scholar]
- Fulton, S.W.; Grant, F.E. Some little known decapod crustacean with a description of a new species. Proc. R. Soc. Vic. 1902, 14, 55–64. [Google Scholar]
- Zeidler, W. Note on the occurrence of the European shore crab Carcinus maenas (Linn., 1758) in Australia. S. Aust. Nat. 1978, 53, 11–12. [Google Scholar]
- Rosenzweig, P.A. A range extension for the European shore crab Carcinus maenas (Linn., 1758) in South Australia. S. Aust. Nat. 1984, 59, 18–19. [Google Scholar]
- Gardner, N.C.; Kwa; Paturusi, A. First recording of the European shore crab Carcinus maenas in Tasmania. Tasman. Nat. J. 1994, 116, 26–28. [Google Scholar]
- Furlani, D.M. A Guide to the Introduced Marine Species in Australian Waters. In Centre for Research on Introduced Marine Pests; Report No. 5.; CSIRO Division of Fisheries; Centre for Research on Introduced Marine Pests, CSIRO Publishing: Victoria, Australia, 1996. [Google Scholar]
- Proctor, C.; Thresher, R. The invasive history and abundance of C. maenas in Australia. In Proceedings of the First International Workshop on the Demography, Impacts and Management of Introduced Populations of the European crab, Carcinus maenas, Hobart, Australia, 20–21 March 1997; CRIMP Technical Report 11. Thresher, R.E., Ed.; CSIRO: Hobart, TAS, Australia, 1997; pp. 31–33. [Google Scholar]
- Thresher, R.; Proctor, C.; Ruiz, G.; Gurney, R.; MacKinnon, C. Invasion dynamics of the European shore crab, Carcinus maenas, in Australia. Mar. Biol. 2003, 142, 867–876. [Google Scholar] [CrossRef]
- Sinclair, M.A. Interactions between native grapsid crabs and Carcinus maenas in Victoria. In Proceedings of the First International Workshop on the Demography, Impacts and Management of Introduced Populations of the European Crab, Carcinus maenas, Hobart, Australia, 20–21 March 1997; Thresher, R.E., Ed.; Technical Report 11. Centre for Research on Introduced Marine Pests: Hobart, TAS, Australia, 1997; pp. 35–41. [Google Scholar]
- Ahyong, S.T.; Wilkens, S.L. Aliens in the Antipodes: Non-indigenous marine crustaceans of New Zealand and Australia. In In the Wrong Place-Alien Marine Crustaceans: Distribution, Biology and Impacts; Springer: Berlin/Heidelberg, Germany, 2011; pp. 451–485. [Google Scholar]
- Ross, D.; Johnson, C.; Hewitt, C.; Ruiz, G.M. Interaction and impacts of two introduced species on a soft-sediment marine assemblage in SE Tasmania. Mar. Biol. 2004, 144, 747–756. [Google Scholar] [CrossRef]
- Ahyong, S.T. Range extension of two invasive crab species in eastern Australia: Carcinus maenas (Linnaeus) and Pyromaia tuberculata (Lockington). Mar. Pollut. Bull. 2005, 50, 460–462. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, P.; Vander-Velde, J.; Keable, S. Baseline survey of the benthic macrofauna of Twofold Bay, N.S.W., with a discussion of the marine species introduced to the bay. Proc. Linn. Soc. NSW 1989, 110, 339–367. [Google Scholar]
- Vinuesa, J.H. Molt and reproduction of the European green crab Carcinus maenas (Decapoda: Portunidae) in Patagonia, Argentina. Revista de Biología Trop. 2007, 55 (Suppl. 1), 49–54. [Google Scholar] [CrossRef]
- Vinuesa, J.H. Distribucion de crustaceos decapodos y estomatopodos del golfo San Jorge, Argentina. Revista de biología marina y oceanografía 2005, 40, 7–21. [Google Scholar] [CrossRef]
- Furota, T.S.; Watanabe, T.; Watanabe, S.; Akiyama, S.; Kinasita, K. Life history of the Mediterranean green crab, Carcinus aestuarii Nardo, in Tokyo Bay, Japan. Crustac. Res. 1999, 28, 5–15. [Google Scholar] [CrossRef] [Green Version]
- deRivera, C.E.; Gray Hitchcock, N.; Teck, S.J.; Steves, B.P.; Hines, A.H.; Ruiz, G.M. Larval development rate predicts range expansion of an introduced crab. Mar. Biol. 2006, 10, 1275–1288. [Google Scholar] [CrossRef]
- Compton, T.J.; Leathwick, J.R.; Inglis, G.J. Thermogeography predicts the potential global range of the invasive European green crab (Carcinus maenas). Divers. Distrib. 2010, 16, 243–255. [Google Scholar] [CrossRef]
- Carlton, J.T. The global dispersal of marine and estuarine crustaceans. In In the Wrong Place—Alien Marine Crustaceans: Distribution, Biology and Impacts; Springer: Berlin/Heidelberg, Germany, 2011; pp. 3–23. [Google Scholar]
- Bulnheim, H.P.; Bahns, S. Genetic variation and divergence in the genus Carcinus (Crustacea, Decapoda). Internationale Revue der gesamten Hydrobiologie und Hydrographie 1996, 81, 611–619. [Google Scholar] [CrossRef]
- Geller, J.; Walton, E.D.; Grosholz, E.D.; Ruiz, G.M. Cryptic invasions of the crab Carcinus detected by molecular phylogeography. Mol. Ecol. 1997, 6, 901–906. [Google Scholar] [CrossRef]
- Pringle, J.M.; Blakeslee, A.M.; Byers, J.E.; Roman, J. Asymmetric dispersal allows an upstream region to control population structure throughout a species’ range. Proc. Natl. Acad. Sci. USA 2011, 108, 15288–15293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascoal, S.; Creer, S.; Taylor, M.I.; Queiroga, H.; Carvalho, G.; Mendo, S. Development and application of microsatellites in Carcinus maenas: Genetic differentiation between northern and central Portuguese populations. PLoS ONE 2009, 4, e7268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tepolt, C.K.; Bagley, M.J.; Geller, J.B.; Blum, M.J. Characterization of microsatellite loci in the European green crab (Carcinus maenas). Mol. Ecol. Notes 2006, 6, 343–345. [Google Scholar] [CrossRef]
- Cosham, J.; Beazley, K.F.; McCarthy, C. Environmental factors influencing local distributions of European green crab (Carcinus maenas) for modeling and management applications. Environ. Rev. 2016, 24, 244–252. [Google Scholar] [CrossRef] [Green Version]
- Hampton, S.L.; Griffiths, C.L. Why Carcinus maenas cannot get a grip on South Africa’s wave-exposed coastline. Afr. J. Mar. Sci. 2007, 29, 123–126. [Google Scholar] [CrossRef]
- Breteler, W.C.M. Settlement, growth and production of the shore crab, Carcinus maenas, on tidal flats in the Dutch Wadden Sea. Neth. J. Sea Res. 1976, 10, 354–376. [Google Scholar] [CrossRef]
- Eriksson, S.; Evans, S.; Tallmark, B. On the coexistence of 0-group Carcinus maenas (L.) from a shallow sandy bottom in Gullmar Fjord, Sweden. Zoon 1975, 3, 65–70. [Google Scholar]
- Eriksson, S.; Edlund, A.-M. On the ecological energetics of 0-group Carcinus maenas (L.) from a shallow sandy bottom in Gullmar Fjord, Sweden. J. Exp. Mar. Biol. Ecol. 1977, 30, 233–248. [Google Scholar] [CrossRef]
- Pihl, L.; Rosenberg, R. Production, abundance, and biomass of mobile epibenthic marine fauna in shallow waters, western Sweden. J. Exp. Mar. Biol. Ecol. 1982, 57, 273–301. [Google Scholar] [CrossRef]
- Ropes, J.W. The food habits of five crab species at Pettaquamscutt River, Rhode Island. Fish. Bull. 1989, 87, 197–204. [Google Scholar]
- Zeng, C.; Naylor, E.; Abelló, P. Endogenous control of timing of metamorphosis in megalopae of the shore crab Carcinus maenas. Mar. Biol. 1997, 128, 299–305. [Google Scholar] [CrossRef]
- Moksnes, P.-O.; Pihl, L.; van Montirans, J. Predation on postlarvae and juveniles of the shore crab Carcinus maenas: Importance of shelter, size and cannibalism. Mar. Ecol. Prog. Ser. 1998, 166, 211–225. [Google Scholar] [CrossRef] [Green Version]
- Hedvall, O.; Moksnes, P.-O.; Pihl, L. Active habitat selection by megalopae and juvenile shore crabs Carcinus maenas: A laboratory study in an annular flume. Hydrobiologia 1998, 375, 89–100. [Google Scholar] [CrossRef]
- Amaral, V.; Paula, J. Carcinus maenas (Crustacea: Brachyura): Influence of artificial substrate type and patchiness on estimation of megalopae settlement. J. Exp. Mar. Biol. Ecol. 2007, 346, 21–27. [Google Scholar] [CrossRef]
- Almeida, M.J.; Flores, A.A.V.; Queiroga, H. Effect of crab size and habitat type on the locomotory activity of juvenile shore crabs, Carcinus maenas. Estuar. Coast. Shelf Sci. 2008, 80, 509–516. [Google Scholar] [CrossRef]
- Almeida, M.J.; Gonzalez-Gordillo, J.I.; Flores, A.A.V.; Queiroga, H. Cannibalism, post-settlement growth rate and size refuge in a recruitment-limited population of the shore crab Carcinus maenas. J. Exp. Mar. Biol. Ecol. 2011, 410, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Gehrels, H.; Knysh, K.M.; Boudreau, M.; Thériault, M.H.; Courtenay, S.C.; Cox, R.; Quijón, P.A. Hide and seek: Habitat-mediated interactions between European green crabs and native mud crabs in Atlantic Canada. Mar. Biol. 2016, 163, 152. [Google Scholar] [CrossRef]
- Todd, P.A.; Briers, R.A.; Ladle, R.J.; Middleton, F. Phenotype-environment matching in the shore crab (Carcinus maenas). Mar. Biol. 2006, 148, 1357–1367. [Google Scholar] [CrossRef]
- Price, N.; Green, S.; Troscianko, J.; Tregenza, T.; Stevens, M. Background matching and disruptive coloration as habitat-specific strategies for camouflage. Sci. Rep. 2019, 9, 7840. [Google Scholar] [CrossRef]
- Thiel, M.; Dernedde, T. Recruitment of shore crabs Carcinus maenas on tidal flats: Mussel clumps as an important refuge for juveniles. Helgoländer Meeresunters. 1994, 48, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Amaral, V.; Cabral, H.N.; Jenkins, S.; Hawkins, S.; Paula, J. Comparing quality of estuarine and nearshore intertidal habitats for Carcinus maenas. Estuar. Coast. Shelf Sci. 2009, 83, 219–226. [Google Scholar] [CrossRef]
- Bessa, F.; Baeta, A.; Martinho, F.; Marques, S.; Pardal, M.A. Seasonal and temporal variations in population dynamics of the Carcinus maenas (L.): The effect of an extreme drought event in a southern European estuary. J. Mar. Biol. Assoc. UK 2010, 90, 867–876. [Google Scholar] [CrossRef]
- Ropes, J.W. The feeding habits of the green crab, Carcinus maenas (L.). Fish. Bull. 1968, 67, 183–203. [Google Scholar]
- Young, A.M.; Elliott, J.A.; Incatasciato, J.M.; Taylor, M.L. Seasonal catch, size, color, and assessment of trapping variables for the European green crab Carcinus maenas (Linnaeus, 1758) (Brachyura: Portunoidea: Carcinidae), a nonindigenous species in Massachusetts, USA. J. Crustac. Biol. 2017, 37, 556–570. [Google Scholar] [CrossRef]
- Jensen, G.C.; McDonald, P.S.; Armstrong, D.A. East meets west: Competitive interactions between green crab Carcinus maenas, and native and introduced shore crab Hemigrapsus spp. Mar. Ecol. Prog. Ser. 2002, 225, 251–262. [Google Scholar] [CrossRef]
- Ameyaw-Akumfi, C.; Naylor, E. Spontaneous and induced components of salinity preference behavior in Carcinus maenas. Mar. Ecol. Prog. Ser. 1987, 37, 153–158. [Google Scholar] [CrossRef]
- Singh, R. Natural Diet of, and Shelter-Use by, the Green Crab, Carcinus maenas (L.) in Nova Scotia. Master’s Thesis, University of New Brunswick, Fredericton, NB, Canada, 1991. [Google Scholar]
- Yonge, C.M. The Sea Shore; Collins: London, UK, 1949. [Google Scholar]
- Dare, P.J.; Edwards, D.B. Underwater television observations on the intertidal movements of shore crabs, Carcinus maenas, across a mudflat. J. Mar. Biol. Assoc. UK 1981, 61, 107–116. [Google Scholar] [CrossRef]
- Green, J. The Biology of Estuarine Animals; Sidgwick and Jackson: London, UK, 1968. [Google Scholar]
- Ingle, R.W. British Crabs; Oxford University Press and British Museum: London, UK, 1980. [Google Scholar]
- Ingle, R.W. Shallow-Water Crabs Synopsis of the British Fauna; Cambridge University Press and the Estuarine and Brackish-water Science Association: Cambridge, UK, 1983. [Google Scholar]
- Elner, R.W. Diet of green crab Carcinus maenas (L.) from Port Hebert, southwestern Nova Scotia. J. Shellfish Res. 1981, 1, 89–94. [Google Scholar]
- Clark, P.F. North-East Atlantic Crabs: An Atlas of Distribution; Marine Conservation Society: Ross-on-Wye, UK, 1986. [Google Scholar]
- Novak, M. Diurnal activity in a group of Gulf of Maine decapods. Crustaceana 2004, 77, 603–620. [Google Scholar] [CrossRef] [Green Version]
- Himes, A.R.; Balschi, W.S.; Pelletier, G.; Frederich, M. Color phase—Specific ion regulation of the European green crab Carcinus maenas in an oscillating salinity environment. J. Shellfish Res. 2017, 36, 465–479. [Google Scholar] [CrossRef]
- MacDonald, A.J.; Kienzle, H.M.; Drolet, D.; Hamilton, D.J. Distribution and habitat use of the invasive Carcinus maenas L. (European green crab) and the native Cancer irroratus (Say) (rock crab) in intertidal zones in the Upper Bay of Fundy, Canada. Northeast. Nat. 2018, 25, 161–180. [Google Scholar] [CrossRef]
- Quinn, B.K. Dramatic decline and limited recovery of a green crab (Carcinus maenas) population in the Minas Basin, Canada after the summer of 2013. PeerJ 2018, 6, e5566. [Google Scholar] [CrossRef] [PubMed]
- Edgell, C.; Hollander, J. The evolutionary ecology of European green crab, Carcinus maenas, in North America. In In the Wrong Place—Alien Marine Crustaceans: Distribution, Biology and Impacts. Invading Nature – Springer Series in Invasion Ecology, Vol. 6; Galil, B.S., Clark, P.F., Carlton, J.T., Eds.; Springer: New York, NY, USA, 2011; pp. 641–659. [Google Scholar]
- Tepolt, C.K.; Somero, G.N. Master of all trades: Thermal acclimation and adaptation of cardiac function in a broadly distributed marine invasive species, the European green crab, Carcinus maenas. J. Exp. Biol. 2014, 217, 1129–1138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahsanullah, M.; Newell, R.C. The effects of humidity and temperature on water loss in Carcinus maenas (L) and Portunus marmoreus (Leach). Comp. Biochem. Physiol. 1977, 56, 593–601. [Google Scholar] [CrossRef]
- Madeira, D.; Narciso, L.; Cabral, H.N.; Vinagre, C.; Diniz, M.S. HSP70 production patterns in coastal and estuarine organisms facing increasing temperatures. J. Sea Res. 2012, 73, 137–147. [Google Scholar] [CrossRef]
- Cuculescu, M.; Hyde, D.; Bowler, K. Thermal tolerance of two species of marine crab, Cancer pagurus and Carcinus maenas. J. Therm. Biol. 1988, 23, 107–110. [Google Scholar] [CrossRef]
- Crisp, D.J. (Ed.) The effects of the severe winter of 1962-63 on marine life in Britain. J. Anim. Ecol. 1964, 33, 165–210. [Google Scholar] [CrossRef]
- Beukema, J.J. The abundance of shore crabs Carcinus maenas (L.) on a tidal flat in the Wadden Sea after cold and mild winters. J. Exp. Mar. Biol. Ecol. 1991, 153, 97–113. [Google Scholar] [CrossRef]
- Dexter, R.W. The marine communities of a tidal inlet at Cape Ann, Massachusetts: A study in bio-ecology. Ecol. Monogr. 1947, 17, 261–294. [Google Scholar] [CrossRef]
- Kienzle, H. Population Dynamics and Interactions between Invasive Green Crabs (Carcinus maenas) and Native Rock Crabs (Cancer irroratus) in Intertidal Zones in the Upper Bay of Fundy. Bachelor’s Thesis, Mount Allison University, Sackville, NB, Canada, 2015; p. 41. [Google Scholar]
- Kelley, A.; De Riviera, C.E.; Buckley, B.A. Cold tolerance of the invasive Carcinus maenas in the east Pacific: Molecular mechanisms and implications for range expansion in a changing climate. Biol. Invasions 2013, 15, 2299–2309. [Google Scholar] [CrossRef]
- Naylor, E. Biological effects of heat effluent in the docks at Swansea. Proc. Zool. Soc. Lond. 1965, 144, 253–268. [Google Scholar] [CrossRef]
- Berrill, M. The life cycle of the green crab Carcinus maenas at the northern end of its range. J. Crustac. Biol. 1982, 2, 31–39. [Google Scholar] [CrossRef]
- McGaw, I.J.; Naylor, E. Distribution and rhythmic locomotor patterns of estuarine and open shore populations of Carcinus maenas. J. Mar. Biol. Assoc. UK 1992, 72, 599–609. [Google Scholar] [CrossRef]
- Atkinson, R.J.A.; Parsons, A.J. Seasonal patterns of migration and locomotion rhythmicity in populations of Carcinus. Neth. J. Sea Res. 1973, 7, 81–93. [Google Scholar] [CrossRef]
- Dries, M.; Adelung, D. Die schlei, ein modell für die verbreitung der strandkrabbe Carcinus maenas. Helgoländer Meeresunter. 1982, 35, 65. [Google Scholar] [CrossRef] [Green Version]
- Hunter, E.; Naylor, E. Intertidal migration by the shore crab Carcinus maenas. Mar. Ecol. Prog. Ser. 1993, 101, 131–138. [Google Scholar] [CrossRef]
- Warman, C.G.; Reid, D.G.; Naylor, E. Variation in the tidal migratory behavior and rhythmic light-responsiveness in the shore crab, Carcinus maenas. J. Mar. Biol. Assoc. UK 1993, 73, 355–364. [Google Scholar] [CrossRef]
- Aagaard, A.; Warman, C.G.; Depledge, M.H. Tidal and seasonal changes in the temporal and spatial distribution of foraging Carcinus maenas in the weakly tidal littoral zone of Kerteminde Fjord, Denmark. Mar. Ecol. Prog. Ser. 1995, 122, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Reid, D.; Abelló, P.; Kaiser, M.; Warman, C. Carapace colour, inter-moult duration and the behavioural and physiological ecology of the shore crab Carcinus maenas. Estuar. Coast. Shelf Sci. 1997, 44, 203–211. [Google Scholar] [CrossRef]
- Audet, D.; Miron, G.; Morivasu, M. Biological characteristics of a newly established green crab population in the southern Gulf of St. Lawrence, Canada. J. Shellfish Res. 2008, 27, 427–441. [Google Scholar] [CrossRef]
- Dawirs, R.R.; Pueschel, C.; Schorn, F. Temperature and growth in Carcinus maenas L. (Decapoda: Portunidae) larvae reared in the laboratory from hatching through metamorphosis. J. Exp. Mar. Biol. Ecol. 1986, 100, 47–74. [Google Scholar] [CrossRef]
- McKnight, A.; Mathews, L.M.; Avery, R.; Lee, K.T. Distribution is correlated with color phase in green crabs, Carcinus maenas (Linnaeus, 1758) in southern New England. Crustaceana 2000, 73, 763–768. [Google Scholar]
- Baeta, A.; Cabral, H.N.; Neto, J.M.; Marques, J.C. Biology, population dynamics and secondary production of the green crab Carcinus maenas (L.) in a temperate estuary. Estuar. Coast. Shelf Sci. 2005, 65, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Dawirs, R.R. Temperature and larval development of Carcinus maenas (Decapoda) in the laboratory; predictions of larval dynamics in the sea. Mar. Ecol. Prog. Ser. 1985, 24, 297–302. [Google Scholar] [CrossRef]
- Nagaraj, M. Combined effects of temperature and salinity on the zoeal development of the green crab, Carcinus maenas (Linnaeus, 1758) (Decapoda: Portunidae). Sci. Mar. 1993, 57, 1–8. [Google Scholar]
- Henry, R.P.; Garrelts, E.E.; McCarty, M.M.; Towle, D.W. Salinity adaptations in the euryhaline green crab, Carcinus maenas. Bull. Mt. Desert Isl. Biol. Lab. 1999, 38, 55. [Google Scholar]
- Perkins, E.J.; Gribbon, E.; Murray, R.B. Some aspects of the biology of Carcinus maenas (L.) II. survival at low salinity. Trans. Dumfries. Galloway Nat. Hist. Antiqu. Soc. 1969, 46, 27–28. [Google Scholar]
- Taylor, A.C. The respiratory responses of Carcinus maenas (L.) to changes in environmental salinity. J. Exp. Mar. Biol. Ecol. 1977, 29, 197–210. [Google Scholar] [CrossRef]
- McGaw, I.J. Behavioural Responses of the Shore Crab Carcinus maenas to Salinity Variation. Ph.D. Thesis, University of Wales, Bangor, UK, 1991. [Google Scholar]
- McGaw, I.; Naylor, E. Salinity preference of the shore crab Carcinus maenas in relation to coloration during intermoult and to prior acclimation. J. Exp. Mar. Biol. Ecol. 1992, 155, 145–159. [Google Scholar] [CrossRef]
- Reid, D.; Abelló, P.; McGaw, I.J.; Naylor, E. Differential tolerances of desiccation and hypo-osmotic stress in sub- and inter-tidal Carcinus maenas. In Phenotypic Responses and Individuality in Aquatic Ectotherms; Aldrich, J.C., Ed.; Japaga Publishers: Wicklow, Ireland, 1989; pp. 89–96. [Google Scholar]
- Lee, K.T.; McKnight, A.; Kellogg, K.; Juanes, F. Salinity tolerance in color phases of female green crabs Carcinus maenas (Linnaeus, 1758). Crustaceana 2003, 76, 247–253. [Google Scholar]
- Styrishave, B.; Andersen, O. Seasonal variations in hepatopancreas fatty acid profiles of two colour forms of shore crabs, Carcinus maenas. Mar. Biol. 2000, 137, 415–422. [Google Scholar] [CrossRef]
- Thomas, N.J.; Lasiak, T.A.; Naylor, E. Salinity preference behavior in Carcinus. Mar. Behav. Physiol. 1981, 7, 277–282. [Google Scholar] [CrossRef]
- McGaw, I.J.; Kaiser, M.J.; Naylor, E.; Hughes, R.N. Intraspecific morphological variation related to the moult-cycle in colour forms of the shore crab Carcinus maenas. J. Zool. Lond. 1992, 228, 351–359. [Google Scholar] [CrossRef]
- Anger, K.; Spivak, E.; Luppi, T. Effects of reduced salinities on development and bioenergetics of early larval shore crab, Carcinus maenas. J. Exp. Mar. Biol. Ecol. 1998, 220, 287–304. [Google Scholar] [CrossRef] [Green Version]
- Lance, J. The salinity tolerance of some estuarine planktonic crustaceans. Biol. Bull. 1964, 127, 108–118. [Google Scholar] [CrossRef]
- Darbyson, E.A. Local Vectors of Spread of the Green Crab (Carcinus maenas) and the Clubbed Tunicate (Styela Clava) in the Southern Gulf of St. Lawrence, Canada. Master’s Thesis, University of Halifax, Halifax, NS, Canada, 2006. [Google Scholar]
- Bohn, G. Sur le renversement du courant respiratoire chez les Décapodes. CR. Acad. Sci. 1897, 125, 539–542. [Google Scholar]
- Reid, D.G.; Aldrich, J.C. Variation in responses to environmental hypoxia of different colour forms of the shore crab Carcinus maenas (L.). Comp. Biochem. Physiol. 1989, 92, 535–539. [Google Scholar] [CrossRef]
- Taylor, A.C.; Butler, P.J. The behavior and physiological responses of the shore crab Carcinus maenas during changes in environmental oxygen tension. Neth. J. Sea Res. 1973, 7, 496–505. [Google Scholar] [CrossRef]
- Aldrich, J.C. The influence of individual variations in metabolic rate and tidal conditions on the response to hypoxia in Carcinus maenas (L.). Comp. Biochem. Physiol. 1986, 83, 53–60. [Google Scholar] [CrossRef]
- Taylor, A.C. The respiratory responses of Carcinus maenas to declining oxygen tension. J. Exp. Biol. 1976, 65, 309–322. [Google Scholar]
- Jungblut, S.; Boos, K.; McCarthy, M.L.; Saborowski, R.; Hagen, W. Invasive versus native brachyuran crabs in a European rocky intertidal: Respiratory performance and energy expenditures. Mar. Biol. 2018, 165, 54. [Google Scholar] [CrossRef] [Green Version]
- McMahon, B.R. Physiological responses to oxygen depletion in intertidal animals. Am. Zool. 1988, 28, 39–53. [Google Scholar] [CrossRef]
- Souza, A.T.; Ilarri, M.I.; Campos, J.; Marques, J.C.; Martins, I. Differences in the neighborhood: Structural variations in the carapace of shore crabs Carcinus maenas (Decapoda: Portunidae). Estuar. Coast. Shelf Sci. 2011, 95, 424–430. [Google Scholar] [CrossRef]
- Wolf, F. Red and green colour forms in the common shore crab Carcinus maenas (L.) (Crustacea: Brachyura: Portunidae): Theoretical predictions and empirical data. J. Nat. Hist. 1998, 32, 1807–1812. [Google Scholar] [CrossRef]
- Styrishave, B.; Rewitz, K.; Andersen, O. Frequency of moulting by shore crabs Carcinus maenas (L.) changes their colour and their success in mating and physiological performance. J. Exp. Mar. Biol. Ecol. 2004, 313, 317–336. [Google Scholar] [CrossRef]
- Styrishave, B.; Aagaard, A.; Andersen, O. In situ studies on physiology and behavior in two colour forms of the shore crab Carcinus maenas in relation to season. Mar. Ecol. Prog. Ser. 1999, 189, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Rewitz, K.; Styrishave, B.; Depledge, M.H.; Andersen, O. Spatial and temporal distribution of shore crabs Carcinus maenas in a small tidal estuary (Looe Estuary), Cornwall, England. J. Crustac. Biol. 2004, 24, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, M.J.; Hughes, R.N.; Reid, D.G. Chelal morphometry, prey-size selection and aggressive competition in green and red forms of Carcinus maenas (L.). J. Exp. Mar. Biol. Ecol. 1990, 140, 121–134. [Google Scholar] [CrossRef]
- Lee, K.T.; Vespoli, J.L. Tracking color change in individual green crabs, Carcinus maenas (L.). Northeast. Nat. 2015, 22, 413–423. [Google Scholar] [CrossRef]
- Queiroga, H. An analysis of the size structure of Carcinus maenas (L.)(Decapoda, Brachyura) from Canal de Mira, Ria de Aveiro, Portugal. Biosis 1993, 1, 89–106. [Google Scholar]
- Poirier, L.A.; Mohan, J.; Speare, R.; Davidson, J.; Quijón, P.A.; St-Hilaire, S. Moulting synchrony in green crabs (Carcinus maenas) from Prince Edward Island, Canada. Mar. Biol. Res. 2016, 12, 969–977. [Google Scholar] [CrossRef]
- Fulton, B.A.; Fairchild, E.A.; Warner, R. Carcinus maenas in two New Hampshire estuaries. Part 2: Assessment of average intermolt period. J. Crustac. Biol. 2013, 33, 339–347. [Google Scholar] [CrossRef] [Green Version]
- McVean, A.; Findlay, I. The incidence of autotomy in an estuarine population of the crab Carcinus maenas. J. Mar. Biol. Assoc. UK 1979, 59, 341–354. [Google Scholar] [CrossRef]
- Bliss, D.E. Autotomy and regeneration. In Physiology of Crustacea, Vol. 1; Waterman, T.H., Ed.; Academic Press: New York, NY, USA, 1961; pp. 561–589. [Google Scholar]
- Sekkelsten, G.I. Effect of handicap on mating success in male shore crabs Carcinus maenas. Oikos 1988, 51, 131–134. [Google Scholar] [CrossRef]
- Brian, J.V. Inter-Population Variability in the Shore Crab (Carcinus maenas L.) and Its Potential Use as a Biomarker of Anthropogenic Effects. Ph.D. Thesis, Napier University, Edinburgh, UK, 2002. [Google Scholar]
- van der Meeren, G.I. Sex- and size-dependent mating tactics in a natural population of shore crabs Carcinus maenas. J. Anim. Ecol. 1994, 63, 307–314. [Google Scholar] [CrossRef]
- d’Acoz, C.D.U. Activités reproductrices saisonnieres des differentes classe de tailes d’une population de crabs verts Carcinus maenas (Linnaeus, 1758) dans le sud de la mer du Nord. Cah. Biol. Mar. 1993, 35, 1–13. [Google Scholar]
- van der Meeren, G.I.; Van der Meeren, C.I. Location of spawning shore crabs, Carcinus maenas (L., 1758) (Decapoda, Brachyura). Crustaceana 1992, 63, 92–94. [Google Scholar] [CrossRef]
- Williamson, H.C. On the larval and early young stages, and rate of growth, of the shore crab (Carcinus maenas Leach). Rep. Fish. Board. Scotl. 1903, 21, 136–139. [Google Scholar]
- Rasmussen, E. Behaviour of sacculinized shore crabs (Carcinus maenas Pennant). Nature 1959, 183, 479–480. [Google Scholar] [CrossRef]
- Wheatly, M.G. The provision of oxygen to developing eggs by female shore crabs (Carcinus maenas). J. Mar. Biol. Assoc. UK 1981, 61, 117–128. [Google Scholar] [CrossRef]
- Almaça, C. Note sur la biologie des populations de Carcinus maenas (L.) de la zone intertidale du Portugal occidental. Quad. Lab. Tecnol. Pesca 1982, 3, 179–185. [Google Scholar]
- Sprung, M. Larval abundance and recruitment of Carcinus maenas L. close to its southern geographic limit: A case of match and mismatch. In Advances in Decapod Crustacean Research; Springer: Berlin/Heidelberg, Germany, 2001; pp. 153–158. [Google Scholar]
- Queiroga, H.; Costlow, J.D.; Moreira, M.H. Larval abundance patterns of Carcinus maenas (Decapoda, Brachyura) in Canal de Mira (Ria de Aveiro, Portugal). Mar. Ecol. Prog. Ser. 1994, 111, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Best, K.; McKenzie, C.H.; Couturier, C. Reproductive biology of an invasive population of European green crab, Carcinus maenas, in Placentia Bay, Newfoundland. Manag. Biol. Invasions 2017, 8, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Sharp, G.; Semple, R.; Connolly, K.; Blok, R.; Audet, D.; Cairns, D.; Courtenay, S. Ecological assessment of the Basin Head Lagoon: A proposed Marine Protected Area. Can. Manuscr. Rep. Fish. Aquat. Sci. 2003, 2641, 76. [Google Scholar]
- Gillespie, G.E.; Phillips, A.C.; Paltzat, D.L.; Therriault, T.W. Status of the European green crab, Carcinus maenas, in British Columbia—2006. Can. Tech. Rep. Fish. Aquat. Sci. 2007, 2700, 39. [Google Scholar]
- Mabin, C.; Wilson, J.R.U.; Le Roux, J.J.; Robinson, T.B. Reassessing the invasion of South African waters by the European shore-crab Carcinus maenas. Afr. J. Mar. Sci. 2017, 39, 259–267. [Google Scholar] [CrossRef]
- Lyons, J.L.; O’Riordan, M.R.; Cross, F.T.; Culloty, C.S. Reproductive biology of the shore crab Carcinus maenas (Decapoda, Portunidae): A macroscopic and histological view. Invertebr. Reprod. Dev. 2012, 56, 144–156. [Google Scholar] [CrossRef]
- Hines, A.H.; Ruiz, G.M.; Hitchcock, N.G.; deRivera, C. Projecting Range Expansion of Invasive European Green Crabs (Carcinus maenas) to Alaska: Temperature and Salinity Tolerance of Larvae; Prince William Sound Regional Citizens’ Advisory Council: Anchorage, AK, USA, 2004; p. 22. [Google Scholar]
- Flores, A.A.V.; Gomes, C.C.; Villano, W.F. Source populations in coastal crabs: Parameters affecting egg production. Aquat. Biol. 2009, 7, 31–43. [Google Scholar] [CrossRef]
- Bate, C.S. On the development of Decapoda Crustacea. Philos. Trans. R. Soc. Lond. 1858, 148, 2–589. [Google Scholar]
- Griffen, B.D. Linking individual diet variation and fecundity in an omnivorous marine consumer. Oecologia 2014, 174, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Klein Breteler, W. Laboratory experiments on the influence of environmental factors on the frequency of moulting and the increase in size at moulting of juvenile shore crabs, Carcinus maenas. Neth. J. Sea Res. 1975, 9, 100–120. [Google Scholar] [CrossRef]
- Starr, M.; Himmelman, J.H.; Therriault, J.-C. Direct coupling of marine invertebrate spawning with phytoplankton blooms. Science 1990, 247, 1071–1074. [Google Scholar] [CrossRef] [PubMed]
- Mohamedeen, H.; Hartnoll, R.G. Larval and postlarval growth of individually reared specimens of the common shore crab Carcinus maenas (L.). J. Exp. Mar. Biol. Ecol. 1989, 134, 1–24. [Google Scholar] [CrossRef]
- Epifanio, C.E.; Cohen, J.H. Behavioral adaptations in larvae of brachyuran crabs; a review. J. Exp. Mar. Biol. Ecol. 2016, 482, 85–105. [Google Scholar] [CrossRef]
- Rasmussen, E. Systematics and ecology of the Isefjord marine fauna (Denmark) with a survey of the eelgrass (Zostera) vegetation and its communities. Ophelia 1973, 11, 1–507. [Google Scholar] [CrossRef]
- Zeng, C.; Naylor, E. Occurrence in coastal water and endogenous tidal swimming rhythms of late megalopae of the shore crab Carcinus maenas: Implications for onshore recruitment. Mar. Ecol. Prog. Ser. 1966, 136, 69–79. [Google Scholar] [CrossRef]
- Queiroga, H. Processos de dispersão e recrutamento das larvas do caranguejo Carcinus maenas (L.) na Ria de Aveiro. Ph.D. Thesis, Universidade de Aveiro, Aveiro, Portugal, 1995. [Google Scholar]
- Queiroga, H. Distribution and drift of the crab Carcinus maenas (L.) (Decapoda, Portunidae) larvae over the continental shelf off northern Portugal in April 1991. J. Plank. Res. 1996, 18, 1981–2000. [Google Scholar] [CrossRef] [Green Version]
- Zeng, C.; Naylor, E. Endogenous tidal rhythms of vertical migration in field collected zoea-1 larvae of the shore crab Carcinus maenas: Implications for ebb tide offshore dispersal. Mar. Ecol. Prog. Ser. 1966, 132, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Crowe, T.P.; Underwood, A.J. Testing behavioural “preferences” for suitable microhabitat. J. Exp. Mar. Biol. Ecol. 1998, 225, 1–11. [Google Scholar] [CrossRef]
- Therriault, T.W.; Herborg, L.M.; Locke, A.; McKindsey, C.W. Risk Assessment for European Green Crab (Carcinus maenas) in Canadian Waters; Fisheries and Oceans Canada, Science Branch: Nanaimo, BC, Canada, 2008. [Google Scholar]
- Orton, J.H. Experiments in the sea on the rate of growth of some Crustacea Decapoda. J. Mar. Biol. Assoc. UK 1936, 20, 673–689. [Google Scholar] [CrossRef] [Green Version]
- Lützen, J. Growth, reproduction, and life span in Sacculina carcini Thompson (Cirripedia: Rhizocephala) in the Isefjord, Denmark. Sarsia 1984, 69, 91–105. [Google Scholar] [CrossRef]
- Behrens Yamada, S.; Dumbauld, B.R.; Kalin, A.; Hunt, C.E.; Figlar-Barnes, R.; Randall, A. Growth and persistence of a recent invader Carcinus maenas in estuaries of the northeastern Pacific. Biol. Invasions 2005, 7, 309–321. [Google Scholar] [CrossRef]
- Werner, M. Prevalence of the parasite Sacculina carcini Thompson 1836 (Crustacea, Rhizocephala) on its host crab Carcinus maenas (L.) on the west coast of Sweden. Ophelia 2001, 55, 101–110. [Google Scholar] [CrossRef]
- Munch-Petersen, S.; Sparre, P.; Hoffman, E. Abundance of the shore crab, Carcinus maenas (L) estimated from mark-recapture experiments. Dana 1982, 2, 97–121. [Google Scholar]
- Heath, J.R.; Barnes, H. Some changes in biochemical composition with season and during the moulting cycle of the common shore crab, Carcinus maenas (L.). J. Exp. Mar. Biol. Ecol. 1970, 5, 199–233. [Google Scholar] [CrossRef]
- Garm, A. Mechanosensory properyies of the mouthpart setae of the European shore crab Carcinus maenas. Mar. Biol. 2005, 147, 1179–1190. [Google Scholar] [CrossRef]
- Pedersen, K.L.; Pedersen, S.N.; Højrup, P.; Andersen, J.S.; Roepstorff, P.; Knudsen, J.; Depledge, M.H. Purification and characterization of a cadmium-induced metallothionein from the shore crab Carcinus maenas (L.). Biochem. J. 1994, 297, 609–614. [Google Scholar] [CrossRef] [Green Version]
- Sneddon, L.U.; Huntingford, F.A.; Taylor, A.C. Weapon size versus body size as a predictor of winning in fights between shore crabs, Carcinus maenas (L.). Behav. Ecol. Sociobiol. 1997, 41, 237–242. [Google Scholar] [CrossRef] [Green Version]
- Abelló, P.; Aagaard, A.; Warman, C.G.; Depledge, M.H. Spatial variability in the population structure of the shore crab Carcinus maenas (Crustacea: Brachyura) in a shallow-water, weakly tidal fjord. Mar. Ecol. Prog. Ser. 1997, 147, 97–103. [Google Scholar] [CrossRef]
- McVean, A. The incidence of autotomy in Carcinus maenas (L.). J. Exp. Mar. Biol. Ecol. 1976, 24, 177–187. [Google Scholar] [CrossRef]
- Fletcher, N.; Hardege, J.D. The cost of conflict: Agonistic encounters influence responses to chemical signals in the European shore crab. Anim. Behav. 2009, 77, 357–361. [Google Scholar] [CrossRef]
- Walne, P.R.; Dean, G.J. Experiments on predation by the shore crab Carcinus maenas on Mytilus and Mercenaria. ICES J. Mar. Sci. 1972, 34, 190–199. [Google Scholar] [CrossRef]
- Elner, R.W. The mechanics of predation by the shore crab, Carcinus maenas (L.), on the edible mussel, Mytilus edulis L. Oecologia 1978, 36, 333–344. [Google Scholar] [CrossRef]
- Elner, R.W.; Hughes, R.N. Energy maximization in the diet of the shore crab, Carcinus maenas. J. Anim. Ecol. 1978, 47, 103–116. [Google Scholar] [CrossRef]
- Huxley, J.S.; Richards, O.W. Relative growth of the abdomen and the carapace of the shore-crab Carcinus maenas. J. Mar. Biol. Assoc. UK 1931, 17, 1001–1015. [Google Scholar] [CrossRef] [Green Version]
- Kamermans, P.; Blankendaal, M.; Perdon, J. Predation of shore crabs (Carcinus maenas (L.)) and starfish (Asterias rubens L.) on blue mussel (Mytilus edulis L.) seed from wild sources and spat collectors. Aquaculture 2009, 290, 256–262. [Google Scholar] [CrossRef]
- Stevens, M.; Lown, A.E.; Wood, L.E. Camouflage and individual variation in shore crabs (Carcinus maenas) from different habitats. PLoS ONE 2014, 9, e115586. [Google Scholar] [CrossRef]
- Spooner, E.H.; Coleman, R.A.; Attrill, M.J. Sex differences in body morphology and multitrophic interactions involving the foraging behavior of the crab Carcinus maenas. Mar. Ecol. 2007, 28, 394–403. [Google Scholar] [CrossRef]
- Tremblay, M.J.; Thompson, A.; Paul, K. Recent Trends in the Abundance of the Invasive Green Crab (Carcinus maenas) in Bras d’Or Lakes and Eastern Nova Scotia Based on Trap Surveys; Fisheries and Ocean Canada, Bedford Institute of Oceanography: Dartmouth, NS, Canada, 2006. [Google Scholar]
- Tummon Flynn, P.; Lynn, K.D.; Cairns, D.K.; Quijon, P.A. The role of the non-indigenous green crab (Carcinus maenas) in the decline of a unique strain of Irish moss (Chondrus crispus): Direct and indirect effects. ICES J. Mar. Sci. 2019. [Google Scholar] [CrossRef]
- Rayner, G. The Behavioural Interactions between the American Lobster (Homarus Americanus) and the Invasive Green Crab (Carcinus maenas). Ph.D. Thesis, Memorial University of Newfoundland, St. John’s, NL, Canada, 2018. [Google Scholar]
- Rayner, G.; McGaw, I.J. Effects of the invasive green crab (Carcinus maenas) on American lobster (Homarus americanus): Food acquisition and trapping behavior. J. Sea Res. 2019, 144, 95–104. [Google Scholar] [CrossRef]
- Aman, J.; Grimes, K.W. Measuring Impacts of Invasive European Green Crabs on Maine Salt Marshes: A Novel Approach. In Report to the Maine Outdoor Heritage Fund; Wells National Estuarine Research Reserve: Wells, ME, USA, 2016; p. 19. [Google Scholar]
- Jamieson, G.S.; Foreman, M.G.G.; Cherniawsky, J.Y.; Levings, C.D. European green crab (Carcinus maenas) dispersal: The Pacific experience. In Crabs in Cold Water Regions: Biology, Management and Economics; Paul, A.J., Dawe, E.G., Elner, R., Jamieson, G.S., Kruse, G.H., Otto, R.S., Sainte-Marie, B., Shirley, T.C., Woodby, D., Eds.; College Program AK-SG-02-01; University of Alaska Sea Grant: College, AK, USA, 2002; pp. 561–576. [Google Scholar]
- Kelley, A.L.; deRivera, C.E.; Grosholz, E.D.; Ruiz, G.M.; Behrens Yamada, S.; Gillespie, G. Thermogeographic variation in body size of Carcinus maenas, the European green crab. Mar. Biol. 2015, 162, 1625–1635. [Google Scholar] [CrossRef]
- DiBacco, C.; Therriault, T.W. Reproductive periodicity and larval vertical migration behavior of European green crab Carcinus maenas in a non-native habitat. Mar. Ecol. Prog. Ser. 2015, 536, 123–134. [Google Scholar] [CrossRef]
- Hidalgo, F.J.; Baron, P.J.; Orensanz, J.M. A prediction come true: The green crab invades the Patagonian coast. Biol. Invasions 2005, 7, 547–552. [Google Scholar] [CrossRef]
- Campbell, R.T.; Baring, R.J.; Dittmann, S. Cracking the cuisine: Invasive European shore crabs (Carcinus maenas) select a menu of soft-shelled mussels over cockles. J. Exp. Mar. Biol. Ecol. 2019, 517, 25–33. [Google Scholar] [CrossRef]
- Jensen, G.C.; McDonald, P.S.; Armstrong, D.A. Biotic resistance to green crab, Carcinus maenas, in California bays. Mar. Biol. 2007, 151, 2231–2243. [Google Scholar] [CrossRef]
- Behrens Yamada, S.; Gillespie, G.E. Will the European green crab (Carcinus maenas) persist in the Pacific Northwest? ICES J. Mar. Sci. 2008, 65, 725–729. [Google Scholar] [CrossRef] [Green Version]
- Grosholz, E.D.; Ruiz, G.M. Biological invasions drive size increases in marine and estuarine invertebrates. Ecol. Lett. 2003, 6, 700–705. [Google Scholar] [CrossRef]
- Bloch, C.P.; Curry, K.D.; Jahoda, J.C. Long-term effects of an invasive shore crab on Cape Cod, Massachusetts. Northeast. Nat. 2015, 22, 178–191. [Google Scholar] [CrossRef] [Green Version]
- Scherer, B.; Reise, K. Significant predation on micro- and macrobenthos by the crab Carcinus maenas L. in the Wadden Sea. Kiel Meeresforsch Sonderh 1981, 5, 490–500. [Google Scholar]
- Jackson, D.; Mason, C.F.; Long, S.P. Macro-invertebrate populations and production on a salt-marsh in east England dominated by Spartina anglica. Oecologia 1985, 65, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.J. Effectiveness of crab and lobster traps. Can. J. Fish. Aquat. Sci. 1990, 47, 1228–1251. [Google Scholar] [CrossRef]
- Grosholz, E.D.; Ruiz, G.M.; Dean, C.A.; Shirley, K.A.; Maron, J.L.; Connors, P.G. The impacts of a nonindigenous marine predator in a California bay. Ecology 2000, 81, 1206–1224. [Google Scholar] [CrossRef]
- Miller, R.J. Saturation of crab traps: Reduced entry and escapement. ICES J. Mar. Sci. 1979, 38, 338–345. [Google Scholar] [CrossRef]
- Bergshoeff, J.A.; McKenzie, C.H.; Best, K.; Zargarpour, N.; Favaro, B. Using underwater video to evaluate the performance of the Fukui trap as a mitigation tool for the invasive European green crab (Carcinus maenas) in Newfoundland, Canada. PeerJ 2018, 6, e4223. [Google Scholar] [CrossRef] [Green Version]
- Bergshoeff, J.A.; McKenzie, C.H.; Favaro, B. Improving the efficiency of the Fukui trap as a capture tool for the invasive European green crab (Carcinus maenas) in Newfoundland, Canada. PeerJ 2019, 7, e6308. [Google Scholar] [CrossRef] [Green Version]
- Waser, A.M.; Dekker, R.; Witte, J.I.; McSweeney, N.; Ens, B.J.; van der Meer, J. Quantifying tidal movements of the shore crab Carcinus maenas on to complex epibenthic bivalve habitats. Estuaries Coasts 2018, 41, 507–520. [Google Scholar] [CrossRef] [Green Version]
- Edwards, E. Observations on growth of the edible crab Cancer pagurus. Rapports et Procés-Verbaux des Réunions 1965, 156, 62–70. [Google Scholar]
- Therriault, T. European green crab in British Columbia—Population and impact. In Proceedings of the Regional Advisory Process on Green Crab, Carcinus maenas, Populations and Mitigations in the Newfoundland and Labrador Region, Canada, 17 March 2010; Can. Sci. Advis. Sec. Proceed. Ser. 2011/020; Clovelly Golf Club: St. John’s, NL, Canada, 2011; pp. 5–7. [Google Scholar]
- Mathieson, S.; Berry, A.J. Spatial, temporal and tidal variation in crab populations in the Forth Estuary, Scotland. J. Mar. Biol. Assoc. UK 1997, 77, 167–183. [Google Scholar] [CrossRef]
- Grundstrom, J.; McAneney, B.; Fregeau, M.; Weston, S.; Buttner, J.; Walton, W.C.; Murphy, D.C. Characterization of Green Crab (Carcinus maenas) Populations in Coastal Waters of Massachusetts; 2004; p. 16, Unpublished report. [Google Scholar]
- Aagaard, A. In situ variation in heart rate of the shore crab Carcinus maenas in relation to environmental factors and physiological condition. Mar. Biol. 1996, 125, 765–772. [Google Scholar] [CrossRef]
- Baeta, A.; Cabral, H.N.; Marques, J.C.; Pardal, M.A. Feeding ecology of the green crab, Carcinus maenas (L., 1758) in a temperate estuary, Portugal. Crustaceana 2006, 79, 1181–1193. [Google Scholar]
- McGaw, I.J.; (Memorial University, St. John’s, NL, Canada). Personal communication, 2018.
- McKenzie, C. Ecological assessment of the invasive European green crab, Carcinus maenas, in Newfoundland: 2007–2009 population dynamics ecological impact. In Proceedings of the Regional Advisory Process on Green Crab, Carcinus maenas, Populations and Mitigations in the Newfoundland and Labrador Region, Canada, 17 March 2010; Can. Sci. Advis. Sec. Proceed. Ser. 2011/020; Clovelly Golf Club: St. John’s, NL, Canada, 2011; pp. 1–3. [Google Scholar]
- Goldstein, J.S.; Morrissey, E.M.; Moretti, E.D.; Watson, W.H. A comparison of the distribution and abundance of European green crabs and American lobsters in the Great Bay Estuary, New Hampshire, USA. Fish. Res. 2017, 189, 10–17. [Google Scholar] [CrossRef]
- Young, T.; Komarow, S.; Deegan, L.; Garritt, R. Population size and summer home range of the green crab, Carcinus maenas, in salt marsh tidal creeks. Biol. Bull. 1999, 197, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Gee, J.M.; Warwick, R.M.; Davey, J.T.; George, C.L. Field experiments on the role of epibenthic predation in determining prey densities in an estuarine mudflat. Estuar. Coast. Shelf Sci. 1985, 21, 439–448. [Google Scholar] [CrossRef]
- Raffaelli, D.; Conacher, A.; McLaclan, H.; Emes, C. The role of epibenthic crustacean predators in an estuarine food web. Estuar. Coast. Shelf Sci. 1989, 28, 149–160. [Google Scholar] [CrossRef]
- Zwarts, L.; Wanink, J.H. How the food supply harvestable by waders in the Wadden Sea depends on the variation in energy density, body weight, biomass, burying depth and behavior of tidal-flat invertebrates. Neth. J. Sea Res. 1993, 31, 441–476. [Google Scholar] [CrossRef]
- Ameyaw-Akumfi, C.; Hughes, R.N. Behaviour of Carcinus maenas feeding on large Mytilus edulis. How do they assess the optimum diet? Mar. Ecol. Prog. Ser. 1987, 38, 213–216. [Google Scholar] [CrossRef]
- Smith, O.R.; Chin, E. The effects of predation on soft clams, Mya arenaria. Natl Shellfish. Assoc. Conv. Address 1951, 42, 37–44. [Google Scholar]
- MacPhail, J.S.; Lord, E.I.; Dickie, L.M. The green crab—A new clam enemy. Fish. Res. Bull. Can. Atl. Prog. Rep. 1955, 63, 3–12. [Google Scholar]
- Jensen, K.T.; Jensen, J.N. The importance of some epibenthic predators on the density of juvenile benthic macrofauna in the Danish Wadden Sea. J. Exp. Mar. Biol. Ecol. 1985, 89, 157–174. [Google Scholar] [CrossRef]
- Beal, B.F. Evaluating factors contributing to mortalities of juveniles of the soft-shell clam (Mya arenaria L.) in Hampton/Seabrook Harbor, New Hampshire. Final Rep. New Hamps. Estuaries Proj. 2002, 111p. Available online: http://scholars.unh.edu/cgi/viewcontent.cgi?article=1315context=prep (accessed on 27 December 2017).
- Tyrell, M.; Harris, L.G. Potential impact of the introduced Asian shore crab, Hemigrapsus sanguineus, in northern New England: Diet, feeding preferences, and overlap with the green crab, Carcinus maenas. In Marine Bioinvasions, Proceedings of the First National Conference, Cambridge, MA, USA, 24–27 January 1999; Pederson, Ed.; MIT SeaGrant College Program: Cambridge, MA, USA, 1999; pp. 208–220. [Google Scholar]
- DeGraaf, J.D.; Tyrrell, M.C. Comparison of the feeding rates of two introduced crab species, Carcinus maenas and Hemigrapsus sanguineus, on the blue mussel, Mytilus edulis. Northeast. Nat. 2004, 11, 163–166. [Google Scholar] [CrossRef]
- Whitlow, W.L. Changes in survivorship, behavior, and morphology in native soft-shell clams induced by invasive green crab predators. Mar. Ecol. 2010, 31, 418–430. [Google Scholar] [CrossRef]
- Department of Fisheries and Oceans (DFO). Ecological Assessment of the Invasive European Green Crab (Carcinus maenas) in Newfoundland 2007–2009. In DFO Canadian Science Advisory Secretariat Science Advisory Report 2010/033; DFO: St. John’s, NL, Canada, 2011; p. 10. [Google Scholar]
- Trussell, G.C.; Ewanchuk, P.J.; Bertness, M.D.; Silliman, B.R. Trophic cascades in rocky shore tide pools: distinguishing lethal and nonlethal effects. Oecologia 2004, 139, 427–432. [Google Scholar] [CrossRef]
- Sigurdsson, G.M.; Rochette, R. Predation by green crab and sand shrimp on settling and recently settled American lobster postlarvae. J. Crustac. Biol. 2013, 33, 10–14. [Google Scholar] [CrossRef] [Green Version]
- Gregory, G.J.; Quijon, P.A. The impact of a coastal invasive predator on infaunal communities: Assessing the roles of density and a native counterpart. J. Sea Res. 2011, 66, 181–186. [Google Scholar] [CrossRef]
- Le Calvez, J.C. Location of the shore crab Carcinus maenas L., in the food web of a managed estuary ecosystem: The Rance Basin (Brittany, France). Investigación Pesquera 1987, 51 (Suppl. 1), 431–442. [Google Scholar]
- Ruiz, G.M.; Rodriguez, L. Preliminary evaluation and predictions of impacts of Carcinus maenas on native crabs in Tasmania. In Proceedings of the First International Workshop on the Demography, Impacts and Management of Introduced Populations of the European Crab, Carcinus maenas, Hobart, Australia, 20–21 March 1997; Thresher, R.E., Ed.; Technical Report 11. Centre for Research on Introduced Marine Pests: Hobart, TAS, Australia, 1997; pp. 51–55. [Google Scholar]
- Griffen, B.D.; Guy, T.; Buck, J.C. Inhibition between invasives: A newly introduced predator moderates the impacts of a previously established invasive predator. J. Anim. Ecol. 2008, 77, 32–40. [Google Scholar] [CrossRef]
- Wallace, J.C. Feeding, starvation and metabolic rate in the shore crab Carcinus maenas. Mar. Biol. 1973, 20, 277–281. [Google Scholar] [CrossRef]
- Crothers, J.H. Dale Fort Marine Fauna. Field Stud. 1966, 2, 169. [Google Scholar]
- Crothers, J.H. The distribution of crabs on rocky shores around the Dale Peninsula. Field. Stud. 1970, 23, 263–274. [Google Scholar]
- Mascaró, M.; Seed, R. Foraging behavior of juvenile Carcinus maenas (L.) and Cancer pagurus L. Mar. Biol. 2001, 139, 1135–1145. [Google Scholar]
- van den Brink, A.; Hutting, S. Clash of the crabs: Interspecific, inter-cohort competition between the native European green crab, Carcinus maenas and the exotic brush clawed crab Hemigrapsus takanoi on artificial oyster reefs. J. Sea Res. 2017, 128, 41–51. [Google Scholar] [CrossRef]
- Schubart, C.D. The East Asian shore crab Hemigrapsus sanguineus (Brachyura: Varunidae) in the Mediterranean Sea: An independent human-mediated introduction. Sci. Mar. 2003, 67, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Dauvin, J.C.; Rius, A.T.; Ruellet, T. Recent expansion of two invasive crab species Hemigrapsus sanguineus (de Haan, 1835) and H. takanoi Asakura and Wataabe 2005 along the Opal Coast, France. Aquat. Invasions 2009, 4, 451–465. [Google Scholar] [CrossRef] [Green Version]
- Miron, G.; Audet, D.; Landry, T.; Moriyasu, M. Predation potential of the invasive green crab (Carcinus maenas) and other common predators on commercial bivalve species found on Prince Edward Island. J. Shellfish Res. 2005, 24, 579–586. [Google Scholar]
- Sungail, J.; Brown, A.C.; Alpert, K.; Maurukas, J. Prey selection by Gulf of Maine green crabs (Carcinus maenas), rock crabs (Cancer irroratus) and American lobsters (Homarus americanus): A laboratory study. J. Exp. Mar. Biol. Ecol. 2013, 449, 294–303. [Google Scholar] [CrossRef]
- Matheson, K.; Gagnon, P. Effects of temperature, body size, and chela loss on competition for a limited food resource between indigenous rock crab (Cancer irroratus Say) and recently introduced green crab (Carcinus maenas L.). J. Exp. Mar. Biol. Ecol. 2012, 428, 49–56. [Google Scholar] [CrossRef]
- Donahue, M.J.; Nichols, A.; Santamaria, C.A.; League-Pike, P.E.; Krediet, C.J.; Perez, K.O.; Shulman, M.J. Predation risk, prey abundance, and the vertical distribution of three brachyuran crabs on Gulf of Maine shores. J. Crustac. Biol. 2009, 29, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Belair, M.-C.; Miron, G. Predation behavior of Cancer irroratus and Carcinus maenas during conspecific and heterospecific challenges. Aquat. Biol. 2009, 6, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Breen, E.; Metaxas, A. Effects of juvenile non-indigenous Carcinus maenas on the growth and condition of juvenile Cancer irroratus. J. Exp. Mar. Biol. Ecol. 2009, 377, 12–19. [Google Scholar] [CrossRef]
- Williams, P.J.; Floyd, T.A.; Rossong, M.A. Agonistic interactions between invasive green crabs, Carcinus maenas (Linnaus), and sub-adult American lobsters, Homarus americanus (Milne Edwards). J. Exp. Mar. Biol. Ecol. 2006, 329, 66–74. [Google Scholar] [CrossRef]
- Rossong, M.A.; Williams, P.J.; Comeau, M.; Mitchell, S.C.; Apaloo, J. Agonistic interactions between the invasive green crab, Carcinus maenas (Linnaeus) and juvenile American lobster, Homarus americanus (Milne Edwards). J. Exp. Mar. Biol. Ecol. 2006, 329, 281–288. [Google Scholar] [CrossRef]
- Rossong, M.A.; Quijon, P.A.; Williams, P.J.; Snelgrove, P.V.R. Foraging and shelter behavior of juvenile American lobster (Homarus americanus): The influence of a non-indigenous crab. J. Exp. Mar. Biol. Ecol. 2011, 403, 75–80. [Google Scholar] [CrossRef]
- Haarr, M.L.; Rochette, R. The effect of geographic origin on interactions between adult invasive green crabs carcinus maenas and juvenile American lobsters Homarus americanus in Atlantic Canada. J. Exp. Mar. Biol. Ecol. 2012, 422, 88–100. [Google Scholar] [CrossRef]
- McDonald, P.S.; Jensen, G.C.; Armstrong, D.A. The competitive and predatory impacts of the nonindigenous crab Carcinus maenas (L.) on early benthic phase Dungeness crab Cancer magister Dana. J. Exp. Mar. Biol. Ecol. 2001, 258, 39–54. [Google Scholar] [CrossRef]
- McDonald, P.S.; Jensen, G.C.; Armstrong, D.A. Green crabs and native predators; possible limitations on the west coast invasion. (Abstract). J. Shellfish Res. 1998, 17, 1283. [Google Scholar]
- MacDonald, J.A.; Roudez, R.; Glover, T.; Weis, J.S. The invasive green crab and Japanese shore crab: Behavioral interactions with a native crab species, the blue crab. Biol. Invasions 2007, 9, 837–848. [Google Scholar] [CrossRef]
- Ahi, R.S.; Moss, S.P. Status of the nonindigenous crab, Hemigrapsus sanguineus, at Greenwich Point, Connecticut. Northeast. Nat. 1999, 6, 221–224. [Google Scholar]
- Lohrer, A.M.; Whitlatch, R.B. Interactions among aliens: Apparent replacement of one exotic species by another. Ecology 2002, 83, 719–732. [Google Scholar] [CrossRef]
- Lohrer, A.M.; Whitlatch, R.B. Ecological studies on the recently introduced Japanese shore crab (Hemigrapsus sanguineus), in eastern Long Island Sound. In Proceedings of the Second Northeast Conference on Nonindigenous Aquatic Nuisance Species, Burlington, VT, USA, 18–19 April 1997; Balcoln, N., Ed.; Connecticut Sea Grant College Program CTSG-97-02. University of Connecticut, Avery Point: Groton, CT, USA, 1997; pp. 49–60. [Google Scholar]
- McDermott, J.J. The western Pacific brachyuran (Hemigrapsus sanguineus: Grapsidae), in its new habitat along the Atlantic coast of the United States: Geographic distribution and ecology. ICES J. Mar. Sci. 1998, 55, 289–298. [Google Scholar] [CrossRef]
- Gerard, V.A.; Cerrato, R.M.; Larson, A.A. Potential impacts of a grapsid crab on intertidal communities of the northwestern Atlantic Ocean. Biol. Invasions 1999, 1, 353–361. [Google Scholar] [CrossRef]
- Kraemer, G.P.; Sellberg, M.; Gordon, A.; Main, J. Eight-year record of Hemigrapsus sanguineus (Asian shore crab) invasion in western Long Island Sound estuary. Northeast. Nat. 2007, 14, 207–224. [Google Scholar] [CrossRef]
- Griffen, B.D. Ecological impacts of replacing one invasive species with another in rocky intertidal areas. In In the Wrong Place—Alien Marine Crustaceans: Distribution, Biology and Impacts. Invading Nature—Springer Series in Invasion Ecology, Vol. 6; Galil, B.S., Clark, P.F., Carlton, J.T., Eds.; Springer: New York, NY, USA, 2011; pp. 687–701. [Google Scholar]
- Young, A.M.; (Salem State University, Salem, MA, USA). Personal observation, 2014.
- Richards, B.; Chief Scientist, Weedoo Greenboat Inc., West Palm Beach, FL, USA. Personal communication, 2018.
- Menge, B.A. Components of predation intensity in the low zone of the New England rocky intertidal region. Oecologia 1983, 58, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Epifanio, C.E. Invasion biology of the Asian shore crab Hemigrapsus sanguineus: A review. J. Exp. Mar. Biol. Ecol. 2013, 441, 33–49. [Google Scholar] [CrossRef]
- Baillie, C.; Grabowski, J.H. Invasion dynamics: Interactions between the European green crab Carcinus maenas and the Asian shore crab Hemigrapsus sanguineus. Biol. Invasions 2019, 21, 787–802. [Google Scholar] [CrossRef]
- Wong, M.C.; Dowd, M. Role of invasive green crabs in the food web of an intertidal sand flat determined from field observations and a dynamic simulation model. Estuaries Coasts 2014, 37, 1004–1016. [Google Scholar] [CrossRef]
- Moksnes, P.-O. Self-regulating mechanisms in cannibalistic populations of juvenile shore crabs Carcinus maenas. Ecology 2004, 85, 1343–1354. [Google Scholar] [CrossRef] [Green Version]
- Dunstone, N.; Birks, J.D.S. The feeding ecology of mink (Mustela vison) in coastal habitat. J. Zool. 1987, 212, 69–83. [Google Scholar] [CrossRef]
- Mason, C.F.; MacDonald, S.M. The winter diet of otters (Lutra zutra) on a Scottish sea loch. J. Zool. 1980, 192, 558–561. [Google Scholar] [CrossRef]
- Sergeant, D.E. The status of the common seal (Phoca vitulina L.) on the East Anglian coast. J. Mar. Biol. Assoc. UK 1951, 29, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Lynch, B.R.; Rochette, R. Spatial overlap and interaction between sub-adult American lobsters, Homarus americanus, and the invasive European green crab Carcinus maenas. J. Exp. Mar. Biol. Ecol. 2009, 369, 127–135. [Google Scholar] [CrossRef]
- Skonberg, D.I.; Perkins, D.L. Nutrient composition of green crab (Carcinus maenas) leg meat and claw meat. Food Chem. 2002, 77, 401–404. [Google Scholar] [CrossRef]
- McNiven, M.A.; Quijon, P.A.; Mitchell, A.W.; Ramsey, A.; St-Hilaire, S. Composition and distribution of the European green crab in Prince Edward Island, Canada. Open J. Anim. Sci. 2013, 3, 295–298. [Google Scholar] [CrossRef] [Green Version]
- St-Hilaire, S. Assessing the Potential for a Soft-Shell Green Crab Industry in PEI. Ph.D. Thesis, University of Prince Edward Island, Charlottetown, PE, Canada, 2016. [Google Scholar]
- Warner, R. Little green invaders. We have met the enemy and his name is Carcinus maenas. Now shall we eat him? The Boston Globe 2015, 287, 16–23. Available online: https://www.bostonglobe.com/magazine/2015/02/12/the-green-crab-problem-shall-eat-enemy/Ahtg6L87Gpxs0RMKntYAoN/story.html (accessed on 22 July 2017).
- Galetti, J.A.; Calder, B.L.; Skonberg, D.I. Mechanical separation of green crab (Carcinus maenas) meat and consumer acceptability of a value-added food product. J. Aquat. Food Prod. Tech. 2017, 26, 172–180. [Google Scholar] [CrossRef]
- McMahon, M. Investigating the viability of a soft-shell green crab industry in New England. In Proceedings of the Green Crab Working Summit Presentation, Portland, ME, USA, 6 June 2018. [Google Scholar]
- Parks, M. Bait to delicacy: Navigating and redefining the green crab market. In Proceedings of the Green Crab Working Summit Presentation, Portland, ME, USA, 6 June 2018. [Google Scholar]
- Skonberg, D. Toward full utilization of the green crab Biomass: Food and feed ingredients. In Proceedings of the Green Crab Working Summit Presentation, Portland, ME, USA, 6 June 2018. [Google Scholar]
- Kang, B.; Myracle, A.D.; Skonberg, D.I. Potential of recovered proteins from invasive green crabs (Carcinus maenas) as a functional food ingredient. J. Sci. Food Agric. 2019, 99, 1748–1754. [Google Scholar] [CrossRef]
- Parks, M.; Thái, T. The Green Crab Cookbook; Ingram Spark Publishing: La Vergne, TN, USA, 2018. [Google Scholar]
- Lee, K.T.; Jivoff, P.; Bishop, R.E. A low cost, reliable method for quantifying coloration in Carcinus maenas (Linnaeus, 1758) (Decapoda, Brachyura). Crustaceana 2005, 78, 579–590. [Google Scholar]
- Nagaraju, G.D.; Borst, D. Methyl farnesoate couples environmental changes to testicular development in the crab Carcinus maenas. J. Exp. Biol. 2008, 211, 2773–2778. [Google Scholar] [CrossRef] [Green Version]
- Abuhagr, A.M.; Blindert, J.L.; Nimitkul, S.; Zander, I.A.; LaBere, S.M.; Chang, S.A.; MacLea, K.S.; Chang, E.S.; Mykles, D.L. Molt regulation in green and red color morphs of the crab Carcinus maenas: Gene expression of molt-inhibiting hormone signaling components. J. Exp. Biol. 2014, 217, 796–808. [Google Scholar] [CrossRef] [Green Version]
- Deese, H.; Arnold, S. Green crabs working as lobster bait in Nova Scotia. Waterfr. Arch. 2014. Available online: http://www.workingwaterfrontarchives.org/2014/03/18/green-crabs-as-lobster-bait-working-in-nova-scotia/ (accessed on 18 March 2014).
- Ghosh, B.; Urban, M.W. Self-repairing oxetane-substituted chitosan polyurethane networks. Science 2009, 323, 1458–1460. [Google Scholar] [CrossRef] [PubMed]
- Morganti, P.; Li, Y.H. Innovation in cosmetic and medical science. The role of chitin nanofibrils composites. J. Appl. Cosmetol. 2015, 33, 9–24. [Google Scholar]
- Gao, Y.; Cranston, R. Recent advances in antimicrobial treatments of textiles. Text. Res. J. 2008, 78, 60–72. [Google Scholar]
- Varshosaz, J.; Jaffari, F.; Karimzadeh, S. Development of bioadhesive chitosan gels for topical delivery of lidocaine. Sci. Pharm. 2006, 74, 209–223. [Google Scholar] [CrossRef] [Green Version]
- Steiger, M.G.; Mach-Aigner, A.R.; Gorsche, R.; Rosenberg, E.E.; Mihovilovic, M.D.; Mach, R.L. Synthesis of an antiviral drug precursor from chitin using a saprophyte as a whole-cell catalyst. Microb. Cell Fact. 2011, 10, 102. [Google Scholar] [CrossRef] [Green Version]
- Teotia, S.; Lata, R.; Gupta, M. Chitosan as a macroaffinity ligand: Purification of chitinases by affinity precipitation and aqueous two-phase extractions. J. Chromatogr. A 2004, 1052, 85–91. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, C. Estuary Therapy: Advances in Coastal Restoration at Kejimkujik National Park Seaside. In Green Crab Summit Presentation; University of Maine: Orono, ME, USA, 2013; Available online: http://www.seagrant.umaine.edu/green-crab-summit (accessed on 16 December 2013).
- Lützen, J.; Jensen, K.H.; Glenner, H. Life history of Sacculina carcini Thompson, 1836 (Cirripedia: Rhizocephala: Sacculinidae) and the intermoult cycle of its host, the shore crab Carcinus maenas (Linnaeus, 1758) (Decapoda: Brachyura: Carcinidae). J. Crustac. Biol. 2018, 38, 413–419. [Google Scholar] [CrossRef]
- Mouritsen, K.N.; Jensen, T. The effect of Sacculina carcini infections on the fouling, burying, behavior and condition of the shore crab, Carcinus maenas. Mar. Biol. Res. 2006, 2, 270–275. [Google Scholar] [CrossRef]
- Bojko, J.; Subramaniam, K.; Waltzek, T.B.; Stentiford, G.D.; Behringer, D.C. Genomic and developmental characterisation of a novel bunyavirus infecting the crustacean Carcinus maenas. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Bojko, J.; Clark, F.; Bass, D.; Dunn, A.M.; Stewart-Clark, S.; Stebbing, P.D.; Stentiford, G.D. Parahepatospora carcini n. gen., n. sp., a parasite of invasive Carcinus maenas with intermediate features of sporogony between the Enterocytozoon clade and other Microsporidia. J. Invertebr. Pathol. 2017, 143, 124–134. [Google Scholar] [CrossRef]
- Torchin, M.E.; Lafferty, K.D.; Kuris, A.M. Release from parasites as natural enemies: Increased performance of a globally introduced marine crab. Biol. Invasions 2001, 3, 333–345. [Google Scholar] [CrossRef]
- Costa, S.; Bessa, F.; Pardal, M.A. The parasite Sacculina carcini Thompson, 1936 (Cirrepedia, Rhizocephala) in the crab Carcinus maenas (Linnaeus, 1758) (Decapoda, Portunidae): Influence of environmental conditions, colour morphotype and sex. Crustaceana 2013, 86, 34–47. [Google Scholar]
- Waser, A.M.; Goedknegt, M.A.; Dekker, R.; McSweeney, N.; Witte, J.I.J.; van der Meer, J.; Thieltges, D.W. Tidal elevation and parasitism: Patterns of infection by the rhizocephalan parasite Sacculina carcini in shore crabs Carcinus maenas. Mar. Ecol. Prog. Ser. 2016, 545, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Rytter Jensen, A.; Schneider, M.R.; Høeg, J.T.; Glenner, H.; Lützen, J. Variation in juvenile stages and success of male acquisition in Danish and French populations of the parasitic barnacle Sacculina carcini (Cirripedia: Rhizocephala) parasitizing the shore crab Carcinus maenas. Mar. Biol. Res. 2019, 15, 191–203. [Google Scholar] [CrossRef]
- Høeg, J.T.; Lützen, J. Life cycle and reproduction in the Cirripedia Rhizocephala. Oceanog. Mar. Biol. Ann. Rev. 1995, 33, 427–485. [Google Scholar]
- Torchin, M.E.; Mitchell, C.E. Parasites, pathogens, and invasions by plants and animals. Front. Ecol. Environ. 2004, 2, 183–190. [Google Scholar] [CrossRef]
- Blakeslee, A.M.H.; Keogh, C.L.; Fowler, A.E.; Griffen, B.D. Assessing the effects of trematode infection on invasive green crabs in eastern North America. PLoS ONE 2015, 10, e0128674. [Google Scholar] [CrossRef] [Green Version]
- Kuris, A. Nemertean egg predators as potential biocontrol agents for Carcinus maenas. In Proceedings of the First International Workshop on the Demography, Impacts and Management of Introduced Populations of the European crab, Carcinus maenas, Hobart, Australia, 20–21 March 1997; Thresher, R.E., Ed.; Technical Report 11. Centre for Research on Introduced Marine Pests: Hobart, TAS, Australia, 1997; pp. 88–90. [Google Scholar]
- Minchin, D. The influence of the parasitic cirripede Sacculina carcini on its brachyuran host Carcinus maenas within its home range. In Proceedings of the First International Workshop on the Demography, Impacts and Management of Introduced Populations of the European crab, Carcinus maenas, Hobart, Australia, 20–21 March 1997; Thresher, R.E., Ed.; Technical Report 11. Centre for Research on Introduced Marine Pests: Hobart, TAS, Australia, 1997; pp. 81–86. [Google Scholar]
- Goddard, J.H.R.; Torchin, M.E.; Kuris, A.M.; Lafferty, K.D. Host specificity of Sacculina carcini, a potential biological control agent of the introduced European green crab Carcinus maenas in California. Biol. Invasions 2005, 7, 895–912. [Google Scholar] [CrossRef]
- Crooks, J.A. Characterizing ecosystem-level consequences of biological invasions: The role of ecosystem engineers. Oikos 2002, 97, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Garbary, D.J.; Miller, A.G.; Williams, J.; Seymour, N.R. Drastic decline of an extensive eelgrass bed in Nova Scotia due to the activity of the invasive green crab (Carcinus maenas). Mar. Biol. 2014, 161, 3–15. [Google Scholar] [CrossRef]
- McDonald, P.S.; Holsman, K.K.; Beauchamp, D.A.; Dumbauld, B.R.; Armstrong, D.A. Bioenergetics modeling to investigate habitat use by the nonindigenous crab, Carcinus maenas, in Willipa Bay, Washington. Estuar. Coast. 2006, 29, 1132–1149. [Google Scholar] [CrossRef]
- Davis, R.C.; Short, F.T.; Burdick, D.M. Quantifying the effects of green crab damage to eelgrass transplants. Rest. Ecol. 1998, 6, 297–302. [Google Scholar] [CrossRef]
- Garbary, D.J.; Miller, A.G. Green crabs (Carcinus maenas) as the grim reaper: Destruction of eelgrass beds in Nova Scotia. J. Shellfish Res. 2006, 25, 728. [Google Scholar]
- Matheson, K.; McKenzie, C.H.; Gregory, R.S.; Robichaud, D.A.; Bradbury, I.A.; Snelgrove, P.V.; Rose, G.A. Linking eelgrass decline and impacts on associated fish communities to European green crab Carcinus maenas invasion. Mar. Ecol. Prog. Ser. 2016, 548, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Trussell, G.C.; Ewanchuk, P.J.; Bertness, M.D. Trait-mediated effects in rocky intertidal food chains: Predator risk cues alter prey feeding rates. Ecology 2003, 84, 629–640. [Google Scholar] [CrossRef]
- Dare, P.J.; Edwards, D.B. Experiments on the survival, growth and yield of relaid seed mussels (Mytilus edulis L.) in the Menai Straits, North Wales. ICES J. Mar. Sci. 1976, 37, 16–28. [Google Scholar] [CrossRef]
- Richards, M.G.; Huxman, M.; Bryant, A. Predation: A causal mechanism for variability in intertidal bivalve populations. J. Exp. Mar. Biol. Ecol. 1999, 241, 159–177. [Google Scholar] [CrossRef]
- Camphuysen, C.J.; Berrevoets, C.M.; Cremers, H.J.W.M.; Dekinga, A.; Dekker, R.; Ens, B.J.; van der Have, T.M.; Kats, R.K.H.; Kuiken, T.; Leopold, M.F.; et al. Mass mortality of common eiders (Somateria mollissima) in the Dutch Wadden Sea, winter 1999/2000: Starvation in a commercially exploited wetland of international imporE.B.ce. Biol. Conserv. 2002, 106, 303–317. [Google Scholar] [CrossRef]
- Welch, W.R. Changes in abundance of the green crab, Carcinus maenas (L.), in relation to recent temperature changes. Fish. Bull. 1968, 67, 337–345. [Google Scholar]
- Tan, E.B.P.; Beal, B.F. Interactions between the invasive European green crab, Carcinus maenas (L.), and juveniles of the soft-shell clam, Mya arenaria L., in eastern Maine, USA. J. Exp. Mar. Biol. Ecol. 2015, 462, 62–73. [Google Scholar] [CrossRef]
- McClenachan, L.; O’Connor, G.; Reynolds, T. Adaptive capacity of co-management systems in the face of environmental change: The soft-shell clam fisher and invasive green crab in Maine. Mar. Policy 2015, 52, 26–32. [Google Scholar] [CrossRef]
- Atlantic Coastal Cooperative Statistics Program. Available online: http://www.accsp.org (accessed on 7 July 2018).
- Beal, B.F.; Kraus, M.G. Interactive effects of initial size, stocking density, and type of predator deterrent netting on survival and growth of cultured juveniles of the soft-shell clam, Mya arenaria L, in eastern Maine. Aquaculture 2002, 208, 81–111. [Google Scholar] [CrossRef]
- Beal, B.F. Biotic and abiotic factors influencing growth and survival of wild and cultured individuals of the softshell clam (Mya arenaria L.) in eastern Maine. J. Shellfish Res. 2006, 25, 461–474. [Google Scholar] [CrossRef]
- Beal, B.F. Relative importance of predation and intraspecific competition in regulating growth and survival of juveniles of the softshell clam, Mya arenaria L., at several spatial scales. J. Exp. Mar. Biol. Ecol. 2006, 336, 1–17. [Google Scholar] [CrossRef]
- Beal, B.F.; Coffin, C.R.; Randall, S.F.; Goodenow, C.A.; Pepperman, K.E.; Ellis, B.W.; Jourdet, C.B.; Protopopescu, G.C. Spatial variability in recruitment of an infaunal bivalve: Experimental effects of predator exclusion on the softshell clam (Mya arenaria L.) along three tidal estuaries in southern Maine, USA. J. Shellfish Res. 2018, 37, 1–27. [Google Scholar] [CrossRef]
- Williams, P.J.; Macsween, C.; Rossong, M.A. Competition between invasive green crab (Carcinus maenas) and American lobster (Homarus americanus). N. Z. J. Mar. Freshw. Res. 2009, 44, 37–41. [Google Scholar] [CrossRef]
- Malyshev, A.; Quijón, P.A. Disruption of essential habitat by a coastal invader: New evidence of the effects of green crabs on eelgrass beds. ICES J. Mar. Sci. 2011, 68, 1852–1856. [Google Scholar] [CrossRef] [Green Version]
- Neckles, H.A. Loss of eelgrass in Casco Bay, linked to green crab disturbance. Northeast. Nat. 2015, 22, 478–500. [Google Scholar] [CrossRef]
- Howard, B.R.; Francis, F.T.; Côté, I.M.; Therriault, T.W. Habitat alteration by invasive European green crab (Carcinus maenas) causes eelgrass loss in British Columbia, Canada. Biol. Invasions 2019, 21, 3607–3618. [Google Scholar] [CrossRef]
- Finger, J. Concerns for the molluscan shellfish industry. In Proceedings of the Exotic Green Crab Presentation Abstract, Vancouver, WA, USA, 9–10 February 1998. [Google Scholar]
- Grosholz, E.D.; Olin, P.G.; Williams, B.; Tinsman, R. Reducing predation on Manila clams by nonindigenous European green crabs. J. Shellfish Res. 2001, 20, 913–919. [Google Scholar]
- Bojko, J.; Stebbing, P.D.; Dunn, A.M.; Bateman, K.S.; Clark, F.; Kerr, R.C.; Stewart-Clark, S.; Johannesen, Á.; Stentiford, G.D. Green crab Carcinus maenas symbiont profiles along a North Atlantic invasion route. Dis. Aquat. Org. 2018, 128, 147–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, G.H.; Bertness, M.D.; Yund, P.O. Crab predation, waterborne cues, and inducible defenses in the blue mussel, Mytilus edulis. Ecology 1999, 80, 1–14. [Google Scholar] [CrossRef]
- Trussell, G.C. Phemotypic plasticity in an intertidal snail – the role of a common crab predator. Evolution 1996, 50, 448–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seeley, R. Intense natural selection caused a rapid morphological transition in a living marine snail. Proc. Natl. Acad. Sci. USA 1986, 83, 6897–6901. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Worldwide distribution of green crab (Carcinus maenas) populations as of 2019. Solid icons represent established populations with dates of introduction. Open icons indicate potential locations for future invasions.
Figure 1.
Worldwide distribution of green crab (Carcinus maenas) populations as of 2019. Solid icons represent established populations with dates of introduction. Open icons indicate potential locations for future invasions.
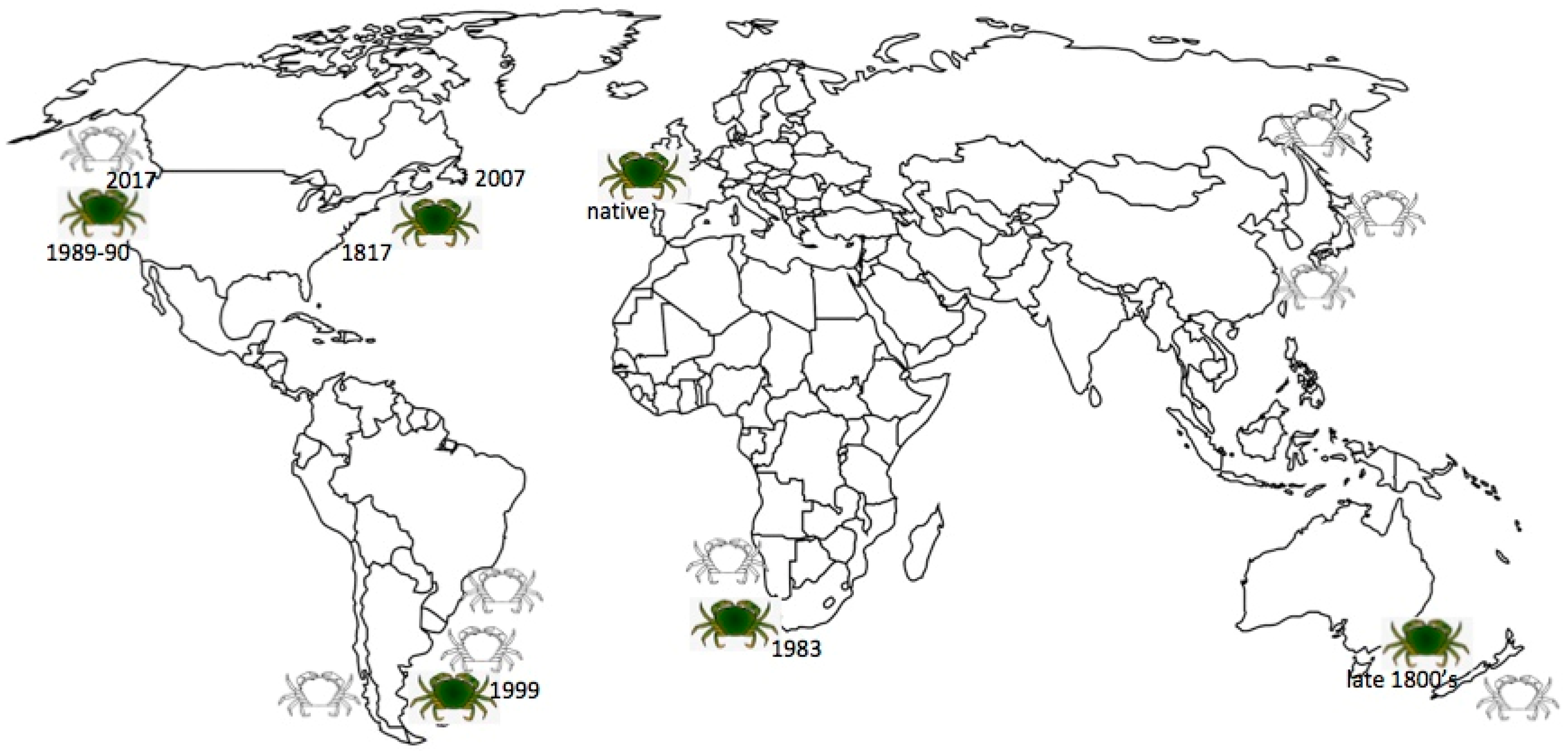

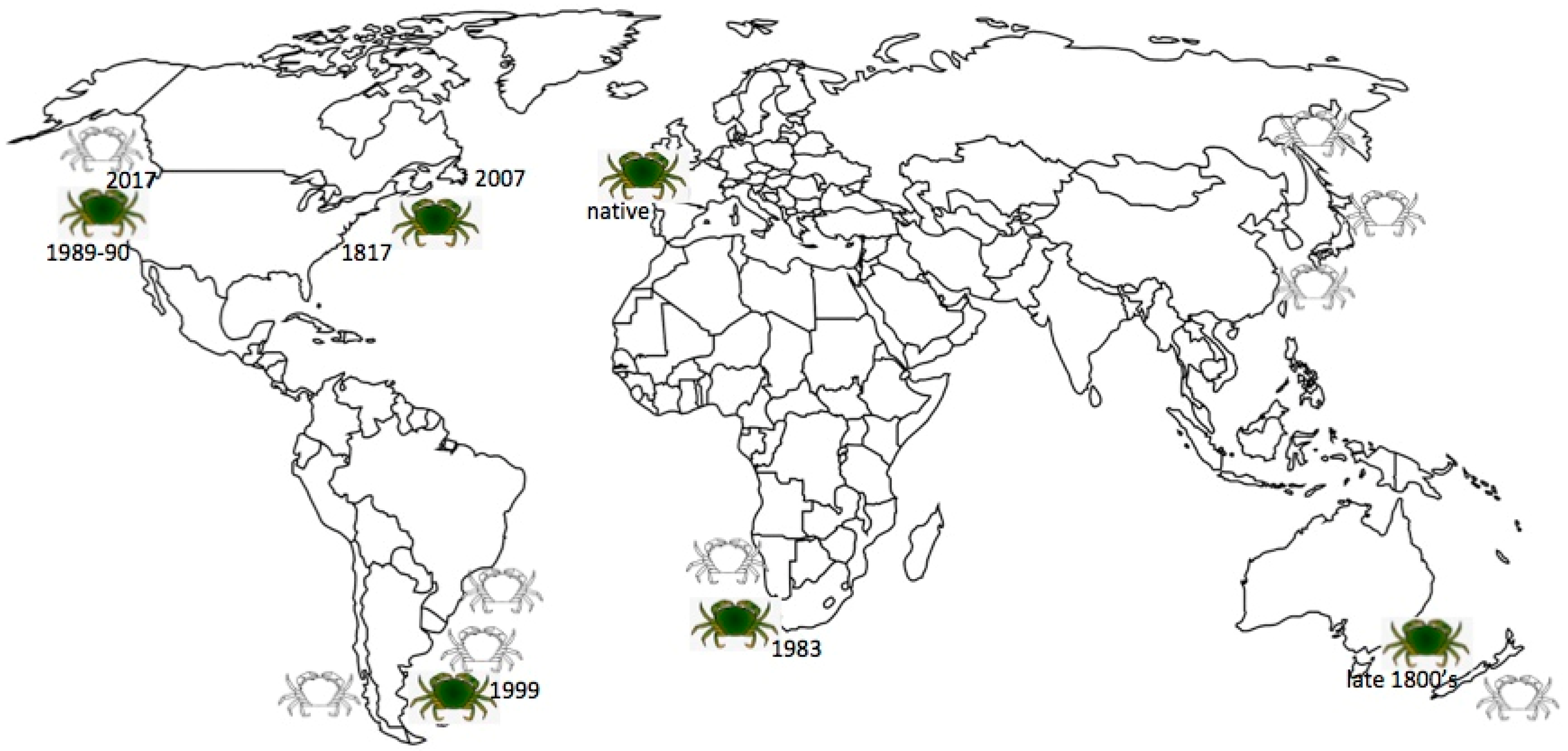{kind=link}
Table 1.
Seasonality of C. maenas reproductive events in different populations. Empty cells indicate no data are available for that location.
Table 1.
Seasonality of C. maenas reproductive events in different populations. Empty cells indicate no data are available for that location.
| Location | Copulation | Females Ovigerous | Larvae Released |
|---|---|---|---|
| Native European Population | |||
| Norway | January [189] | ||
| Scotland | All year (Peaks October to Spring Months) [190] | April to end of July [190] | |
| Denmark | May and June [191] | ||
| Baltic Germany | August [142] | ||
| United Kingdom | July-September [192] | November to December [192] | May-June [192] |
| Wales | August to September [5] | February to June (Peak in March and April) [5] | February to July [5] |
| The Netherlands | July to September (Peaks in August) [3] | November and December, and during the spring and early summer [3] | After winter season and in June [3] |
| Belgium | June to October [188] | December to August [188] | March to mid-October [188] |
| France | May to November (Peak in August and September) [7] | November to July (Peak in April) [7] | |
| Portugal | April-May and September-December [193] | All year [150]; October to June (Peaks in January-February) [193] | All year [150]; October through May [194]; February to June [195] |
| Northwestern Atlantic Nonindigenous Population | |||
| Newfoundland, Canada | Spring and Late Summer [196] | July and August [196] | Late May or Early June to August (Peak in June and July) [196] |
| Prince Edward Island, Canada | Peak from mid-August to September [147] | July to mid-September; (Peak in early July) [147,197] | August to December [147,197] |
| Maine, US | July to October (Peaks in August) [139] | April to August (Peaks from May to June) [139]; December to August, peak in summer months [125] | Peaks in September [139] |
| Massachusetts, US | November to January, and May to August [113] | ||
| Northeastern Pacific Nonindigenous Population | |||
| British Columbia, Canada | July [9,198] | April and May [9,198] | May [9,198] |
| South Africa Nonindigenous Population | |||
| Bloubergstrand and Sea Point | July to December [65] | ||
| Table Bay | June to January, peak in October [199] | ||
| Argentina Nonindigenous Population | |||
| San Jorge Gulf, Patagonia, Argentina | May to September [82] | ||
Table 2.
Maximum size (CW) reported and minimum size at sexual maturity in various populations of C. maenas. When not stated in the reference, CW was estimated from graphs. (C) = combined sexes or sex not indicated. Empty cells indicate no data are available. No data are available for nonindigenous population in Australia. Latitude rounded to nearest whole degree.
Table 2.
Maximum size (CW) reported and minimum size at sexual maturity in various populations of C. maenas. When not stated in the reference, CW was estimated from graphs. (C) = combined sexes or sex not indicated. Empty cells indicate no data are available. No data are available for nonindigenous population in Australia. Latitude rounded to nearest whole degree.
| Location | Latitude | Maximum Size (mm) | Sexual Maturity (mm) | Reference | ||
|---|---|---|---|---|---|---|
| Male | Female | Male | Female | |||
| Native European Population | ||||||
| Herdla, NO | 61° N | 80 | 44 | 28–30 | [189] | |
| Herdla, NO | 61° N | 80 | >20 | [185] | ||
| west coast of Sweden | 58° N | 72 | [219] | |||
| Western Sweden | 58–59° N | 100 | Moksnes, unpublished data, cited in Behrens Yamada [218] | |||
| Kattegat, DK | 57° N | 71 | 62 | [220] | ||
| Isefjord, DK | 55–56° N | 92 | [209] | |||
| Millport, SL | 56° N | 80 | [221] | |||
| Orensund, DK | 56° N | 80 (C) | [222] | |||
| Odense Fjord, DK | 56° N | 80 | [223] | |||
| Clyde Sea Area, SL | 56° N | 80 | [224] | |||
| Kerteminde Fjord, DK | 55° N | 81 | [225] | |||
| Schlei Fjord, Baltic Sea, DE | 54° N | 75 | 60 | [142] | ||
| Wadden Sea, DE | 54° N | 71 | [174] | |||
| Whitby Harbor and Robin Hood’s Bay, ENG | 53° N | 82.5 | 75 | 16–31 | [226] | |
| Boggle Hole, ENG | 54° N | 75 | [227] | |||
| Isle of Man, UK | 54° N | 27 | [207] | |||
| Llandudno and Conway, North WAL | 53° N | 75 | [228] | |||
| Menai Straits, North WAL | 53° N | 83.1 | 74.1 | [52] | ||
| Menai Straits, North WAL | 53° N | 75 | [229] | |||
| Menai Straits, North WAL | 53° N | 74 | [230] | |||
| Den Helder, NL | 53° N | 86 | 70 | 36–42 | [3] | |
| Essex, ENG | 52° N | 62 | [231] | |||
| Dale Peninsula, SW WAL | 51° N | 86 | 70 | 25 | 15 | [1] |
| Oosterschelde, NL | 52° N | 63 | [232] | |||
| Bullens Bay, IE | 52° N | 86.5 | 38.6 | [200] | ||
| Swansea, South WAL | 52° N | 86 | 70 | 44 | [5] | |
| Ostend, BE | 51° N | 44 | 23–45 | [188] | ||
| Cornwall, ENG | 50–51° N | 21 | 28 | [233] | ||
| Plymouth, ENG | 50° N | 57 | 65 | [231] | ||
| Mountbatten, Plymouth, ENG | 50° N | 65 (C) | [234] | |||
| Baltic Germany | 50° N | 75 | 60 | [8] | ||
| Luc-sur-Mer, FR | 49° N | 12 | [7] | |||
| Minho Estuary, PT | 42° N | 65 | 56 | 30 | [173] | |
| Canal de Mira, PT | 41° N | 27 | [195] | |||
| Mondego estuary, PT | 41° N | 71 | 65 | 29 | [150] | |
| Ericeira and Parede, PT | 39° N | 27 | 21.5 | [193] | ||
| Northwestern Atlantic Nonindigenous Population | ||||||
| Bras d’Or Lakes, Nova Scotia, CA | 46° N | 91 | 69 | 40 | [235] | |
| Placentia Bay, Newfoundland, CA | 48° N | 79 | 72 | 32 | 37 | [196] |
| Newfoundland west coast, CA | 48° N | 30 | McKenzie unpublished data, cited in Best et al. [196] | |||
| Basin Head, Prince Edward Island, CA | 46° N | 89.76 | 76.44 | 21.3 | 28.66 | [147] |
| Basin Head, Prince Edward Island, CA | 46° N | 80 | [236] | |||
| Basin Head Lagoon, Prince Edward Island, CA | 46° N | 42.7 | [197] | |||
| Long Harbor, Placentia Bay, Newfoundland, CA | 45° N | 78 | [237,238] | |||
| Boothbay Harbor, Maine, US | 44° N | 82 | 70 | 34 | 34 | [139] |
| Days Cove, Damariscotta, Maine, US | 44° N | 82 (C) | [239] | |||
| Broad Cove, Yarmouth, Maine, US | 44° N | 85 (C) | [239] | |||
| Webhannet River, Wells, Maine, US | 43° N | 78 (C) | [239] | |||
| Great Bay Estuary, New Hampshire, US | 43° N | 91 | 88 | [182] | ||
| Hampton-Seabrook Estuary, New Hampshire, US | 43° N | 36 | [182] | |||
| Pavilion Beach, Ipswich, Massachusetts, US | 42° N | 22 | A.M. Young, personal observation | |||
| Salem Sound, Massachusetts, US | 42° N | 80.9 | 85.9 | [113] | ||
| Northeastern Pacific Nonindigenous Population | ||||||
| Little Espinoza Inlet, British Columbia, CA | 50° N | 80.3 | 58.4 | [240] | ||
| Pipestem Inlet, Barkley Sound, British Columbia, CA | 49° N | 113 | 87 | [241] | ||
| Barkley Sound, British Columbia, CA | 49° N | 101.1 | 85.4 | [52] | ||
| Barkley Sound, British Columbia, CA | 49° N | 114 | 93 | 41 | [59] | |
| British Columbia, CA | 49 ° N | 106 | Therriault & DiBacco personal communication to Iain McGaw | |||
| Pipestem Inlet, Barkley Sound, British Columbia, CA | 49° N | 95 (C) | [242] | |||
| Vancouver Island, British Columbia, CA | 49° N | 98 | 76 | [198] | ||
| Oregon/Washington, US | 46–48° N | 99.6 | 79 | [218] | ||
| Tillamock Bay, Oregon, US | 46° N | 92 | 85 | [241] | ||
| Netarts Bay, Oregon, US | 45° N | 98 | 89 | [241] | ||
| Yaquina Bay, Oregon, US | 45° N | 96 | 83 | [241] | ||
| Yaquina Bay, Oregon, US | 45° N | 96 | 79 | [8] | ||
| Oregon, US | 45° N | 99 (C) | Behrens Yamada personal communication to Jamieson [240] | |||
| Coos Bay, Oregon, US | 43° N | 98 | 88 | [241] | ||
| Bodega Harbor, California, US | 38° N | 40 | [30] | |||
| Bodega Harbor, California, US | 38° N | 34 | [8] | |||
| Bodega Bay, California, US | 38° N | 102 | 88 | [241] | ||
| Tomales Bay, California, US | 38° N | 86 | 73 | [241] | ||
| Sea Drift Lagoon, California, US | 38° N | 88 | 88 | [241] | ||
| San Francisco Bay | 38° N | 83 | 80 | [241] | ||
| Elkhorn Slough, California, US | 37° N | 89 | 89 | [241] | ||
| Argentina Nonindigenous Population | ||||||
| Camarones Bay, AR | 45° S | 81.4 | 71.4 | 39.9 | [243] | |
| San Jorge Gulf, Patagonia, AR | 45° S | 73.4 | 64.3 | 45.8 | [82] | |
| South Africa Nonindigenous Population | ||||||
| Table Bay Docks, Cape Town, ZA | 34° S | 84 | 56 | [65] | ||
| Australia Nonindigenous Population | ||||||
| Gulf St. Vincent, South Australia, AU | 35° S | 80 | 67 | [244] | ||
Table 3.
Sex ratios (M:F) for various populations of C. maenas. Value > 1 indicates a higher proportion of males; value < 1 indicates a higher proportion of females.
Table 3.
Sex ratios (M:F) for various populations of C. maenas. Value > 1 indicates a higher proportion of males; value < 1 indicates a higher proportion of females.
| Location | Latitude (to Nearest Degree) | Sex Ratio (M:F) | Reference |
|---|---|---|---|
| Native European Population | |||
| Herdla, Norway | 61° N | 1.55; 1.48 | [187] |
| Herdia, Norway | 61° N | 1.49 | [185] |
| Kattegat, Denmark | 57° N | 51.63 * (shallow) 21.99 * (deeper) | [220] |
| Forth Estuary, Scotland | 56° N | 0.68 | [259] |
| Firth of Forth, Scotland1 | 56° N | 0.85 | [107] |
| Kerteminde Fjord, Denmark | 55° N | 2.99 | [145] |
| Kerteminde Fjord, Denmark | 55° N | 1.01 | [225] |
| Menai Strait, Anglesey, UK | 53° N | 1.86 | [263] |
| Whitby Harbor & Robin Hood’s Bay, England | 53° N | 0.45 | [226] |
| Ynys Faelog, Menai Strait, North Wales | 53° N | 2.33 | [143] |
| Den Helder, Netherlands | 53° N | 0.62 | [3] |
| Bullens Bay, Ireland | 51° N | 1.89 | [200] |
| Looe Estuary, Cornwall, England | 50° N | 0.64 | [233] |
| Mondego estuary, Portugal | 41° N | 0.89 | [150] |
| northwestern Atlantic nonindigenous population | |||
| Placentia Bay, Newfoundland, Canada | 48° N | 1.00 (September) > 1.00 (June) | [264] |
| Placentia Bay, Newfoundland, Canada | 48° N | 1.76 (fall) 1.33 (spring) 1.29 (summer) | [196] |
| Basin Head, PEI, Canada | 46° N | 1.63 | [147] |
| Bras d’Or Lakes, Nova Scotia, Canada | 46° N | 2.70 | [235] |
| Port L’Hebert, Nova Scotia, Canada | 44° N | 1.18 (May) 0.79 (August) | [122] |
| Boothbay Harbor, Maine | 44° N | 1.29 | [139] |
| Days Cove, Damariscotta, Maine | 44° N | 1.57 | [239] |
| Broad Cove, Yarmouth, Maine | 44° N | 0.60 | [239] |
| Biddeford Pool, Maine | 43° N | < 1.00 in 74.4% of samples | [125] |
| Webhannet River, Wells, Maine | 43° N | 0.94 | [239] |
| Great Bay Estuary, New Hampshire, US | 43° N | 1.00 | [265] |
| Great Bay Estuary, New Hampshire, US | 43° N | 0.37 | [182] |
| Houlton-Seabrook Estuary, New Hampshire, US | 43° N | 0.98 | [182] |
| Plum Island Sound, Massachusetts, US | 43° N | 0.34 (upstream) 0.90 (downstream) | [112] |
| Great Marsh, Rowley, Massachusetts, US | 43° N | 0.73 | [260] |
| Great Marsh, Rowley, Massachusetts, US | 43° N | 1.11 | [266] |
| Salem Sound, Massachusetts, US | 42° N | 0.37 | [113] |
| Nauset Marsh, Eastham, Massachusetts, US | 42° N | 0.26 | [260] |
| northeastern Pacific nonindigenous population | |||
| Nootka Sound, British Columbia, Canada | 49° N | 4.18 | [198] |
| Clayoquot Sound, British Columbia, Canada | 49 ° N | 3.88 | [198] |
| Barkley Sound, British Columbia, Canada | 49° N | 1.58 | [198] |
| Barkley Sound, British Columbia, Canada | 49° N | 2.45 | [263] |
| Barkley Sound, British Columbia, Canada | 49° N | 1.61 | [59] |
| Pipestem Inlet, British Columbia, Canada | 49° N | 1.71 | [241] |
| Tillamook Bay, Oregon, US | 45° N | 8.25 | [241] |
| Netarts Bay, Oregon, US | 45° N | 4.04 | [241] |
| Yaquina Bay, Oregon, US | 44° N | 6.00 | [241] |
| Coos Bay, Oregon, US | 43° N | 3.45 | [241] |
| Bodega Harbor, California, US | 38° N | 3.07 | [241] |
| Tomales Bay, California, US | 38° N | 2.36 | [241] |
| Seadrift lagoon, California, US | 38° N | 0.73 | [241] |
| San Francisco Bay, California, US | 37° N | 0.63 | [241] |
| Elkhorn Slough, California, US | 36° N | 0.72 | [241] |
| Argentina nonindigenous population | |||
| San Jorge Gulf, Patagonia, Argentina | 45° S | 2.36 | [82] |
* ratio calculated from percentages of males and females.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Young, A.M.; Elliott, J.A. Life History and Population Dynamics of Green Crabs (Carcinus maenas). Fishes 2020, 5, 4. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5010004
AMA Style
Young AM, Elliott JA. Life History and Population Dynamics of Green Crabs (Carcinus maenas). Fishes. 2020; 5(1):4. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5010004
Chicago/Turabian StyleYoung, Alan M., and James A. Elliott. 2020. "Life History and Population Dynamics of Green Crabs (Carcinus maenas)" Fishes 5, no. 1: 4. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5010004